Abstract
Dysfunction of the neuroglia can have profound consequences on the blood–brain barrier (BBB). Studies have shown that the disruption of astrocytic–endothelial interaction can compromise the permeability of BBB and its effectiveness in selectively regulating the exchange of substances. Microglia have recently been recognized to have a significant role in the initiation of chronic pain and in its interactions with various nerve blockers and anesthetic agents. Microglia have a role in pain resolution via a pathway that involves Cannabinoid receptor type 2 activation and MAP kinase phosphorylation. Understanding the role of these cells in the context of neuropathic pain and neurological disorders can aid in improving clinical outcomes and the challenging nature of managing pain. Advancing studies have proposed pharmacological and genetic modulation of microglia as a potential treatment option for patients with chronic pain.
1. Introduction
The blood–brain barrier (BBB) plays a crucial role in protecting the central nervous system (CNS). The BBB is comprised of tightly sealed endothelial cells lining the blood vessels of the brain, which serve to strictly block the passage of undesired molecules into the brain. The role of the BBB is to safeguard what molecules, drugs, and nutrients are able to cross from the peripheral bloodstream into the brain tissue [1]. The semipermeable nature of the BBB selectively allows the passage of nutrients and desired molecules into the brain tissue and prevents the passage of potentially harmful toxins, pathogens, or autoantibodies [2].
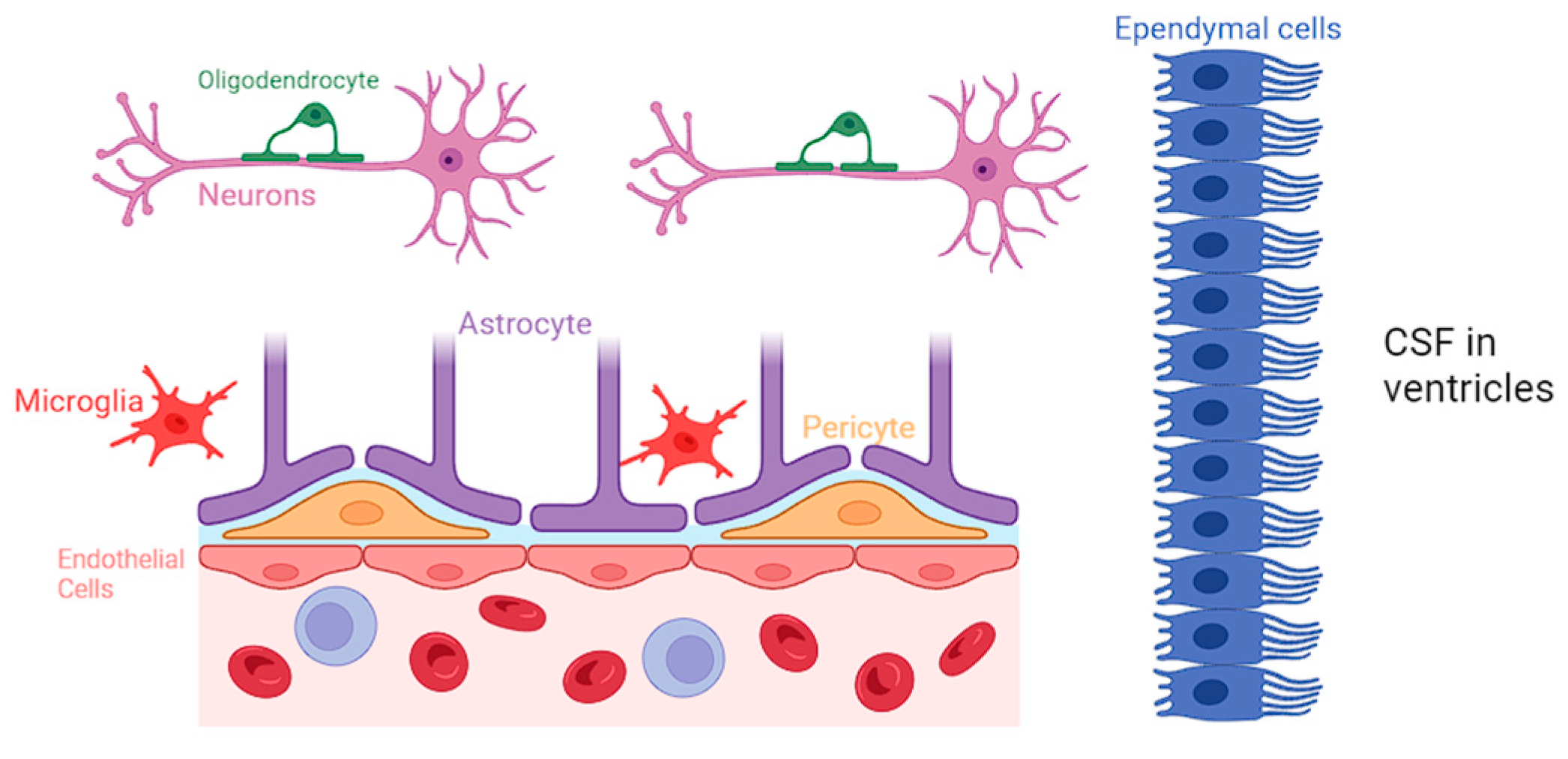
Neuroglia constitute a diverse group of cells within the CNS, comprising astrocytes, microglia, oligodendrocytes, and ependymal cells [3]. While neurons are traditionally considered the primary functional units of the nervous system, neuroglia provide crucial support, maintaining the structural and functional integrity of the neural environment [3]. A comprehensive representation of all the various types of neuroglial cells is presented in Figure 1.
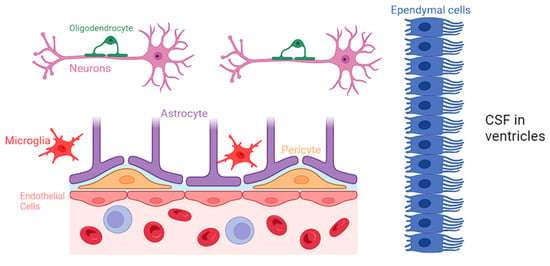
Figure 1.
Biochemical specifics of the BBB and the specific role of each neuroglial cell.
Astrocytes are the most abundant type of neuroglia and are particularly noteworthy for their intricate association with the BBB. They closely interact with the endothelial cells of the CNS and regulate the permeability of BBB by releasing various signaling molecules, including growth factors and cytokines, and maintaining the tight junctions in the blood vessels [4]. Astrocytes have a protrusion called “end-feet”, which allows them to sheath brain vasculature to maintain tight junctions and regulate blood flow [5]. Moreover, astrocytes play a role in regulating the transport of nutrients and waste products across the BBB, contributing to the overall homeostasis of the neural environment [6].
Microglia are traditionally known for their immune surveillance and response functions. However, recent research has shed light on the intricate involvement of microglia in BBB regulation [7]. They actively participate in the modulation of tight junctions between endothelial cells, contributing to the selective permeability of the BBB. Studies have identified that following dysfunction of the BBB, microglia release factors such as transforming growth factor-beta (TGF-β) and interleukin-10 (IL-10), which have been shown to enhance the integrity of the BBB by promoting tight junction formation [8,9]. Microglia are also able to phagocytize cellular debris and pathogens that may be causing BBB disruption, and clearance of such materials can help return the BBB to a more normal state of functioning. Additionally, microglia can interact with astrocytes to promote the secretion of growth factors that can help heal damaged components of the BBB [10,11,12].
Oligodendrocytes play a pivotal role in the CNS by producing myelin and facilitating rapid electrical signal transmission. While the primary function of oligodendrocytes lies in myelination, emerging research suggests their involvement in BBB regulation. Studies have demonstrated that oligodendrocytes are closely associated with endothelial cells of the BBB, and they may participate in the modulation of barrier permeability. The interactions between oligodendrocytes and endothelial cells at the BBB highlight the complexity of neuroglial contributions to the maintenance of a tightly regulated neural microenvironment [13,14].
