
Temporal and Spatial Signatures of Scylla paramamosain Transcriptome Reveal Mechanistic Insights into Endogenous Ovarian Maturation under Risk of Starvation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
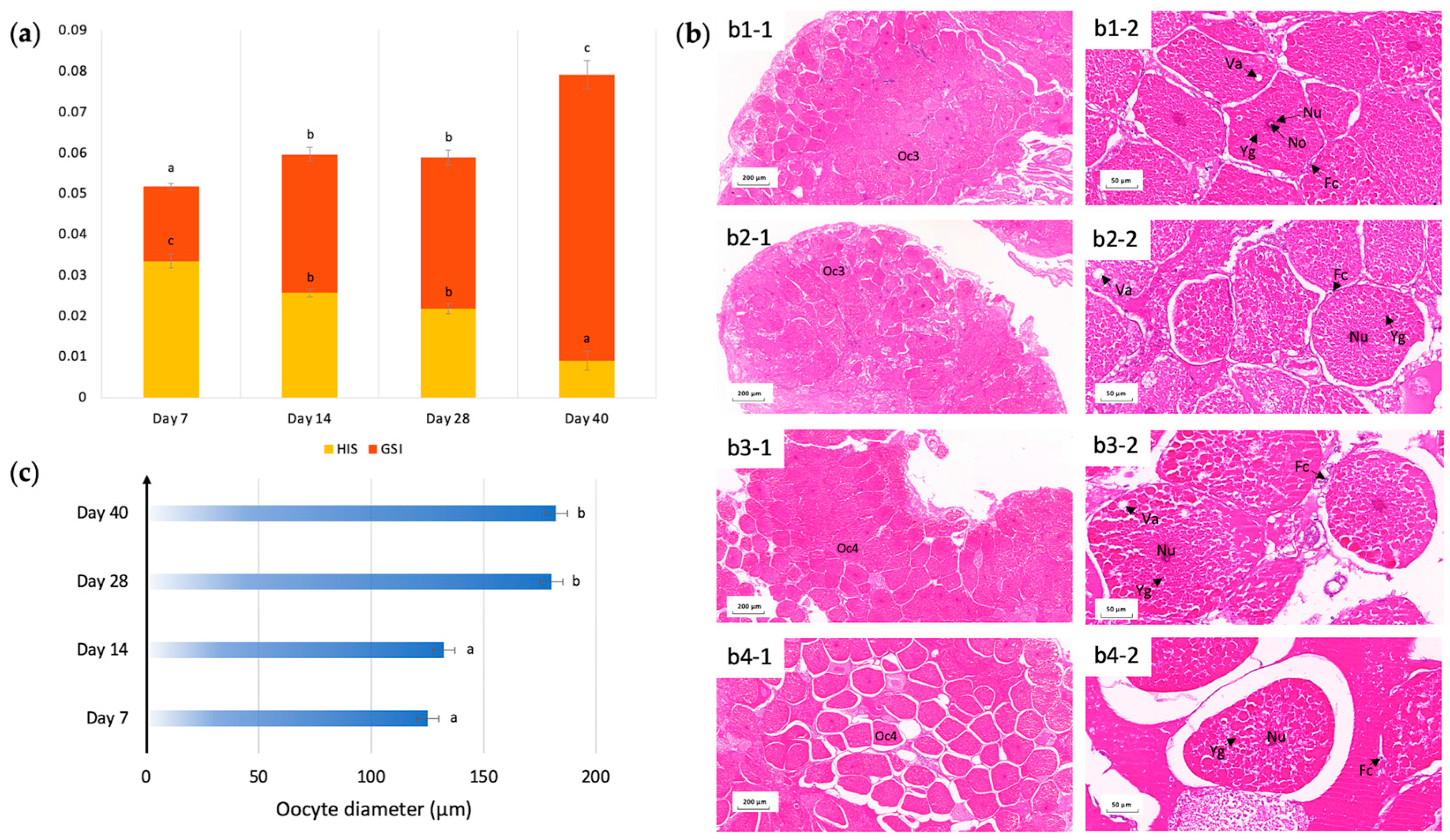
2.1. Maintenance of Ovarian Maturation under Imposed Starvation
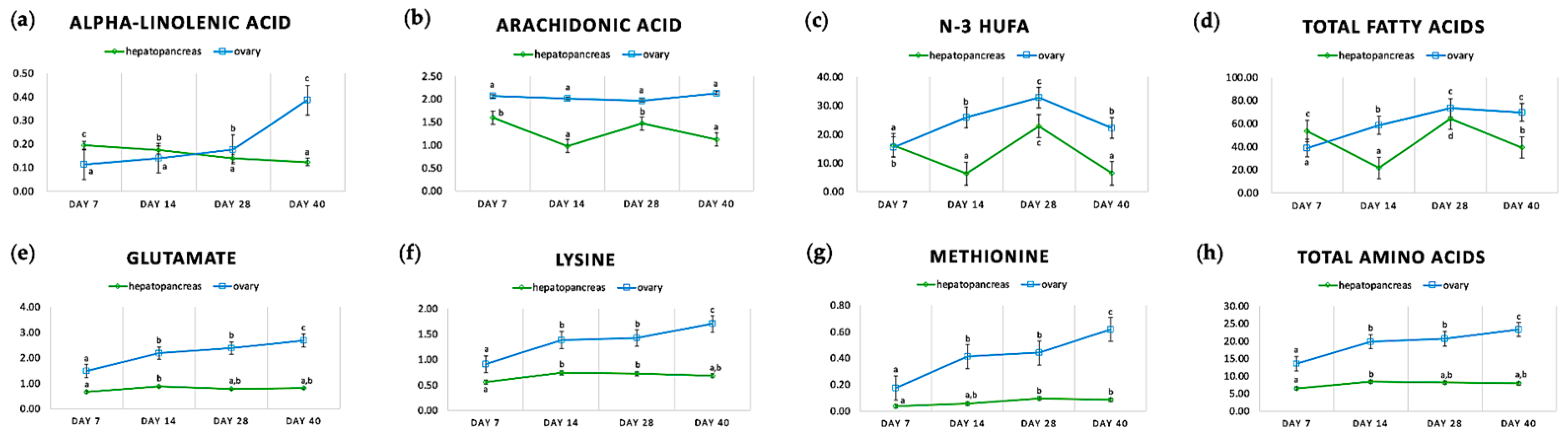
2.2. Starvation during Ovarian Maturation Resulted in Chemical and Biochemical Changes in Hepatopancreas, Ovary and Serum
2.3. The Landscape of the Scylla paramamosain Hepatopancreas and Ovary Transcriptomes
2.4. K-Means Analysis Revealed Time-Specific Gene Clusters
2.5. Starvation Alters the Expression of Genes Relevant to Epigenetic Modification
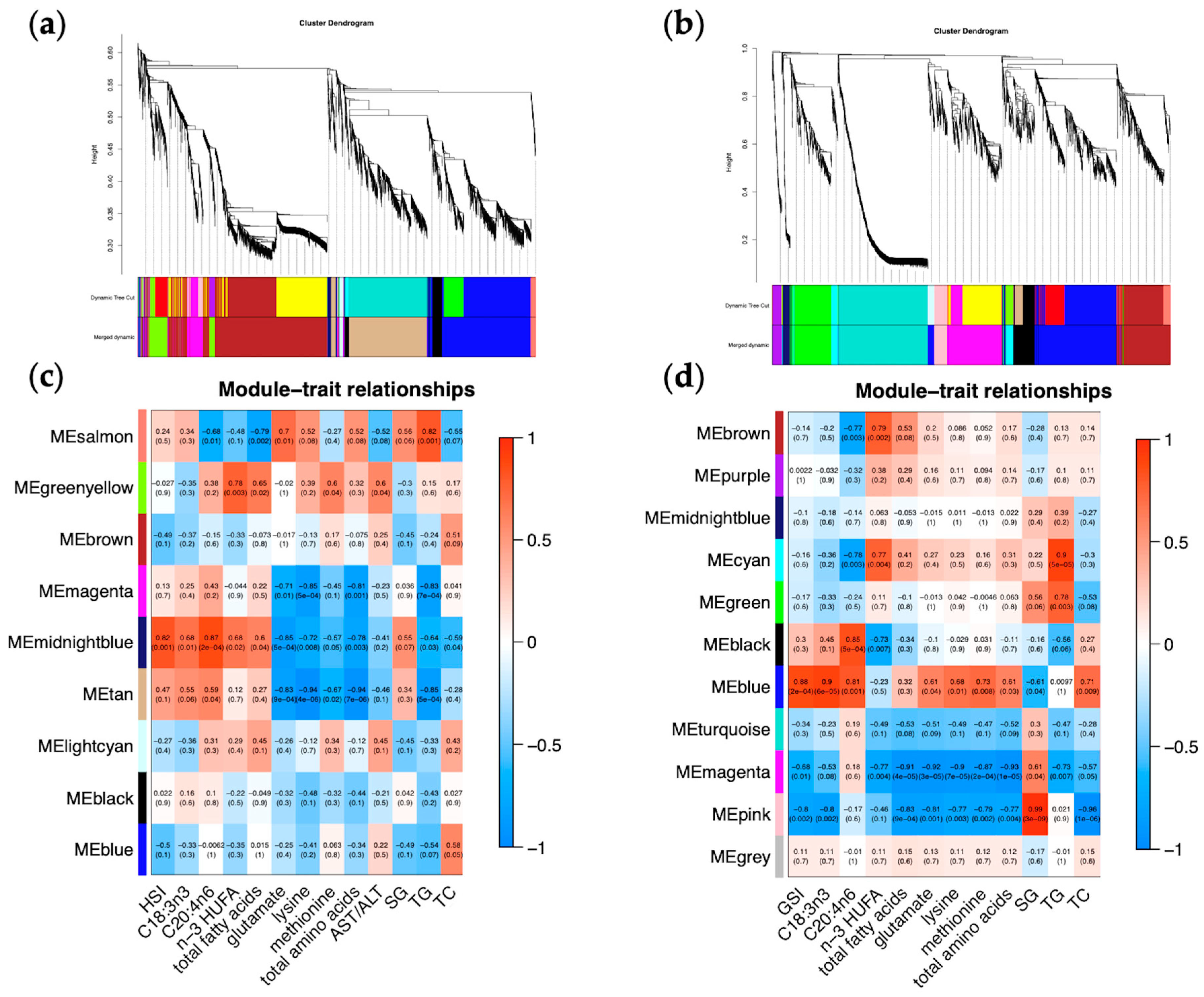
2.6. Starvation-Responsive Genes Are Highly Linked to Tissue-Specific Traits
3. Discussion
3.1. Terminal Investment of Scylla paramamosain into Ovarian Maturation
3.2. Phase Specificity of Ovarian Development under Imposed Starvation
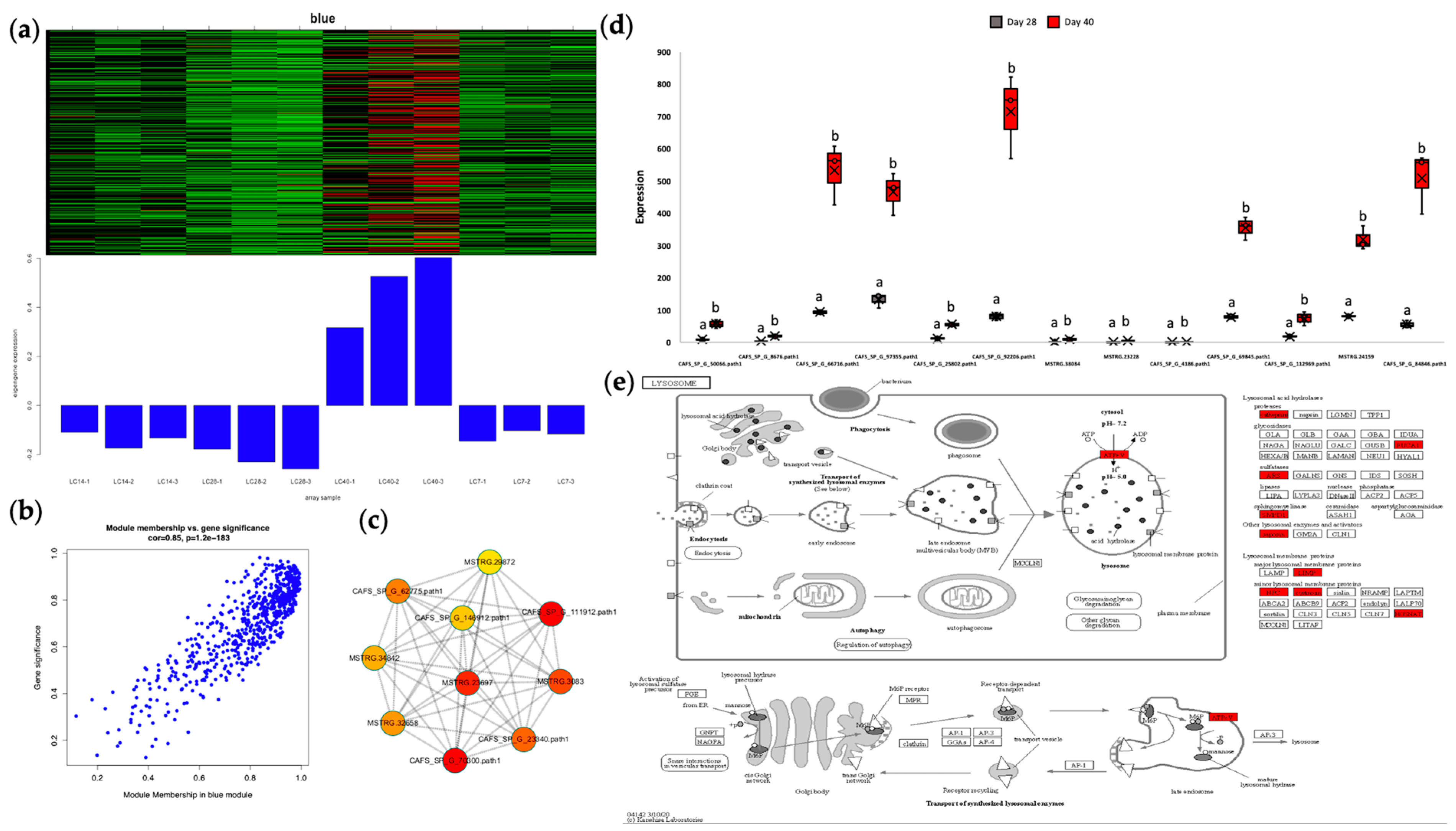
3.3. Autophagy at the Core of Regulatory Pathways
4. Materials and Methods
4.1. Ethics Statement
4.2. Animal Materials and Starvation Experiment
4.3. Hemolymph Collection and Serum Biochemistry
4.4. Tissue Collection and Histology
4.5. Chemical Analyses
4.5.1. GC–MS Analysis of Fatty Acids
4.5.2. HPLC Analysis of Amino Acids
4.6. Statistical Analysis
4.7. RNA Isolation, Library Preparation, and RNA-Seq
4.8. Sequence Alignment and Functional Annotation
4.9. Analysis of Gene Expression Patterns
4.10. Identification of Gene Modules Related to Ovarian Development under Imposed Starvation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McKenna, A.J. Rapid Evolution of Starvation Resistance in Drosophila: Physiological and Molecular Mechanisms. Ph.D. Thesis, University of Nevada, Las Vegas, NV, USA, 2020. [Google Scholar] [CrossRef]
