
Pattern Formation Mechanisms of Spatiotemporally Discrete Activator–Inhibitor Model with Self- and Cross-Diffusions


Abstract
1. Introduction
2. The CML Model and Stability Analysis
2.1. Development of CML Model
2.2. Stability Analysis of the Homogeneous Stationary State
- It is a saddle if one of and holds:
- It is an unstable node if one of , , and holds:
- It is a stable node if one of and holds:
3. Analysis of the Bifurcation Behaviors of the Homogeneous Stationary State
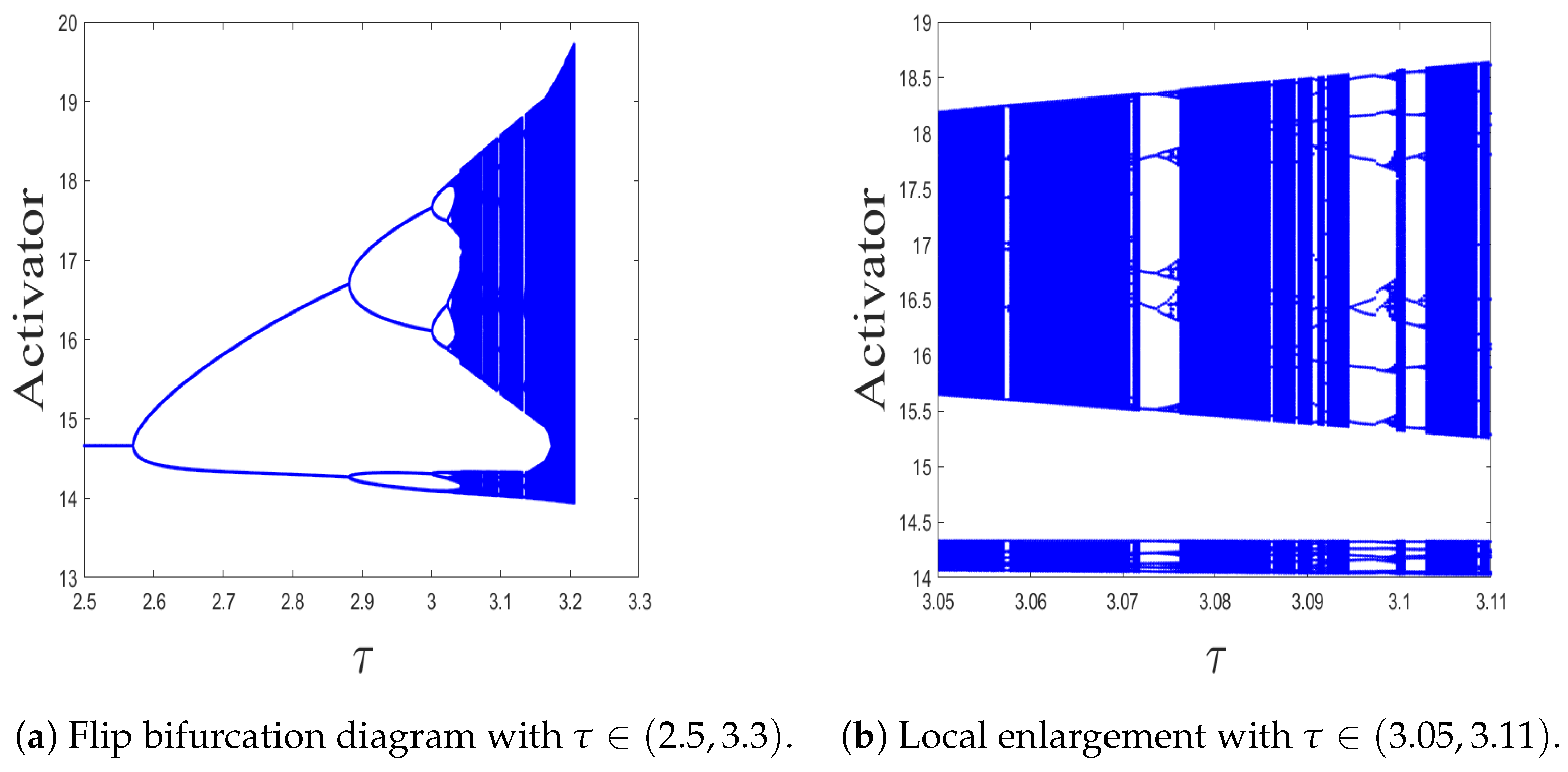
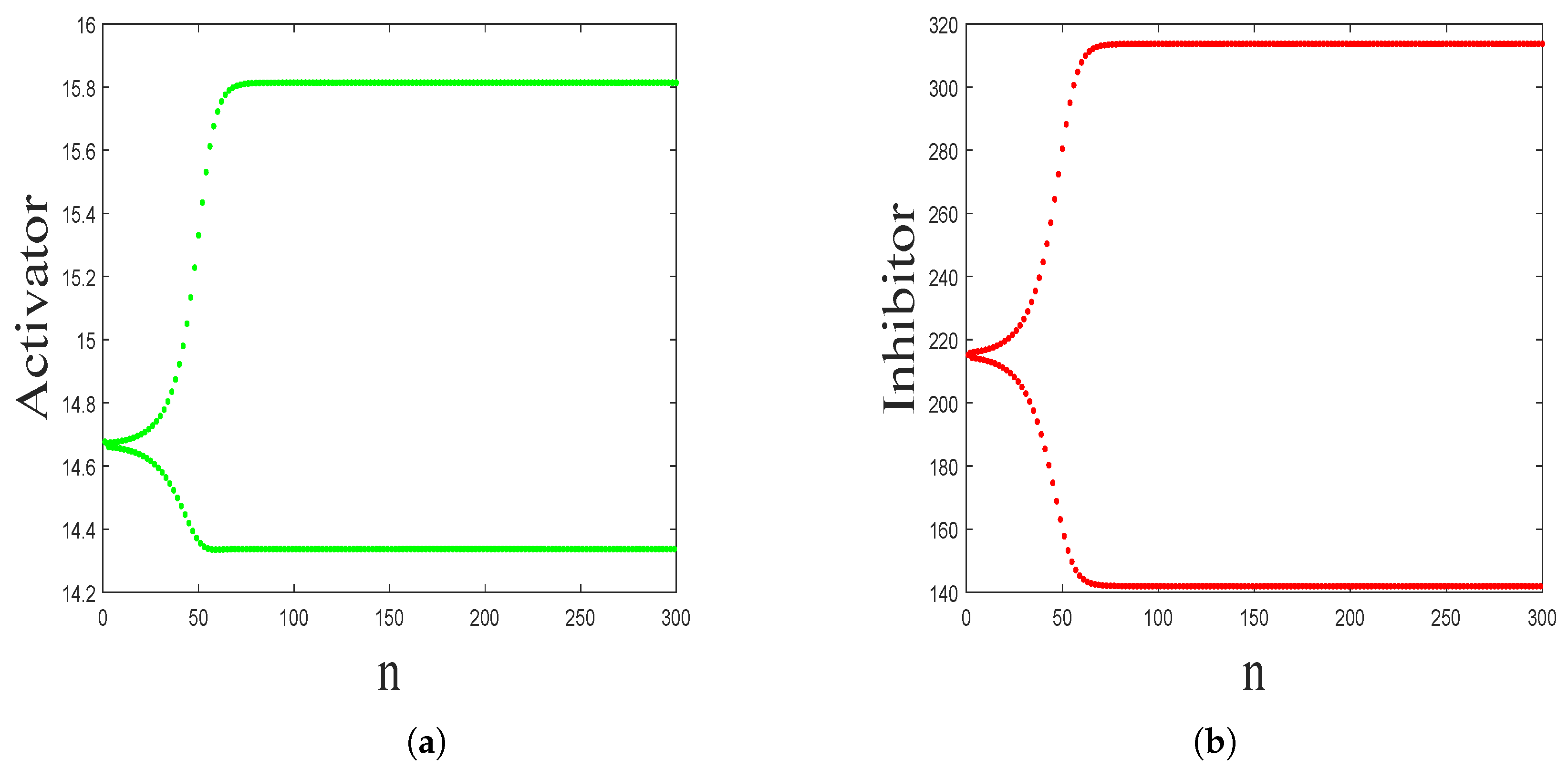
3.1. Flip Bifurcation Analysis
3.2. Turing Bifurcation Analysis
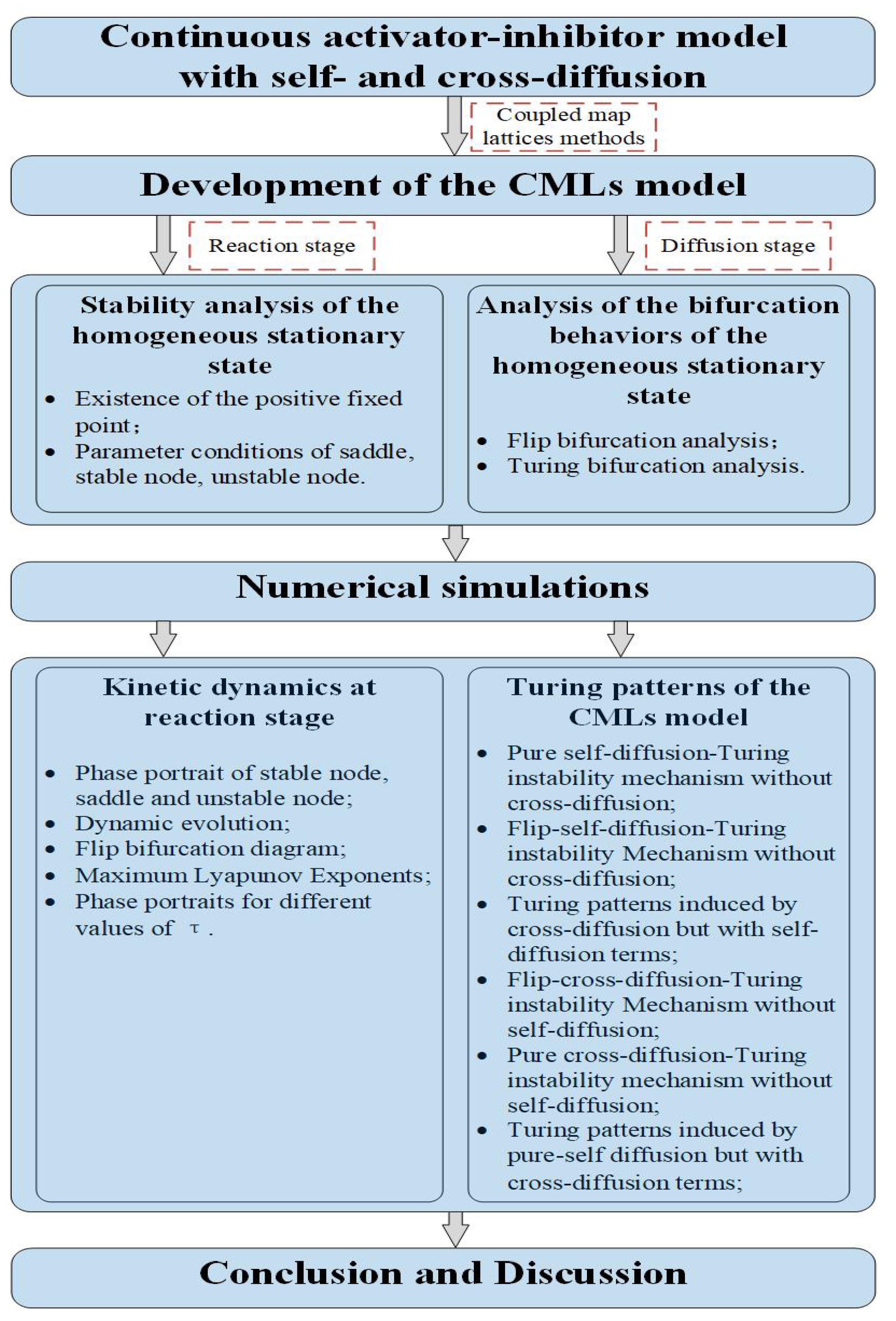
4. Numerical Simulations
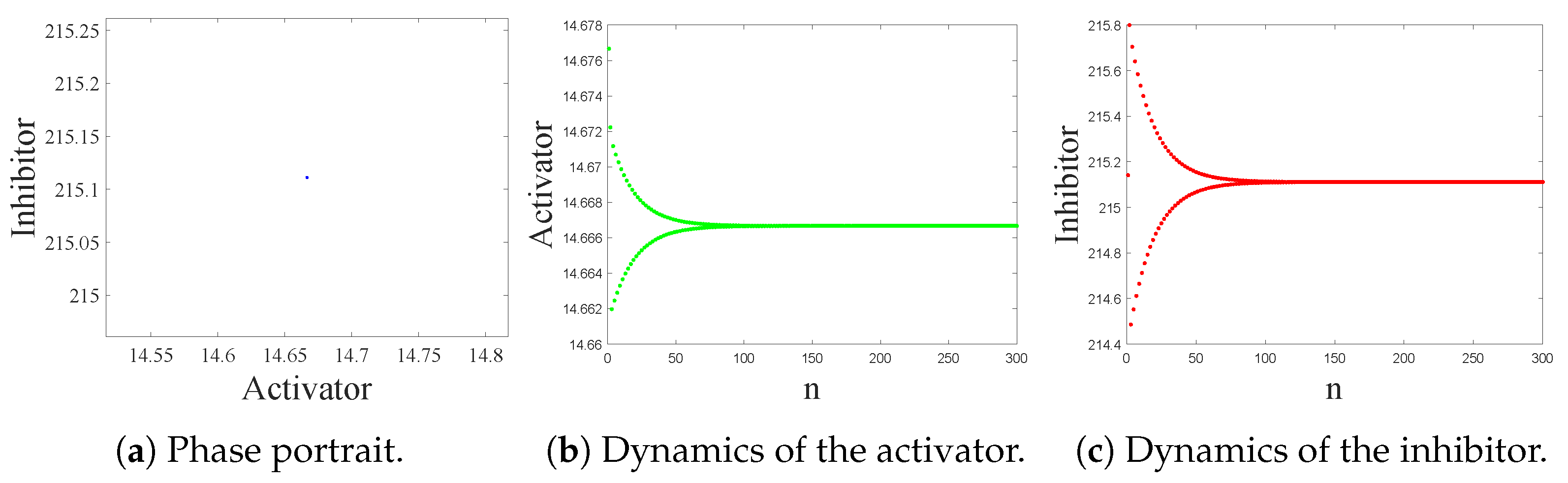
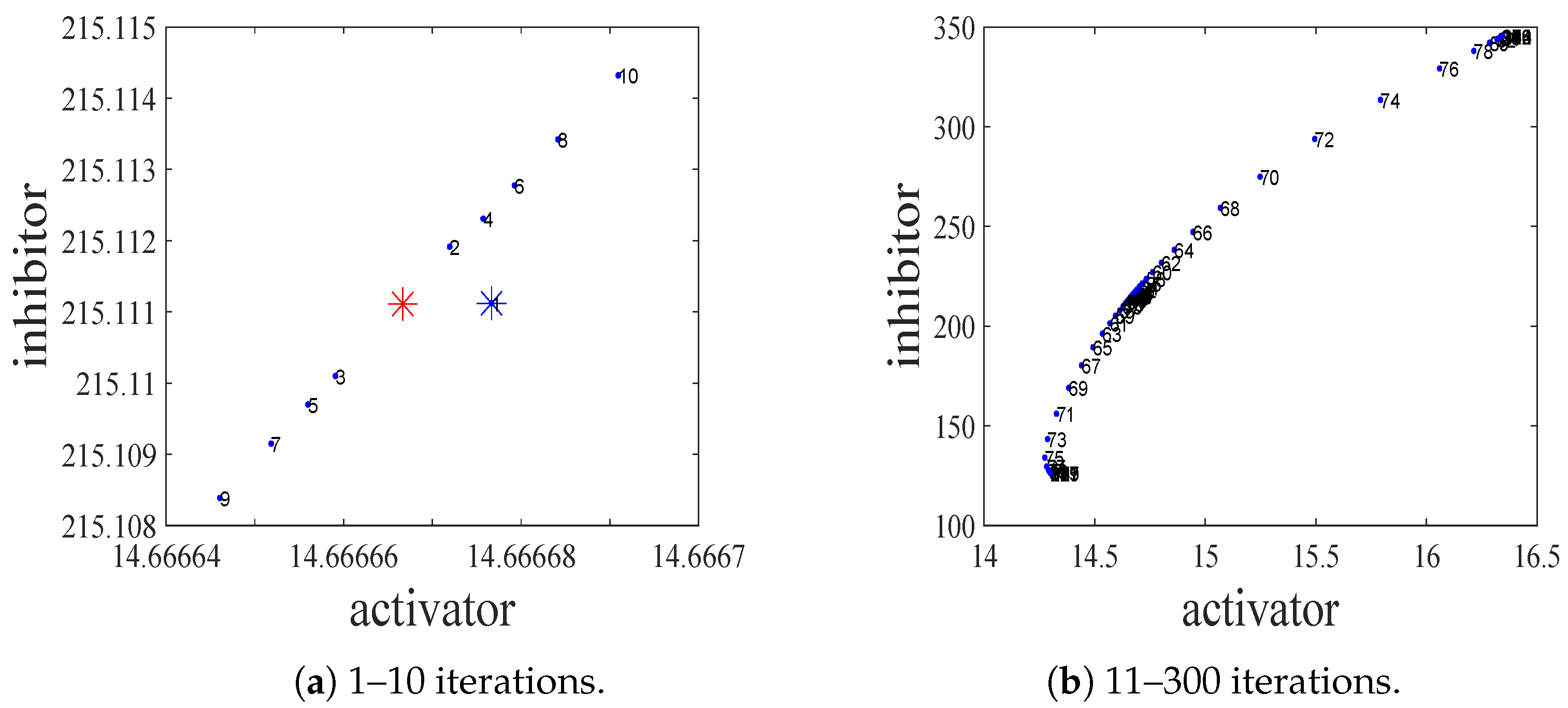
4.1. Kinetic Dynamics at Reaction Stage
4.2. Turing Patterns of the CML Model
5. Discussion and Conclusions
- •
- It is worth noting that, compared to the paper [46], our model in this paper includes an additional cross-diffusion term. In terms of spatially homogeneous dynamic behavior, the authors in [46] have already proven the existence and stability of a positive fixed point and have conducted an analysis of flip bifurcation and Neimark–Sacker bifurcation. Our work is similar to the spatially homogeneous dynamic behavior discussed in [46], while we have further discussed the types of the fixed point and determined the precise parameter conditions for the fixed point to be a saddle, a stable node, and an unstable node. For spatially heterogeneous dynamic behavior, the authors in [46] only explored the instability phenomena caused by self-diffusion, whereas our research not only analyzed the instability caused by self-diffusion but also considered the instability induced by cross-diffusion. Therefore, the mechanisms of pattern formation we have revealed are more comprehensive, and the types of patterns observed are also more abundant. For example, in our paper, the pure-self-Turing instability mechanism leads to a colorful mottled grid pattern with winding and twisted bands. The pure-cross-Turing instability mechanism leads to dense patches and twisted bands nested together. These patterns cannot be generated by the model in [46]. Furthermore, our system can also undergo a Neimarck–Sacker bifurcation. We have conducted related research on this and discovered some very interesting phenomena such as a plum blossom-shaped chaotic attractor. Moving forward, we will continue to delve deeper into this field. Compared to the continuous Gierer–Meinhardt model, our CML-based Gierer–Meinhardt model exhibits a richer dynamic behavior. Our model undergoes a flip bifurcation, which is not observed in the corresponding continuous model. The mechanisms of pattern formation in our model are more diverse than those in the corresponding continuous models [32,33,44,45]. Additionally, the types of patterns we observe are also more abundant.