Ependymal cells contribute to the functionality of BBB primarily through their role in cerebrospinal fluid (CSF) production and circulation but also via their involvement in modulating the BBB’s permeability. Recent studies have highlighted the expression of tight junction proteins in ependymal cells, indicating their potential contribution to the barrier function [15]. Ependymal cells can also work along with other glial cells, influencing the overall regulation of brain homeostasis.
Furthermore, pericytes serve the BBB by promoting blood vessel formation, maintaining the BBB, regulating immune cell entry, and controlling blood flow [16,17]. Pericytes stimulate angiogenesis through the secretion of vascular endothelial growth factor (VEGF), platelet-derived growth factor (PDGF), and angiopoietin-1, which can support the development of new capillaries. Additionally, pericytes control vasodilation and vasoconstriction, which allows them to control blood flow corresponding to the needs of the tissue. This helps tissue regenerate from damage and ensures the strength of the vasculature making up the BBB [18,19,20].
Dysfunction of the neuroglia can have profound consequences on the BBB. Studies have shown that the disruption of astrocytic–endothelial interaction can compromise the permeability of BBB and its effectivity on selectively regulating the exchange of substances between blood and the brain [7]. This compromised barrier function allows for the uncontrolled entry of potentially harmful molecules, including inflammatory mediators and toxins, into the brain parenchyma. This disruption can contribute to the development of neuroinflammatory diseases like multiple sclerosis (MS), neuromyelitis optica (NMO), and systemic lupus erythematosus (SLE) [2]. Dysfunction of the BBB plays a particular role in autoimmune neurological disorders due to effector molecules of the peripheral immune system being able to enter the brain and stimulate an inflammatory response, leading to disruption of normal neural function [21].
2. Pathophysiology of Pain
Pain is a nuanced and intricate experience, functioning as a major alarm system that signifies a potential threat or injury, which serves an adaptive role in protecting the body from harm. Yet, in certain cases, chronic pain can evolve into a maladaptive condition, causing significant personal and economic burdens [22,23,24,25,26]. Thus, effective pain management becomes crucial in mitigating these challenges, as it not only improves the individual’s quality of life but also reduces the broader societal impact associated with healthcare costs, productivity loss, and emotional distress. However, the dynamics of pain involve a more complex interaction among biological, psychological, and emotional systems [27], and the perception of pain varies significantly among individuals, highlighting the high degree of inter- and intra-patient variability [28]. Recent advances in imaging modalities, such as functional magnetic resonance imaging (fMRI) and positron emission tomography (PET), have significantly enhanced our understanding of the central role played by the brain in the intricate processes of perceiving and modulating pain signals through the ascending and descending pathways.
These advances have also shed light on the interactions that macrophages have on our ability to feel pain. Many research studies have been conducted to analyze macrophages and their interactions with various nociceptors in our body. The Toll-like receptors that begin to circulate in our body after activation of nociceptors have been shown to activate macrophages and lengthen the pain that we feel. This is imperative to understanding when thinking about the differences between the ascending and descending pathways.
2.1. Ascending Pathway
Specialized neurons within the peripheral nervous system detect potentially harmful stimuli such as pain, temperature, and noxious chemicals through the activation of receptors like nociceptors [27,29]. These nociceptors transmit signals to the spinal cord, initiating the first leg of the ascending pathway [29]. Within the spinal cord, these signals synapse with secondary neurons that carry the information up to the brain, where the stimulus is further processed [29]. This process uses the ventrolateral system, alternatively termed the spinothalamic tract, which serves as a specialized tract in the transmission of pain and temperature signals to the thalamus—a central relay station for sensory information—and subsequently to the cortex of the brain [29,30]. As the signals ascend further, they reach various regions of the brain, including the somatosensory cortex, where the conscious perception of pain occurs [29,30]. Simultaneously, other brain areas, such as the limbic system, contribute to the emotional and affective aspects of pain [29]. The ascending pathways serve to relay information about the location, intensity, and quality of the pain, enabling the brain to generate an appropriate response [30].
2.2. Descending Pathway
In addition to the ascending pathways, the brain possesses a sophisticated system of descending pathways that play a crucial role in modulating the perception of pain. These pathways originate in higher brain regions, such as the periaqueductal gray (PAG) and the rostral ventromedial medulla (RVM) [22,30]. The PAG, in particular, acts as a key hub for pain modulation. Descending pathways exert both inhibitory and facilitatory influences on the transmission of pain signals [30,31]. Endogenous opioids, such as endorphins, are released in response to stress or injury, acting as natural pain relievers by binding to receptors in the spinal cord and dampening the transmission of nociceptive signals [32]. Alternatively, descending pathways can also enhance pain transmission, emphasizing the dynamic and nuanced nature of pain modulation [30].
2.3. Advances in Pain Management
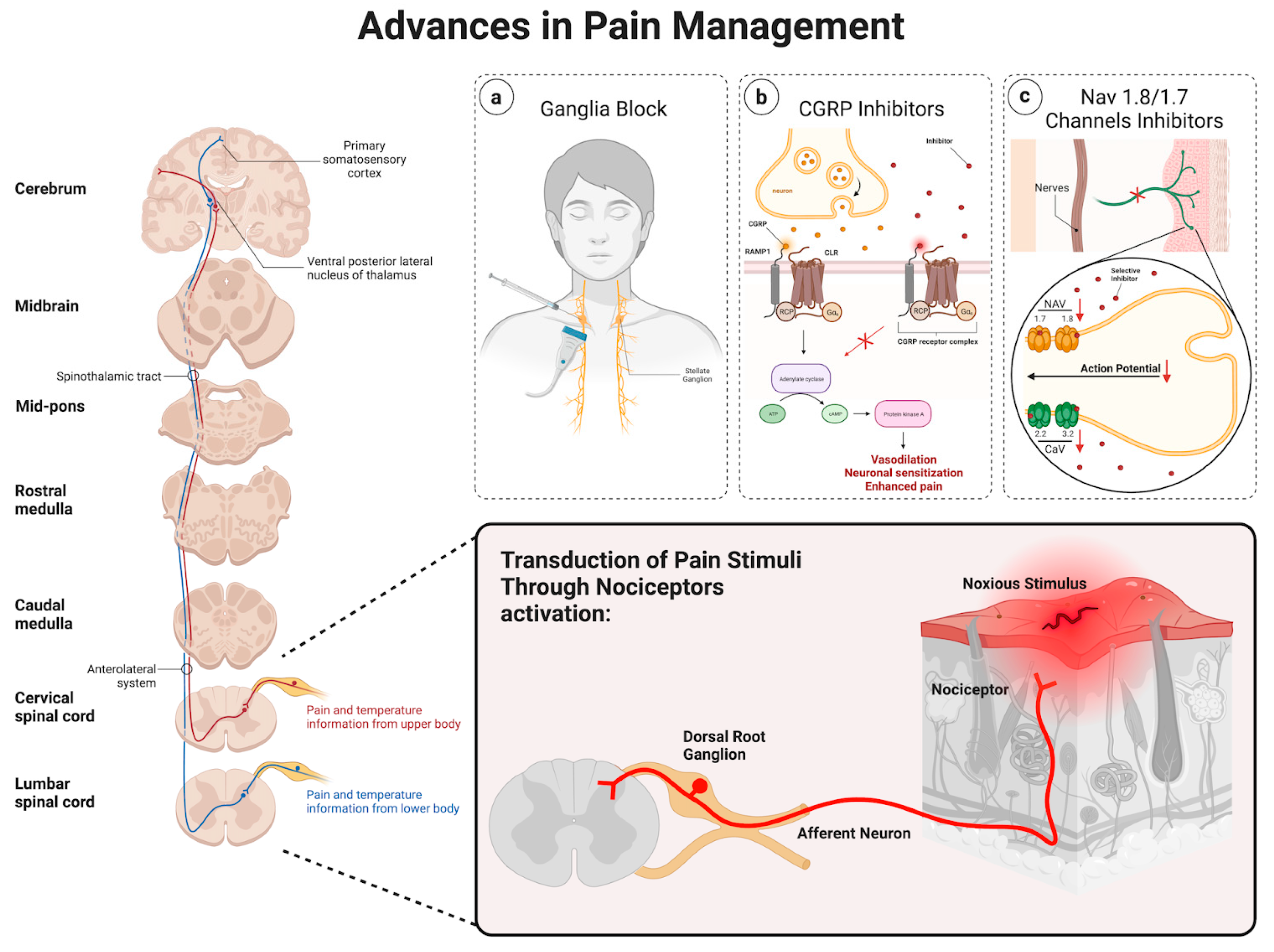
Chronic pain is maladaptive and remains a substantial burden for patients. In many cases, lifestyle modifications, as well as opiates, have been used as initial treatments. However, these approaches may prove ineffective or be linked to adverse systemic side effects, emphasizing the need for more precise and targeted interventional strategies. Thus, advancements in our understanding of pain perception and modulation allowed us to utilize different approaches, including ganglia injections, the role of calcitonin gene-related peptide (CGRP), and the involvement of Nav 1.8/1.7 channels, as promising targets in pain management, as shown in Figure 2.
