
A Simple Protocol for Isolating Hemolymph from Single Drosophila melanogaster Adult Flies

Abstract
:1. Introduction
2. Experimental Design
2.1. Materials
2.2. Equipment
2.3. Statistical Analysis
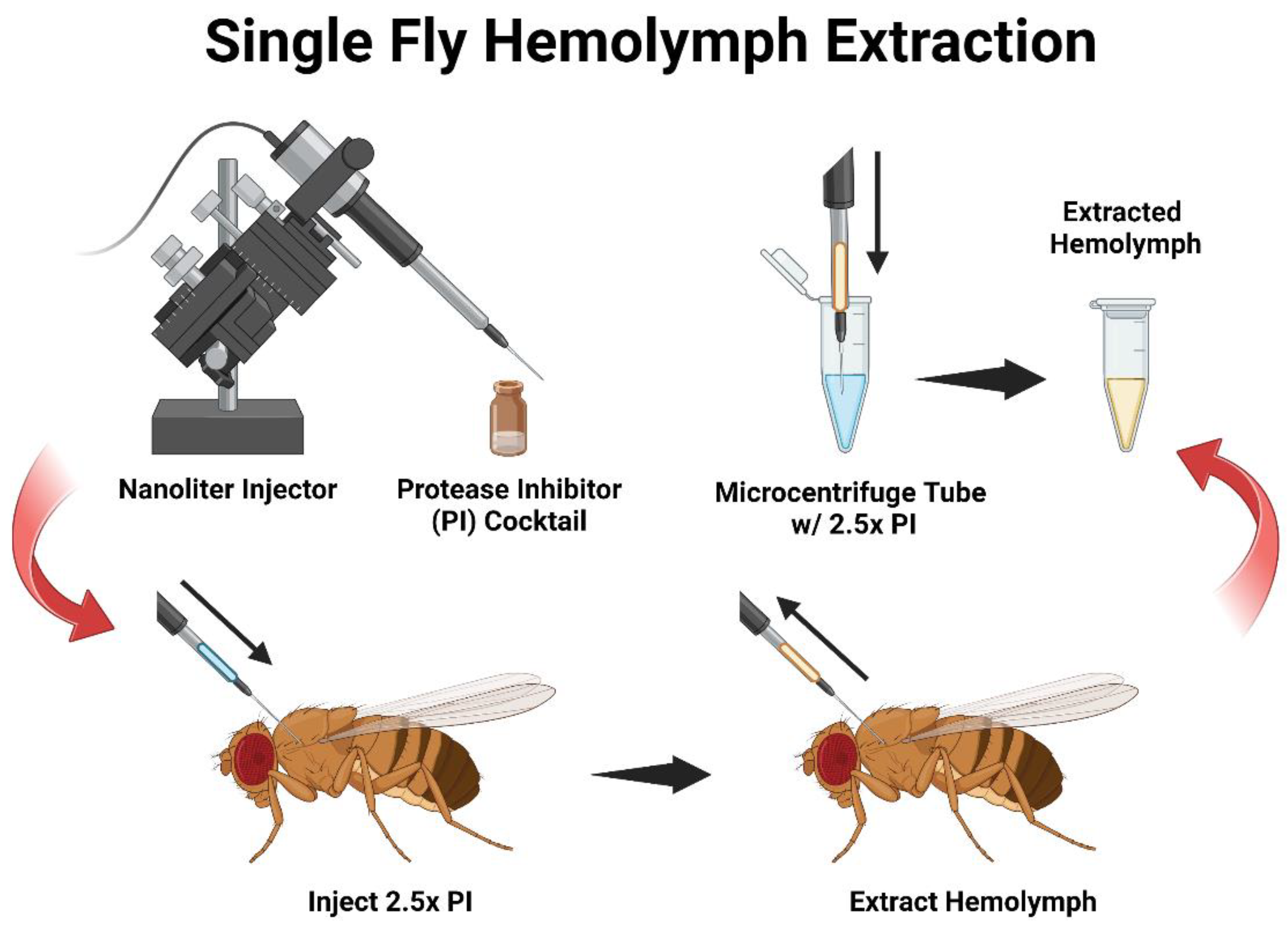
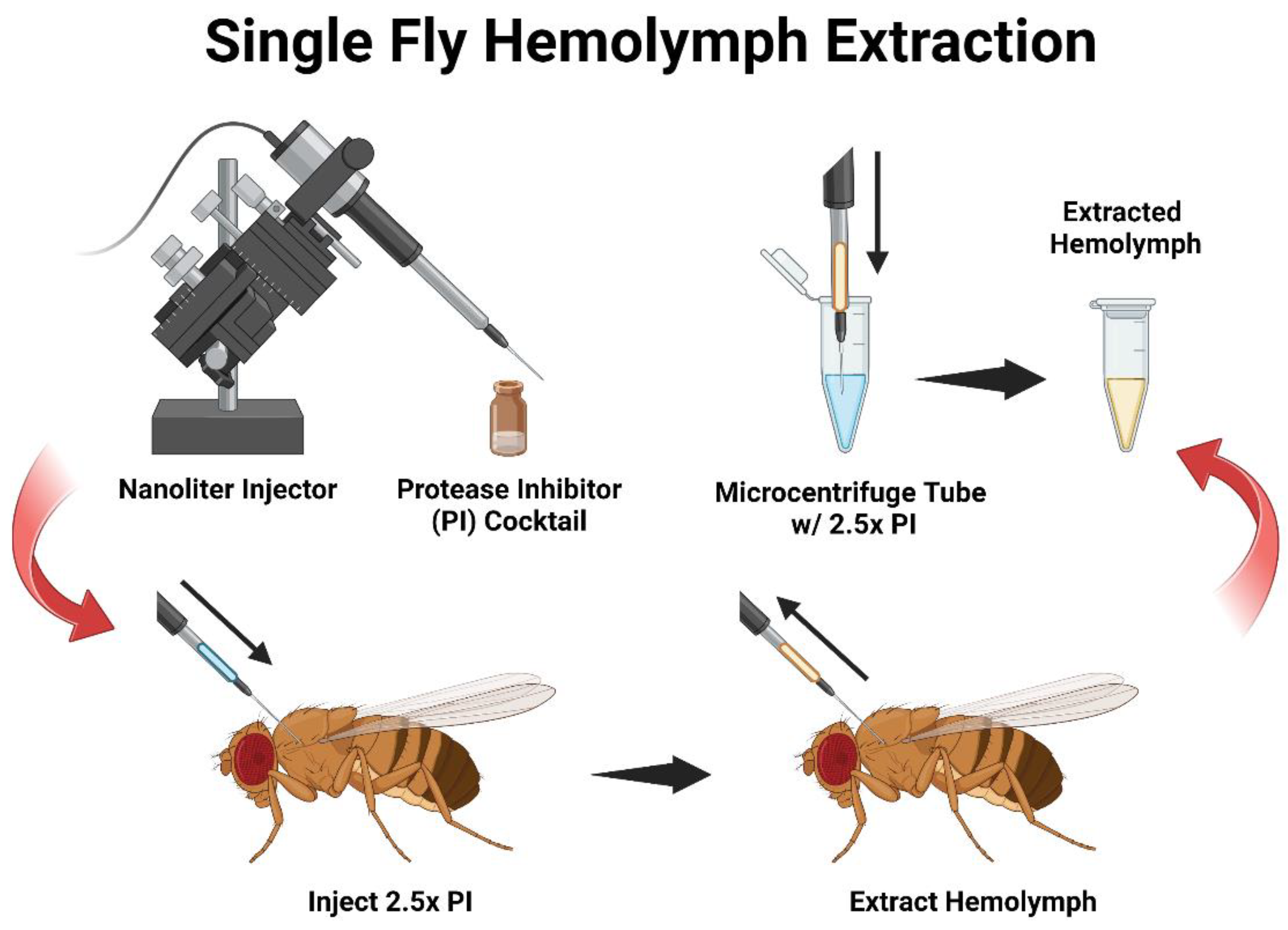
3. Procedure
- Prepare the nanoinjector per the manufacturer’s instructions and attach a glass capillary filled with mineral oil. Set the injector to “empty” to remove excess oil from the needle, and then fill the capillary needle with 5 μL of the diluted 2.5× protease inhibitor (PI) in 0.1 M of the sodium phosphate buffer (pH 7.4) per the manufacturer’s protocol. The flow rate is set at 69 nL/s and the fill rate at 30 nL/s.
