
Applying Acoustic Scattering Layer Descriptors to Depict Mid-Trophic Pelagic Organisation: The Case of Atlantic African Large Marine Ecosystems Continental Shelf


 , and
, and
Abstract
1. Introduction
2. Materials and Methods
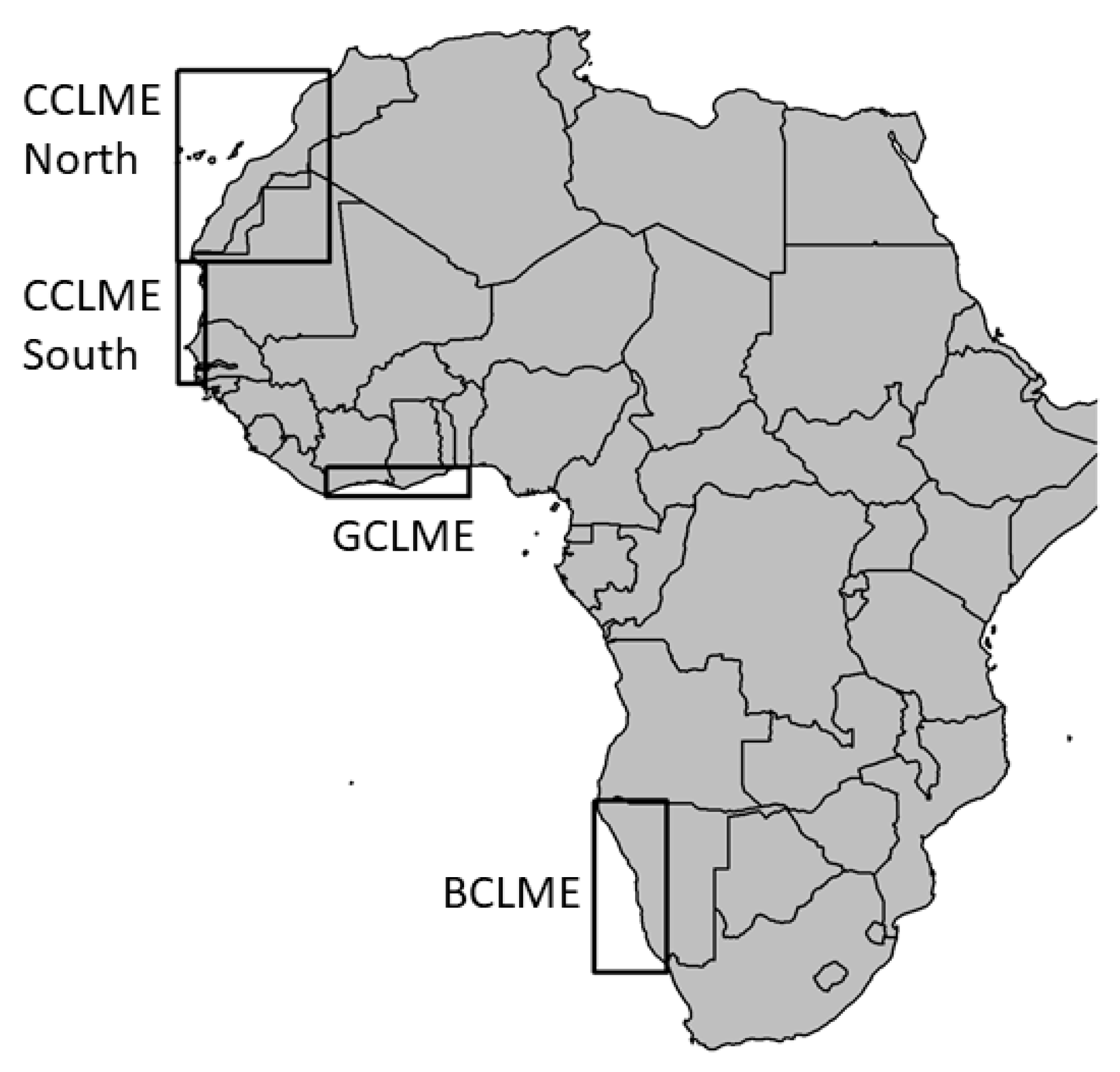
2.1. Material: Annual Acoustics Survey in African Atlantic Large Marine Ecosystems
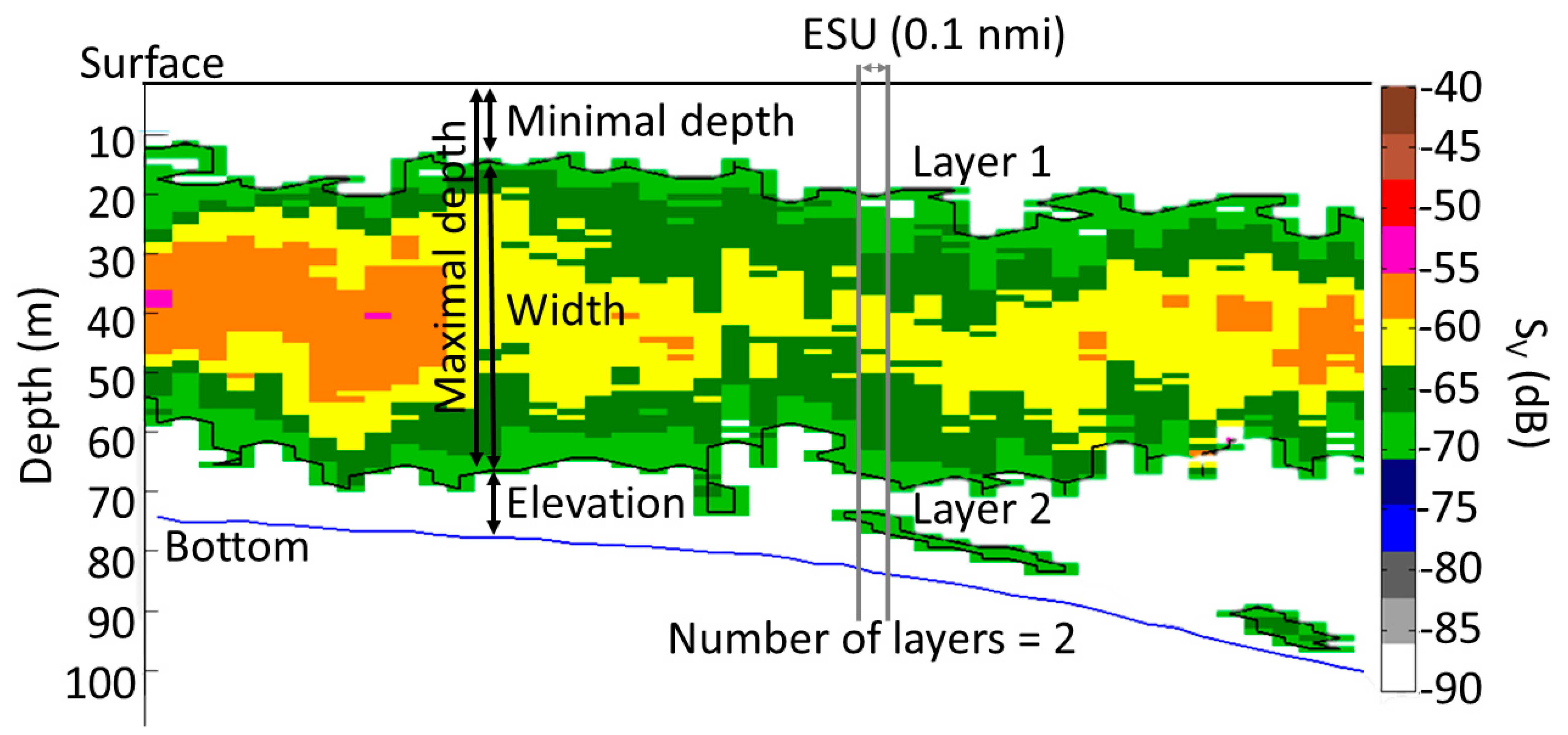
2.2. Methods: Fisheries Acoustics and Sound Scattering Layers
3. Results
3.1. Spatial Variability of Sound Scattering Layers
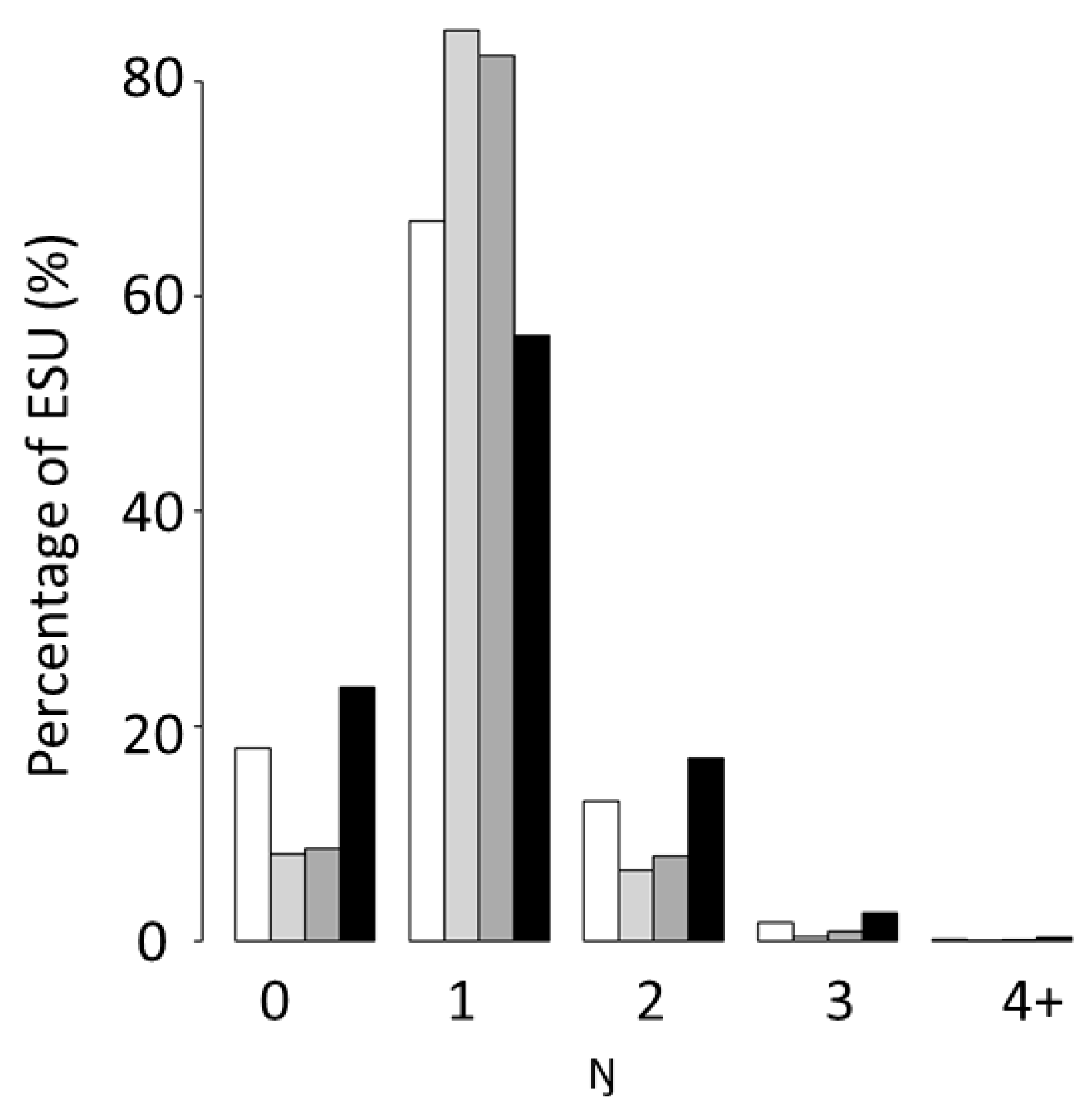
3.1.1. Number of Sound Scattering Layers (Ŋ) in the Water Column
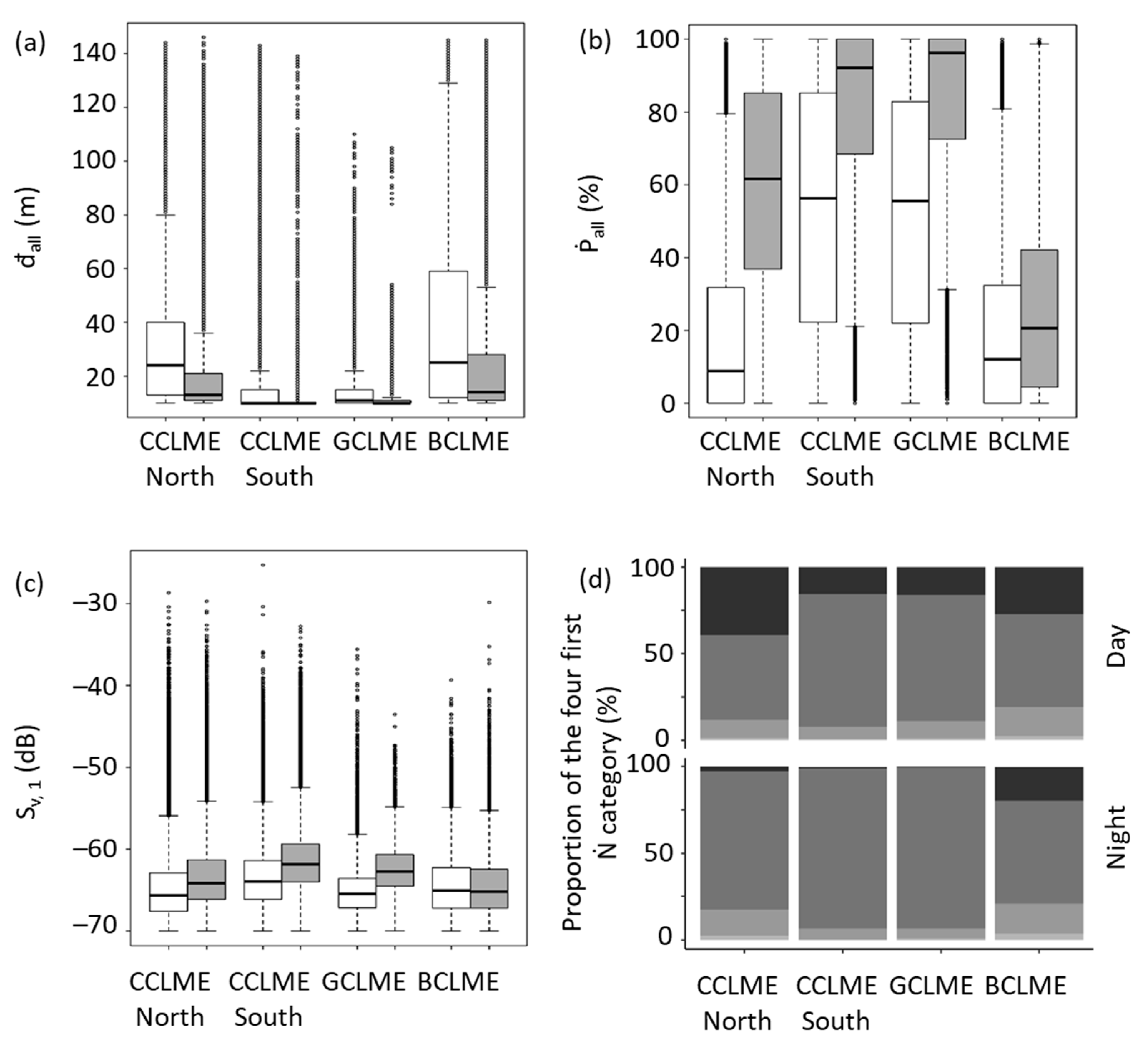
3.1.2. Minimum Depth of All SSLs (đall)
3.1.3. Proportion of Water Column Occupied (Ṗall)
3.1.4. Mean Volume Backscattering Strength of First Sound Scattering Layer (Sv, 1)
3.2. Comparative Analysis of Diel Differences among Marine Ecosystems
3.3. Inter-Annual Variability and Trends in the Atlantic African Large Marine Ecosystems
3.4. Global Analyse of Correlations between SSL Variables
4. Discussion
4.1. Relevance of Choice of Variables
4.2. Comparative Analysis of Sound-Scattering-Layer Variables within and between Atlantic African Large Marine Ecosystems
4.3. Comparison of Diel Vertical Migration
4.4. Annual Variability and Trends in Atlantic African Large Marine Ecosystems
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brehmer, P.A.J.-P. Fisheries Acoustics: Theory and Practice, 2nd ed. Fish Fish. 2006, 7, 227–228. [Google Scholar] [CrossRef]
- Simmonds, E.J.; MacLennan, D.N. Fisheries Acoustics: Theory and Practice, 2nd ed.; Fish and Aquatic Resources Series; Blackwell Science: Oxford, UK; Ames, IA, USA, 2005; ISBN 978-0-632-05994-2. [Google Scholar]
- Draštík, V.; Godlewska, M.; Balk, H.; Clabburn, P.; Kubečka, J.; Morrissey, E.; Hateley, J.; Winfield, I.J.; Mrkvička, T.; Guillard, J. Fish Hydroacoustic Survey Standardization: A Step Forward Based on Comparisons of Methods and Systems from Vertical Surveys of a Large Deep Lake. Limnol. Oceanogr. Methods 2017, 15, 836–846. [Google Scholar] [CrossRef]
- Sund, O. Echo Sounding in Fishery Research. Nature 1935, 135, 953. [Google Scholar] [CrossRef]
- Klemas, V. Fisheries Applications of Remote Sensing: An Overview. Fish. Res. 2013, 148, 124–136. [Google Scholar] [CrossRef]
- MacLennan, D.N.; Fernandes, P.G.; Dalen, J. A Consistent Approach to Definitions and Symbols in Fisheries Acoustics. ICES J. Mar. Sci. 2002, 59, 365–369. [Google Scholar] [CrossRef]
- Ballón, M.; Bertrand, A.; Lebourges-Dhaussy, A.; Gutiérrez, M.; Ayón, P.; Grados, D.; Gerlotto, F. Is There Enough Zooplankton to Feed Forage Fish Populations off Peru? An Acoustic (Positive) Answer. Prog. Oceanogr. 2011, 91, 360–381. [Google Scholar] [CrossRef]
- Cabreira, A.G.; Madirolas, A.; Brunetti, N.E. Acoustic Characterization of the Argentinean Short-Fin Squid Aggregations. Fish. Res. 2011, 108, 95–99. [Google Scholar] [CrossRef]
- Korneliussen, R.J.; Ona, E. An Operational System for Processing and Visualizing Multi-Frequency Acoustic Data. ICES J. Mar. Sci. 2002, 59, 293–313. [Google Scholar] [CrossRef]
- Ritz, D.A.; Hobday, A.J.; Montgomery, J.C.; Ward, A.J.W. Chapter Four—Social Aggregation in the Pelagic Zone with Special Reference to Fish and Invertebrates. In Advances in Marine Biology; Lesser, M., Ed.; Advances in Marine Biology; Academic Press: Cambridge, MA, USA, 2011; Volume 60, pp. 161–227. [Google Scholar]