- •
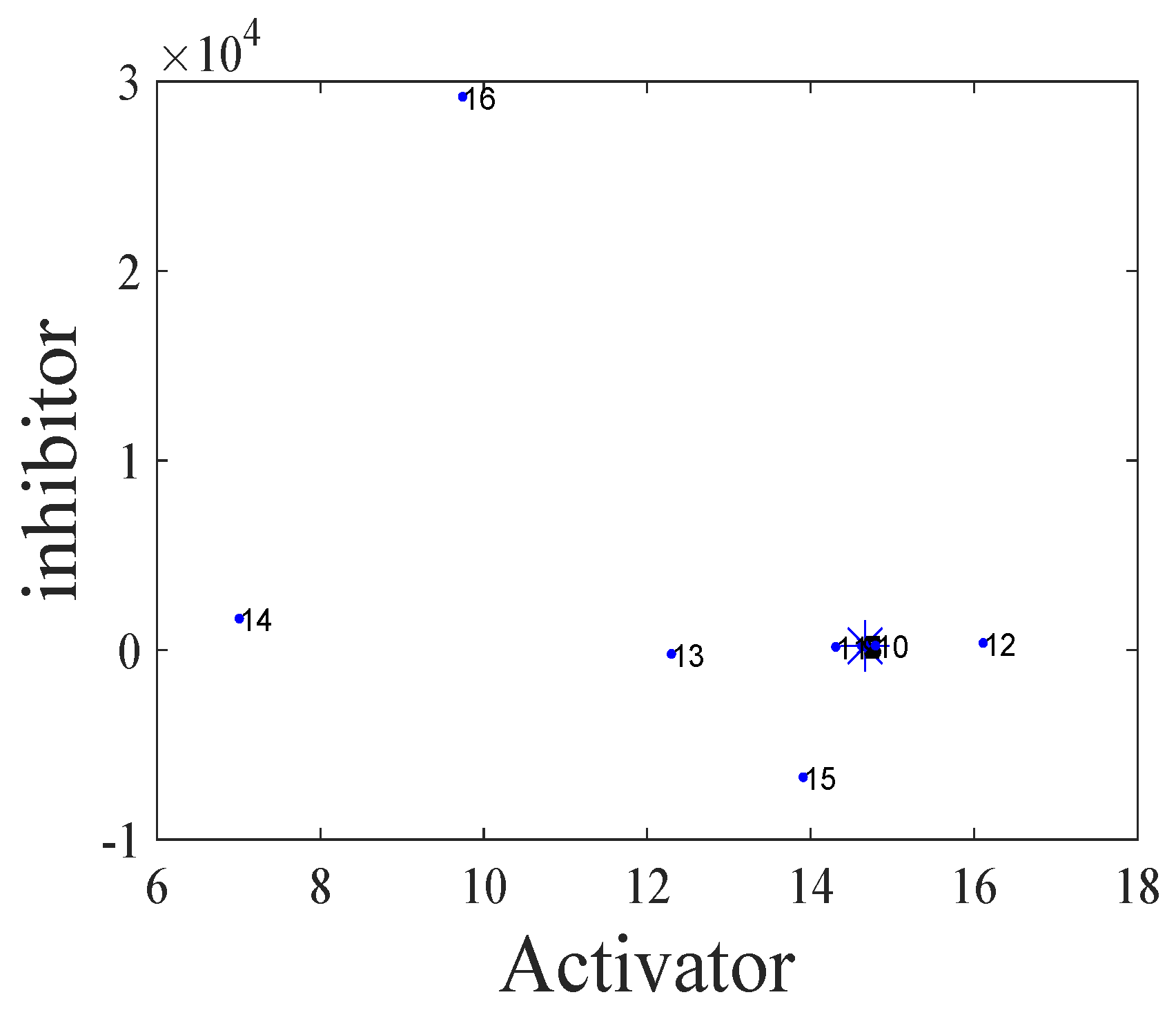
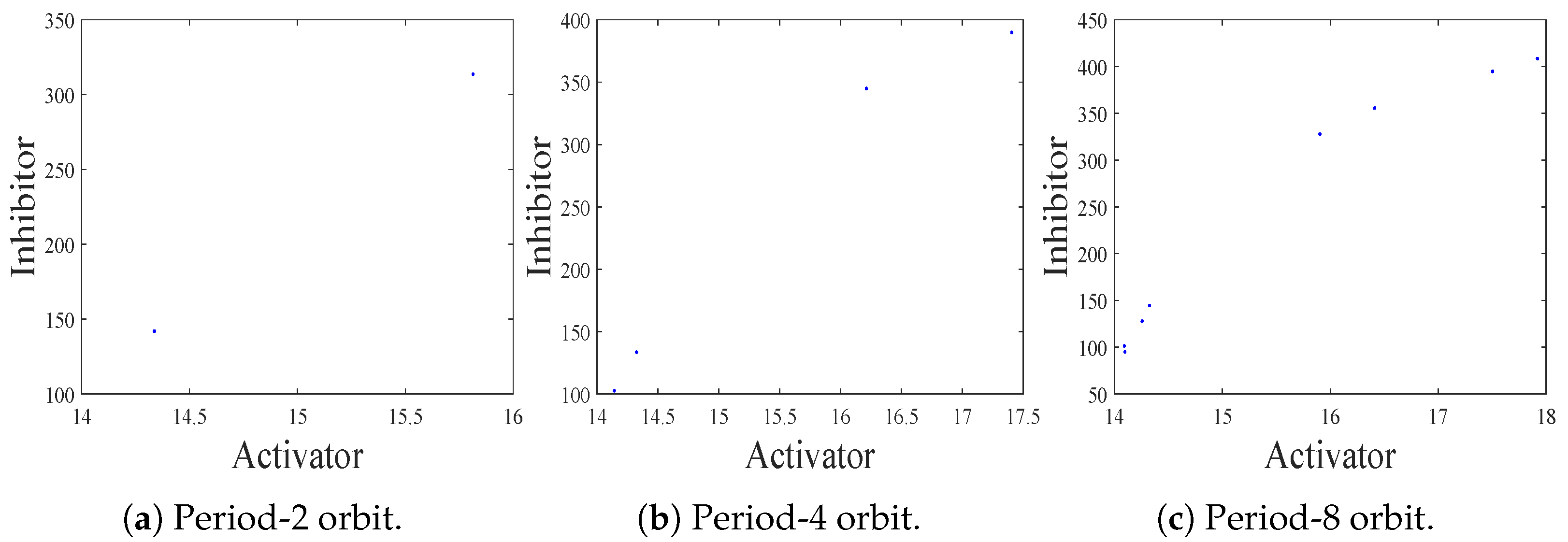
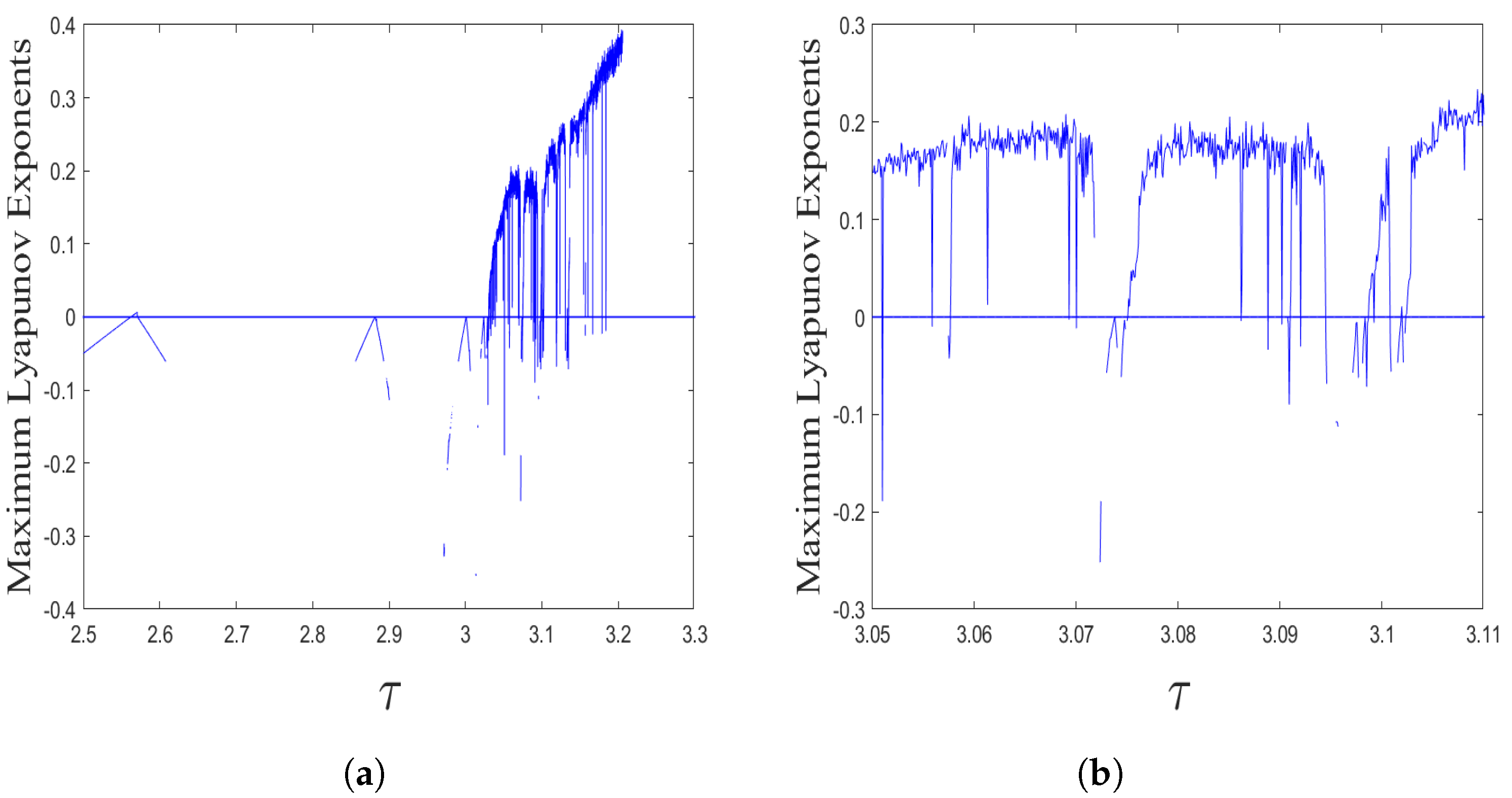
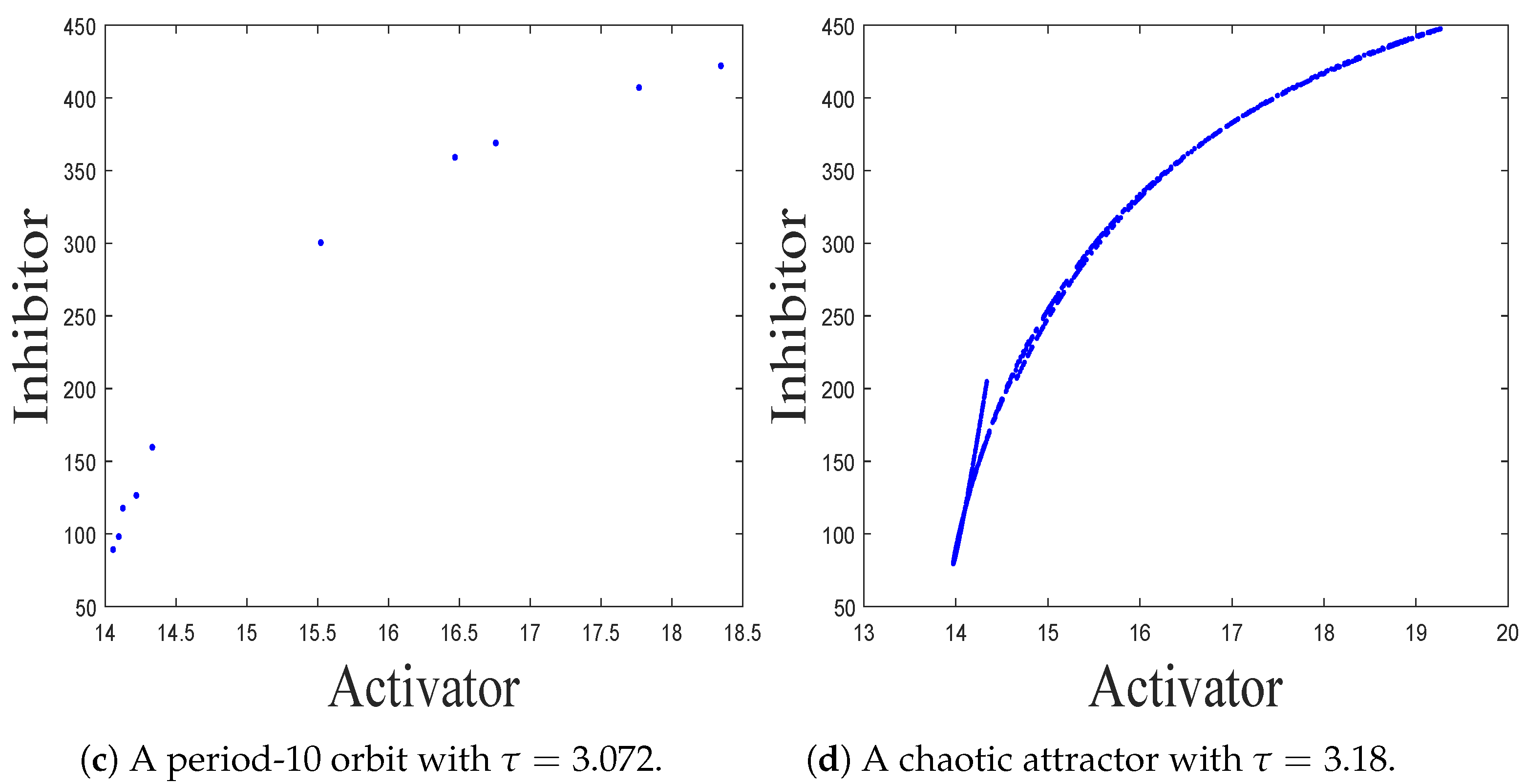
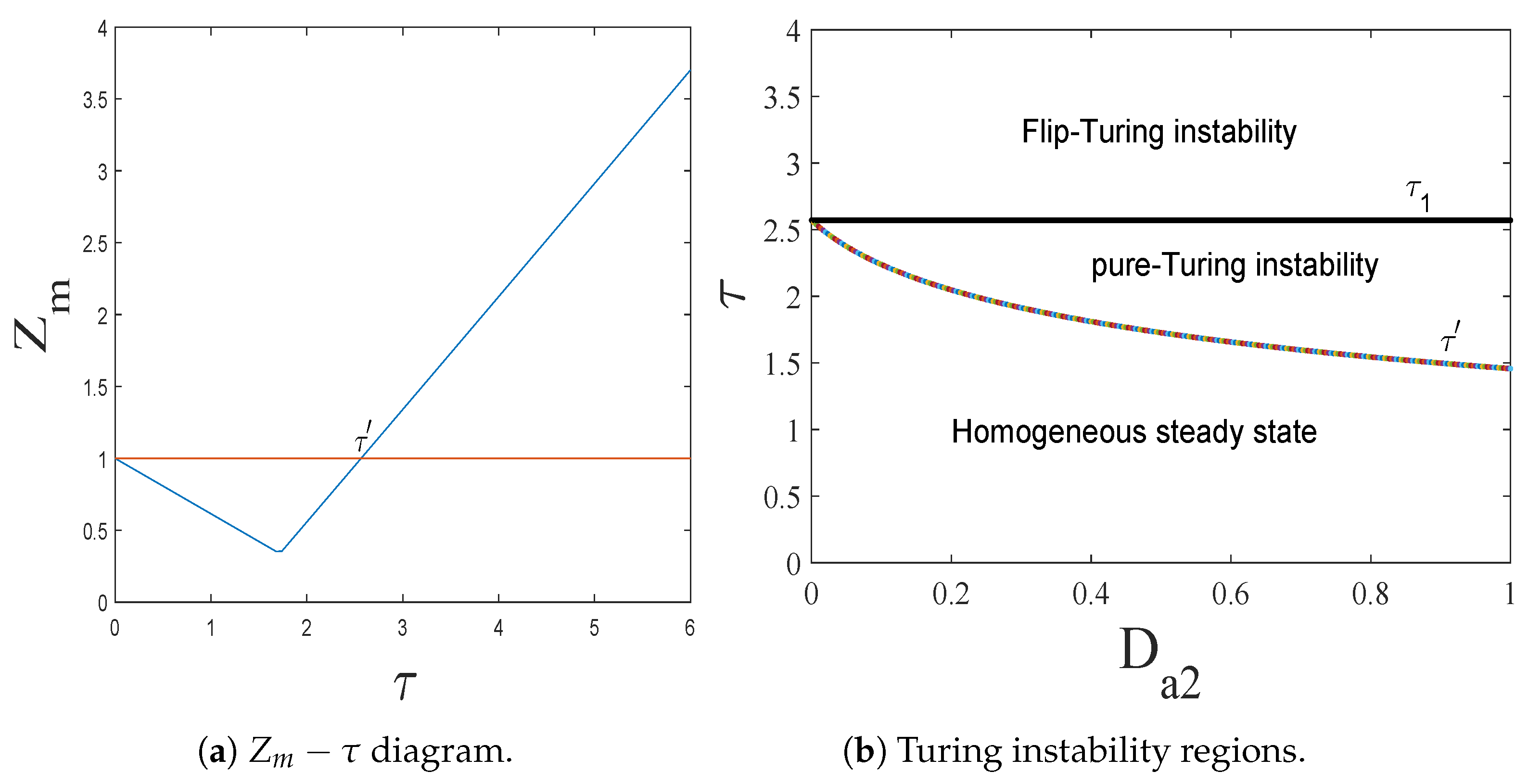
- The spatiotemporal dynamics of the CML model (4)–(7) consists of two parts: spatially homogeneous and spatially heterogeneous dynamics. The spatially homogeneous dynamics includes a stable homogeneous stationary state (stable node) as well as homogeneous oscillatory states generated by flip bifurcation. The spatially heterogeneous dynamics include the patterns induced by Turing instability. In this paper, we give the precise parameter conditions for the system to experience flip bifurcation, which induces a continuous period-doubling oscillatory process and introduces a path to chaos, allowing the system to enter a chaotic oscillatory state eventually. The maximum Lyapunov exponents can help us to distinguish the regular and irregular behaviors. On the road to chaos, we can see many periodic windows. In each window, there exists a periodic orbit.
