Abstract
Fire injury stresses Douglas-fir trees (Pseudotsuga menziesii) that survive a wildfire event, allowing subsequent Douglas-fir beetle (Dendroctonus pseudotsugae) infection to kill trees that may have otherwise survived. This study aimed to determine how fire injury, stand, and tree characteristics drive Douglas-fir beetle host tree selection five years post-fire. We paired 28 adjacent beetle-infected and uninfected stands (infected N = 14) and 140 Douglas-fir trees (infected N = 70) within the 416 Fire burn area in Southwest Colorado. We found no statistically significant differences between infected and uninfected stand characteristics. Individual tree height, DBH, and bark char severity index were significantly higher in infected versus uninfected trees. We created a regression decision tree model to determine the influence of fire injury and tree characteristics on the probability of infection. Trees with a height ≥ 27 m, bark char height < 2.3 m, and DBH < 80 cm had the greatest probability of attack (100%). Trees with a height < 27 m, bark char severity index < 5.5, and DBH < 49 cm had the lowest probability of attack (3.7%). Understanding the influence of fire on Douglas-fir beetle host selection allows land managers to model potential epidemic outbreaks and guide proactive management actions that may reduce beetle outbreak severity or preserve high-value trees not killed by fire.
1. Introduction
Fire and insect outbreaks are major disturbances that affect the structure and function of mixed conifer forests. Mixed conifer forests of the Southwestern United States represent diverse forest assemblages and ecotypes. These forests can be classified along a spectrum from warm/dry, dominated by ponderosa pine (Pinus ponderosa); cool/moist, dominated by Douglas-fir (Pseudotsuga menziesii); to cold/wet, dominated by Engelmann spruce (Picea engelmanii), blue spruce (Picea pungens), and subalpine fir (Abies lasiocarpa) [1]. Historically, mixed conifer forests experienced diverse fire regimes depending on their forest type, with frequent (4 to 30 years) low-intensity surface fires in warm/dry mixed conifer, less frequent (35 to 100+ years) mixed-severity fires in cool/moist mixed conifer and stand-replacing fires occurring relatively infrequently (150 to 300+ years) in cold/wet mixed conifer forests [1,2]. Various native defoliating and phloem-feeding insects have also historically affected mixed conifer forests, occasionally erupting from endemic to epidemic population levels and influencing forest composition and structure [3,4]. With Euro-American settlement of the Rocky Mountain West in the mid-to-late 1800s, grazing, logging, fire suppression, and a trending warming climate over the past few decades has led to widespread changes in warm/dry and cool/moist mixed conifer disturbance regimes [1,5]. Denser, more homogenous forests, moisture stress and shorter, warmer winters have resulted in more frequent, intense, and expansive fires and insect outbreaks [4,6]. Fire and insect disturbances can interact, impacting mixed conifer forests depending on the disturbances’ sequential, temporal, and spatial characteristics [7]. Understanding the mechanisms driving fire/insect disturbance interactions is crucial to predicting future mixed conifer trajectories and implementing post-disturbance forest management.
Douglas-fir and Douglas-fir beetle (Dentroctonus pseudotsugae) are prevalent species in mixed conifer forests. Douglas-fir is widely distributed across Western North America, is the dominant tree species found in cool/moist mixed conifer forests, and is often found alongside ponderosa pine in warm/dry mixed conifer forests [8]. Douglas-fir is economically important as a timber source and provides a habitat for a range of wildlife [9]. Douglas-fir beetle parasitizes Douglas-fir, laying eggs under the bark and feeding on the phloem during its larval stage, with phloem destruction often resulting in tree death [10]. Douglas-fir beetles are native to the Rocky Mountain West and typically found in low-endemic populations. At endemic levels, Douglas-fir beetles tend to attack recently downed trees, such as wind throw, and trees that are stressed from disease, drought, or defoliating insects [11,12,13]. Given favorable conditions, such as abundant downed or weakened trees, Douglas-fir beetle populations can quickly increase to epidemic levels and begin attacking and overwhelming vigorously growing trees, causing large-scale Douglas-fir mortality, altering stand and understory composition and structure, and reducing Douglas-fir abundance [3,14,15].
Fire also plays a vital role in Douglas-fir forest dynamics. Historically, Douglas-fir-dominated forests experienced periodic mixed severity fires every 35 to 100+ years, with patches of low-severity surface fires interspersed with high-severity stand-replacing crown fires, creating a mosaic of stands with various age classes and overstory and understory communities [1]. Mature Douglas-fir possesses several physiological adaptations to withstand low- and medium-intensity fires, such as cork-like bark that grows thicker as the tree ages; a deep tap root system that protects the cambium and roots from heat injury; and lower branches that shed as the tree grows taller, making it harder for flames to enter the canopy. However, immature Douglas-fir lacks these adaptations and easily succumbs to fire [16]. Despite their adaptations, Douglas-firs that survive a fire can be significantly stressed due to partial cambium, root, and crown injuries. Beetles actively target stressed trees, which may be less capable of fending off beetle infection [12,17,18]. Trees that survive an initial fire event may die due to subsequent beetle infection [19,20].
Understanding Douglas-fir beetle’s host-tree selection has been an active area of research, as it is a major factor in the interaction of beetle/fire disturbances and can be used to predict the extent of tree mortality and stand replacement in the years following a fire [12,18,19,20]. Studies have found that various tree-, stand-, and landscape-level variables can increase the likelihood of Douglas-fir beetle attacks and the potential for epidemic outbreaks. Some studies have found increasing tree diameter at breast height (DBH), tree height, age, phloem thickness, and bark thickness to coincide with increased rates of beetle attack [12,21]. Trees experiencing reduced growth rates in recent years, perhaps due to stressors such as drought, have also been found to be more susceptible [12,21]. On a stand level, higher Douglas-fir basal area and higher overall stand density increased the odds of beetle infection [12,22,23], and landscape-level topographical features that create drier conditions, such as aspect, slope, and elevation, correlate with increased Douglas-fir beetle infestations [13].
Research on Douglas-fir beetle/fire interaction has found that in the first-year post-fire, DBH, stand density, and tree height remain important predictors of beetle infestation and that beetles prefer trees with a moderate degree of fire injury to their bole and high levels of crown scorch [17,18,19,20]. In successive years post-fire, as large fire-injured trees become depleted and beetle populations increase, beetles gradually colonize smaller, lightly fire-injured, and unburned green trees [17,19].
Other studies on Douglas-fir beetle’s host-tree selection have been conducted in unburned stands or years immediately after a fire event [17,18,19]. Our research objective was to determine if Douglas-fir beetle’s host-tree selection patterns in mixed conifer forests are influenced by burn severity, fire injury, stand structure, or individual tree characteristics in Southwestern Colorado’s 416 Fire burn area five years post-fire. We then utilized these infection data to develop a model that land managers may use to guide proactive management actions. Post-fire tree mortality is an important factor that land managers must consider when attempting to establish desired forest conditions [7,24,25], and the interactions between fire and tree-boring insects must be included in these considerations. Actions such as the deployment of anti-aggregation hormone packets or the targeted removal of high-risk trees may reduce the likelihood of epidemic outbreaks and preserve individual high economic or social value trees [26,27,28,29]. Using a machine learning methodology, we developed a recursive partitioning decision tree model that uses fire injury, tree, and stand characteristics as parameters. This model may be used by land managers to assess the probability that a tree will be infected by Douglas-fir beetles post-fire and evaluate the need for management intervention.
2. Methods
Our study area was located approximately 21 km north of Durango, Colorado, in the southern portion of the San Juan National Forest adjacent to Hermosa Creek within the Hermosa Special Management Area and Hermosa Wilderness [30]. The study area ranges in elevation from 2277 m to 2470 m on steep slopes that range from 30 to 45 degrees. Average daily temperatures range from a maximum of 26.7 °C in July to a minimum of −12.2 °C in January.