- Remond, B. Les Couches Diffusantes Du Golfe de Gascogne: Caractérisation Acoustique, Composition Spécifique et Distribution Spatiale. Ph.D. Thesis, Université Pierre et Marie Curie, Paris, France, 2015. [Google Scholar]
- Greene, C.H.; Wiebe, P.H.; Pelkie, C.; Benfield, M.C.; Popp, J.M. Three-Dimensional Acoustic Visualization of Zooplankton Patchiness. Deep Sea Res. Part II Top. Stud. Oceanogr. 1998, 45, 1201–1217. [Google Scholar] [CrossRef]
- Lawson, G.L.; Wiebe, P.H.; Ashjian, C.J.; Gallager, S.M.; Davis, C.S.; Warren, J.D. Acoustically-Inferred Zooplankton Distribution in Relation to Hydrography West of the Antarctic Peninsula. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2004, 51, 2041–2072. [Google Scholar] [CrossRef]
- Béhagle, N.; du Buisson, L.; Josse, E.; Lebourges-Dhaussy, A.; Roudaut, G.; Ménard, F. Mesoscale Features and Micronekton in the Mozambique Channel: An Acoustic Approach. Deep Sea Res. Part II Top. Stud. Oceanogr. 2014, 100, 164–173. [Google Scholar] [CrossRef]
- Peña, M.; Olivar, M.P.; Balbín, R.; López-Jurado, J.L.; Iglesias, M.; Miquel, J. Acoustic Detection of Mesopelagic Fishes in Scattering Layers of the Balearic Sea (Western Mediterranean). Can. J. Fish. Aquat. Sci. 2014, 71, 1186–1197. [Google Scholar] [CrossRef]
- Warren, J.D.; Smith, J.N. Density and Sound Speed of Two Gelatinous Zooplankton: Ctenophore (Mnemiopsis leidyi) and Lion’s Mane Jellyfish (Cyanea capillata). J. Acoust. Soc. Am. 2007, 122, 574–580. [Google Scholar] [CrossRef]
- Benoit-Bird, K.J.; Moline, M.A.; Waluk, C.M.; Robbins, I.C. Integrated Measurements of Acoustical and Optical Thin Layers I: Vertical Scales of Association. Ecol. Oceanogr. Thin Plankton Layers 2010, 30, 17–28. [Google Scholar] [CrossRef]
- Bok, T.-H.; Na, J.; Paeng, D.-G. Diel Variation in High-Frequency Acoustic Backscatter from Cochlodinium Polykrikoides. J. Acoust. Soc. Am. 2013, 134, EL140–EL146. [Google Scholar] [CrossRef]
- Churnside, J.H.; Marchbanks, R.D.; Lee, J.H.; Shaw, J.A.; Weidemann, A.; Donaghay, P.L. Airborne Lidar Detection and Characterization of Internal Waves in a Shallow Fjord. J. Appl. Remote Sens. 2012, 6, 063611. [Google Scholar] [CrossRef]
- Marshall, N.B. Bathypelagic Fishes as Sound Scatterers in the Ocean. J. Mar. Res. 1951, 10, 1–17. [Google Scholar]
- Benoit-Bird, K.J.; Moline, M.A.; Southall, B.L. Prey in Oceanic Sound Scattering Layers Organize to Get a Little Help from Their Friends: Schooling within Sound Scattering Layers. Limnol. Oceanogr. 2017, 62, 2788–2798. [Google Scholar] [CrossRef]
- Bianchi, D.; Stock, C.; Galbraith, E.D.; Sarmiento, J.L. Diel Vertical Migration: Ecological Controls and Impacts on the Biological Pump in a One-Dimensional Ocean Model. Glob. Biogeochem. Cycles 2013, 27, 478–491. [Google Scholar] [CrossRef]
- Hays, G.C.; Richardson, A.J.; Robinson, C. Climate Change and Marine Plankton. Trends Ecol. Evol. 2005, 20, 337–344. [Google Scholar] [CrossRef]
- Richardson, A.J.; Bakun, A.; Hays, G.C.; Gibbons, M.J. The Jellyfish Joyride: Causes, Consequences and Management Responses to a More Gelatinous Future. Trends Ecol. Evol. 2009, 24, 312–322. [Google Scholar] [CrossRef]
- Boersch-Supan, P.H.; Rogers, A.D.; Brierley, A.S. The Distribution of Pelagic Sound Scattering Layers across the Southwest Indian Ocean. Deep Sea Res. Part II Top. Stud. Oceanogr. 2017, 136, 108–121. [Google Scholar] [CrossRef]