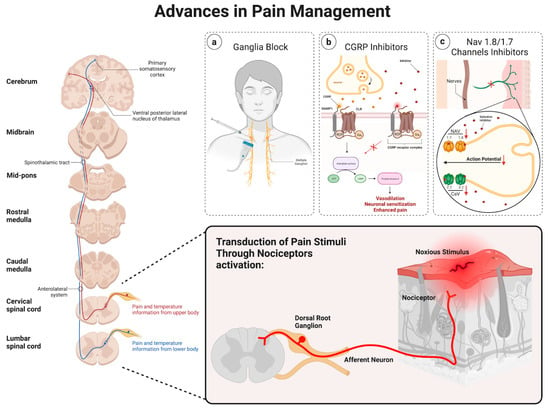
Figure 2.
Schematic of the various advancements in pain management.
2.4. Ganglia Injections
Ganglia injections are considered a promising approach when it comes to pain management. Sympathetic ganglia injections as a pain management approach have been described previously [33]. The mechanisms believed to contribute to sympathetic pain involve the impairment of inhibitory pain control as well as increased adrenergic excitability [33,34]. Thus, through the precise application of substances like local anesthetics (i.e., bupivacaine) or neuroleptic agents (i.e., alcohol) to specific ganglia, this method seeks to disrupt the positive feedback loop, thereby reducing central hyperexcitability and delivering a focused and targeted intervention [33]. Sympathetic blocks are increasingly used in managing both painful and nonpainful conditions, including postherpetic neuralgia [35], posttraumatic stress disorder (PTSD) [36], and hyperhidrosis [33]. Image-guided stellate ganglion injections have demonstrated effectiveness in addressing certain types of sympathetically maintained and visceral pain [35,36,37,38,39,40]. However, the lumbar sympathetic block is employed to address various types of pain, particularly those associated with the lower extremities, such as complex regional pain syndrome or sympathetic-mediated leg pain [41,42,43]. On the other hand, visceral abdominal and pelvic pain can be relieved by sympathetic blocks of the celiac plexus block [44,45,46,47,48,49], superior hypogastric [50,51,52,53,54], and ganglion impar [55,56,57,58]. This method holds potential for various pain conditions, offering a more precise and minimally invasive alternative to traditional systemic treatments.
2.5. Calcitonin Gene-Related Peptide
CGRP, a neuropeptide primarily found in sensory nerves C and Aδ [59,60], has gained attention for its pivotal role in pain modulation [61]. Along with its vasodilatory action [62], recent research suggests that CGRP plays a crucial part in mediating pain signals through the facilitation of nociceptive transmission as well as peripheral and central sensitization [63,64,65]. Thus, therapies targeting CGRP receptors are being explored as potential treatments for chronic pain conditions. For instance, CGRP has been described in migraine pathology [66], and the subsequent development of CGRP antagonists was proven successful in treating acute episodes of migraine [67]. Furthermore, the later development of monoclonal antibodies against CGRP showed promise in treating chronic migraines [68,69,70]. Currently, three monoclonal antibodies (mAbs) have received approval for the prevention and management of migraines. Among these, two are designed to target the CGRP peptide, while the third is specifically directed toward the CGRP receptor [71]. Thus, blocking CGRP signaling may offer a novel avenue for pain relief without the side effects associated with traditional analgesics.
2.6. NaV 1.8/1.7 Channels
Also, Nav 1.8 and Nav 1.7 channels, specific sodium channels found in sensory neurons, have been identified as key players in pain transmission [72]. It is believed that the increased activation and/or expression of NaV1.7/1.8 channels have a role in the development and persistence of many forms of neuropathic pain [73]. Thus, since these channels contribute to the generation and propagation of pain signals, their modulation represents a promising therapeutic strategy. Even though agents selectively blocking Nav1.3, Nav1.7, Nav1.8, Cav3.2, and HCN2, as well as activators of Kv7.2, have demonstrated efficacy in alleviating indicators of neuropathic pain within animal models, their translation to clinical application has yet to prove successful [73,74,75]. Certain compounds fall short of achieving therapeutic endpoints, while others exhibit dose-limiting side effects, thus impeding their successful clinical implementation. Efforts to develop selective inhibitors for NaV 1.7 and NaV 1.8 channels are underway, aiming to provide effective pain relief while minimizing adverse effects on other physiological processes [74].
The experience of pain is a multifaceted phenomenon intricately woven into the fabric of human perception. The interplay between ascending and descending pathways within the brain illustrates the sophisticated mechanisms that contribute to the sensation, interpretation, and modulation of pain. A comprehensive understanding of these pathways is crucial not only for unraveling the mysteries of pain but also for developing targeted interventions to alleviate suffering and improve the quality of life for individuals experiencing pain. Thus, the exploration of ganglia injections, CGRP, and Nav 1.8/1.7 channels as mechanisms in pain modulation signifies a growing understanding of the intricate processes involved in pain perception. These emerging avenues hold promise for the development of targeted and effective interventions for patients with acute or chronic pain.
3. Introduction to Various Nerve Blockers Used in Anesthesia
The peripheral nerve blocks terminate pain signals the cerebral cortex receives from the spinal cord. Perioperative anesthetic nerve blocks can manage pain after procedures and reduce the need for postsurgical opioid consumption [76] when administered along with general anesthesia or autonomously in less complex surgeries using ultrasound-guided techniques [77]. Diagnostic nerve blocks in chronic pain can determine the anatomical source of pain signals and provide therapeutic utility [78]. Nerve blocks can reduce inflammation and provide temporary pain relief for acute and chronic upper and lower extremity pain. Damage to a sympathetic nerve chain can be used as a target for sympathetic nerve blocks when autonomic function damage and sympathetically mediated pain (SMP) occurs. Stellate ganglion blocks further identify upper limb, head, and neck region nerve damage and block neural connections, improving the blood supply of the region and reducing adrenal hormone plasma concentration [79]. To diagnose facet joints as a source of pain, placebo-controlled zygapophysial blocks can be a cost-effective alternative to lumbar medial branch neurotomy [80]. We have summarized several types of common nerve blocks depending on the injury, clinical indications, and side effects in Table 1.

Table 1.
Summary of the various types of common nerve blocks depending on the injury, clinical indications, and side effects.