- Group Collection: Anesthetize the flies under carbon dioxide and inject individuals with 100 nL of PI. After all individuals are injected, set the nanoinjector to the “fill” setting.
![Mps 06 00100 i001]() CRITICAL STEP TIP: Before withdrawing hemolymph from the flies, there should be PI left in the needle (approximately 10 nL) to come into contact with the extracted hemolymph and preserve it.
CRITICAL STEP TIP: Before withdrawing hemolymph from the flies, there should be PI left in the needle (approximately 10 nL) to come into contact with the extracted hemolymph and preserve it.- Reinsert the capillary into the mesothorax region of the adult fly and slightly pulse the nanoinjector for one to two seconds, creating a small vacuum allowing the hemolymph to freely draw upward into the needle.
- Once hemolymph is extracted from all individuals and the contents of the needle are emptied into a 1.5 mL microcentrifuge tube containing 20 μL of 2.5× PI on ice.
- OPTIONAL STEP: If extracting hemolymph from a large number of flies, periodically empty the collected hemolymph (approximately every 5–10 individuals) into the microcentrifuge tube on ice to preserve the hemolymph and repeat until all individuals are processed.
- Individual Collection: While flies are anesthetized under CO2, prepare the nanoinjector similarly to 3.1 but fill the capillary with 2.5 μL of PI instead of 5 μL. Inject a single individual with 100 nL of PI and then immediately reverse the flow of the nanoinjector to collect the hemolymph.