Average annual precipitation is 58.4 cm, with the greatest amounts occurring in July and August due to summer thunderstorm activity. Precipitation from November to March is dominated by snowfall (Western Regional Climate Center, Tacoma, 1971–2000, www.wrcc.dri.edu accessed on 3 July 2019). Forest types in the study area vary from ponderosa pine and pine-oak forest to warm/dry and cool/moist mixed conifer. Quaking aspen (Populus tremuloides) is present in the study area, and continuous aspen stands exist adjacent to plots; however, quaking aspen is only a minor component of overstory trees in plots. The study area has never been logged and has a high proportion of large-diameter trees for all species, with many stands having old-growth characteristics [31]. Ponderosa pine, Douglas-fir, white fir, subalpine fir, blue spruce, limber pine (Pinus flexilis), and quaking aspen are the common tree species. Common sprouting shrubs include Gambel oak (Quercus gambelii), snowberry (Symphoricarpos oreophilus), chokecherry (Prunus virginiana), and Utah serviceberry (Amalanchier alnifolia). In 2008, portions of the study area were burned in a broadcast-prescribed fire, using aerial ignitions. Ten years later, in 2018, the study area was burned by an unplanned, artificial ignition that burned 54,130 acres (416 Fire). The 416 Fire burned during an extreme drought year, resulting in mixed soil burn severities from overall moderate fire behavior driven primarily by available fuels and topography. The daily rate of spread and flaming front intensities varied based on daily fire weather and only sometimes exhibited a rapid rate of spread indicative of extreme intensity. Given the large area burned over ~four months, little can be quantified for fire intensities experienced at an individual tree or stand level, making char height an okay proxy for flame length at an individual tree. Suppression efforts focused on the wildland–urban interface, and no slurry drops or direct attack measures were taken in the study area (Communications with the Incident Section Chief). Before the 2018 wildfire, the study area had minimal previous disturbance, and the overall tree density was lower than that of other similar forest types in the area that had previously been logged [31].
We conducted sampling from May to September 2023. We located study plots in Douglas-fir beetle-infected areas in the 416 Fire. To identify stands within the 416 Fire that were potentially affected by Douglas-fir beetle, we used United States Forest Service 2022 Aerial Detection Survey data (ADS) [32]. We then inspected trees in the ADS-identified areas for signs of Douglas-fir beetle attacks (boreholes, reddish-orange bore dust on the bole, reddish-brown needles) until we found a grouping of Douglas-fir beetle-infected trees. This resulted in no plots in high-severity burn areas, <1% of plots in moderate burn severity, and 85.7% of plots in low-severity burn areas, with the remainder of plots being in unburned areas. We randomly selected the plot center, and we sampled the first tree nearest the plot center and then a tree in each cardinal direction of the initial tree until five total trees were sampled. Within a 22.36 m diameter circle (0.04 ha) surrounding the plot center, we recorded the following stand characteristics: stand-level burn severity, stand density by species, basal area by species, % alive/declining/dead, and % Douglas-fir infected. Plots were considered uninfected if the number of Douglas-fir trees in the stand infected by Douglas-fir beetles was <20%. We established 14 plots in beetle-infested Douglas-fir stands, which were paired with 14 nearby uninfected Douglas-fir stands, for a total of 28 plots (infected N = 14) and 140 sampled Douglas-fir trees (infected N = 70). Paired plots were 30–100 m from each other.
We designated all sampled trees with the following condition classes: green (needles 100% green), partially brown (<50% of needles are brown), brown (100% of needles are brown), and defoliated (<50% needles are remaining). We then recorded their DBH and height. We assessed the sampled tree’s degree of fire injury by using bark char and bark char height. We assessed bark char visually and assigned a bark char severity index (BCSI) classification of none = 0, superficial = 1, moderate = 2, or severe = 3 (Table 1). We viewed trees from four sides (north, south, east, and west) and gave each quadrant a bark char classification. We then summed the BCSI for each quadrant for a tree’s overall BCSI (maximum BCSI = 12). We based the BCSI on the guidelines presented by Ryan (1982). We also recorded the height of the tallest bark char extent on the tree’s bole.

Table 1.
Bark char severity index (BCSI) was adapted from Ryan (1982). We recorded BCSI at the bole base for all four cardinal directions and summed the four quadrants (max value = 12 if all quadrants were rated as severe).
We assessed the level of beetle attack by visually inspecting the tree’s circumference and then estimating the percentage with evidence of beetle attack. Trees with ≥75% of their circumference affected by beetles were classified as mass attacked, and <75% were classified as strip attacked. We confirmed Douglas-fir beetle presence by removing a 15 cm × 30 cm section of bark and then inspected the cambium for Douglas-fir beetle egg gallery patterns for a random sampling of attacked trees. We photographed the egg galleries for later confirmation.
We paired plots and ran Mann–Whitney U tests to determine which stand, tree, and fire injury variables had statistically significant differences between uninfected and infected stands. We used Pearson correlation tests to determine the correlation between DBH and tree height, BCSI, and bark char height and used a Bonferroni post hoc correction. We used a machine learning-based recursive partitioning hierarchal decision tree to model the influence of stand and tree characteristics and burn injury on the probability of infection. We included all possible stand- and tree-level variable combinations as separate models and used the Akaike information criterion (AIC) to select the best-performing model [33]. Multiple models exist to achieve this goal; however, because logistic model trees predict the probability of a given outcome, we chose this approach over classification-based approaches that deal with categorical or continuous outcomes [34]. We included tree height, DBH, BCSI, and bark char height as our model’s input parameters, and only mass-attacked trees were included in our infected dataset. We randomly selected and retained 20% of our tree dataset for model cross-validation. We conducted Mann–Whitney U tests in SPSS version 27 software and used the tidymodels and glmtree packages in Program R, RStudio Build 524 for decision tree modeling [35,36,37].
3. Results
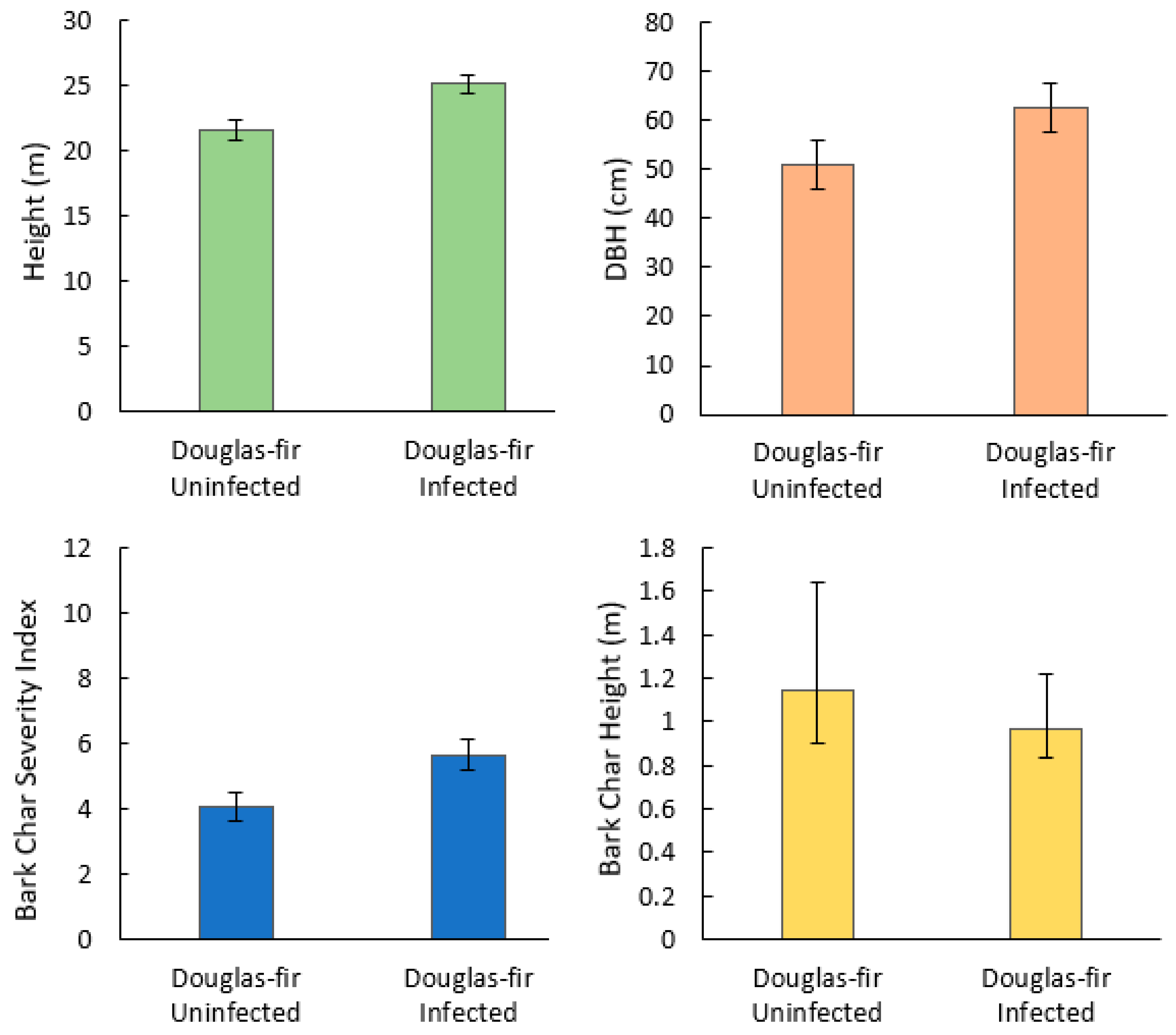
Bark char severity index (p = 0.02), tree height (p < 0.001), and DBH (p < 0.001) were significantly higher in infected trees than uninfected trees, while bark char height (p = 0.5) was not significantly different (Figure 1).