- D’Elia, M.; Warren, J.D.; Rodriguez-Pinto, I.; Sutton, T.T.; Cook, A.; Boswell, K.M. Diel Variation in the Vertical Distribution of Deep-Water Scattering Layers in the Gulf of Mexico. Deep Sea Res. Part Oceanogr. Res. Pap. 2016, 115, 91–102. [Google Scholar] [CrossRef]
- Bertrand, A.; Grados, D.; Habasque, J.; Fablet, R.; Ballon, M.; Castillo, R.; Gutierrez, M.; Chaigneau, A.; Josse, E.; Roudaut, G.; et al. Routine Acoustic Data as New Tools for a 3D Vision of the Abiotic and Biotic Components of Marine Ecosystem and Their Interactions. In Proceedings of the 2013 IEEE/OES Acoustics in Underwater Geosciences Symposium, Rio de Janeiro, Brazil, 24–26 July 2013; pp. 1–3. [Google Scholar]
- Proud, R.; Cox, M.J.; Wotherspoon, S.; Brierley, A.S. A Method for Identifying Sound Scattering Layers and Extracting Key Characteristics. Methods Ecol. Evol. 2015, 6, 1190–1198. [Google Scholar] [CrossRef]
- Demer, D.A.; Andersen, L.N.; Bassett, C.; Berger, L.; Chu, D.; Condiotty, J.; Cutter, G.R., Jr.; Hutton, B.; Korneliussen, R.; Le Bouffant, N.; et al. 2016 USA–Norway EK80 Workshop Report: Evaluation of a Wideband Echosounder for Fisheries and Marine Ecosystem Science; ICES Cooperative Research Report 336; International Council for the Exploration of the Sea: Copenhagen, Denmark, 2017. [Google Scholar] [CrossRef]
- Urmy, S.S.; Horne, J.K.; Barbee, D.H. Measuring the Vertical Distributional Variability of Pelagic Fauna in Monterey Bay. ICES J. Mar. Sci. 2012, 69, 184–196. [Google Scholar] [CrossRef]
- Diogoul, N.; Brehmer, P.; Demarcq, H.; El Ayoubi, S.; Thiam, A.; Sarre, A.; Mouget, A.; Perrot, Y. On the Robustness of an Eastern Boundary Upwelling Ecosystem Exposed to Multiple Stressors. Sci. Rep. 2021, 11, 1908. [Google Scholar] [CrossRef] [PubMed]
- Berraho, A.; Somoue, L.; Hernández-León, S.; Valdés, L. Zooplankton in the Canary Current Large Marine Ecosystem. In Oceanographic and Biological Features in the Canary Current Large Marine Ecosystem; Valdés, L., Déniz-Gonzalez, I., Eds.; Paris IOC Technical Series; IOC-UNESCO: Paris, France, 2015; pp. 183–195. [Google Scholar]
- Sambe, B.; Tandstad, M.; Caramelo, A.M.; Brown, B.E. Variations in Productivity of the Canary Current Large Marine Ecosystem and Their Effects on Small Pelagic Fish Stocks. Environ. Dev. 2016, 17, 105–117. [Google Scholar] [CrossRef]
- Arístegui, J.; Barton, E.D.; Álvarez-Salgado, X.A.; Santos, A.M.P.; Figueiras, F.G.; Kifani, S.; Hernández-León, S.; Mason, E.; Machú, E.; Demarcq, H. Sub-Regional Ecosystem Variability in the Canary Current Upwelling. Prog. Oceanogr. 2009, 83, 33–48. [Google Scholar] [CrossRef]
- Wiafe, G.; Yaqub, H.B.; Mensah, M.A.; Frid, C.L.J. Impact of Climate Change on Long-Term Zooplankton Biomass in the Upwelling Region of the Gulf of Guinea. ICES J. Mar. Sci. 2008, 65, 318–324. [Google Scholar] [CrossRef]
- Binet, D.; Marchal, E. The Large Marine Ecosystem of Shelf Areas in the Gulf of Guinea: Long-Term Variability Induced by Climatic Changes. In Large Marine Ecosystems: Stocks, Mitigation and Sustainability; Shermon, K., Alexander, L.M., Gold, B.D., Eds.; Institut de Recherche pour le Développement: Paris, France, 1993; pp. 104–118. [Google Scholar]
- Shannon, V.; Hempel, G.; Malanotte-Rizzoli, P.; Moloney, C.; Woods, J. (Eds.) Benguela: Predicting a Large Marine Ecosystem; Elsevier Science: Amsterdam, The Netherlands, 2006; Volume 14, ISBN 978-0-444-52759-2. [Google Scholar]
- Verheye, H.M.; Lamont, T.; Huggett, J.A.; Kreiner, A.; Hampton, I. Plankton Productivity of the Benguela Current Large Marine Ecosystem (BCLME). Environ. Dev. 2016, 17, 75–92. [Google Scholar] [CrossRef]