- •
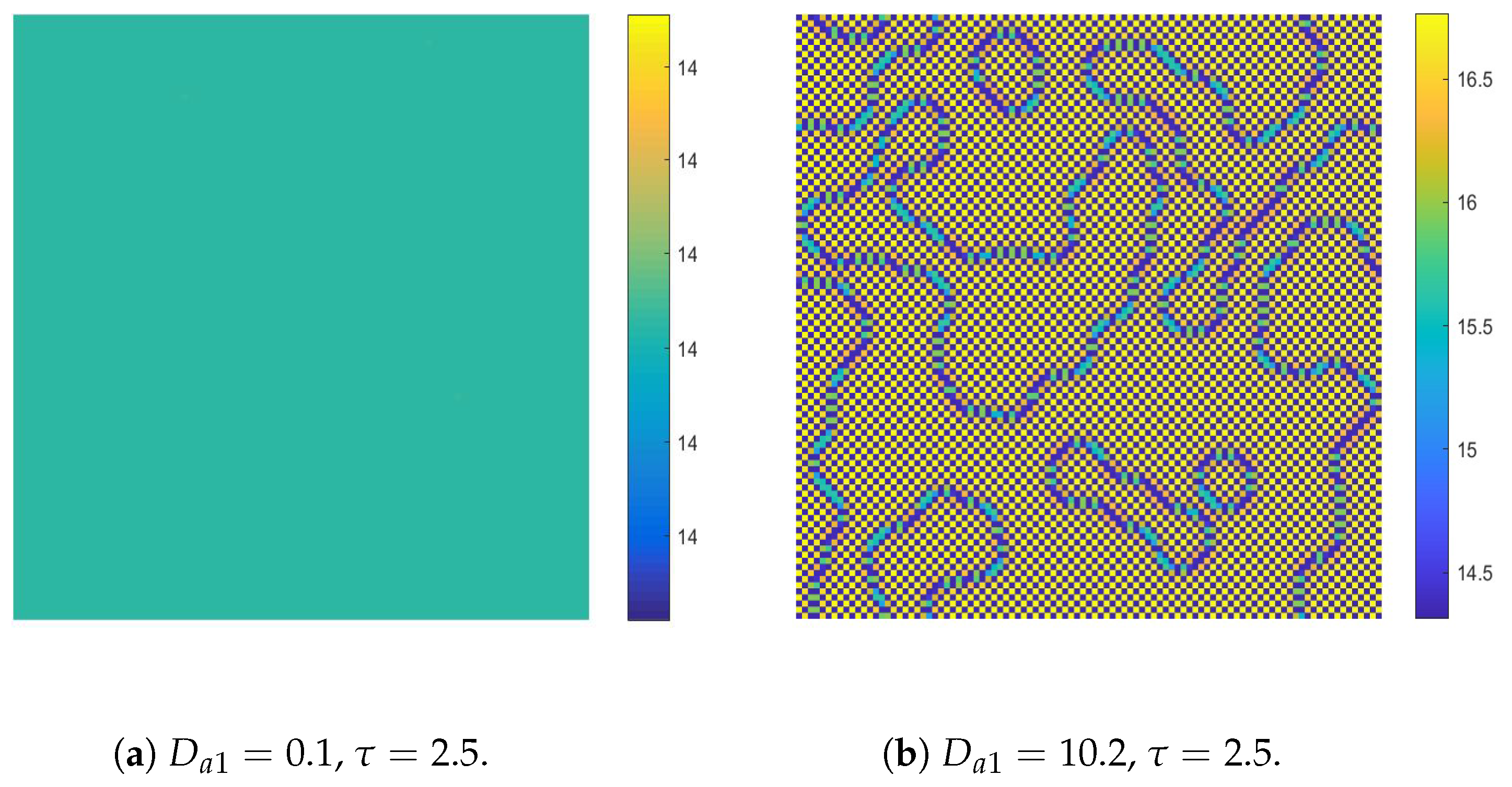
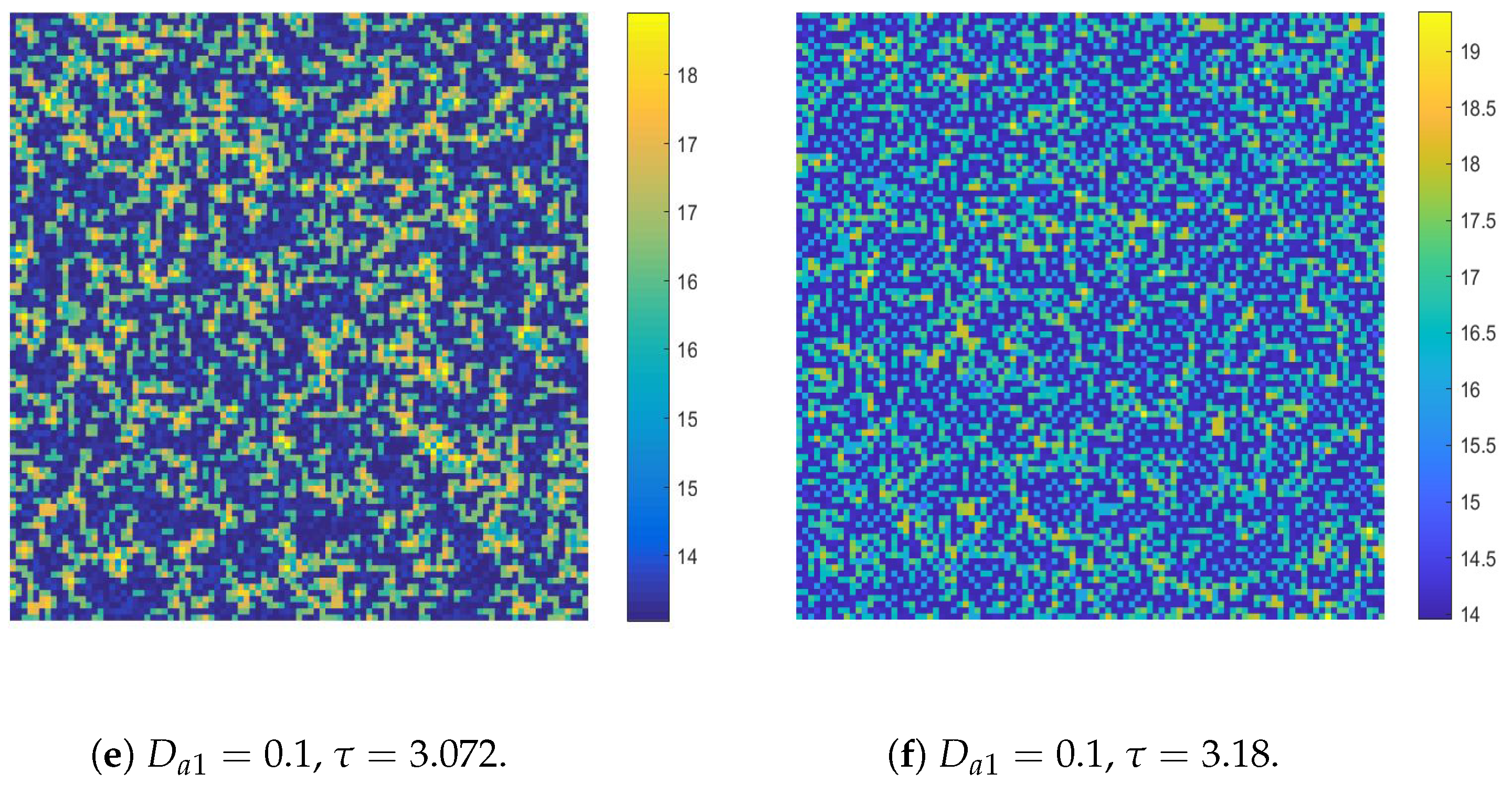
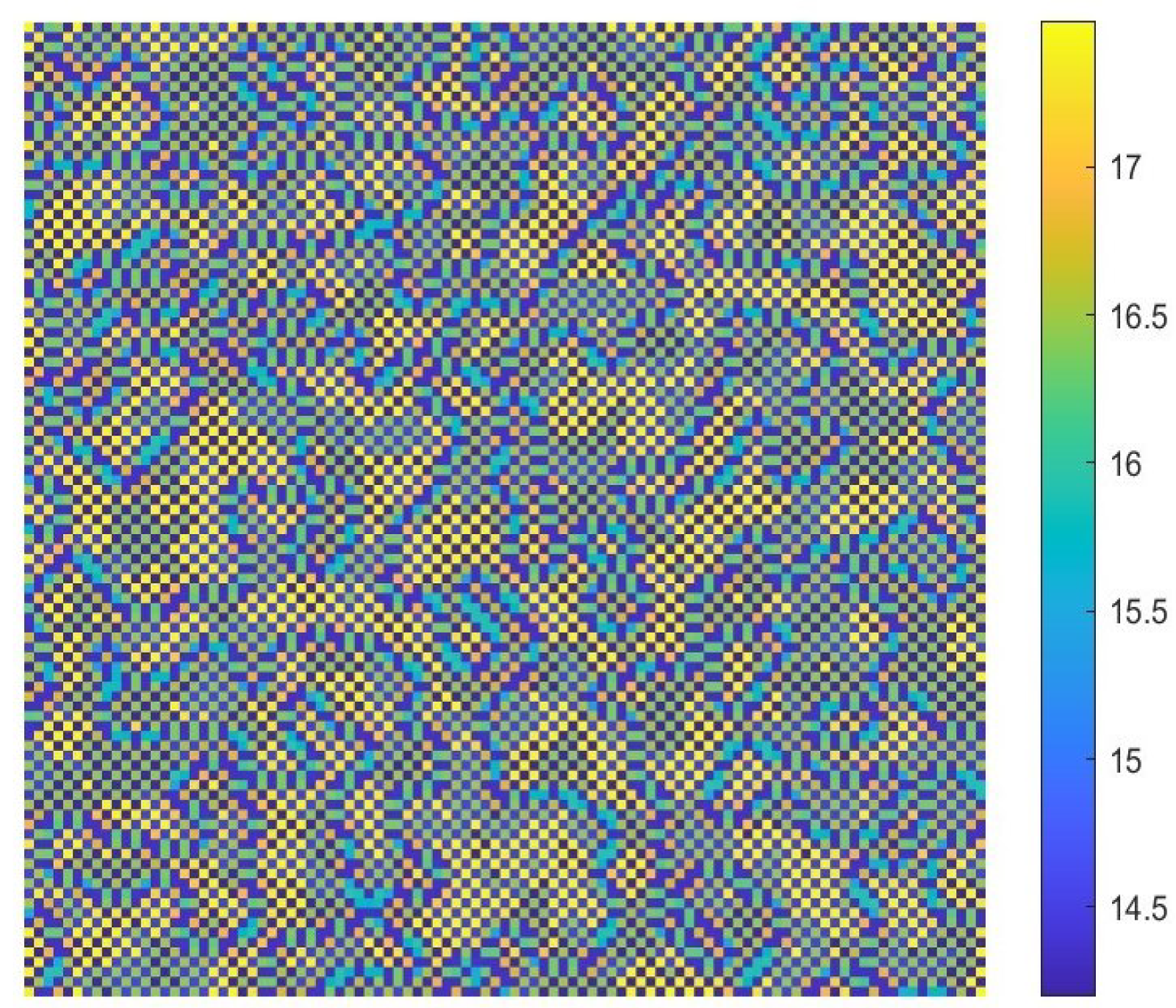
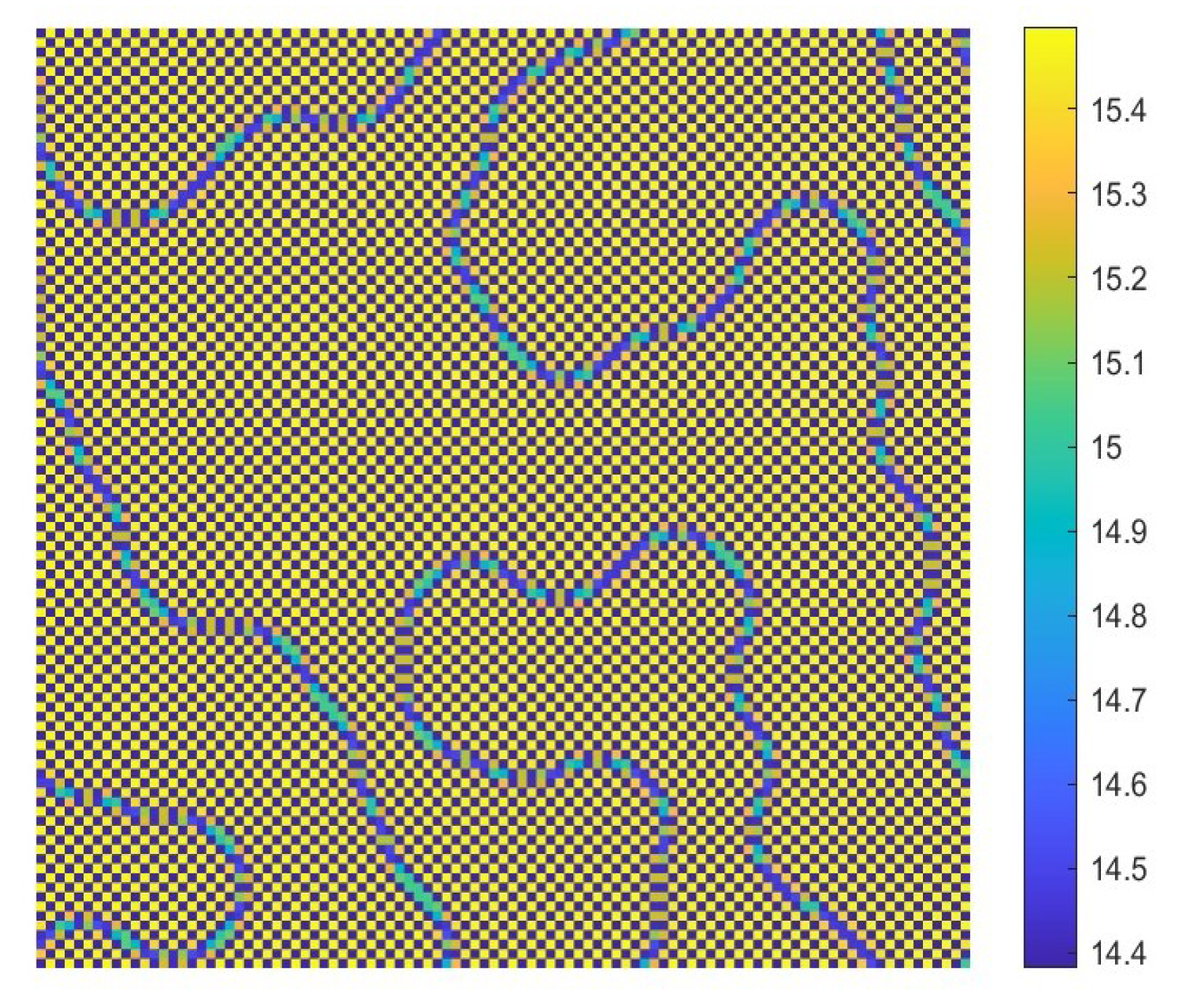
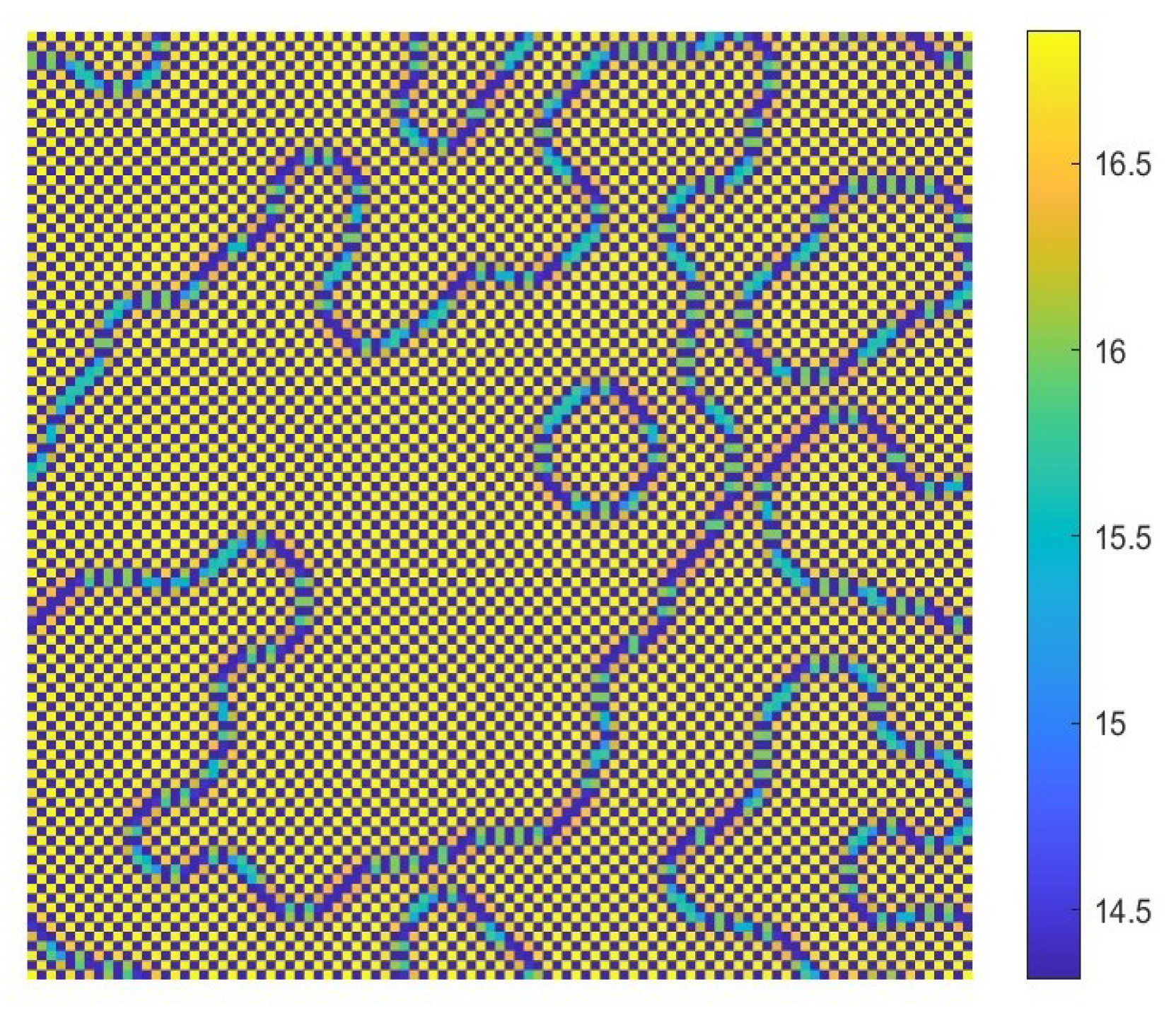
- The diffusion effects offer the possibility for the emergence of Turing bifurcation, while the homogeneous state of kinetic dynamical behavior offers the options for the CML model. During the numerical simulations, we find that the size of the Turing region is related to the spatial step . The mechanisms for the pattern formation of the CML model are divided into two broad categories: pure Turing instability mechanism and flip-Turing instability mechanism. The pure Turing instability mechanism can be further categorized into pure-self-diffusion-Turing instability mechanism and pure-cross-diffusion-Turing instability mechanism. The flip-Turing instability mechanism can be further categorized into the flip-self-diffusion-Turing instability mechanism and flip-cross-diffusion-Turing instability mechanism. The pure-self-Turing instability mechanism leads to a colorful, mottled grid pattern with winding and twisted bands. The pure-cross-Turing instability mechanism leads to dense patches and twisted bands nested together. The flip-Turing instability mechanism leads to multi-state intertwined patterns. The chaos-self-diffusion-Turing instability mechanism leads to labyrinthine and mosaic patterns. The flip-Turing instability mechanism on chaotic paths induces a spatial multiply periodization process. It leads to a pattern transition of the CML model from spatiotemporally ordered patterns to spatiotemporally disordered patterns.
- •
- Self-diffusion and cross-diffusion can both lead to Turing instability, thereby forming Turing patterns. When the system only has self-diffusion or only cross-diffusion, the resulting spatial patterns are shown in Figure 10b and Figure 15. They are all mottled grid patterns with winding and twisted bands. The patterns present natural curves in the bands and patchy distributions. However, when the system has both self-diffusion and cross-diffusion if the instability phenomenon and the formation of the patterns are primarily caused by cross-diffusion, as shown in Figure 12, the resulting spatial patterns are dense patches and twisted bands nested together. If the instability phenomenon and the formation of the patterns are primarily caused by self-diffusion, as shown in Figure 16, the resulting spatial patterns are also mottled grid patterns with winding and twisted bands. The complexity of the pattern structure in Figure 12 is higher than that in Figure 16.
- •
- The Turing patterns formed each time under random perturbations are qualitatively consistent but not identical in shape. From the simulations, we find that the patterns induced by pure-cross-diffusion-Turing instability mechanism are more coplexed than the patterns induced by pure-self-diffusion-Turing instability mechanism. It is important to note in particular that when in the second case. For the map (10), there exists a chaotic attractor, as shown in Figure 8d. But when we consider the effects of the pure cross-diffusion effects, though the CML model (4)–(7) experiences Turing instability, while we can not find the corresponding mosaic patterns similar to the patterns shown in Figure 11 which are induced by the pure-self-diffusion effects.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviation
| CMLs | Copuled map lattices. |
Appendix A
References
- Murray, J.D. Mathematical Biology I: An Introduction, 3rd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Turing, A.M. The chemical basis of morphogenesis. Philpos. Trans. R. Soc. Lond. Ser. B 1952, 237, 37–72. [Google Scholar]