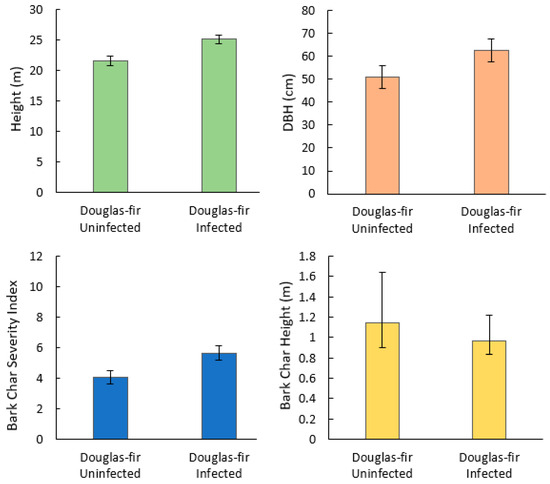
Figure 1.
Mean and 95% CI for tree-level variables of Douglas-fir beetle-infected and -uninfected Douglas-fir trees (N = 140) in the 416 Fire burn area, five years post-fire. Mann–Whitney U showed a statistically significant difference between infected and uninfected height, DBH, and bark char severity index (p < 0.001, p = 0.016, p < 0.001, respectively). Bark char height was not statistically significant.
For uninfected trees, 31% had no visible bark char, 22% had a BCSI of superficial, 22% rated moderate, and 14% rated severe. Of the infected trees, 21% had no visible bark char, 17% rated superficial, 34% rated moderate, and 27% rated severe. Ninety-two percent of the uninfected trees were in the green condition class, and 7% were defoliated. Of the infected trees, 18% were green, 17% were partially brown, 44% were brown, and 20% were defoliated.
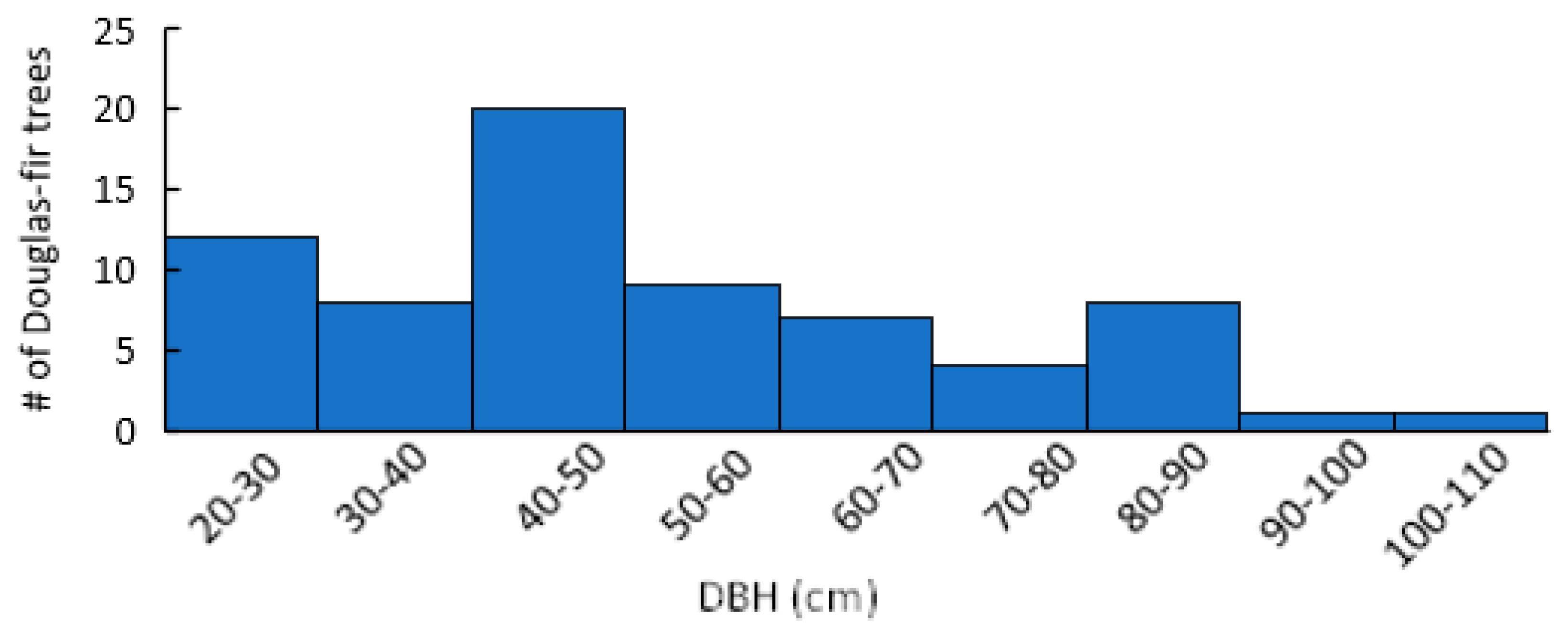
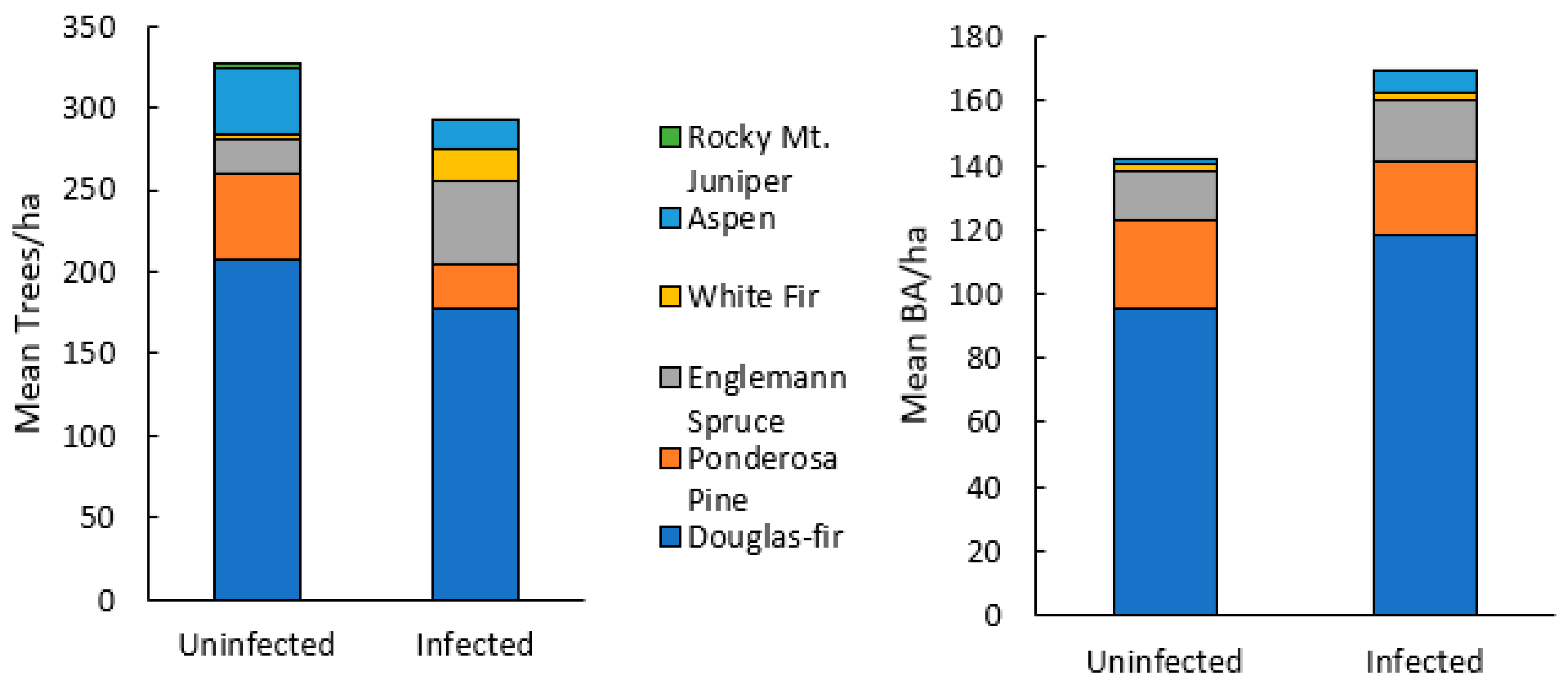
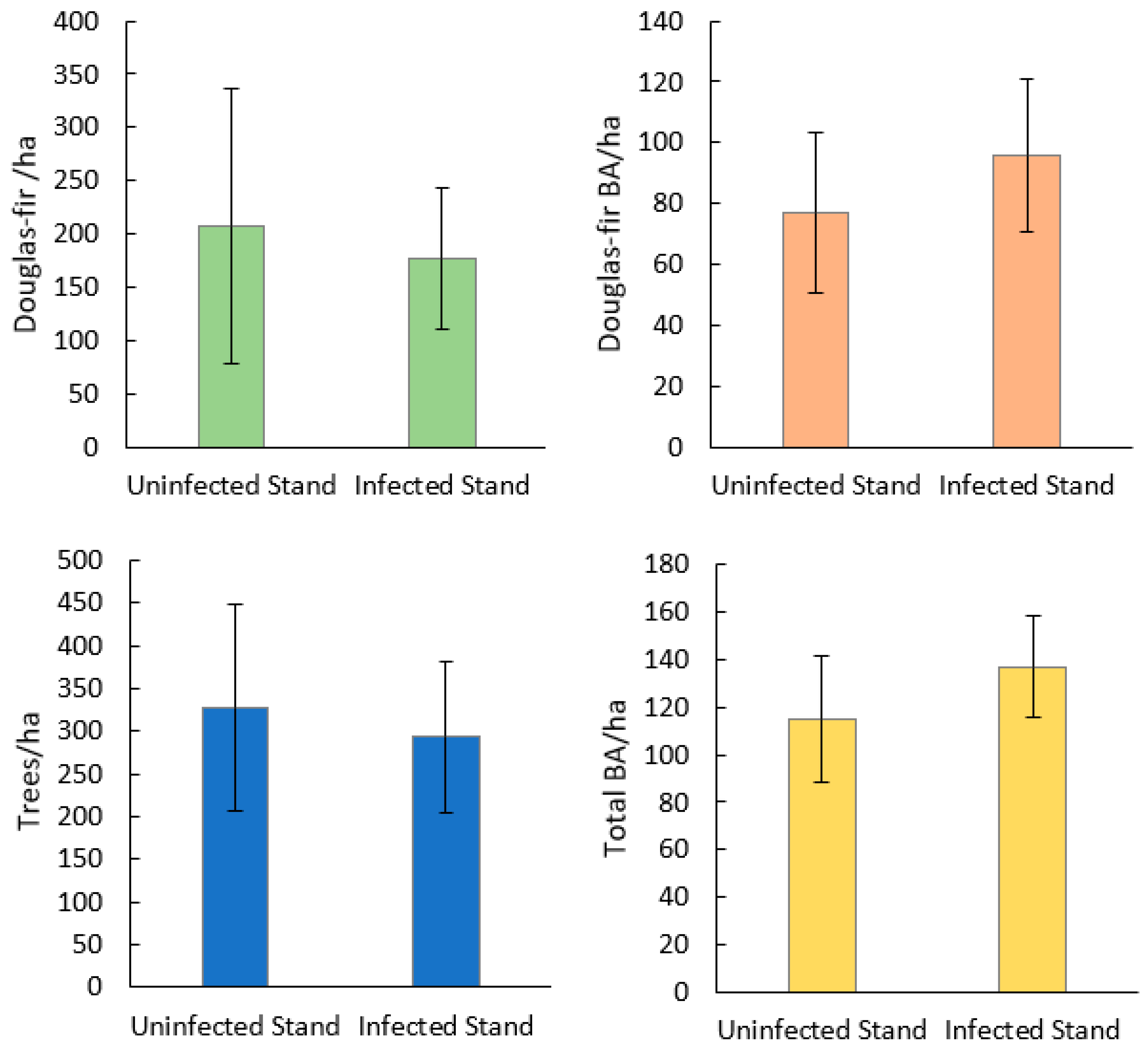
Seventy-seven percent of Douglas-fir within plots were found to show mature or old growth characteristics (DBH > 40 cm) (Figure 2) [38]. Stand species composition was similar between infected and uninfected stands, with no significant differences in the mean density or BA of any single species (Figure 3). Mean stand density (trees/ha), Douglas-fir