- Hagen, E.; Feistel, R.; Agenbag, J.J.; Ohde, T. Seasonal and Interannual Changes in Intense Benguela Upwelling (1982–1999). Oceanol. Acta 2001, 24, 557–568. [Google Scholar] [CrossRef]
- Mason, E.; Colas, F.; Molemaker, J.; Shchepetkin, A.F.; Troupin, C.; McWilliams, J.C.; Sangrà, P. Seasonal Variability of the Canary Current: A Numerical Study. J. Geophys. Res. Oceans 2011, 116. [Google Scholar] [CrossRef]
- Brierley, A.S. Diel Vertical Migration. Curr. Biol. 2014, 24, R1074–R1076. [Google Scholar] [CrossRef] [PubMed]
- Fock, H.O.; Matthiessen, B.; Zidowitz, H.; Westernhagen, H.v. Diel and Habitat-Dependent Resource Utilisation by Deep-Sea Fishes at the Great Meteor Seamount: Niche Overlap and Support for the Sound Scattering Layer Interception Hypothesis. Mar. Ecol. Prog. Ser. 2002, 244, 219–233. [Google Scholar] [CrossRef]
- Krakstad, J.-O.; Olsen, M.; Sarré, A.; Mbye, E.M. Survey of the Pelagic Fish Resources off North West Africa; Institute of Marine Research: Sénégal, The Gambia, 2006; p. 54. [Google Scholar]
- Foote, K.G. Calibration of Acoustic Instruments for Fish Density Estimation: A Practical Guide; International Council for the Exploration of the Sea: Copenhagen, Denmark, 1987. [Google Scholar]
- Bertrand, A.; Borgne, R.L.; Josse, E. Acoustic Characterisation of Micronekton Distribution in French Polynesia. Mar. Ecol. Prog. Ser. 1999, 191, 127–140. [Google Scholar] [CrossRef]
- Kloser, R.J.; Ryan, T.E.; Young, J.W.; Lewis, M.E. Acoustic Observations of Micronekton Fish on the Scale of an Ocean Basin: Potential and Challenges. ICES J. Mar. Sci. 2009, 66, 998–1006. [Google Scholar] [CrossRef]
- McClatchie, S.; Dunford, A. Estimated Biomass of Vertically Migrating Mesopelagic Fish Off New Zealand. Deep Sea Res. Part I Oceanogr. Res. Pap. 2003, 50, 1263–1281. [Google Scholar] [CrossRef]
- Cade, D.E.; Benoit-Bird, K.J. An Automatic and Quantitative Approach to the Detection and Tracking of Acoustic Scattering Layers. Limnol. Oceanogr. Methods 2014, 12, 742–756. [Google Scholar] [CrossRef]
- Burd, B.J.; Thomson, R.E.; Jamieson, G.S. Composition of a Deep Scattering Layer Overlying a Mid-Ocean Ridge Hydrothermal Plume. Mar. Biol. 1992, 113, 517–526. [Google Scholar] [CrossRef]
- Opdal, A.F.; Godø, O.R.; Bergstad, O.A.; Fiksen, Ø. Distribution, Identity, and Possible Processes Sustaining Meso- and Bathypelagic Scattering Layers on the Northern Mid-Atlantic Ridge. Deep Sea Res. Part II Top. Stud. Oceanogr. 2008, 55, 45–58. [Google Scholar] [CrossRef]
- Sherman, K. The Large Marine Ecosystem Concept: Research and Management Strategy for Living Marine Resources. Ecol. Appl. 1991, 1, 349–360. [Google Scholar] [CrossRef]
- Sarré, A.; Demarcq, H.; Faye, S.; Brehmer, P.; Krakstad, J.-O.; Thiao, D.; Elayoubi, S. Climate-Driven Shift of Sardinella Aurita Stock in Northwest Africa Ecosystem as Evidenced by Robust Spatial Indicators. Presented at the ICAWA: International Conference AWA, Dakar, Senegal, 13–15 December 2016. [Google Scholar]
- Benazzouz, A.; Mordane, S.; Orbi, A.; Chagdali, M.; Hilmi, K.; Atillah, A.; Lluís Pelegrí, J.; Hervé, D. An Improved Coastal Upwelling Index from Sea Surface Temperature Using Satellite-Based Approach—The Case of the Canary Current Upwelling System. Cont. Shelf Res. 2014, 81, 38–54. [Google Scholar] [CrossRef]