- Felix, D.A.; Gutiérrez-Gutiérrez, Ó.; Espada, L.; Thems, A.; González-Estévez, C. It is not all about regeneration: Planarians striking power to stand starvation. Semin. Cell Dev. Biol. 2019, 87, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Ji, P.; Ao, S.; Gao, X.; Zhang, X. Effects of Starvation and Refeeding on Glucose Metabolism and Immune Responses in Macrobrachium rosenbergii. Mar. Biotechnol. 2023, 25, 447–462. [Google Scholar] [CrossRef] [PubMed]
- Szuwalski, C.S.; Aydin, K.; Fedewa, E.J.; Garber-Yonts, B.; Litzow, M.A. The collapse of eastern Bering Sea snow crab. Science 2023, 382, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Colson-Proch, C.; Morales, A.; Hervant, F.; Konecny, L.; Moulin, C.; Douady, C.J. First cellular approach of the effects of global warming on groundwater organisms: A study of the HSP70 gene expression. Cell Stress Chaperones 2010, 15, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, X.; Wang, C.; Song, Y.; Pan, J.; Shi, Q.; Qin, J.; Chen, L. Gamma-aminobutyric acid regulates glucose homeostasis and enhances the hepatopancreas health of juvenile Chinese mitten crab (Eriocheir sinensis) under fasting stress. Gen. Comp. Endocrinol. 2021, 303, 113704. [Google Scholar] [CrossRef] [PubMed]
- Thongrod, S.; Wanichanon, C.; Kankuan, W.; Siangcham, T.; Phadngam, S.; Morani, F.; Isidoro, C.; Sobhon, P. Autophagy-Associated Shrinkage of the Hepatopancreas in Fasting Male Macrobrachium rosenbergii Is Rescued by Neuropeptide F. Front. Physiol. 2018, 9, 613. [Google Scholar] [CrossRef] [PubMed]
- Comoglio, L.; Goldsmit, J.; Amin, O. Starvation effects on physiological parameters and biochemical composition of the hepatopancreas of the southern king crab Lithodes santolla (Molina, 1782). Rev. Biol. Mar. Y Oceanogr. 2008, 43, 345–353. [Google Scholar] [CrossRef]
- Donelson, J.M.; Munday, P.L.; McCormick, M.I.; Pankhurst, N.W.; Pankhurst, P.M. Effects of elevated water temperature and food availability on the reproductive performance of a coral reef fish. Mar. Ecol. Prog. Ser. 2010, 401, 233–243. [Google Scholar] [CrossRef]
- Pretterebner, K.; Pardo, L.M. All or nothing: Switch to high current reproductive investment under risk of starvation in male kelp crab. Ecol. Evol. 2020, 10, 3383–3391. [Google Scholar] [CrossRef]