density (DF/ha), and Douglas-fir basal area (DF BA/ha) were not significantly different between infected and uninfected stands. Mean total basal area (BA/ha) was higher in infected stands than in uninfected but was not significant (p = 0.07). The range of data for BA/ha for infected stands included much higher values (upper CI = 119, lower CI = 64) compared to uninfected stands (upper 95% CI = 108, lower 95% CI =48). Additionally, infected stands had a higher percentage of Douglas-fir (mean = 66%, upper 95% CI =84%, lower 95% CI = 50%) compared to uninfected stands (mean =14%, upper 95% CI = 26%, lower 95% CI = <1%) (Figure 4). Mean live tree density was significantly higher in uninfected stands than in infected stands (p < 0.01), while mean declining and dead trees were significantly higher in infected stands (p = 0.03 and p < 0.01, respectively). Of the uninfected stands, 71% were in low-severity burn areas, 14% were in moderate-severity burn areas, and 14% were in unburned areas. Eighty-five percent of the infected stands were in a low-severity area, 7% were in a moderate-severity area, and 7% were in an unburned area. None of the infected or uninfected stands was in a high-burn-severity area.
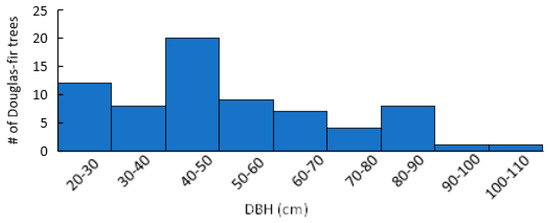
Figure 2.
DBH (cm) distribution classes for sampled Douglas-fir trees in the 416 Fire burn area five years post-fire. N = 140.