- Gierer, A.; Meinhardt, H. A theory of biological pattern formation. Kybernetik 1972, 12, 30–39. [Google Scholar] [CrossRef]
- Gao, S.P.; Chang, L.L.; Perc, M.; Wang, Z. Turing patterns in simplicial complexes. Phys. Rev. E 2023, 107, 014216. [Google Scholar] [CrossRef]
- Vanag, V.K.; Epstein, I.R. Pattern formation in a tunable medium: The Belousov-Zhabotinsky reaction in an aerosol OT microemulsion. Phys. Rev. Lett. 2001, 87, 169–177. [Google Scholar] [CrossRef]
- Tan, Z.; Chen, S.; Peng, X.; Zhang, L.; Gao, C. Polyamide membranes with nanoscale Turing structures for water purification. Science 2018, 360, 518–521. [Google Scholar] [CrossRef]
- Karig, D.; Martini, K.M.; Lu, T.; Delateur, N.A.; Goldenfeld, N.; Weiss, R. Stochastic Turing patterns in a synthetic bacterial population. Proc. Natl. Acad. Sci. USA 2018, 115, 6572–6577. [Google Scholar] [CrossRef]
- Wakano, J.Y.; Nowak, M.A.; Hauert, C. Spatial dynamics of ecological public goods. Proc. Natl. Acad. Sci. USA 2009, 106, 7910–7914. [Google Scholar] [CrossRef]
- Short, M.B.; Bertozzi, A.L.; Brantingham, P.J. Nonlinear patterns in urban crime: Hotspots, bifurcations, and suppression. SIAM J. Appl. Dyn. Syst. 2010, 9, 462–483. [Google Scholar] [CrossRef]
- Fuseya, Y.; Katsuno, H.; Behnia, K.; Kapitulnik, A. Nanoscale Turing patterns in a bismuth monolayer. Nat. Phys. 2021, 17, 1031–1036. [Google Scholar] [CrossRef]
- Taylor, N.P.; Kim, H.; Krause, A.L.; Van Gorder, R.A. A non-local cross-diffusion model of population dynamics I: Emergent spatial and spatiotemporal patterns. Bull. Math. Biol. 2020, 82, 112. [Google Scholar] [CrossRef]
- Hata, S.; Nakao, H.; Mikhailov, A.S. Dispersal-induced destabilization of metapopulations and oscillatory Turing patterns in ecological networks. Sci. Rep. 2014, 4, 3585. [Google Scholar] [CrossRef]
- Kondo, S.; Miura, T. Reaction-diffusion model as a framework for understanding biological pattern formation. Science 2010, 329, 1616–1620. [Google Scholar] [CrossRef]
- Nakamasu, A.; Takahashi, G.; Kanbe, A.; Kondo, S. Interactions between zebrafish pigment cells responsible for the generation of Turing patterns. Proc. Natl. Acad. Sci. USA 2009, 106, 8429–8434. [Google Scholar] [CrossRef]
- Alessio, B.M.; Gupta, A. Diffusiophoresis-enhanced Turing patterns. Sci. Adv. 2023, 9, eadj2457. [Google Scholar] [CrossRef]
- Rui, X.; Gao, Q.Y.; Azaele, S.; Sun, Y.Z. Effects of noise on the critical points of Turing instability in complex ecosystems. Phys. Rev. E 2023, 108, 014407. [Google Scholar]
- Huang, T.S.; Zhang, H.Y. Bifurcation, chaos and pattern formation in a space- and time-discrete predator-prey system. Chaos Solitons Fractals 2016, 91, 92–107. [Google Scholar] [CrossRef]
- Li, Y.; Cao, J.J.; Sun, Y.; Song, D.; Wu, X.Y. Spatiotemporal patterns induced by four mechanisms in a tussock sedge model with discrete time and space variables. Adv. Differ. Equ. 2021, 399, 92–107. [Google Scholar] [CrossRef]
- Nakao, H.; Mikhailov, A.S. Turing patterns in network organized activator inhibitor systems. Nat. Phys. 2010, 6, 544–550. [Google Scholar] [CrossRef]
- Horsthemke, W.; Lam, K.; Moore, P.K. Network topology and Turing instabilities in small arrays of diffusively coupled reactors. Phys. Lett. A 2004, 328, 444–451. [Google Scholar] [CrossRef]
- Zheng, Q.Q.; Shen, J.W.; Horsthemke, Y.W.; Lam, K.; Moore, P.K. Turing instability in the reaction-diffusion network. Phys. Rev. E 2020, 102, 1–9. [Google Scholar] [CrossRef]
- Li, Z.J.; Fang, S.Y.; Ma, M.L.; Wang, M.J. Bursting Oscillations and Experimental Verification of a Rucklidge System. Int. J. Bifurcat. Chaos 2021, 31, 1–13. [Google Scholar] [CrossRef]