Impact of Lack of Neuroglia in Neurologic Disorders
The dysfunction or absence of neuroglial cells is associated with the development of certain neurological diseases. Astrocytes are neuroglial cells associated with several central nervous system diseases when damaged or absent. Chronic migraines are a disease characterized by astrocyte dysfunction [102]. Patients with chronic migraines have headaches for at least 15 days in a month, with eight of those migraines fulfilling migraine criteria [103]. The relationship between astrocyte dysfunction and the development of chronic migraines may have to do with calcitonin gene-related peptide (CGRP). CGRP acts on astrocyte receptors to modulate neuropathic pain. The interaction of CGRP on astrocyte receptors causes Histone H3 lysine 9 acetylation, which is associated with inflammatory gene expression [104]. Pain can be experienced as a result of the response to the inflammatory processes by glial cells.
In addition to migraines, Amyotrophic Lateral Sclerosis (ALS) is another disease associated with neuroglial dysfunction. ALS is a neurological disease characterized by upper and lower motor neuron damage. With time, this disease progressively leads to muscle atrophy and paralysis. Microglial cells are involved in the pathogenesis of ALS. The early stages of ALS are associated with a decrease in the number of microglial cells. Interestingly, one study by Gerber et al. showed that the number of astrocytes is not altered at early symptomatic stages, but their intraspinal repartition is modified at symptom onset. [105,106]. In addition to the association of microglia with ALS pathogenesis, microglia also have a role in pain modulation. Neuromodulators produced by microglia can affect synaptic pruning, inducing pain after tissue or nerve injury. In contrast, microglia have a role in pain resolution via a pathway that involves Cannabinoid receptor type 2 activation and MAP kinase phosphorylation [107]. Although pain is not a characteristic finding in ALS, the decreased number of microglial cells may affect their pain-modulation abilities.
Multiple sclerosis (MS) is another neurological disorder associated with glial cell dysfunction. MS is characteristically defined as a CNS demyelinating disorder due to an autoimmune response against myelin. This autoimmune response is typically associated with T-cells and B-cells, but microglia have been suspected to play a role as well. Activation of microglia leads to the release of cytotoxic nitrous oxide and superoxide radicals, causing CNS injury [108]. Additionally, oligodendrocyte injury is noted to occur in MS. A recent study supports a relationship between oligodendrocyte absence and pain after finding that genetic oligodendrocyte ablation rapidly triggers sensory changes that resemble central neuropathic pain [109,110]. This may explain the chronic pain symptoms experienced by some individuals with MS.
Similar to MS, Charcot–Marie–Tooth (CMT) disease is a demyelinating disorder of the nervous system. However, CMT differs in how it occurs due to dysfunction of the Schwann cells, which are located in the peripheral nervous system. Patients with CMT frequently experience pain or musculoskeletal deformities. More than one-third of patients with CMT manage their pain with analgesics such as NSAIDs or Acetaminophen [111]. Some patients may elect to undergo surgery to treat deformities and pain in their feet. The amount of studies reporting peripheral nerve blocks in patients with CMT is scarce, but one study evaluated the analgesic effect of a catheter-based sciatic nerve block in patients with CMT for postoperative pain control. The authors of this study concluded that peripheral nerve block in patients with CMT is safe and effective [112]. This is a reassuring finding, as patients with pre-existing neuropathies may experience complications from anesthesia [113].
Several of the discussed neurological diseases associated with glial cells have microglial dysfunction in common. As mentioned above, microglia have recently been recognized to have a significant role in the initiation of chronic pain. Advancing studies have proposed pharmacological and genetic modulation of microglia as a potential treatment option for patients with chronic pain. Even though there are no FDA-approved drugs that specifically target microglia, there are some available medications that have a degree of microglia modulation. These drugs can be used as analgesics for certain chronic pain syndromes [114]. Patients with neurological disorders involving microglial dysfunction may have unexpected effects with microglial-modulating drugs. Therefore, these drugs may not be an option for these individuals.
4. Future Developments in Alternative Nerve-Blockage Therapies
Botulinum neurotoxins (BoNTs) have been used for many years in patients with neuropathic pain [115]. BoNTs exert their effect by blocking the release of certain neurotransmitters, such as glutamate, substance P, and CGRP. Some of these neurotransmitters are associated with the activation of glial cells. Therefore, BoNTs also play a role in pain modulation by inhibiting the activation of glial cells associated with chronic pain [116].
Recent advances in the treatment of chronic neuropathic pain with paresthesia-free side effects include Spinal Cord Stimulation (SCS). Although this therapy suppresses central neuron excitability and causes a reduction in pain scores, it is associated with serious adverse effects of hematoma secondary to dural punctures. SCS is preferred over deep brain stimulation or motor cortex stimulation due to its moderate efficacy and minimal side effects [117]. Peripheral nerve stimulation has shown success in treating acute post-surgical pain by applying current to large-diameter myelinated afferent fibers and interfering with central pain signals. PNS can provide advantages in comparison to epidural local anesthetic injections for short-term pain relief without risk of infection, local anesthetic exhaustion, urinary retention, or motor weakness [112].
Novel, minimally invasive, painless therapies for alternative neuropathic pain include Transcranial Direct Current Stimulation (tDCS) and Remote Electrical Neuromodulation (REN) [118]. tDCS reduces overall pain intensity in diabetic polyneuropathy patients with MRI-guided targeted tissue ablation and magnetic stimulation of the motor cortex. [119] REN therapy is effective for the acute treatment of migraines by inducing conditioned pain modulation from upper arm peripheral nerves [120]. Immersive Virtual Reality, although challenging for clinicians with limited technological experience and expensive, was found by a preliminary study examining 3D mirror feedback therapy to significantly reduce chronic upper extremity neuropathic pain for a brief period [121]. Further investigations regarding the role of glial cell response to neuromodulation in IVR can identify target glial cells for other regions of pain. To prevent the degeneration of injured nerves and replace lost neural cells, stem cell regeneration with transplantation has been studied with GABAergic neuron transplants in an injured spinal cord and reduces clinical symptoms of hyperalgesia and spontaneous pain. [122] Understanding the role of neuroglia in the context of neuropathic pain and neurological disorders can aid in improving clinical outcomes and the challenging nature of managing pain.
There have been advances in various clinical trials for the potential to manipulate microglia for patients suffering from chronic pain. One drug called Minocycline is a microglial inhibitor that has been indicated for low back pain, but they are currently in the recruiting status of the study [114]. Another drug called Tetrahydrocannabinol is a cannabinoid-receptor agonist that serves to modulate pain. This clinical trial is currently active [114]. A completed clinical trial involves the use of low-dose Naltrexone (a TLR4 antagonist) for fibromyalgia [114]. However, this drug is still pending FDA approval. There was another clinical study filled with patients suffering from post-herpetic neuralgia [114]. Unfortunately, the effects of the drug to decrease pain had minimal impact on the majority of the patients.
In conclusion, the neuroglia plays an imperative role in administrating nerve blockers and anesthetic agents to patients. Our review focused on microglia having a role in pain resolution via various pathways, such as the activation of cannabinoid receptor type 2 and MAP kinase phosphorylation. We described the intricate relationship between cells that create the blood–brain barrier and the impact that anesthetic agents and nerve blocks have on them. Future studies do need to be conducted in order to further augment the science behind proposed pharmacological and genetic modulation of microglia as potential treatment options that can be offered to patients suffering from chronic pain.