- Capet, X.; Estrade, P.; Machu, E.; Ndoye, S.; Grelet, J.; Lazar, A.; Marié, L.; Dausse, D.; Brehmer, P. On the Dynamics of the Southern Senegal Upwelling Center: Observed Variability from Synoptic to Superinertial Scales. J. Phys. Oceanogr. 2017, 47, 155–180. [Google Scholar] [CrossRef]
- Houghton, R.W. Seasonal Variations of the Subsurface Thermal Structure in the Gulf of Guinea. J. Phys. Oceanogr. 1983, 13, 2070–2081. [Google Scholar] [CrossRef][Green Version]
- Ndoye, S.; Capet, X.; Estrade, P.; Sow, B.; Machu, E.; Brochier, T.; Döring, J.; Brehmer, P. Dynamics of a “Low-Enrichment High-Retention” Upwelling Center over the Southern Senegal Shelf. Geophys. Res. Lett. 2017, 44, 5034–5043. [Google Scholar] [CrossRef]
- Perrot, Y.; Brehmer, P.; Habasque, J.; Roudaut, G.; Behagle, N.; Sarré, A.; Lebourges-Dhaussy, A. Matecho: An Open-Source Tool for Processing Fisheries Acoustics Data. Acoust. Aust. 2018, 46, 241–248. [Google Scholar] [CrossRef]
- Béhagle, N.; Cotté, C.; Ryan, T.E.; Gauthier, O.; Roudaut, G.; Brehmer, P.; Josse, E.; Cherel, Y. Acoustic Micronektonic Distribution Is Structured by Macroscale Oceanographic Processes across 20–50° S Latitudes in the South-Western Indian Ocean. Deep Sea Res. Part I Oceanogr. Res. Pap. 2016, 110, 20–32. [Google Scholar] [CrossRef]
- Lehodey, P.; Conchon, A.; Senina, I.; Domokos, R.; Calmettes, B.; Jouanno, J.; Hernandez, O.; Kloser, R. Optimization of a Micronekton Model with Acoustic Data. ICES J. Mar. Sci. 2015, 72, 1399–1412. [Google Scholar] [CrossRef]
- Domokos, R. Environmental Effects on Forage and Longline Fishery Performance for Albacore (Thunnus alalunga) in the American Samoa Exclusive Economic Zone. Fish. Oceanogr. 2009, 18, 419–438. [Google Scholar] [CrossRef]
- Sabarros, P.S.; Ménard, F.; Lévénez, P.; Tew-Kai, P.; Ternon, J.F. Mesoscale Eddies Influence Distribution and Aggregation Patterns of Micronekton in the Mozambique Channel. Mar. Ecol. Prog. Ser. 2009, 395, 101–107. [Google Scholar] [CrossRef]
- Weill, A.; Scalabrin, C.; Diner, N. MOVIES-B: An Acoustic Detection Description Software. Application to Shoal Species’ Classification. Aquat. Living Resour. 1993, 6, 255–267. [Google Scholar] [CrossRef]
- Woillez, M.; Poulard, J.-C.; Rivoirard, J.; Petitgas, P.; Bez, N. Indices for Capturing Spatial Patterns and Their Evolution in Time, with Application to European Hake (Merluccius merluccius) in the Bay of Biscay. ICES J. Mar. Sci. 2007, 64, 537–550. [Google Scholar] [CrossRef]
- Bez, N.; Rivoirard, J. Transitive Geostatistics to Characterise Spatial Aggregations with Diffuse Limits: An Application on Mackerel Ichtyoplankton. Fish. Res. 2001, 50, 41–58. [Google Scholar] [CrossRef]
- Benoit-Bird, K.J.; Au, W.W.L. Prey Dynamics Affect Foraging by a Pelagic Predator (Stenella Longirostris) over a Range of Spatial and Temporal Scales. Behav. Ecol. Sociobiol. 2003, 53, 364–373. [Google Scholar] [CrossRef]
- Lawson, G.L.; Barange, M.; Fréon, P. Species Identification of Pelagic Fish Schools on the South African Continental Shelf Using Acoustic Descriptors and Ancillary Information. ICES J. Mar. Sci. 2001, 58, 275–287. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Sheather, S.J.; Jones, M.C. A Reliable Data-Based Bandwidth Selection Method for Kernel Density Estimation. J. R. Stat. Soc. Ser. B Methodol. 1991, 53, 683–690. [Google Scholar] [CrossRef]
- McHugh, M.L. The Chi-Square Test of Independence. Biochem. Med. 2013, 23, 143–149. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Hempel, G. The Canary Current: Studies of an Upwelling System. In Proceedings of the Rapports et Procès-Verbaux de Réunions, Las Palmas, Spain, 11 April 1978; Volume 180, p. 10. [Google Scholar]