- Kawabata, T.; Yoshimori, T. Beyond starvation: An update on the autophagic machinery and its functions. J. Mol. Cell. Cardiol. 2016, 95, 2–10. [Google Scholar] [CrossRef]
- Chang, Y.Y.; Neufeld, T.P. Autophagy takes flight in Drosophila. FEBS Lett. 2010, 584, 1342–1349. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, I.M.; Frederich, M.; Bagwe, R.; Lannig, G.; Sukhotin, A.A. Energy homeostasis as an integrative tool for assessing limits of environmental stress tolerance in aquatic invertebrates. Mar. Environ. Res. 2012, 79, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zamal, H.; Ollevier, F. Effect of feeding and lack of food on the growth, gross biochemical and fatty acid composition of juvenile catfish. J. Fish Biol. 2010, 46, 404–414. [Google Scholar] [CrossRef]
- Comoglio, L.; Smolko, L.; Amin, O. Effects of starvation on oxygen consumption, ammonia excretion and biochemical composition of the hepatopancreas on adult males of the False Southern King crab Paralomis granulosa (Crustacea, Decapoda). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2005, 140, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zhang, F.; Ma, C.; Wang, W.; Liu, Z.; Chen, W.; Zhao, M.; Ma, L. Comparative Metabolomics and Lipidomics of Four Juvenoids Application to Scylla paramamosain Hepatopancreas: Implications of Lipid Metabolism During Ovarian Maturation. Front. Endocrinol. 2022, 13, 886351. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Wang, K. Characterization of the innate immunity in the mud crab Scylla paramamosain. Fish Shellfish Immunol. 2019, 93, 436–448. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Paz, A.; García-Carreño, F.; Muhlia-Almazán, A.; Peregrino-Uriarte, A.B.; Hernández-López, J.; Yepiz-Plascencia, G. Usage of energy reserves in crustaceans during starvation: Status and future directions. Insect Biochem. Mol. Biol. 2006, 36, 241–249. [Google Scholar] [CrossRef]
- Chen, Z.; Ma, Y.; Cai, J.; Sun, M.; Zeng, L.; Wu, F.; Zhang, Y.; Hu, M. Serum biomarkers for liver fibrosis. Clin. Chim. Acta 2022, 537, 16–25. [Google Scholar] [CrossRef]