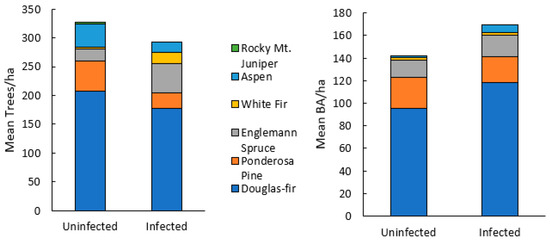
Figure 3.
Stand structure of live and dead trees by species for Douglas-fir beetle uninfected and infected stands in the 416 Fire burn area, five years post-fire (N = 28). There were no significant differences in mean trees/ha or mean BA/ha of any species using Mann Whitney U tests.
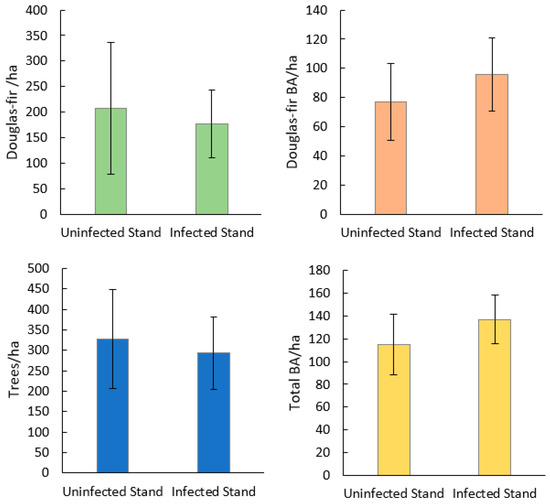
Figure 4.
Mean and 95% CI for stand-level variables of Douglas-fir beetle-infected and -uninfected Douglas-fir stands in the 416 Fire burn area, five years post-fire (N = 28). Mann–Whitney U did not find statistically significant differences between Douglas-fir trees/ha, Douglas-fir BA/ha, or trees/ha. Total BA/ha was found to be marginally significantly higher in infected stands than uninfected stands (p = 0.07).
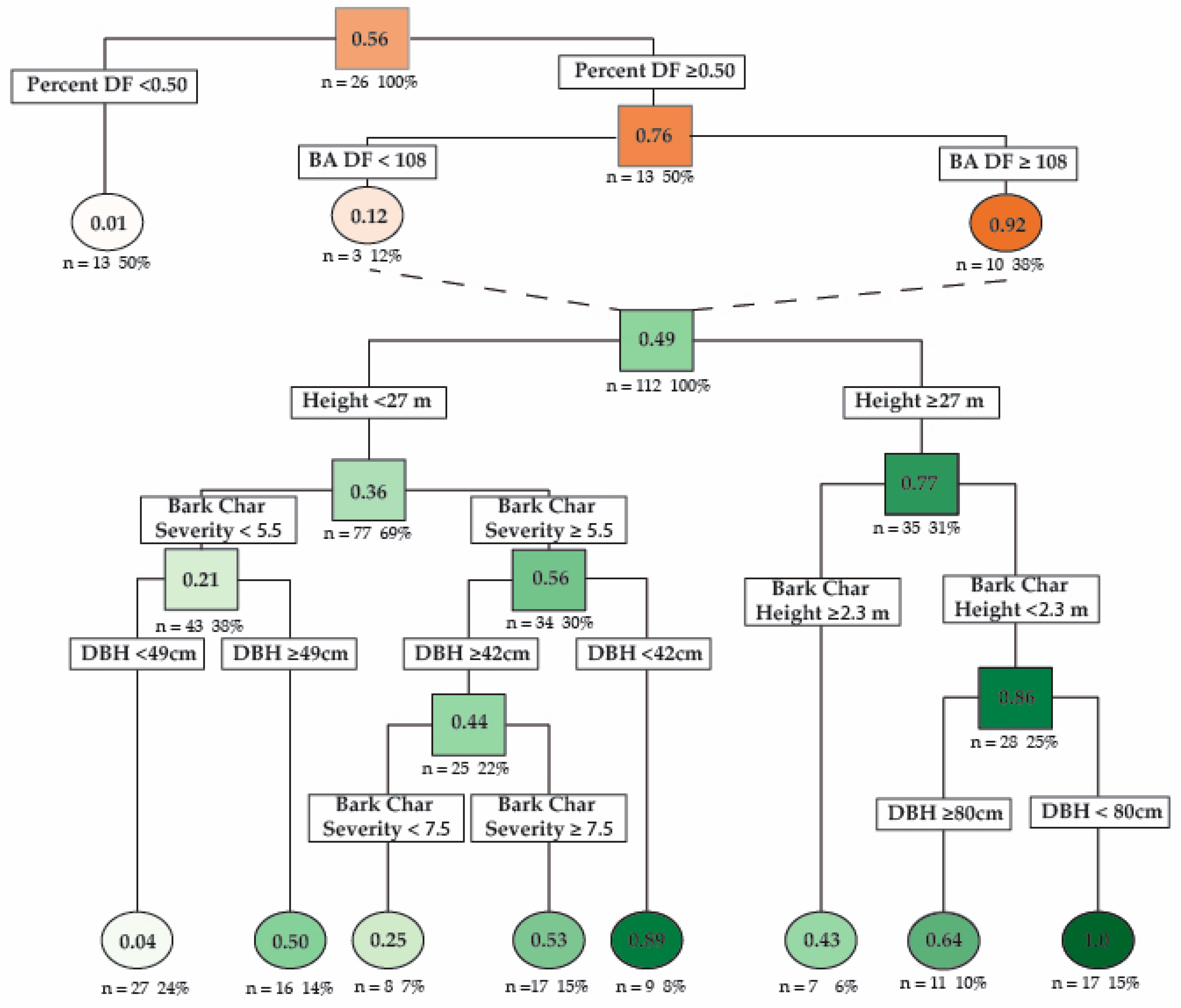
Using recursive partitioning, a hierarchal logistic regression decision tree was used to model the probability of Douglas-fir beetle infection. The best performing model included the basal area of Douglas-fir and the percent of basal area of Douglas-fir as stand-level predictors and DBH, tree height, BCSI, and bark char height as tree-level predictors (Table 2). This model had two nodes at the stand level and seven nodes at the tree level (Figure 5). Cross-validation showed that the model could predict whether a tree would be infected by Douglas-fir beetle with a 78% predictive accuracy (RSME = 0.473, R2 = 0.54). The model predicts that stands with greater than 50% of the basal area composed of Douglas-fir are more likely to have trees infected with Douglas-fir beetle (infection probability = 76%), and within these stands, if Douglas-fir basal area exceeds 108 m2/ha, then the stand is highly likely to have trees infected by Douglas-fir beetle (infection probability 92%). The model found that the strongest predictor of Douglas-fir beetle infection was a combination of a tree height ≥ 27 m, bark char height < 2.3 m, and DBH < 80 cm (infection probability = 100%). For trees <27 m, the strongest predictors were a combination of BCSI ≥ 5.5 and DBH ≥ 42 cm (infection probability = 89%). Trees with the lowest probability of infection were trees < 27 m in height, BCSI < 5.5, and DBH < 49 cm (infection probability = 3.4%). Tree DBH and height were highly correlated (R2 = 0.71, p < 0.001), while DBH and BCSI were poorly correlated (R2 = 0.06, p = 0.15), as were DBH and bark char height (R2 = 0.13, p = 0.05). BCSI and bark char height were moderately correlated (R2 = 0.472, p < 0.001).
Table 2.
Model variables and resulting score indices and RSME and R2 values for an iterative leave one out of the hierarchical logistic tree. The best-performing model is bolded. AIC, Akaike information criterion.
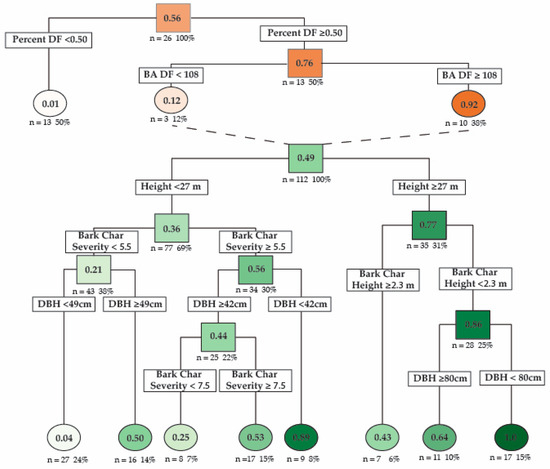
Figure 5.