- Wiafe, G.; Dovlo, E.; Agyekum, K. Comparative Productivity and Biomass Yields of the Guinea Current LME. Environ. Dev. 2016, 17, 93–104. [Google Scholar] [CrossRef]
- Hutchings, L.; van der Lingen, C.D.; Shannon, L.J.; Crawford, R.J.M.; Verheye, H.M.S.; Bartholomae, C.H.; van der Plas, A.K.; Louw, D.; Kreiner, A.; Ostrowski, M.; et al. The Benguela Current: An Ecosystem of Four Components. Prog. Oceanogr. 2009, 83, 15–32. [Google Scholar] [CrossRef]
- Baussant, T.; Ibanez, F.; Dallot, S.; Etienne, M. Diurnal Mesoscale Patterns of 50-Khz Scattering Layers across the Ligurian Sea Front (NW Mediterranean-Sea). Oceanol. Acta 1992, 15, 3–12. [Google Scholar]
- Marchal, E.; Gerlotto, F.; Stequert, B. On the Relationship between Scattering Layer, Thermal Structure and Tuna Abundance in the Eastern Atlantic Equatorial Current System. Oceanol. Acta 1993, 16, 261–272. [Google Scholar]
- Urmy, S.S.; Horne, J.K. Multi-Scale Responses of Scattering Layers to Environmental Variability in Monterey Bay, California. Deep Sea Res. Part I Oceanogr. Res. Pap. 2016, 113, 22–32. [Google Scholar] [CrossRef]
- Zabala, M.; Riera, T.; Gili, J.M.; Barange, M.; Lobo, A.; Peñuelas, J. Water Flow, Trophic Depletion, and Benthic Macrofauna Impoverishment in a Submarine Cave from the Western Mediterranean. Mar. Ecol. 1989, 10, 271–287. [Google Scholar] [CrossRef]
- Brehmer, P.; Gerlotto, F.; Laurent, C.; Cotel, P.; Achury, A.; Samb, B. Schooling Behaviour of Small Pelagic Fish: Phenotypic Expression of Independent Stimuli. Mar. Ecol. Prog. Ser. 2007, 334, 263–272. [Google Scholar] [CrossRef]
- Cochrane, K.L.; Augustyn, C.; Fairweather, T.; Japp, D.; Kilongo, K.; Iitembu, J.; Moroff, N.; Roux, J.-P.; Shannon, L.; Van Zyl, B.; et al. Benguela Current Large Marine Ecosystem—Governance and Management for an Ecosystem Approach to Fisheries in the Region. Coast. Manag. 2009, 37, 235–254. [Google Scholar] [CrossRef]
- Sakko, A.L. The Influence of the Benguela Upwelling System on Namibia’s Marine Biodiversity. Biodivers. Conserv. 1998, 7, 419–433. [Google Scholar] [CrossRef]
- Shannon, L.V.; Pillar, S.C. The Benguela Ecosystem. Part III. Plankton. In Oceanography and Marine Biology, 1st ed.; CRC Press: Boca Raton, FL, USA, 1986; Volume 24, pp. 65–170. [Google Scholar]
- Tiedemann, M.; Brehmer, P. Larval Fish Assemblages across an Upwelling Front: Indication for Active and Passive Retention. Estuar. Coast. Shelf Sci. 2017, 187, 118–133. [Google Scholar] [CrossRef]
- Dekshenieks, M.; Donaghay, P.; Sullivan, J.; Rines, J.; Osborn, T.; Twardowski, M. Temporal and Spatial Occurrence of Thin Phytoplankton Layers in Relation to Physical Processes. Mar. Ecol. Prog. Ser. 2001, 223, 61–71. [Google Scholar] [CrossRef]
- McManus, M.; Alldredge, A.; Barnard, A.; Boss, E.; Case, J.; Cowles, T.; Donaghay, P.; Eisner, L.; Gifford, D.; Greenlaw, C.; et al. Characteristics, Distribution and Persistence of Thin Layers over a 48 Hour Period. Mar. Ecol. Prog. Ser. 2003, 261, 1–19. [Google Scholar] [CrossRef]
- Demarcq, H. Trends in Primary Production, Sea Surface Temperature and Wind in Upwelling Systems (1998–2007). Prog. Oceanogr. 2009, 83, 376–385. [Google Scholar] [CrossRef]
- Bakun, A. Coastal Ocean Upwelling. Science 1990, 247, 198–201. [Google Scholar] [CrossRef]