- Huang, X.; Feng, Y.; Duan, J.; Xiong, G.; Fan, W.; Liu, S.; Zhong, L.; Wang, K.; Geng, Y.; Ouyang, P.; et al. Antistarvation Strategies of E. sinensis: Regulatory Networks under Hepatopancreas Consumption. Oxidative Med. Cell. Longev. 2020, 2020, 6085343. [Google Scholar] [CrossRef]
- Schwartz, T.S.; Pearson, P.; Dawson, J.; Allison, D.B.; Gohlke, J.M. Effects of fluctuating temperature and food availability on reproduction and lifespan. Exp. Gerontol. 2016, 86, 62–72. [Google Scholar] [CrossRef]
- Duffield, K.R.; Bowers, E.K.; Sakaluk, S.K.; Sadd, B.M. A dynamic threshold model for terminal investment. Behav. Ecol. Sociobiol. 2017, 71, 185. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Lv, N.; Shi, Q.; Hu, X.; Wu, J. Reproductive adaptation in alate adult morphs of the English grain aphid Sitobion avenae under starvation stress. Sci. Rep. 2019, 9, 2023. [Google Scholar] [CrossRef] [PubMed]
- Krams, I.A.; Krama, T.; Moore, F.R.; Rantala, M.J.; Mänd, R.; Mierauskas, P.; Mänd, M. Resource availability as a proxy for terminal investment in a beetle. Oecologia 2015, 178, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Judge, K.A.; De Luca, P.; Morris, G.K. Food limitation causes female haglids to mate more often. Can. J. Zool. 2011, 89, 992–998. [Google Scholar] [CrossRef]
- Barrett, E.L.; Hunt, J.; Moore, A.J.; Moore, P.J. Separate and combined effects of nutrition during juvenile and sexual development on female life-history trajectories: The thrifty phenotype in a cockroach. Proc. R. Soc. B Biol. Sci. 2009, 276, 3257–3264. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Feng, Y.; Yang, M.; Huang, Y.; Li, M.; Geng, Y.; Ouyang, P.; Chen, D.; Yang, S.; Yin, L.; et al. Starvation induces hepatopancreas atrophy in Chinese mitten crab (Eriocheir sinensis) by inhibiting angiogenesis. BMC Genom. 2023, 24, 612. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Paz, A.; Garcia-Carreno, F.; Hernandez-Lopez, J.; Muhlia-Almazan, A.; Yepiz-Plascencia, G. Effect of short-term starvation on hepatopancreas and plasma energy reserves of the Pacific white shrimp (Litopenaeus vannamei). J. Exp. Mar. Biol. Ecol. 2007, 340, 184–193. [Google Scholar] [CrossRef]