Author Contributions
Conceptualization, A.P. and B.L.-W.; methodology, A.P. and B.L.-W.; validation, A.P. and B.L.-W.; formal analysis, A.P.; investigation, B.L.-W.; writing—original draft preparation, A.P., R.A.-B., R.T., D.P., D.F., J.B., M.P. and B.L.-W.; writing—review and editing, A.P., R.A.-B., R.T., D.P., D.F., J.B., M.P. and B.L.-W.; visualization, A.P.; supervision, B.L.-W. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gawdi, R.; Shumway, K.R.; Emmady, P.D. Physiology, Blood Brain Barrier. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Obermeier, B.; Verma, A.; Ransohoff, R.M. The blood-brain barrier. Handb. Clin. Neurol. 2016, 133, 39–59. [Google Scholar] [CrossRef]
- Cichorek, M.; Kowiański, P.; Lietzau, G.; Lasek, J.; Moryś, J. Neuroglia-development and role in physiological and pathophysiological processes. Folia Morphol. 2021, 80, 766–775. [Google Scholar] [CrossRef]
- Cekanaviciute, E.; Buckwalter, M.S. Astrocytes: Integrative Regulators of Neuroinflammation in Stroke and Other Neurological Diseases. Neurother. J. Am. Soc. Exp. NeuroTherapeutics 2016, 13, 685–701. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.D.; Bordey, A. The astrocyte odyssey. Prog. Neurobiol. 2008, 86, 342–367. [Google Scholar] [CrossRef] [PubMed]
- Daneman, R.; Prat, A. The blood-brain barrier. Cold Spring Harb. Perspect. Biol. 2015, 7, a020412. [Google Scholar] [CrossRef]
- Abbott, N.J.; Patabendige, A.A.; Dolman, D.E.; Yusof, S.R.; Begley, D.J. Structure and function of the blood-brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Cabezas, R.; Avila, M.; Gonzalez, J.; El-Bachá, R.S.; Báez, E.; García-Segura, L.M.; Jurado Coronel, J.C.; Capani, F.; Cardona-Gomez, G.P.; Barreto, G.E. Astrocytic modulation of blood brain barrier: Perspectives on Parkinson’s disease. Front. Cell. Neurosci. 2014, 8, 211. [Google Scholar] [CrossRef]
- Rochfort, K.D.; Cummins, P.M. The blood-brain barrier endothelium: A target for pro-inflammatory cytokines. Biochem. Soc. Trans. 2015, 43, 702–706. [Google Scholar] [CrossRef] [PubMed]
- Ronaldson, P.T.; Davis, T.P. Regulation of blood-brain barrier integrity by microglia in health and disease: A therapeutic opportunity. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2020, 40 (Suppl. S1), S6–S24. [Google Scholar] [CrossRef]
- Gullotta, G.S.; Costantino, G.; Sortino, M.A.; Spampinato, S.F. Microglia and the Blood-Brain Barrier: An External Player in Acute and Chronic Neuroinflammatory Conditions. Int. J. Mol. Sci. 2023, 24, 9144. [Google Scholar] [CrossRef]
- da Fonseca, A.C.; Matias, D.; Garcia, C.; Amaral, R.; Geraldo, L.H.; Freitas, C.; Lima, F.R. The impact of microglial activation on blood-brain barrier in brain diseases. Front. Cell. Neurosci. 2014, 8, 362. [Google Scholar] [CrossRef]
- Simons, M.; Nave, K.A. Oligodendrocytes: Myelination and Axonal Support. Cold Spring Harb. Perspect. Biol. 2015, 8, a020479. [Google Scholar] [CrossRef]
- Seo, J.H.; Maki, T.; Maeda, M.; Miyamoto, N.; Liang, A.C.; Hayakawa, K.; Pham, L.D.; Suwa, F.; Taguchi, A.; Matsuyama, T.; et al. Oligodendrocyte precursor cells support blood-brain barrier integrity via TGF-β signaling. PLoS ONE 2014, 9, e103174. [Google Scholar] [CrossRef]
- Obermeier, B.; Daneman, R.; Ransohoff, R.M. Development, maintenance and disruption of the blood-brain barrier. Nat. Med. 2013, 19, 1584–1596. [Google Scholar] [CrossRef]
- Attwell, D.; Mishra, A.; Hall, C.N.; O’Farrell, F.M.; Dalkara, T. What is a pericyte? J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2016, 36, 451–455. [Google Scholar] [CrossRef]
- Jäkel, S.; Dimou, L. Glial Cells and Their Function in the Adult Brain: A Journey through the History of Their Ablation. Front. Cell. Neurosci. 2017, 11, 24. [Google Scholar] [CrossRef]
- Stapor, P.C.; Sweat, R.S.; Dashti, D.C.; Betancourt, A.M.; Murfee, W.L. Pericyte dynamics during angiogenesis: New insights from new identities. J. Vasc. Res. 2014, 51, 163–174. [Google Scholar] [CrossRef]
- Gaceb, A.; Özen, I.; Padel, T.; Barbariga, M.; Paul, G. Pericytes secrete pro-regenerative molecules in response to platelet-derived growth factor-BB. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2018, 38, 45–57. [Google Scholar] [CrossRef]
- Brown, L.S.; Foster, C.G.; Courtney, J.M.; King, N.E.; Howells, D.W.; Sutherland, B.A. Pericytes and Neurovascular Function in the Healthy and Diseased Brain. Front. Cell. Neurosci. 2019, 13, 282. [Google Scholar] [CrossRef]
- Sonar, S.A.; Lal, G. Blood-brain barrier and its function during inflammation and autoimmunity. J. Leukoc. Biol. 2018, 103, 839–853. [Google Scholar] [CrossRef]
- De Ridder, D.; Adhia, D.; Vanneste, S. The anatomy of pain and suffering in the brain and its clinical implications. Neurosci. Biobehav. Rev. 2021, 130, 125–146. [Google Scholar] [CrossRef]
- Cohen, S.P.; Vase, L.; Hooten, W.M. Chronic pain: An update on burden, best practices, and new advances. Lancet 2021, 397, 2082–2097. [Google Scholar] [CrossRef] [PubMed]
- Thanh, N.X.; Tanguay, R.L.; Manhas, K.J.P.; Kania-Richmond, A.; Kashuba, S.; Geyer, T.; Pereira, J.X.; Wasylak, T. Economic burden of chronic pain in Alberta, Canada. PLoS ONE 2022, 17, e0272638. [Google Scholar] [CrossRef] [PubMed]
- Yong, R.J.; Mullins, P.M.; Bhattacharyya, N. Prevalence of chronic pain among adults in the United States. Pain 2022, 163, e328–e332. [Google Scholar] [CrossRef]
- Maniadakis, N.; Gray, A. The economic burden of back pain in the UK. Pain 2000, 84, 95–103. [Google Scholar] [CrossRef]
- Dubin, A.E.; Patapoutian, A. Nociceptors: The sensors of the pain pathway. J. Clin. Investig. 2010, 120, 3760–3772. [Google Scholar] [CrossRef] [PubMed]
- Lacroix-Fralish, M.L.; Mogil, J.S. Progress in genetic studies of pain and analgesia. Annu. Rev. Pharmacol. Toxicol. 2009, 49, 97–121. [Google Scholar] [CrossRef]
- Basbaum, A.I.; Bautista, D.M.; Scherrer, G.; Julius, D. Cellular and molecular mechanisms of pain. Cell 2009, 139, 267–284. [Google Scholar] [CrossRef]
- Bourne, S.; Machado, A.G.; Nagel, S.J. Basic anatomy and physiology of pain pathways. Neurosurg. Clin. N. Am. 2014, 25, 629–638. [Google Scholar] [CrossRef]
- Millan, M.J. Descending control of pain. Prog. Neurobiol. 2002, 66, 355–474. [Google Scholar] [CrossRef]
- Bagley, E.E.; Ingram, S.L. Endogenous opioid peptides in the descending pain modulatory circuit. Neuropharmacology 2020, 173, 108131. [Google Scholar] [CrossRef]
- Gunduz, O.H.; Kenis-Coskun, O. Ganglion blocks as a treatment of pain: Current perspectives. J. Pain Res. 2017, 10, 2815–2826. [Google Scholar] [CrossRef]