- Barton, E.D.; Field, D.B.; Roy, C. Canary Current Upwelling: More or Less? Prog. Oceanogr. 2013, 116, 167–178. [Google Scholar] [CrossRef]
- McGregor, H.V.; Dima, M.; Fischer, H.W.; Mulitza, S. Rapid 20th-Century Increase in Coastal Upwelling off Northwest Africa. Science 2007, 315, 637–639. [Google Scholar] [CrossRef]
- Lamont, T.; García-Reyes, M.; Bograd, S.J.; van der Lingen, C.D.; Sydeman, W.J. Upwelling Indices for Comparative Ecosystem Studies: Variability in the Benguela Upwelling System. J. Mar. Syst. 2018, 188, 3–16. [Google Scholar] [CrossRef]
- Ariza, A.; Landeira, J.M.; Escánez, A.; Wienerroither, R.; Aguilar de Soto, N.; Røstad, A.; Kaartvedt, S.; Hernández-León, S. Vertical Distribution, Composition and Migratory Patterns of Acoustic Scattering Layers in the Canary Islands. J. Mar. Syst. 2016, 157, 82–91. [Google Scholar] [CrossRef]
- Sarmiento, J.L.; Gruber, N. Ocean Biogeochemical Dynamics; Princeton University Press: Princeton, NJ, USA, 2006; ISBN 978-0-691-01707-5. [Google Scholar]
- Klevjer, T.A.; Irigoien, X.; Røstad, A.; Fraile-Nuez, E.; Benítez-Barrios, V.M.; Kaartvedt, S. Large Scale Patterns in Vertical Distribution and Behaviour of Mesopelagic Scattering Layers. Sci. Rep. 2016, 6, 19873. [Google Scholar] [CrossRef]
- Brehmer, P.; Demarcq, H.; Béhagle, N.; Tiedemann, M.; Perrot, Y.; Mouget, A.; Migayrou, C.; Kouassi, A.M.; Uanivi, U.; El Ayoubi, S.; et al. Comparative Analyses of Spatiotemporal Macrozooplankton Distributions Along Three LMEs of the West Coast of Africa Related to Environmental Parameters; Horizon IRD: Brest, France, 2018; p. 148. [Google Scholar]
- Bertrand, A.; Grados, D.; Colas, F.; Bertrand, S.; Capet, X.; Chaigneau, A.; Vargas, G.; Mousseigne, A.; Fablet, R. Broad Impacts of Fine-Scale Dynamics on Seascape Structure from Zooplankton to Seabirds. Nat. Commun. 2014, 5, 5239. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LME | Geographic Position | Sampled Years | Transect (nmi) | Number of Analysed ESU |
|---|---|---|---|---|
| Canary Current | 34° N; 7° W to 12° N; 17° W | 1994–2006, 2011, 2015 | 96,788 | 588,459 |
| Canary Current North | 20.8° N; 7.2° W to 34.1° N; 17.7° W | 1994–2006, 2011, 2015 | 58,965 | 368,782 |
| Canary Current South | 12.2° N; 16.1° W to 20.8° N; 17.7° W | 1994–2006, 2011, 2015 | 37,822 | 219,677 |
| Guinea Current | 4° N; 8° W to 6° N; 3° E | 1999–2006 | 12,908 | 86,841 |
| Benguela Current | 17° S; 9° E to 31° S; 17° E | 1994–2001 | 39,368 | 1086 |
| Calculated Variable | Symbol | Unit | Formula | Reference(s) |
|---|---|---|---|---|
| Bottom depth at ESU j | Dj | m | N/A | - |
| Number of SSL at ESU j | Ŋj | - | N/A | [30,62,63] |
| Minimum depth of all SSLs at ESU j | đall,j | m | N/A | [62,64] |
| Maximum depth of the shallowest SSL at ESU j | Đ1,j | m | N/A | Adapted from [62,64] |
| Maximum depth of all SSLs at ESU j | Đall,j | m | Adapted from [62] | |
| Minimal elevation of the shallowest SSL at ESU j | Å1,j | m | Adapted from [62,65] | |
| Minimal elevation of all SSLs at ESU j | Åall,j | m | Adapted from [62,65] | |
| Width of shallowest SSL at ESU j | Ẇ1,j | m | [28,62] | |
| Sv of shallowest SSL at ESU j | Sv, 1,j | dB re 1 m−1 | Adapted from [6] | |
| Mean Sv of all SSLs at ESU j | Sv, all,j | dB re 1 m−1 | Adapted from [6] | |
| sA of shallowest SSL at ESU j | sA, 1,j | m2 nmi−2 | Adapted from [6] | |
| Mean sA of all SSLs at ESU j | sA, all,j | m2 nmi−2 | Adapted from [6] | |
| Proportion of the water column occupied by the shallowest SSL at ESU j | Ṗ1,j | % | Present study | |