- Mistro, D.C.; Rodrigues, L.A.D.; Petrovskii, S. Spatiotempral complexity of biological invasion in a space and time discrete predator-prey system with strong Allee effect. Ecol. Complex 2012, 9, 16–32. [Google Scholar] [CrossRef]
- Rodrigues, L.A.D.; Mistro, D.C.; Petrovskii, S. Pattern formation in a space- and time-discrete predator-prey system with strong Allee effect. Theor. Ecol. 2012, 12, 43–57. [Google Scholar] [CrossRef]
- Kaneko, K. Pattern dynamics in spatiotemporal chaos: Pattern secletion, diffusion of defect and pattern competition intermettency. Physica D 1989, 34, 1–41. [Google Scholar] [CrossRef]
- Kaneko, K. Spatiotemporal chaos in one and two dimensional coupled map lattices. Physica D 1989, 37, 60–82. [Google Scholar] [CrossRef]
- Han, Y.T.; Han, B.; Zhang, L.; Xu, L.; Li, M.F.; Zhang, G. Turing instability and wave patterns for a symmetric discrete competitive Lotka-Volteera system. WSEAS Trans. Math. 2011, 10, 181–189. [Google Scholar]
- May, R.M. Simple mathematical models with very complicated dynamics. Nature 1976, 261, 459–467. [Google Scholar] [CrossRef]
- Liu, X.; Xiao, D. Complex dynamic behaviors of a discrete-time predator prey system. Chaos Solition Fractals 2007, 32, 80–94. [Google Scholar] [CrossRef]
- Woodward, D.E.; Tyson, R.; Myerscough, M.R.; Murray, J.D.; Budrene, E.O.; Berg, H.C. Spatio-Temporal Patterns Generated by Salmonella typhimurium. Biophys. J. 1995, 68, 2181–2189. [Google Scholar] [CrossRef]
- Brenner, M.P.; Levitov, L.S.; Budrene, E.O. Physical Mechanisms for Chemotactic Pattern Formation by Bacteria. Biophys. J. 1998, 74, 1677–1693. [Google Scholar] [CrossRef]
- Li, Y.; Wang, J.L.; Hou, X.J. Stripe and spot patterns for general Gierer-Meinhardt model with common sources. Int. J. Bifurcat. Chaos 2017, 27, 1750018. [Google Scholar] [CrossRef]
- Li, Y.; Wang, J.L.; Hou, X.J. Stripe and spot patterns for Gierer-Meinhardt model with saturated activator production. J. Math. Anal. Appl. 2017, 449, 1863–1879. [Google Scholar] [CrossRef]
- Liu, J.; Yi, F.; Wei, J. Multiple bifurcation analysis and spatiotemporal patterns in a 1-d geierer meinhardt model of morphogenesis. Int. J. Bifurcat. Chaos 2010, 20, 1007–1025. [Google Scholar] [CrossRef]
- Wu, R.; Zhou, Y.; Shao, Y.; Chen, L. Bifurcation and turing patterns of reaction-diffusion activator-inhibitor model. Physica A 2017, 482, 597–610. [Google Scholar] [CrossRef]
- Zhong, S.; Wang, J.L.; Xia, J.D.; Li, Y. Spatiotemporal dynamics and pattern formations of an activator-substrate model with double saturation terms. Int. J. Bifurc. Chaos 2021, 31, 2150129. [Google Scholar] [CrossRef]
- Yang, R.; Yu, X. Turing-Hopf bifurcation in diffusive Gierer-Meinhardt model. Int. J. Bifurc. Chaos 2022, 32, 2250046. [Google Scholar] [CrossRef]
- Ma, X.Y.; Wang, J.L.; Zhu, Y.H.; Wang, Z.W.; Sun, Y. Turing-Hopf Bifurcation Coinduced by Diffusion and Delay in Gierer-Meinhardt Systems. Int. J. Bifurc. Chaos 2024, 34, 2450162. [Google Scholar] [CrossRef]
- Zhao, S.; Wang, H.; Jiang, W. Turing-Hopf bifurcation and spatiotemporal patterns in a Gierer-Meinhardt system with gene expression delay. Nonlinear Anal. Model. Control 2021, 26, 461–481. [Google Scholar] [CrossRef]
- Zhao, S.; Wang, H. Turing–Turing bifurcation and multi-stable patterns in a Gierer–Meinhardt system. Appl. Math. Model 2022, 112, 632–648. [Google Scholar] [CrossRef]
- Wu, R.C.; Yang, L.L. Bogdanov-Takens Bifurcation of Codimension 3 in the Gierer-Meinhardt Model. Int. J. Bifurc. Chaos 2023, 33, 2350163. [Google Scholar] [CrossRef]
- Lv, J.P.; Jing, H.F. Analyzing the dynamic behavior of the Gierer-Meinhardt model using finite difference method. AIP Adv. 2024, 14, 085215. [Google Scholar] [CrossRef]