- Flatt, T. Survival costs of reproduction in Drosophila. Exp. Gerontol. 2011, 46, 369–375. [Google Scholar] [CrossRef]
- Ehrlich, S. Effect of fertility and infertility on longevity. Fertil. Steril. 2015, 103, 1129–1135. [Google Scholar] [CrossRef]
- Wang, X.; Wang, L.; Zhang, H.; Liu, R.; Song, L. The carbohydrate metabolism of scallop Chlamys farreri in the immune response against acute challenge of Vibrio anguillarum. Aquacult. Int. 2015, 23, 1141–1155. [Google Scholar] [CrossRef]
- Kankuan, W.; Wanichanon, C.; Titone, R.; Engsusophon, A.; Sumpownon, C.; Suphamungmee, W.; Morani, F.; Masini, M.; Novelli, M.; Isidoro, C.; et al. Starvation Promotes Autophagy-Associated Maturation of the Ovary in the Giant Freshwater Prawn, Macrobrachium rosenbergii. Front. Physiol. 2017, 8, 300. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, V.K.; Timmons, A.K.; McCall, K. Diversity of cell death pathways: Insight from the fly ovary. Trends Cell Biol. 2013, 23, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Takahata, Y.; Miyakawa, H. Developmental Staging of Sexual Egg Formation in Daphnia pulex: Unmated Females Resorb Meiotic Oocytes to Resist Starvation. Zool. Sci. 2022, 39, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.Y.; Dong, S.Z.; Ye, G.Y.; Li, K.; Zhu, J.Y.; Fang, Q.; Hu, C. Oosorption in the endoparasitoid, Pteromalus puparum. J. Insect Sci. 2011, 11, 90. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Chen, Y.; Tooze, S.A. Autophagy pathway: Cellular and molecular mechanisms. Autophagy 2018, 14, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.; Rubinsztein, D.C.; Walker, D.W. Autophagy as a promoter of longevity: Insights from model organisms. Nat. Rev. Mol. Cell Biol. 2018, 19, 579–593. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Khawar, M.B.; Li, W. Essential role of autophagy in resource allocation during sexual reproduction. Autophagy 2020, 16, 18–27. [Google Scholar] [CrossRef]