A hierarchical logistic regression decision tree model created using recursive partitioning that depicts the probability that a Douglas-fir tree will be infected by Douglas-fir beetle depending on diameter at breast height (DBH cm), tree height (Height, m), bark char severity index (Bark Char Severity), and bark char height (m). The orange nodes represent stand-level data and the probability of a beetle infection occurring within a stand. The green nodes represent tree-level characteristics within an infected stand. The top orange node represents all plots sampled (13 infected and 13 uninfected). Moving down the tree, lower nodes depict the probability of a stand being infected given the above characteristics. The dashed lines connect stands with infections to tree-level probability of infections in the hierarchical model. The top green node represents all trees sampled, excluding 20% of samples retained for model cross-validation (66 infected/66 uninfected). Moving down the tree, lower nodes depict the probability that a tree with the proceeding characteristics will be infected by Douglas-fir beetle. Under each node is the number and percentage of sampled trees with those characteristics. Predictive accuracy = 78%; RSME = 0.473; R2 = 0.54.
4. Discussion
Five years post-fire, Douglas-fir trees within the 416 Fire burn area experienced a 37% average beetle infection rate, with infected stands experiencing a 67% average infection rate, while some stands had up to a 100% infection rate. Other studies have found similarly high rates of Douglas-fir beetle infection post-fire [39,40]. None of the infected stands we encountered was in a high-severity burn area, <1% were in moderate-severity burn areas, and 85.7% were in low-severity burn areas. Trees in high-severity burn areas were killed during the fire, or their phloem may have been significantly damaged, rendering them unsuitable hosts for Douglas-fir beetle brood production.
Because sampling occurred during a single season and our model included all trees showing signs of mass infection in our dataset, regardless of condition class, our study does not measure the progressive patterns of Douglas-fir beetle infection over 5 years since the time of fire but rather host tree selection patterns five years post-fire. Despite this, we can make some inferences about infection patterns over the five-year time period based on tree foliage conditions. We found that 36% of infected trees had all or a significant percentage of green needles, suggesting that these trees had been infected during the 4th and 5th post-fire years [41], though there may be considerable variability in the timing of foliar change after Douglas-fir beetle infection [42]. Hood and Bentz [19] tracked beetle infections for four successive years immediately post-fire and found that most beetle infections occurred in the first one-to-two years, and Cunningham et al. [17] found that the importance of burn injury decreased two years post-fire. Nevertheless, based on foliar condition alone, we found many infections occurring 4-to-5 years post-fire within the 416 Fire burn area.
At the stand level, we found stands with higher proportions of basal area composed of Douglas-fir and basal area of Douglas-fir higher than 108 m2/ha to be the highest probability of having Douglas-fir beetle infestations. This makes sense because these stands are stands that have a strong Douglas-fir component in their composition, and thus, the probability of having trees that are preferred by Douglas-fir beetle increases. This stand-level information provides some guidance for silvicultural opportunities to control stand composition to reduce the likelihood of stands being susceptible to Douglas-fir beetle. Given the high proportion of plots with old-growth characteristics, it is important to note that passive management is still an appropriate tool with the acceptance that mortality from fire or bark beetles following fire can help develop old-growth characteristics [43]. The decision to manage a stand to reduce the likelihood of infection by Douglas-fir beetle should depend on the goals and broader social, ecological, and economic context of the stand being considered, but here we provide managers with a useful decision tree to help inform their decisions about the risk for infection by Douglas-fir beetle.
Our model found tree height to be the most important variable in driving the probability of beetle infection, but tree DBH can be substituted, as tree height and DBH were highly correlated. Beetles may prefer larger trees because they have a thicker phloem, which is needed to sustain developing larvae [21]. This finding is consistent with other studies on Douglas-fir beetle’s host-tree selection, both with the influence of fire [17,18,19,44] and without [12,22,39]. For trees ≥ 27 m, a lower bark char height was the most driving infection variable. This pattern may be due to Douglas-fir beetles preferring trees with moderate burn injury, as observed in other studies [17,18,19]. Moderately fire-injured trees may provide a balance of compromised tree defenses with undamaged phloem, making them the most attractive hosts to beetles. Another possibility is that these larger trees are more stressed than might be expected from their low bark char height. We observed a ring of high-severity bark char around the bottom portion of many larger and infected trees, often only a few cm high, likely caused by accumulated duff burning for longer residence times. A similar pattern of fire injury was seen by Gibson [45], who found that trees with little apparent damage to their boles may be completely girdled at the root collar. In trees less than 27 m in height with low-severity BCSI, those with larger diameters were more likely to be infected by bark beetles than those with smaller diameters. Shorter trees that are also smaller in diameter (less than 42 cm) were only likely to be infected when they had moderate–severe BCSI, thus suggesting that, without burn injury or with only low-severity burn injury, larger trees are preferred, whereas when moderate or greater burn injury occurs, beetles sometimes begin to select for more mid–small-diameter trees. Again, this may be a function of the beetle’s selection for a balance between compromised tree defenses and undamaged phloem. We found an increased probability of infection for smaller trees (DBH < 42 cm) at a lower BCSI than larger trees. This is likely due to smaller trees having thinner bark and requiring less intense scorching before reaching the fire-injury threshold favored by Douglas-fir beetle; however, it would be interesting to measure bark thickness [39,40,46] directly.
Due to our study’s timing and other constraints, our model does not include several factors that may drive the probability of Douglas-fir beetle infection. Other studies found percent crown volume scorch to be an important fire injury variable in predicting Douglas-fir beetle infection [17,18,19]. Five years post-fire, we found that 64% of infected trees were partially or completely defoliated, and distinguishing between loss of needles due to beetle effects and crown scorch was impossible. The distance from other Douglas-fir beetle outbreaks may also significantly affect the probability of post-fire infection, as beetles have a maximum range from established populations that they are capable of dispersing to colonize new areas [47]. If established populations are too far from potential hosts, the probability of infection may be reduced to zero. In their post-fire tree mortality model, Scott and others (2002) included distance from known beetle outbreaks; however, pre-fire Douglas-fir beetle outbreak information surrounding our study area was unavailable [48]. Other studies have also found Douglas-fir BA and stand density to predict Douglas-fir beetle infection [12,19,22]. Our study found no stand-level variables to be statistically significant, though total stand BA was marginally higher in infected than uninfected stands. A small sample size may have impacted our results, but we did not include stand-level variables in our model due to a lack of statistically significant differences.
Our finding that large-diameter trees were more likely to be infected if they had a lower bark char height lends credence to duff removal around tree boles before prescribed fire treatment as a method of preservation, particularly old-growth trees in forests where fire has been excluded for extend periods [49,50]. Fine fuel reduction through targeted grazing could also reduce scorching around tree root collars and help reduce the probability of beetle infection in high-value trees [51].
Post-fire management priorities vary depending on ecological and economic objectives, safety, and land ownership. Our decision tree model provides added precision to decisions made by land managers and private landowners when managing desired post-fire conditions or targeting trees for removal and preservation measures. Our study also exemplifies the importance of the stand level, tree level, and specifics regarding fire behavior in order to accurately predict infestation of Douglas-fir beetles following a fire. During post-fire salvage logging operations, efforts should be made to eliminate the adverse ecological impacts of heavy equipment on recovering soil and regenerating vegetation. Our model facilitates precision targeting of high-risk stands and trees within these stands for removal while leaving low-risk trees as seed regeneration sources, potentially reducing the microsite impacts of salvage logging [52]. Our model can also inform the deployment of anti-aggregation pheromones (MHC) to defend high-risk trees near recreational sites, near residential areas, on private property, or as post-fire regeneration seed sources [53]. Though additional variables could be included in Douglas-fir beetle infection models, such as distance from known infections or stand structure characteristics, it may reduce the usability of the model by non-experts. Our simplified model includes parameters that a minimally trained individual could quickly collect in situ to evaluate a tree’s infection probability. Our model also includes both stand-level and individual tree-level characteristics that can help users identify at-risk stands and the at-risk trees within those stands. Our study focused on stand-level scales and thus does not include landscape parameters and should not be used in isolation of landscape-level considerations. Our study occurred in an area with evidence of ongoing Douglas-fir beetle outbreaks adjacent to the study site and, therefore, is dependent on the presence of beetles within flight distance of the stands in question. Other sites may be further away from known beetle activity and thus have a lower innate risk based on the landscape patterns of Douglas-fir beetle distribution. Future studies should consider developing additional models with simplified parameters, such as ours, with the intention of ease of use for the end user and with a more robust sample size than we were able to employ to ensure model accuracy and generalized applicability across broad landscapes.