- Ramer, M.S.; Bisby, M.A. Adrenergic innervation of rat sensory ganglia following proximal or distal painful sciatic neuropathy: Distinct mechanisms revealed by anti-NGF treatment. Eur. J. Neurosci. 1999, 11, 837–846. [Google Scholar] [CrossRef]
- Makharita, M.Y.; Amr, Y.M.; El-Bayoumy, Y. Effect of early stellate ganglion blockade for facial pain from acute herpes zoster and incidence of postherpetic neuralgia. Pain Physician 2012, 15, 467–474. [Google Scholar]
- Lipov, E.; Ritchie, E.C. A review of the use of stellate ganglion block in the treatment of PTSD. Curr. Psychiatry Rep. 2015, 17, 599. [Google Scholar] [CrossRef]
- O’Connell, N.E.; Wand, B.M.; Gibson, W.; Carr, D.B.; Birklein, F.; Stanton, T.R. Local anaesthetic sympathetic blockade for complex regional pain syndrome. Cochrane Database Syst. Rev. 2016, 7, CD004598. [Google Scholar] [CrossRef]
- Abdi, S.; Zhou, Y.; Patel, N.; Saini, B.; Nelson, J. A new and easy technique to block the stellate ganglion. Pain Physician 2004, 7, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Ackerman, W.E.; Zhang, J.M. Efficacy of stellate ganglion blockade for the management of type 1 complex regional pain syndrome. South. Med. J. 2006, 99, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.D.; Jung, S.S.; Kim, H.S.; Yun, D.H.; Kim, D.H.; Chon, J.; Hong, D.W. Efficacy of ultrasonography guided stellate ganglion blockade in the stroke patients with complex regional pain syndrome. Ann. Rehabil. Med. 2012, 36, 633–639. [Google Scholar] [CrossRef]
- Meier, P.M.; Zurakowski, D.; Berde, C.B.; Sethna, N.F. Lumbar sympathetic blockade in children with complex regional pain syndromes: A double blind placebo-controlled crossover trial. Anesthesiology 2009, 111, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Abramov, R. Lumbar sympathetic treatment in the management of lower limb pain. Curr. Pain Headache Rep. 2014, 18, 403. [Google Scholar] [CrossRef]
- Chaturvedi, A.; Dash, H.H. Sympathetic blockade for the relief of chronic pain. J. Indian Med. Assoc. 2001, 99, 698–703. [Google Scholar] [PubMed]
- Erdine, S. Celiac ganglion block. Agri Agri (Algoloji) Dernegi’nin Yayin. Organidir = J. Turk. Soc. Algol. 2005, 17, 14–22. [Google Scholar]
- Nitschke, A.M.; Ray, C.E., Jr. Percutaneous neurolytic celiac plexus block. Semin. Interv. Radiol. 2013, 30, 318–321. [Google Scholar] [CrossRef]
- Wang, P.J.; Shang, M.Y.; Qian, Z.; Shao, C.W.; Wang, J.H.; Zhao, X.H. CT-guided percutaneous neurolytic celiac plexus block technique. Abdom. Imaging 2006, 31, 710–718. [Google Scholar] [CrossRef]
- Yasuda, I.; Wang, H.P. Endoscopic ultrasound-guided celiac plexus block and neurolysis. Dig. Endosc. Off. J. Jpn. Gastroenterol. Endosc. Soc. 2017, 29, 455–462. [Google Scholar] [CrossRef]
- Eisenberg, E.; Carr, D.B.; Chalmers, T.C. Neurolytic celiac plexus block for treatment of cancer pain: A meta-analysis. Anesth. Analg. 1995, 80, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Rana, M.V.; Candido, K.D.; Raja, O.; Knezevic, N.N. Celiac plexus block in the management of chronic abdominal pain. Curr. Pain Headache Rep. 2014, 18, 394. [Google Scholar] [CrossRef]
- Bosscher, H. Blockade of the superior hypogastric plexus block for visceral pelvic pain. Pain Pract. Off. J. World Inst. Pain 2001, 1, 162–170. [Google Scholar] [CrossRef]
- Waldman, S.D.; Wilson, W.L.; Kreps, R.D. Superior hypogastric plexus block using a single needle and computed tomography guidance: Description of a modified technique. Reg. Anesth. 1991, 16, 286–287. [Google Scholar]
- Plancarte, R.; Amescua, C.; Patt, R.B.; Aldrete, J.A. Superior hypogastric plexus block for pelvic cancer pain. Anesthesiology 1990, 73, 236–239. [Google Scholar] [CrossRef]
- de Leon-Casasola, O.A.; Kent, E.; Lema, M.J. Neurolytic superior hypogastric plexus block for chronic pelvic pain associated with cancer. Pain 1993, 54, 145–151. [Google Scholar] [CrossRef]
- Plancarte, R.; de Leon-Casasola, O.A.; El-Helaly, M.; Allende, S.; Lema, M.J. Neurolytic superior hypogastric plexus block for chronic pelvic pain associated with cancer. Reg. Anesth. 1997, 22, 562–568. [Google Scholar]
- Scott-Warren, J.T.; Hill, V.; Rajasekaran, A. Ganglion impar blockade: A review. Curr. Pain Headache Rep. 2013, 17, 306. [Google Scholar] [CrossRef]
- Wemm, K., Jr.; Saberski, L. Modified approach to block the ganglion impar (ganglion of Walther). Reg. Anesth. 1995, 20, 544–545. [Google Scholar] [PubMed]
- Foye, P.M.; Buttaci, C.J.; Stitik, T.P.; Yonclas, P.P. Successful injection for coccyx pain. Am. J. Phys. Med. Rehabil. 2006, 85, 783–784. [Google Scholar] [CrossRef] [PubMed]
- Gunduz, O.H.; Sencan, S.; Kenis-Coskun, O. Pain Relief due to Transsacrococcygeal Ganglion Impar Block in Chronic Coccygodynia: A Pilot Study. Pain Med. 2015, 16, 1278–1281. [Google Scholar] [CrossRef] [PubMed]
- Lawson, S.N.; Crepps, B.; Perl, E.R. Calcitonin gene-related peptide immunoreactivity and afferent receptive properties of dorsal root ganglion neurones in guinea-pigs. J. Physiol. 2002, 540 Pt 3, 989–1002. [Google Scholar] [CrossRef] [PubMed]
- Ruscheweyh, R.; Forsthuber, L.; Schoffnegger, D.; Sandkühler, J. Modification of classical neurochemical markers in identified primary afferent neurons with Abeta-, Adelta-, and C-fibers after chronic constriction injury in mice. J. Comp. Neurol. 2007, 502, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Iyengar, S.; Ossipov, M.H.; Johnson, K.W. The role of calcitonin gene-related peptide in peripheral and central pain mechanisms including migraine. Pain 2017, 158, 543–559. [Google Scholar] [CrossRef] [PubMed]
- Brain, S.D.; Williams, T.J.; Tippins, J.R.; Morris, H.R.; MacIntyre, I. Calcitonin gene-related peptide is a potent vasodilator. Nature 1985, 313, 54–56. [Google Scholar] [CrossRef]
- Wimalawansa, S.J. Calcitonin gene-related peptide and its receptors: Molecular genetics, physiology, pathophysiology, and therapeutic potentials. Endocr. Rev. 1996, 17, 533–585. [Google Scholar] [CrossRef]
- Benemei, S.; Nicoletti, P.; Capone, J.G.; Geppetti, P. CGRP receptors in the control of pain and inflammation. Curr. Opin. Pharmacol. 2009, 9, 9–14. [Google Scholar] [CrossRef]
- Russell, F.A.; King, R.; Smillie, S.J.; Kodji, X.; Brain, S.D. Calcitonin gene-related peptide: Physiology and pathophysiology. Physiol. Rev. 2014, 94, 1099–1142. [Google Scholar] [CrossRef]
- Edvinsson, L. CGRP and migraine: From bench to bedside. Rev. Neurol. 2021, 177, 785–790. [Google Scholar] [CrossRef]