- Suzuki, S.W.; Onodera, J.; Ohsumi, Y. Starvation induced cell death in autophagy-defective yeast mutants is caused by mitochondria dysfunction. PLoS ONE 2011, 6, e17412. [Google Scholar] [CrossRef]
- Zhang, N.; Zhao, Y. Other Molecular Mechanisms Regulating Autophagy. Adv. Exp. Med. Biol. 2019, 1206, 261–271. [Google Scholar] [CrossRef]
- Mizushima, N. A brief history of autophagy from cell biology to physiology and disease. Nat. Cell Biol. 2018, 20, 521–527. [Google Scholar] [CrossRef]
- Ma, L.; Xu, J.; Tao, X.; Wu, J.; Wang, W.; Pu, Y.; Yang, G.; Fang, Y.; Liu, L.; Li, X.; et al. Genome-Wide Identification of C2H2 ZFPs and Functional Analysis of BRZAT12 under Low-Temperature Stress in Winter Rapeseed (Brassica rapa). Int. J. Mol. Sci. 2022, 23, 12218. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Li, F.; Wang, Y.; Wang, Y.; Lin, Z.; Cheng, X.; Liu, J.; Chen, C.; Pan, L. Molecular basis of ubiquitin recognition by the autophagy receptor CALCOCO2. Autophagy 2015, 11, 1775–1789. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.-P.; Xiao, J.; Wu, Y.Q. The Role of PI3K/AKT/FOXO Signaling Pathway-Mediated Autophagy in Diabetes-Associated Cognitive Dysfunction. Chin. J. Biochem. Mol. Biol. 2021, 37, 588–594. [Google Scholar]
- Xie, Z.; Zhang, D.; Chung, D.; Tang, Z.; Huang, H.; Dai, L.; Qi, S.; Li, J.; Colak, G.; Chen, Y.; et al. Metabolic Regulation of Gene Expression by Histone Lysine β-Hydroxybutyrylation. Mol. Cell 2016, 62, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Medvedeva, Y.A.; Lennartsson, A.; Ehsani, R.; Kulakovskiy, I.V.; Vorontsov, I.E.; Panahandeh, P.; Khimulya, G.; Kasukawa, T.; Drabløs, F. EpiFactors: A comprehensive database of human epigenetic factors and complexes. Database 2015, 2015, bav067. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Oh, S.; Abmayr, S.M.; Workman, J.L. When histones are under glucose starvation. J. Biosci. 2020, 45, 17. [Google Scholar] [CrossRef]
- Xu, Y.; Wan, W. Acetylation in the regulation of autophagy. Autophagy 2023, 19, 379–387. [Google Scholar] [CrossRef]