Author Contributions
Conceptualization, M.Y. and J.K.; methodology, M.Y.; data collection M.Y.; data curation, M.Y.; software, M.Y. and J.K.; validation, M.Y., J.K., and M.R.; formal analysis, M.Y. and M.R.; writing—original draft preparation, M.Y.; writing—review and editing, J.K. and M.R.; visualization, M.Y. and M.R.; supervision, J.K.; project administration, J.K.; funding acquisition, M.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fort Lewis College Undergraduate Research.
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not Applicable.
Data Availability Statement
Contact the corresponding author for access to complete dataset.
Acknowledgments
Thank you to the Fort Lewis College Biology Department, and Amy Lockner (USFS) for your support. We also thank two anonymous reviewers for their comments, which substantively improved this manuscript.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Romme, W.H.; Floyd, L.M.; Crist, M.R. Historical Range of Variability and Current Landscape Condition Analysis: South Central Highlands Section, Southwestern Colorado & Northwestern New Mexico; Colorado Forest Restoration Institute: Fort Collins, CO, USA, 2009. [Google Scholar]
- Schoennagel, T.; Veblen, T.T.; Romme, W.H.; Sibold, J.S. ENSO and PDO Variability Affect Drought-Induced Fire Occurrence in Rocky Mountain Subalpine Forests. Ecol. Appl. 2005, 15, 2000–2014. [Google Scholar] [CrossRef]
- Rodman, K.C.; Andrus, R.A.; Butkiewicz, C.L.; Chapman, T.B.; Gill, N.S.; Harvey, B.J.; Kulakowski, D.; Tutland, N.J.; Veblen, T.T.; Hart, S.J. Effects of Bark Beetle Outbreaks on Forest Landscape Pattern in the Southern Rocky Mountains, U. S.A. Remote Sens. 2021, 13, 1089. [Google Scholar] [CrossRef]
- Bentz, B.J.; Rgnire, J.; Fettig, C.J.; Hansen, E.M.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.G.; Negron, J.F.; Seybold, S.J. Climate Change and Bark Beetles of the Western United States and Canada: Direct and Indirect Effects. Bioscience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Grissino-Mayer, H.D.; Romme, W.H.; Floyd, M.L.; Hanna, D.D. Climatic and human influences on fire regimes of the southern San Juan mountains, Colorado, USA. Ecology 2004, 85, 1708–1724. [Google Scholar] [CrossRef]
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and Earlier Spring Increase Western U. S. Forest Wildfire Activity. Science 2006, 313, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Kane, J.M.; Varner, J.M.; Metz, M.R.; van Mantgem, P.J. Characterizing Interactions between Fire and Other Disturbances and Their Impacts on Tree Mortality in Western U. S. Forests. Ecol Manag. 2017, 405, 188–199. [Google Scholar] [CrossRef]
- Remke, M.J.; Chambers, M.E.; Tuten, M.C.; Pelz, K.A. Mixed Conifer Forests in the San Juan Mountains Region of Colorado, USA: The Status of Our Knowledge and Management Implications; Colorado Forest Restoration Institute Report—2110; Colorado Forest Restoration Institute: Fort Collins, CO, USA, 2021. [Google Scholar]
- Lousier, J.D.; Joseph, D.; Kessler, W.B. Ecology and Management of Interior Douglas-Fir (Pseudotsuga Menziesii Var Glauca) at the Northern Extreme of Its Range: Proceedings of a Workshop Held 7–9 October, 1996; Faculty of Natural Resources & Environmental Studies, University of Northern British Columbia: Fort St. James, BC, USA, 1999; ISBN 0968633609. [Google Scholar]
- Schmitz, R.; Gibson, K. Douglas-Fir Beetle. In USDA Forest Service, Forest Insect and Disease Leaflet; Report R1-96-87; Utah State University: Logan, UT, USA, 1996; pp. 5–7. [Google Scholar]
- McMullen, L.H.; Atkins, M.D. On the Flight and Host Selection of the Douglas-Fir Beetle, Dendroctonus Pseudotsugae Hopk. (Cdeoptera: Scolytidae). Can. Entomol. 1962, 94, 1309–1325. [Google Scholar] [CrossRef]
- Negron, J.F. Probability of Infestation and Extent of Mortality Associated with the Douglas-Fir Beetle in the Colorado Front Range. Ecol Manag. 1998, 107, 71–85. [Google Scholar] [CrossRef]
- Powers, J.S.; Sollins, P.; Harmon, M.E.; Jones, J.A. Plant-Pest Interactions in Time and Space: A Douglas-Fir Bark Beetle Outbreak as a Case Study. Landsc. Ecol. 1999, 14, 105–120. [Google Scholar] [CrossRef]
- Dodds, K.J.; Garman, S.L.; Ross, D.W. Landscape Analyses of Douglas-Fir Beetle Populations in Northern Idaho. Ecol. Manag. 2006, 231, 119–130. [Google Scholar] [CrossRef]
- Mcmillin, J.D.; Allen, K.K. Effects of Douglas-fir beetle (Coleoptera: Scolytidae) infestations on forest overstory and understory conditions in western Wyoming. West. N. Am. Nat. 2003, 63, 498–506. [Google Scholar]
- Allen, G.S.; Owens, J.N. The Life History of Douglas Fir. Environment Canada Forestry Service: Ottawa, ON, Canada, 1972. [Google Scholar]
- Cunningham, C.A.; Jenkins, M.J.; Roberts, D.W. Attack and brood production by the Douglas-fir beetle (Coleoptera: Scolytidae) in Douglas-fir, Pseudotsuga menziesii var. Glauca (Pinaceae), following a wildfire. West. N. Am. Nat. 2005, 65, 70–79. [Google Scholar]
- Furniss, M.M. Susceptibility of Fire-Injured Douglas-Fir to Bark Beetle Attack in Southern Idaho. J. For. 1965, 63, 8–11. [Google Scholar]
- Hood, S.; Bentz, B. Predicting Postfire Douglas-Fir Beetle Attacks and Tree Mortality in the Northern Rocky Mountains. Can. J. For. Res. 2007, 37, 1058–1069. [Google Scholar] [CrossRef]
- Kitchens, K.A.; Peng, L.; Daniels, L.D.; Carroll, A.L. Patterns of Infestation by Subcortical Insects (Coleoptera: Buprestidae, Cerambycidae) after Widespread Wildfires in Mature Douglas-Fir (Pseudotsuga menziesii) Forests. Ecol Manag. 2022, 513, 120203. [Google Scholar] [CrossRef]
- Shore, T.L.; Safranyik, L.; Riel, W.G.; Ferguson, M.; Castonguay, J. Evaluation of factors affecting tree and stand susceptibility to the Douglas-fir beetle (Coleoptera: Scolytidae). Can. Entomol. 1999, 131, 831–839. [Google Scholar] [CrossRef]
- Sturdevant, N.; Haavik, L.; Negrón, J.F. Douglas-Fir Tree Mortality Caused by the Douglas-Fir Beetle in Thinned and Unthinned Stands in Montana, USA. For. Sci. 2022, 68, 145–151. [Google Scholar] [CrossRef]
- Negrón, J.F.; Lynch, A.M.; Schaupp, W.C.; Mercado, J.E. Douglas-Fir Tussock Moth- and Douglas-Fir Beetle-Caused Mortality in a Ponderosa Pine/Douglas-Fir Forest in the Colorado Front Range, USA. Forests 2014, 5, 3131–3146. [Google Scholar] [CrossRef]
- Ganio, L.M.; Progar, R.A. Mortality Predictions of Fire-Injured Large Douglas-Fir and Ponderosa Pine in Oregon and Washington, USA. Ecol Manag. 2017, 390, 47–67. [Google Scholar] [CrossRef]
- Hood, S.M.; McHugh, C.W.; Ryan, K.C.; Reinhardt, E.; Smith, S.L. Evaluation of a Post-Fire Tree Mortality Model for Western USA Conifers. Int. J. Wildland Fire 2007, 16, 679–689. [Google Scholar] [CrossRef]
- Ross, D.W.; Daterman, G.E. Integrating Pheromone and Silvicultural Methods for Managing the Douglas-Fir Beetle. In Proceedings of the Integrating cultural tactics into the management of bark beetle and reforestation pests. In USFS General Technical Report NE-236; USDA Forest Service: Washington, DC, USA, 2018; pp. 135–145. [Google Scholar]
- Ross, D.W. Douglas-Fir Beetle Response to Artificial Creation of Down Wood in the Oregon Coast Range. West. J. Appl. For. 2006, 21, 117–122. [Google Scholar] [CrossRef]
- Ross, D.W.; Daterman, G.E. Using Pheromone-Baited Traps To Control the Amount and Distribution of Tree Mortality During Outbreaks of the Douglas-Fir Beetle. For. Sci. 1997, 43, 65–70. [Google Scholar]
- Bernal, A.A.; Kane, J.M.; Knapp, E.E.; Zald, H.S.J. Tree Resistance to Drought and Bark Beetle-Associated Mortality Following Thinning and Prescribed Fire Treatments. Ecol Manag. 2023, 530, 120758. [Google Scholar] [CrossRef]
- BAER 416 Fire Executive Summary. In USDA Forest Service Report; San Juan National Forest: Durango, CO, USA, 2018.