| Proportion of the water column occupied by all SSLs at ESU j | Ṗall,j | % | Adapted from [30] | |
| Contribution of the shallowest SSL in the proportion of the water column occupied at ESU j | Ċ1,j | - | Present study |
| Sound-Scattering Layers Variables (SSL) | CCLME | GCLME | BCLME | |
|---|---|---|---|---|
| North | South | |||
| Minimum depth of all SSLs (đall) | R2 = 7.45 × 10−3 (p < 2.2 × 10−16) | R2 = 4.56 × 10−3 (p < 2.2 × 10−16) | NS | NS |
| Maximum depth of first SSL (Đ1) | R2 = 3.46 × 10−3 (p < 2.2 × 10−16) | NS | NS | NS |
| Width of first SSL (Ẇ1) | R2 = 1.35 × 10−2 (p < 2.2 × 10−16) | NS | NS | NS |
| Minimum elevation of first SSL (Å1) | NS | R2 = 7.19 × 10−3 (p < 2.2 × 10−16) | NS | NS |
| Minimum elevation of all SSLs (Åall) | NS | R2 = 5.11 × 10−3 (p < 2.2 × 10−16) | NS | NS |
| Mean volume backscattering strength of first SSL (Sv, 1) | NS | NS | R2 = 2.71 × 10−2 (p < 2.2 × 10−16) | NS |
| Mean volume backscattering strength of all SSLs (Sv, all) | R2 = 4.73 × 10−3 (p = 0.090) | NS | R2 = 2.76 × 10−2 (p < 2.2 × 10−16) | NS |
| Nautical area scattering strength of first SSL (sA, 1) | NS | NS | NS | NS |
| Mean nautical area scattering strength of all SSLs (sA, all) | NS | NS | NS | NS |
| Proportion of the water column occupied by first SSL (Ṗ1) | R2 = 4.60 × 10−2 (p < 2.2 × 10−16) | NS | R2 = 2.37 × 10−2 (p < 2.2 × 10−16) | R2 = 6.64 × 10−2 (p < 2.2 × 10−16) |
| Proportion of the water column occupied by all SSLs (Ṗall) | R2 = 4.45 × 10−2 (p < 2.2 × 10−16) | R2 = 4.22 × 10−3 (p = 0.084) | R2 = 1.81 × 10−2 (p < 2.2 × 10−16) | R2 = 7.18 × 10−2 (p < 2.2 × 10−16) |
| Contribution of first SSL in the proportion of the water column occupied by (Ċ1) | R2 = 8.43 × 10−3 (p < 2.2 × 10−16) | NS | R2 = 3.80 × 10−3 (p < 2.2 × 10−16) | NS |
| Number of SSLs (Ŋ) | R2 = 2.55 × 10−3 (p = 4.83 × 10−3) | R2 = 2.37 × 10−3 (p = 0.0242) | NS | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mouget, A.; Brehmer, P.; Perrot, Y.; Uanivi, U.; Diogoul, N.; El Ayoubi, S.; Jeyid, M.A.; Sarré, A.; Béhagle, N.; Kouassi, A.M.; et al. Applying Acoustic Scattering Layer Descriptors to Depict Mid-Trophic Pelagic Organisation: The Case of Atlantic African Large Marine Ecosystems Continental Shelf. Fishes 2022, 7, 86. https://doi.org/10.3390/fishes7020086
Mouget A, Brehmer P, Perrot Y, Uanivi U, Diogoul N, El Ayoubi S, Jeyid MA, Sarré A, Béhagle N, Kouassi AM, et al. Applying Acoustic Scattering Layer Descriptors to Depict Mid-Trophic Pelagic Organisation: The Case of Atlantic African Large Marine Ecosystems Continental Shelf. Fishes. 2022; 7(2):86. https://doi.org/10.3390/fishes7020086
Chicago/Turabian StyleMouget, Anne, Patrice Brehmer, Yannick Perrot, Uatjavi Uanivi, Ndague Diogoul, Salahedine El Ayoubi, Mohamed Ahmed Jeyid, Abdoulaye Sarré, Nolwenn Béhagle, Aka Marcel Kouassi, and et al. 2022. "Applying Acoustic Scattering Layer Descriptors to Depict Mid-Trophic Pelagic Organisation: The Case of Atlantic African Large Marine Ecosystems Continental Shelf" Fishes 7, no. 2: 86. https://doi.org/10.3390/fishes7020086
APA StyleMouget, A., Brehmer, P., Perrot, Y., Uanivi, U., Diogoul, N., El Ayoubi, S., Jeyid, M. A., Sarré, A., Béhagle, N., Kouassi, A. M., & Feunteun, E. (2022). Applying Acoustic Scattering Layer Descriptors to Depict Mid-Trophic Pelagic Organisation: The Case of Atlantic African Large Marine Ecosystems Continental Shelf. Fishes, 7(2), 86. https://doi.org/10.3390/fishes7020086