- Lee, K.K.; Workman, J.L. Histone acetyltransferase complexes: One size doesn’t fit all. Nat. Rev. Mol. Cell Biol. 2007, 8, 284–295. [Google Scholar] [CrossRef]
- Shen, Q.; Shi, Y.; Liu, J.; Su, H.; Huang, J.; Zhang, Y.; Peng, C.; Zhou, T.; Sun, Q.; Wan, W.; et al. Acetylation of STX17 (syntaxin 17) controls autophagosome maturation. Autophagy 2021, 17, 1157–1169. [Google Scholar] [CrossRef]
- Suo, S.; Kimura, Y.; Van Tol, H.H. Starvation induces cAMP response element-binding protein-dependent gene expression through octopamine-Gq signaling in Caenorhabditis elegans. J. Neurosci. 2006, 26, 10082–10090. [Google Scholar] [CrossRef]
- Choi, S.; Kim, W.; Chung, J. Drosophila salt-inducible kinase (SIK) regulates starvation resistance through cAMP-response element-binding protein (CREB)-regulated transcription coactivator (CRTC). J. Biol. Chem. 2011, 286, 2658–2664. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Wang, W.; Fu, Y.; Liu, Z.; Zhao, M.; Xu, L.; Zhan, T.; Huang, T.; Luo, M.; Chen, W.; et al. Comparative Transcriptome Analysis Identifies MAPK Signaling Pathway Associated with Regulating Ovarian Lipid Metabolism during Vitellogenesis in the Mud Crab, Scylla paramamosain. Fishes 2023, 8, 145. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, W.; Liu, Z.; Luo, M.; Fu, Y.; Zhang, F.; Ma, C.; Zhao, M.; Chen, W.; Jiang, K.; et al. Insight of vitellogenesis patterns: A comparative analysis of the differences between the primary and secondary vitellogenesis period in the ovary, hepatopancreas, and muscle of mud crab, Scylla paramamosain. Front. Genet. 2022, 13, 965070. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Waiho, K.; Huang, Z.; Li, S.; Zheng, H.; Zhang, Y.; Ikhwanuddin, M.; Lin, F.; Ma, H. Growth performance and biochemical composition dynamics of ovary, hepatopancreas and muscle tissues at different ovarian maturation stages of female mud crab, Scylla paramamosain. Aquaculture 2020, 515, 734560. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, W.; Zhang, F.; Ma, C.; Liu, Z.; Yang, M.H.; Chen, W.; Li, Q.; Cui, M.; Jiang, K.; et al. A chromosome-level genome of the mud crab (Scylla paramamosain estampador) provides insights into the evolution of chemical and light perception in this crustacean. Mol. Ecol. Resour. 