- Korb, J.E.; Fulé, P.Z.; Wu, R. Variability of warm/dry mixed conifer forests in southwestern Colorado, USA: Implications for ecological restoration. For. Ecol. Manag. 2013, 304, 182–191. [Google Scholar] [CrossRef]
- Coleman, T.; Graves, A.; Heath, Z.; Flowers, R. Accuracy of aerial detection surveys for mapping insect and disease disturbances in the United States. Ecol Manag. 2018, 430, 321–336. [Google Scholar] [CrossRef]
- Portet, S. A primer on model selection using the Akaike Information Criterion. Infect. Dis. Model. 2020, 5, 111–128. [Google Scholar] [CrossRef]
- Chen, W.; Xie, X.; Wang, J.; Pradhan, B.; Hong, H.; Bui, D.T.; Duan, Z.; Ma, J. A comparative study of logistic model tree, random forest, and classification and regression tree models for spatial prediction of landslide susceptibility. CATENA 2017, 151, 147–160. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 20 September 2023).
- Hothorn, T.; Hornik, K.; Zeileis, A. Unbiased Recursive Partitioning: A Conditional Inference Framework. J. Comput. Graph. Stat. 2006, 15, 651–674. [Google Scholar] [CrossRef]
- Kuhn, M.; Wickham, H. Tidymodels: A Collection of Packages for Modeling and Machine Learning Using Tidyverse Principles. 2020. Available online: https://www.tidymodels.org (accessed on 21 September 2023).
- Mehl, M. Old-Growth Descriptions for the Major Forest Cover Types in the Rocky Mountain Regions. In Proceedings of the Old-Growth Forests in the Southwest and Rocky Mountain Regions, Portal, AZ, USA, 9 March 1992; USFS: Portal, AZ, USA; pp. 106–120. [Google Scholar]
- Furniss, M. Infestation Patterns of Douglas-Fir Beetle in Standing and Windthrown Trees in Southern Idaho. J. Econ. Entomol. 1992, 55, 486–491. [Google Scholar] [CrossRef]
- Ryan, K.C.; Peterson, D.L.; Reinhardt, E.D. Modeling Long-Term Fire-Caused Mortality of Douglas-Fir. For. Sci. 1988, 34, 190–199. [Google Scholar]
- Rasmussen, L.; Amman, G.D.; Vandygriff, J.C.; Oakes, R.D.; Munson, S.S.; Gibson, K. Bark Beetle and Wood Borer Infestation in the Greater Yellowstone Area During Four Postfire Years; USDA Forest Service, Intermountain Forest and Range Experiment Station: Washington, DC, USA, 1996. [Google Scholar]
- Bedard, W.D. The Douglas-Fir Beetle; Circular No. 817; USDA: Washington, DC, USA, 1950. [Google Scholar]
- Harrington, T.; Peter, D.; Marshall, D.; DeBell, D. Ten-year Douglas-fir regeneration and stand productivity differ among contrasting silvicultural regimes in western Washington, USA. For. Ecol. Manag. 2022, 510, 120102. [Google Scholar] [CrossRef]
- Weatherby, J.; Progar, R.A.; Mocettini, P. Evaluation of Tree Survival on the Payette National Forest 1995–1999; Intermountain Region Report; USDA Forest Service: Washington, DC, USA, 2001. [Google Scholar]
- Gibson, K.; Negron, J. Fire and Bark Beetle Interactions. In The Western Bark Beetle Research Group: A Unique Collaboration with Forest Health Protection-Proceedings of a Symposium at the 2007 Society of American Foresters Conference, Portland, OR, USA, 23–28 October 2009; USDA Forest Service: Washington, DC, USA, 2009. [Google Scholar] [CrossRef]
- Maguire, D.A.; Hann, D.W. Bark Thickness and Bark Volume in Southwestern Oregon Douglas-Fir. West. J. Appl. For. 1990, 5, 5–8. [Google Scholar] [CrossRef]
- Jones, K.L.; Shegelski, V.A.; Marculis, N.G.; Wijerathna, A.N.; Evenden, M.L. Factors Influencing Dispersal by Flight in Bark Beetles (Coleoptera: Curculionidae: Scolytinae): From Genes to Landscapes. Can. J. For. Res. 2019, 49, 1024–1041. [Google Scholar] [CrossRef]
- Scott, D.W.; Schmitt, C.L.; Spiegel, L.H. Factors Affecting Survival of Fire Injured Trees: A Rating System For Determining Relative Probability of Survival of Conifers in the Blue and Wallowa Mountains; USDA Forest Service, Blue Mountains Pest Management Service Center: Washington, DC, USA, 2002. [Google Scholar]
- Fowler, J.F.; Sieg, C.H.; Wadleigh, L.L. Effectiveness of Litter Removal to Prevent Cambial Kill-Caused Mortality in Northern Arizona Ponderosa Pine. For. Sci. 2010, 56, 166–172. [Google Scholar]
- Hood, S.M. Mitigating Old Tree Mortality in Long-Unburned, Fire-Dependent Forests: A Synthesis; Gen. Technical Report; USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2010. [Google Scholar]
- Bruegger, R.A.; Varelas, L.A.; Howery, L.D.; Torell, L.A.; Stephenson, M.B.; Bailey, D.W. Targeted Grazing in Southern Arizona: Using Cattle to Reduce Fine Fuel Loads. Rangel. Ecol. Manag 2016, 69, 43–51. [Google Scholar] [CrossRef]
- Marcolin, E.; Marzano, R.; Vitali, A.; Garbarino, M.; Lingua, E. Post-Fire Management Impact on Natural Forest Regeneration through Altered Microsite Conditions. Forests 2019, 10, 1014. [Google Scholar] [CrossRef]
- Ross, D.W.; Gibson, K.; Daterman, G.E. Using MCH to Protect Trees and Stands from Douglas-Fir Beetle Infestation; USDA Forest Service, Forest Health Technology Enterprise Team: Washington, DC, USA, 2015. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).