- Mai, F.X.; Qin, L.J.; Zhang, G. Turing instability for a semi-discrete Gierer Meinhardt system. Physica A 2012, 391, 2014–2022. [Google Scholar] [CrossRef]
- Wang, J.L.; Li, Y.; Zhong, S.H.; Hou, X.J. Analysis of bifurcation, chaos and pattern formation in a discrete time and space Gierer Meinhardt system. Chaos Solitons Fractals 2019, 118, 1–17. [Google Scholar] [CrossRef]
- Zhu, Y.H.; Li, Y.; Ma, X.Y.; Sun, Y.; Wang, Z.W.; Wang, J.L. Bifurcation and Turing Pattern Analysis for a Spatiotemporal Discrete Depletion Type Gierer-Meinhardt Model with Self-Diffusion and Cross-Diffusion. J. Appl. Anal. Comput. 2025, 15, 705–733. [Google Scholar]
- Liu, B.; Wu, R.C. Bifurcation and Patterns Analysis for a Spatiotemporal Discrete Gierer-Meinhardt System. Mathematics 2022, 10, 243. [Google Scholar] [CrossRef]
- Nakata, K.; Sokabe, M.; Suzuki, R. The application of the Gierer Meinhardt equations to the development of the retinotectal projection. Biol. Cybern 1979, 35, 235–241. [Google Scholar] [CrossRef]
- Meinhardt, H. Models of Biological Pattern Formation; Academic Press: Cambridge, MA, USA, 1982. [Google Scholar]
- Meinhardt, H. The Algorithmic Beauty of Sea Shells; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Koch, A.; Meinhardt, H. Biological pattern formation: From basic mechanisms to complex structures. Rev. Mod. Phys. 1994, 66, 1481. [Google Scholar] [CrossRef]
- Punithan, D.; Kim, D.K.; Mckay, R. Spatio-temporal dynamics and quantification of daisyworld in two-dimensional coupled map lattices. Ecol. Complex 2012, 9, 43–57. [Google Scholar] [CrossRef]
- Nayfeh, A.H.; Balachandran, B. Applied Nonlinear Dynamics: Analytical, Computational, and Experimental Methods, 1st ed.; Wiley: Weinheim, Germany, 1952. [Google Scholar]
- Guckenheimer, J.; Holmes, P. Nonlinear Oscillations, Dynamical Systems and Bifurcations of Vector Fields; Springer: New York, NY, USA, 1983; pp. 117–165. [Google Scholar]
- Abid, W.; Yafia, R.; Aziz-Alaoui, M.A.; Bouhata, H.; Abichou, A. Diffusion driven instability and Hopf bifurcation in spatial predator-prey model on a circualr domain. Appl. Math. Comput. 2015, 260, 292–313. [Google Scholar]
- Chang, L.; Sun, G.Q.; Jin, Z. Rich dynamics in a spatial predator-prey model with delay. Appl. Math. Comput. 2015, 256, 540–550. [Google Scholar] [CrossRef]
- Bai, L.; Zhang, G. Nontrival solutions for a nonlinear discrete elliptic equation with periodic boundary conditions. Appl. Math. Comput. 2009, 210, 321–333. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | ||||||||
|---|---|---|---|---|---|---|---|---|
| 2.57048 | 5.1871 | −0.7495 | ||||||
| 2.57048 | 5.1871 | −0.7495 | ||||||
| 2.57048 | 5.1871 | −0.7495 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Sun, Y.; Luo, J.; Pang, J.; Liu, B. Pattern Formation Mechanisms of Spatiotemporally Discrete Activator–Inhibitor Model with Self- and Cross-Diffusions. Fractal Fract. 2024, 8, 743. https://doi.org/10.3390/fractalfract8120743
Li Y, Sun Y, Luo J, Pang J, Liu B. Pattern Formation Mechanisms of Spatiotemporally Discrete Activator–Inhibitor Model with Self- and Cross-Diffusions. Fractal and Fractional. 2024; 8(12):743. https://doi.org/10.3390/fractalfract8120743
Chicago/Turabian StyleLi, You, Ying Sun, Jingyu Luo, Jiayi Pang, and Bingjie Liu. 2024. "Pattern Formation Mechanisms of Spatiotemporally Discrete Activator–Inhibitor Model with Self- and Cross-Diffusions" Fractal and Fractional 8, no. 12: 743. https://doi.org/10.3390/fractalfract8120743
APA StyleLi, Y., Sun, Y., Luo, J., Pang, J., & Liu, B. (2024). Pattern Formation Mechanisms of Spatiotemporally Discrete Activator–Inhibitor Model with Self- and Cross-Diffusions. Fractal and Fractional, 8(12), 743. https://doi.org/10.3390/fractalfract8120743