- Olesen, J.; Diener, H.C.; Husstedt, I.W.; Goadsby, P.J.; Hall, D.; Meier, U.; Pollentier, S.; Lesko, L.M.; BIBN 4096 BS Clinical Proof of Concept Study Group. Calcitonin gene-related peptide receptor antagonist BIBN 4096 BS for the acute treatment of migraine. N. Engl. J. Med. 2004, 350, 1104–1110. [Google Scholar] [CrossRef] [PubMed]
- Bigal, M.E.; Dodick, D.W.; Krymchantowski, A.V.; VanderPluym, J.H.; Tepper, S.J.; Aycardi, E.; Loupe, P.S.; Ma, Y.; Goadsby, P.J. TEV-48125 for the preventive treatment of chronic migraine: Efficacy at early time points. Neurology 2016, 87, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Bigal, M.E.; Edvinsson, L.; Rapoport, A.M.; Lipton, R.B.; Spierings, E.L.; Diener, H.C.; Burstein, R.; Loupe, P.S.; Ma, Y.; Yang, R.; et al. Safety, tolerability, and efficacy of TEV-48125 for preventive treatment of chronic migraine: A multicentre, randomised, double-blind, placebo-controlled, phase 2b study. Lancet Neurol. 2015, 14, 1091–1100. [Google Scholar] [CrossRef] [PubMed]
- Bigal, M.E.; Escandon, R.; Bronson, M.; Walter, S.; Sudworth, M.; Huggins, J.P.; Garzone, P. Safety and tolerability of LBR-101, a humanized monoclonal antibody that blocks the binding of CGRP to its receptor: Results of the Phase 1 program. Cephalalgia Int. J. Headache 2014, 34, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Edvinsson, L.; Haanes, K.A.; Warfvinge, K.; Krause, D.N. CGRP as the target of new migraine therapies-successful translation from bench to clinic. Nat. Rev. Neurol. 2018, 14, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, G.; McMahon, S.B. The physiological function of different voltage-gated sodium channels in pain. Nat. Rev. Neurosci. 2021, 22, 263–274. [Google Scholar] [CrossRef]
- Alles, S.R.A.; Smith, P.A. Peripheral Voltage-Gated Cation Channels in Neuropathic Pain and Their Potential as Therapeutic Targets. Front. Pain Res. 2021, 2, 750583. [Google Scholar] [CrossRef]
- Hameed, S. Nav1.7 and Nav1.8: Role in the pathophysiology of pain. Mol. Pain 2019, 15, 1744806919858801. [Google Scholar] [CrossRef] [PubMed]
- Emery, E.C.; Luiz, A.P.; Wood, J.N. Nav1.7 and other voltage-gated sodium channels as drug targets for pain relief. Expert Opin. Ther. Targets 2016, 20, 975–983. [Google Scholar] [CrossRef]
- Stewart, J.W.; Dickson, D.; Van Hal, M.; Aryeetey, L.; Sunna, M.; Schulz, C.; Alexander, J.C.; Gasanova, I.; Joshi, G.P. Ultrasound-guided erector spinae plane blocks for pain management after open lumbar laminectomy. Eur. Spine J. 2023. Correction in Eur. Spine J. 2023. [Google Scholar] [CrossRef]
- Mehlmann, F.M.G.; Ferraro, L.H.C.; Sousa, P.C.C.B.D.; Cunha, G.P.; Bergamaschi, E.C.Q.A.; Takeda, A. Ultrasound-guided selective nerve blocks for trigger finger surgeries to maintain flexion/extension of fingers-Case series. Braz. J. Anesthesiol. 2019, 69, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Curatolo, M.; Bogduk, N. Diagnostic blocks for chronic pain. Scand. J. Pain. 2010, 1, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Piraccini, E.; Munakomi, S.; Chang, K.V. Stellate Ganglion Blocks. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK507798/ (accessed on 24 December 2023).
- Bogduk, N. On diagnostic blocks for lumbar zygapophysial joint pain. F1000 Med. Rep. 2010, 2, 57. [Google Scholar] [CrossRef]
- Gupta, R.; Madanat, L.; Jindal, V.; Gaikazian, S. Celiac Plexus Block Complications: A Case Report and Review of the Literature. J. Palliat. Med. 2021, 24, 1409–1412. [Google Scholar] [CrossRef]
- Erdine, S. Complications of Splanchnic and Celiac Plexus Block. In Complications of Pain-Relieving Procedures; Erdine, S., Staats, P.S., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2022. [Google Scholar] [CrossRef]
- John, R.S.; Dixon, B.; Shienbaum, R. Celiac Plexus Block. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK531469/ (accessed on 24 December 2023).
- Avila Hernandez, A.N.; Singh, P. Epidural Anesthesia. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK542219/ (accessed on 24 December 2023).
- Kompoliti, K.; Horn, S.S. Chapter 55—Drug-Induced and Iatrogenic Neurological Disorders. In Textbook of Clinical Neurology, 3rd ed.; Goetz, C.G., Ed.; W.B. Saunders: Philadelphia, PA, USA, 2007; pp. 1285–1318. [Google Scholar] [CrossRef]
- Bhattaram, S.; Shinde, V.S. Novel use of motor-sparing genicular nerve blocks for knee injuries in the emergency department. Am. J. Emerg. Med. 2024, 75, 196.e1–196.e4. [Google Scholar] [CrossRef] [PubMed]
- Dass, R.M.; Kim, E.; Kim, H.K.; Lee, J.Y.; Lee, H.J.; Rhee, S.J. Alcohol neurolysis of genicular nerve for chronic knee pain. Korean J. Pain. 2019, 32, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Choi, S.S.; Yoon, S.H.; So-Hee, L.; Seo, D.K.; Lee, I.G.; Woo-Jong, C.; Jin-Woo, S. Ultrasound-Guided Genicular Nerve Block for Knee Osteoarthritis: A Double-Blind, Randomized Controlled Trial of Local Anesthetic Alone or in Combination with Corticosteroid. Pain Physician 2018, 21, 41–52. [Google Scholar] [PubMed]
- Baxter, C.S.; Singh, A.; Ajib, F.A.; Fitzgerald, B.M. Intercostal Nerve Block. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK482273/ (accessed on 24 December 2023).
- Guerra-Londono, C.E.; Privorotskiy, A.; Cozowicz, C.; Hicklen, R.S.; Memtsoudis, S.G.; Mariano, E.R.; Cata, J.P. Assessment of Intercostal Nerve Block Analgesia for Thoracic Surgery: A Systematic Review and Meta-analysis. JAMA Netw. Open 2021, 4, e2133394. [Google Scholar] [CrossRef] [PubMed]
- Alexander, C.E.; De Jesus, O.; Varacallo, M. Lumbar Sympathetic Block. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK431107/ (accessed on 24 December 2023).
- Carroll, I.; Clark, J.D.; Mackey, S. Sympathetic block with botulinum toxin to treat complex regional pain syndrome. Ann. Neurol. 2009, 65, 348–351. [Google Scholar] [CrossRef] [PubMed]
- Barreto Junior, E.P.S.; Nascimento, J.D.S.; Castro, A.P.C.R. Neurolytic block of the lumbar sympathetic chain improves chronic pain in a patient with critical lower limb ischemia. Braz. J. Anesthesiol. 2018, 68, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Austin, M.; Hinson, M.R. Occipital Nerve Block. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK580523/ (accessed on 24 December 2023).
- Castillo-Álvarez, F.; Hernando de la Bárcena, I.; Marzo-Sola, M.E. Greater occipital nerve block in the treatment of headaches. Review of evidence. Med. Clín. (Engl. Ed.) 2023, 161, 113–118. [Google Scholar] [CrossRef]
- Ghanavatian, S.; Leslie, S.W.; Derian, A. Pudendal Nerve Block. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK551518/ (accessed on 24 December 2023).