2021, 21, 1299–1317. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, X.; Tang, B.; Li, C.; Kou, Z.; Li, L.; Liu, W.; Wu, Y.; Kou, X.; Li, J.; et al. Protein Expression Landscape of Mouse Embryos during Pre-implantation Development. Cell Rep. 2017, 21, 3957–3969. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Chin, C.H.; Chen, S.H.; Wu, H.H.; Ho, C.W.; Ko, M.T.; Lin, C.Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8 (Suppl. S4), S11. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Y.; Zhang, F.; Wang, W.; Xu, J.; Zhao, M.; Ma, C.; Cheng, Y.; Chen, W.; Su, Z.; Lv, X.; et al. Temporal and Spatial Signatures of Scylla paramamosain Transcriptome Reveal Mechanistic Insights into Endogenous Ovarian Maturation under Risk of Starvation. Int. J. Mol. Sci. 2024, 25, 700. https://doi.org/10.3390/ijms25020700
Fu Y, Zhang F, Wang W, Xu J, Zhao M, Ma C, Cheng Y, Chen W, Su Z, Lv X, et al. Temporal and Spatial Signatures of Scylla paramamosain Transcriptome Reveal Mechanistic Insights into Endogenous Ovarian Maturation under Risk of Starvation. International Journal of Molecular Sciences. 2024; 25(2):700. https://doi.org/10.3390/ijms25020700
Chicago/Turabian StyleFu, Yin, Fengying Zhang, Wei Wang, Jiayuan Xu, Ming Zhao, Chunyan Ma, Yongxu Cheng, Wei Chen, Zhixing Su, Xiaokang Lv, and et al. 2024. "Temporal and Spatial Signatures of Scylla paramamosain Transcriptome Reveal Mechanistic Insights into Endogenous Ovarian Maturation under Risk of Starvation" International Journal of Molecular Sciences 25, no. 2: 700. https://doi.org/10.3390/ijms25020700
APA StyleFu, Y., Zhang, F., Wang, W., Xu, J., Zhao, M., Ma, C., Cheng, Y., Chen, W., Su, Z., Lv, X., Liu, Z., Ma, K., & Ma, L. (2024). Temporal and Spatial Signatures of Scylla paramamosain Transcriptome Reveal Mechanistic Insights into Endogenous Ovarian Maturation under Risk of Starvation. International Journal of Molecular Sciences, 25(2), 700. https://doi.org/10.3390/ijms25020700