- Cok, O.Y.; Eker, H.E.; Cok, T.; Akin, S.; Aribogan, A.; Arslan, G. Transsacral S2-S4 Nerve Block For Vaginal Pain Due To Pudendal Neuralgia. J. Minim. Invasive Gynecol. 2011, 18, 401–404. [Google Scholar] [CrossRef]
- Goel, V.; Patwardhan, A.M.; Ibrahim, M.; Howe, C.L.; Schultz, D.M.; Shankar, H. Complications associated with stellate ganglion nerve block: A systematic review. Reg. Anesth. Pain Med. 2019. [Google Scholar] [CrossRef]
- Jeon, Y. Therapeutic potential of stellate ganglion block in orofacial pain: A mini-review. J. Dent. Anesth. Pain Med. 2016, 16, 159–163. [Google Scholar] [CrossRef]
- Jacques, N.; Karoutsos, S.; Marais, L.; Nathan-Denizot, N. Quality of life after trigeminal nerve block in refractory trigeminal neuralgia: A retrospective cohort study and literature review. J. Int. Med. Res. 2022, 50, 3000605221132027. [Google Scholar] [CrossRef]
- Kumar, A.; Sinha, C.; Kumar, A.; Kumari, P.; Mukul, S.K. Ultrasound-guided trigeminal nerve block and its comparison with conventional analgesics in patients undergoing faciomaxillary surgery: Randomised control trial. Indian J. Anaesth. 2018, 62, 871–875. [Google Scholar] [CrossRef] [PubMed]
- Vila-Pueyo, M.; Gliga, O.; Gallardo, V.J.; Pozo-Rosich, P. The Role of Glial Cells in Different Phases of Migraine: Lessons from Preclinical Studies. Int. J. Mol. Sci. 2023, 24, 12553. [Google Scholar] [CrossRef]
- Mungoven, T.J.; Henderson, L.A.; Meylakh, N. Chronic Migraine Pathophysiology and Treatment: A Review of Current Perspectives. Front. Pain Res. 2021, 2, 705276. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; An, Q.; Li, R.; Chen, S.; Gu, X.; An, S.; Wang, Z. Calcitonin gene-related peptide induces the histone H3 lysine 9 acetylation in astrocytes associated with neuroinflammation in rats with neuropathic pain. CNS Neurosci. Ther. 2021, 27, 1409–1424. [Google Scholar] [CrossRef]
- E Hirbec, H.; Noristani, H.N.; Perrin, F.E. Microglia Responses in Acute and Chronic Neurological Diseases: What Microglia-Specific Transcriptomic Studies Taught (and did Not Teach) Us. Front. Aging Neurosci. 2017, 9, 227. [Google Scholar] [CrossRef]
- Gerber, Y.N.; Sabourin, J.C.; Rabano, M.; Vivanco, M.; Perrin, F.E. Early functional deficit and microglial disturbances in a mouse model of amyotrophic lateral sclerosis. PLoS ONE 2012, 7, e36000. [Google Scholar] [CrossRef]
- Chen, G.; Zhang, Y.Q.; Qadri, Y.J.; Serhan, C.N.; Ji, R.R. Microglia in Pain: Detrimental and Protective Roles in Pathogenesis and Resolution of Pain. Neuron 2018, 100, 1292–1311. [Google Scholar] [CrossRef] [PubMed]
- Tafti, D.; Ehsan, M.; Xixis, K.L. Multiple Sclerosis. [Updated 7 September 2022]. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK499849/ (accessed on 2 January 2024).
- Murphy, K.L.; Bethea, J.R.; Fischer, R. Neuropathic Pain in Multiple Sclerosis—Current Therapeutic Intervention and Future Treatment Perspectives. In Multiple Sclerosis: Perspectives in Treatment and Pathogenesis [Internet]; Zagon, I.S., McLaughlin, P.J., Eds.; Codon Publications: Brisbane, AU, USA, 27 November 2017; Chapter 4. Available online: https://www.ncbi.nlm.nih.gov/books/NBK470151/ (accessed on 2 January 2024). [CrossRef]
- Gritsch, S.; Lu, J.; Thilemann, S.; Wörtge, S.; Möbius, W.; Bruttger, J.; Karram, K.; Ruhwedel, T.; Blanfeld, M.; Vardeh, D.; et al. Oligodendrocyte ablation triggers central pain independently of innate or adaptive immune responses in mice. Nat. Commun. 2014, 5, 5472. [Google Scholar] [CrossRef]
- Pisciotta, C.; Saveri, P.; Pareyson, D. Challenges in Treating Charcot-Marie-Tooth Disease and Related Neuropathies: Current Management and Future Perspectives. Brain Sci. 2021, 11, 1447. [Google Scholar] [CrossRef]
- Kaye, A.D.; Ridgell, S.; Alpaugh, E.S.; Mouhaffel, A.; Kaye, A.J.; Cornett, E.M.; Chami, A.A.; Shah, R.; Dixon, B.M.; Viswanath, O.; et al. Peripheral Nerve Stimulation: A Review of Techniques and Clinical Efficacy. Pain Ther. 2021, 10, 961–972. [Google Scholar] [CrossRef]
- Schmitt, H.J.; Huberth, S.; Huber, H.; Münster, T. Catheter-based distal sciatic nerve block in patients with Charcot-Marie-Tooth disease. BMC Anesthesiol. 2014, 14, 8. [Google Scholar] [CrossRef]
- Haight, E.S.; Forman, T.E.; Cordonnier, S.A.; James, M.L.; Tawfik, V.L. Microglial Modulation as a Target for Chronic Pain: From the Bench to the Bedside and Back. Anesth. Analg. 2019, 128, 737–746. [Google Scholar] [CrossRef]
- Park, J.; Park, H.J. Botulinum Toxin for the Treatment of Neuropathic Pain. Toxins 2017, 9, 260. [Google Scholar] [CrossRef] [PubMed]
- Luvisetto, S. Botulinum Neurotoxins beyond Neurons: Interplay with Glial Cells. Toxins 2022, 14, 704. [Google Scholar] [CrossRef] [PubMed]
- Crosby, N.D.; Janik, J.J.; Grill, W.M. Modulation of activity and conduction in single dorsal column axons by kilohertz-frequency spinal cord stimulation. J. Neurophysiol. 2017, 117, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Hebl, J.R.; Horlocker, T.T.; Schroeder, D.R. Neuraxial anesthesia and analgesia in patients with preexisting central nervous system disorders. Anesth. Analg. 2006, 103, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Jeanmonod, D.; Werner, B.; Morel, A.; Michels, L.; Zadicario, E.; Schiff, G.; Martin, E. Transcranial magnetic resonance imaging-guided focused ultrasound: Noninvasive central lateral thalamotomy for chronic neuropathic pain. Neurosurg. Focus 2012, 32, E1. [Google Scholar] [CrossRef] [PubMed]
- Ailani, J.; Rabany, L.; Tamir, S.; Ironi, A.; Starling, A. Real-World Analysis of Remote Electrical Neuromodulation (REN) for the Acute Treatment of Migraine. Front. Pain Res. 2022, 2, 753736. [Google Scholar] [CrossRef] [PubMed]
- Mouraux, D.; Brassinne, E.; Sobczak, S.; Nonclercq, A.; Warzée, N.; Sizer, P.S.; Tuna, T.; Penelle, B. 3D augmented reality mirror visual feedback therapy applied to the treatment of persistent, unilateral upper extremity neuropathic pain: A preliminary study. J. Man. Manip. Ther. 2017, 25, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Chakravarthy, K.; Chen, Y.; He, C.; Christo, P.J. Stem Cell Therapy for Chronic Pain Management: Review of Uses, Advances, and Adverse Effects. Pain Physician 2017, 20, 293–305. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).