- Empty the contents of the needle into a 0.5 mL microcentrifuge tube containing 2.5 μL of 2.5× PI on ice.
- Repeat this process for all individuals in the trial.
- OPTIONAL STEP: You may change the needle in between individuals to avoid cross-contamination.
- Gently vortex the tube with extracted hemolymph and store it on ice until the hemocytes are ready for analysis.
- TROUBLESHOOTING: After creating the vacuum, you may need to slightly pull back on the needle in order for the hemolymph to flow backward into the needle.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
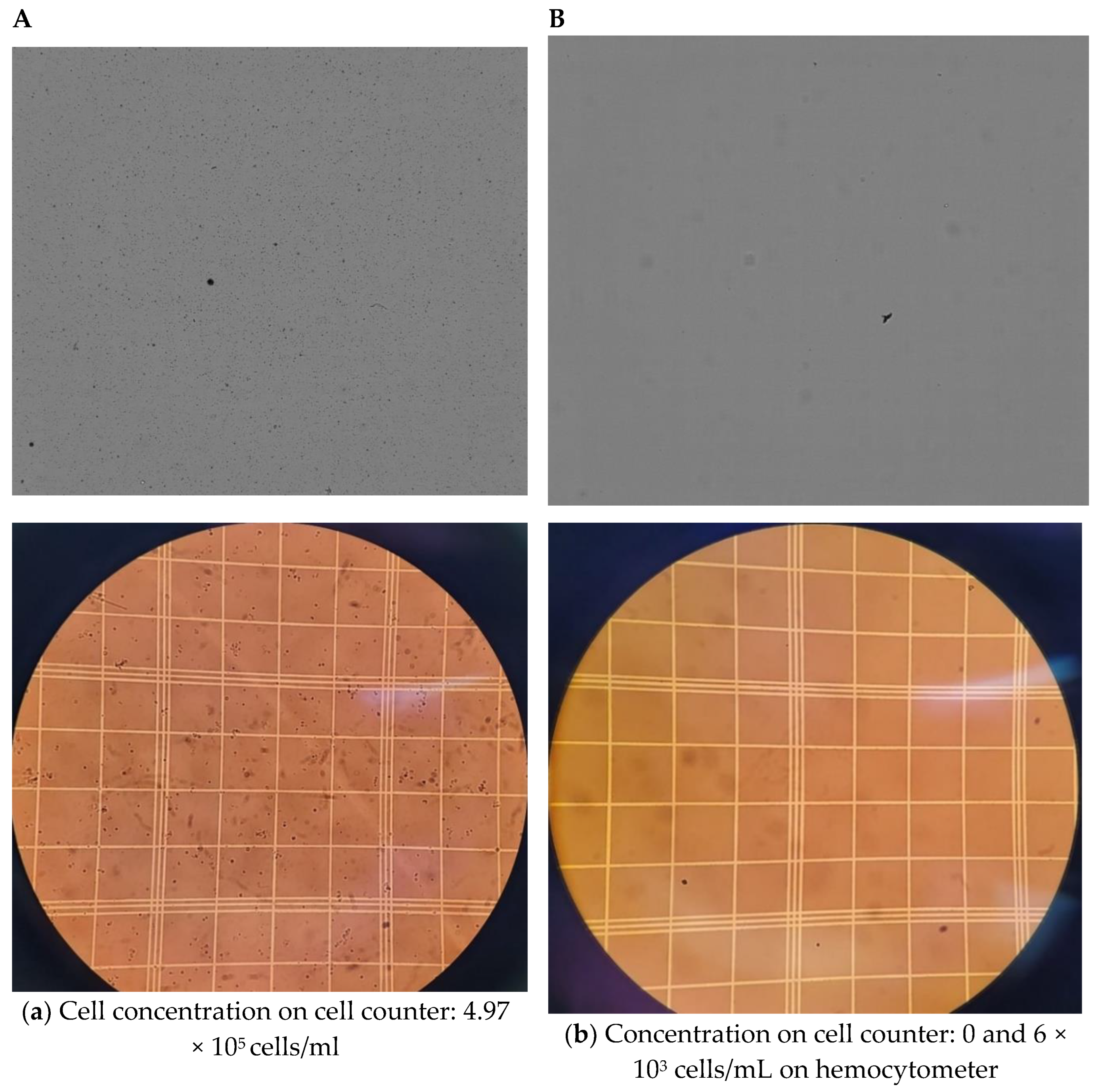
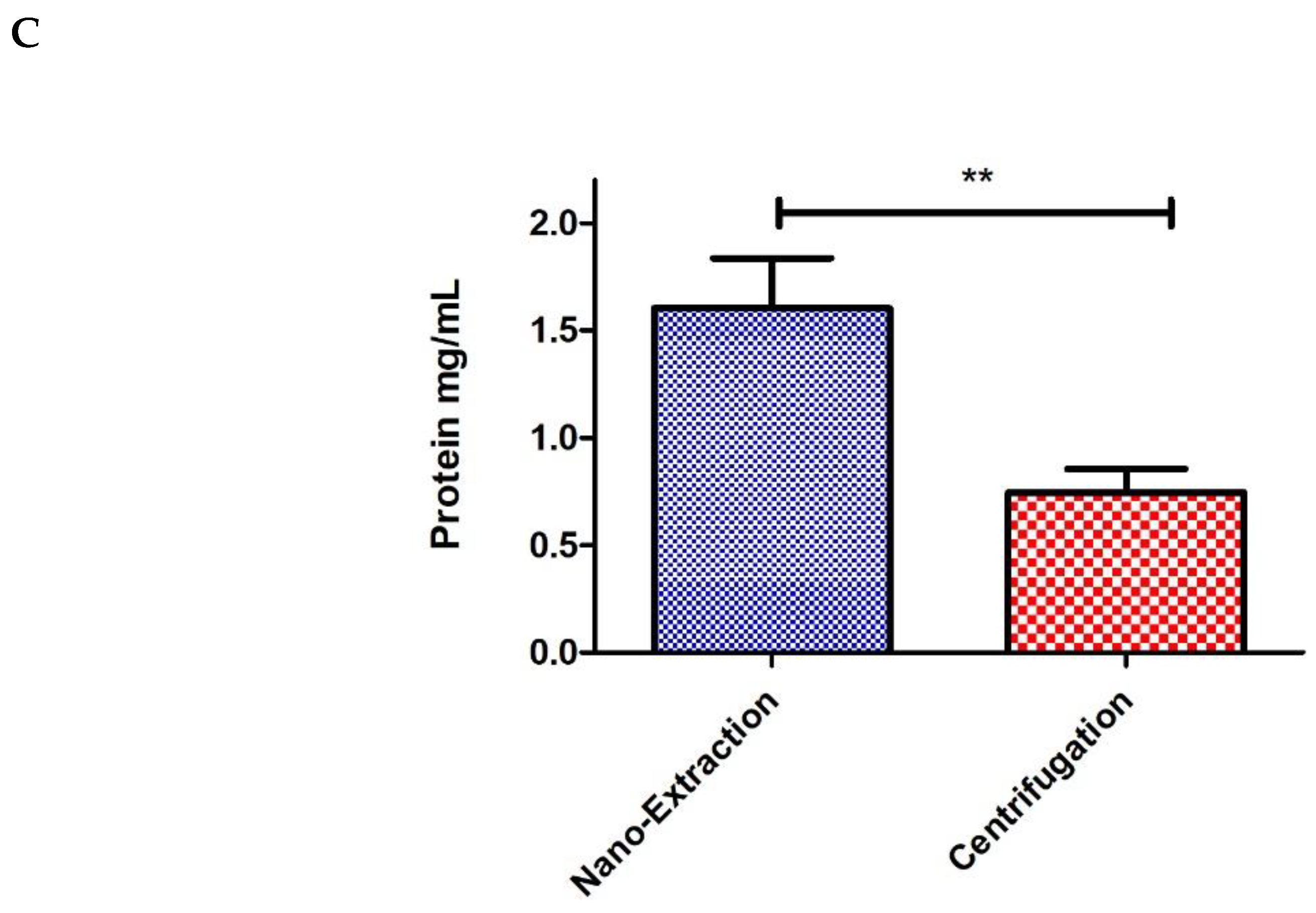
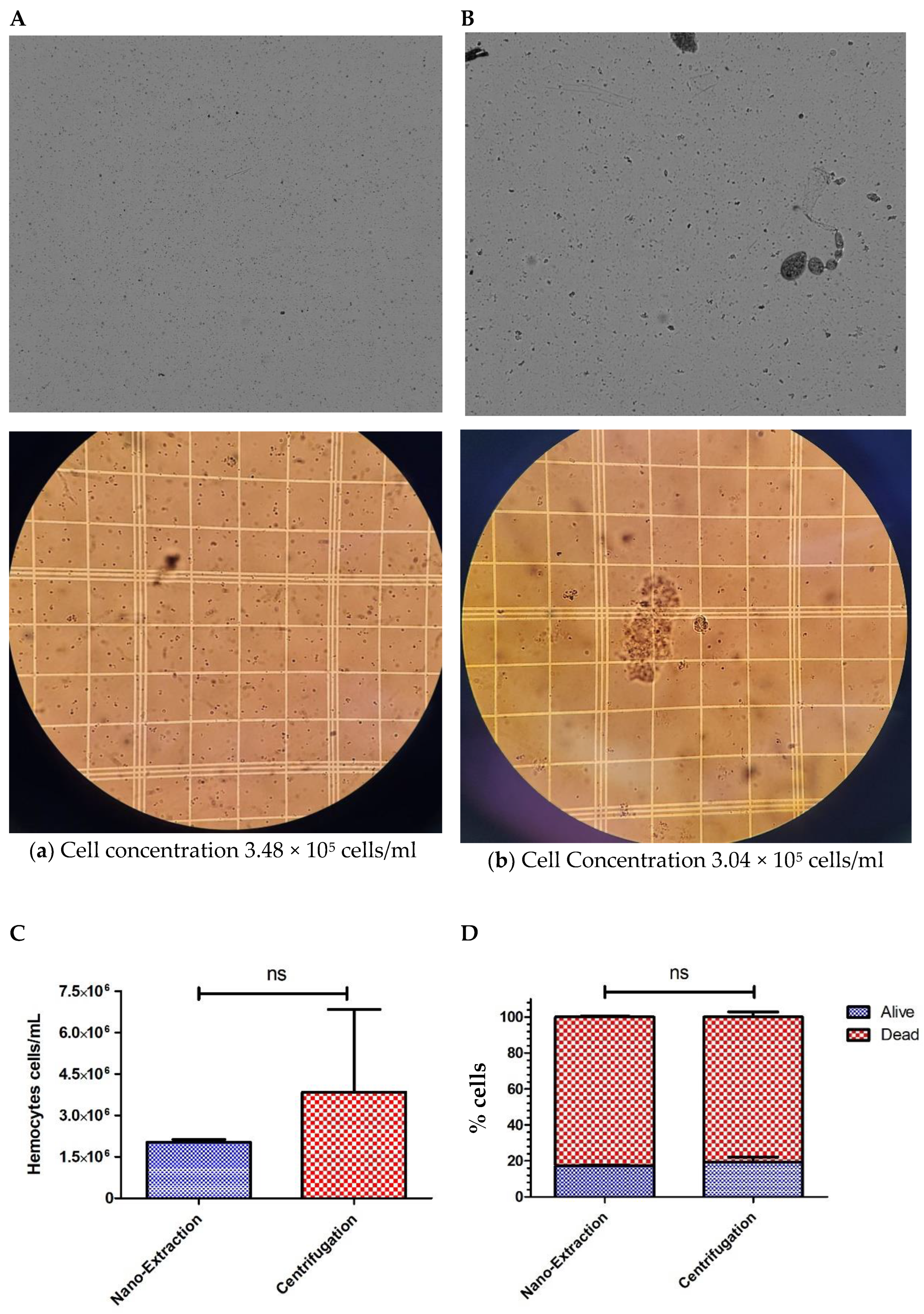
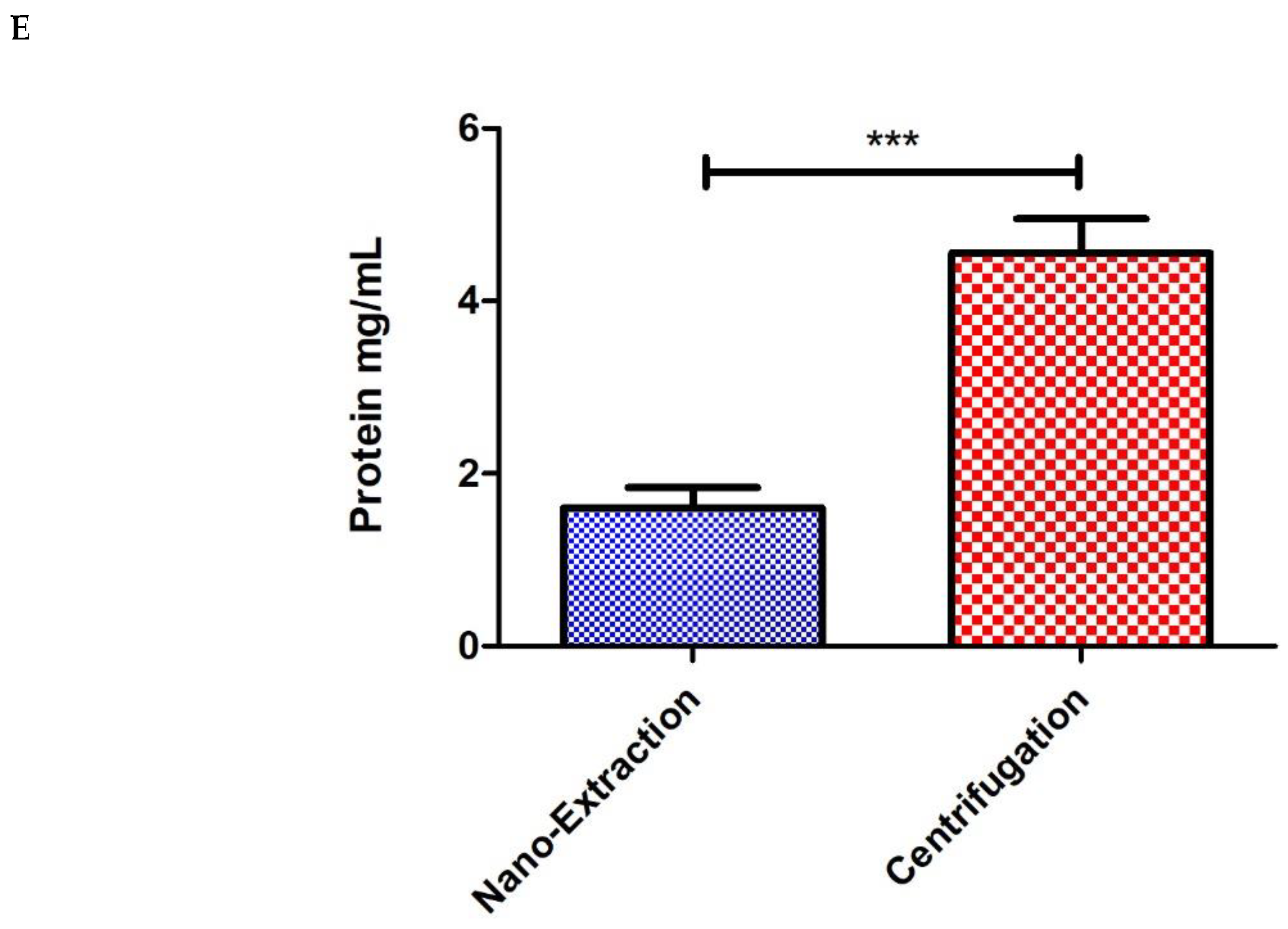
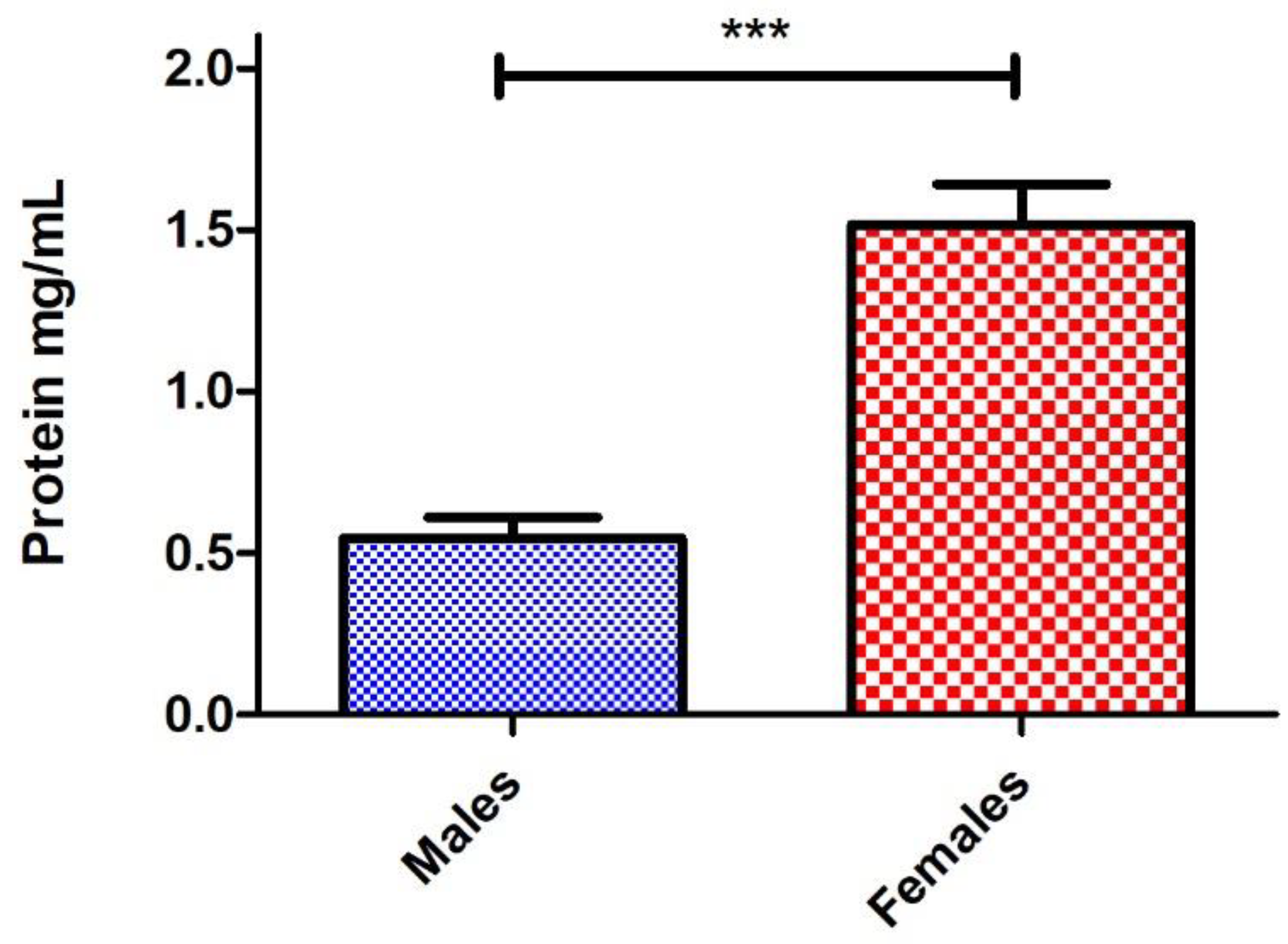
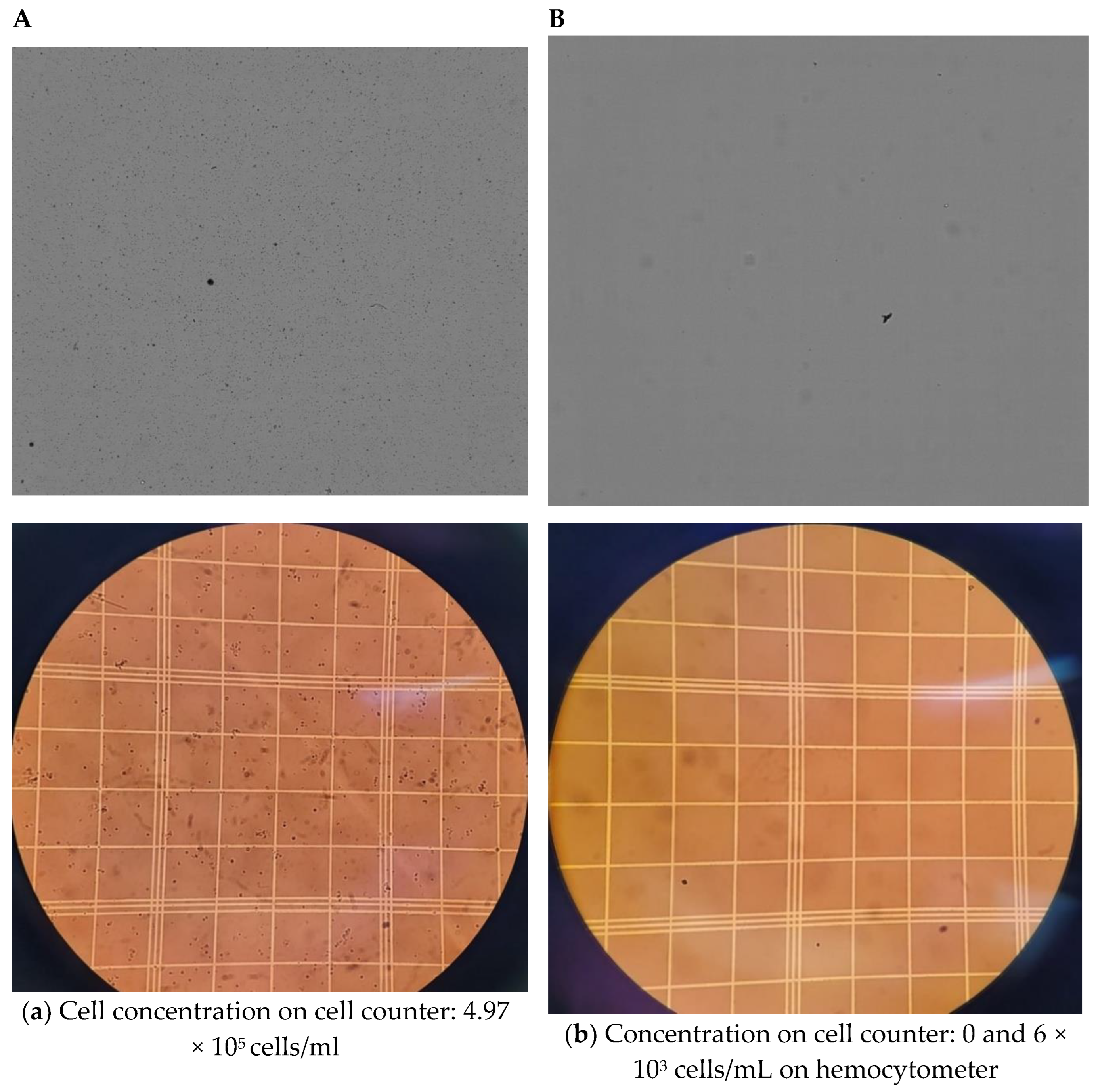
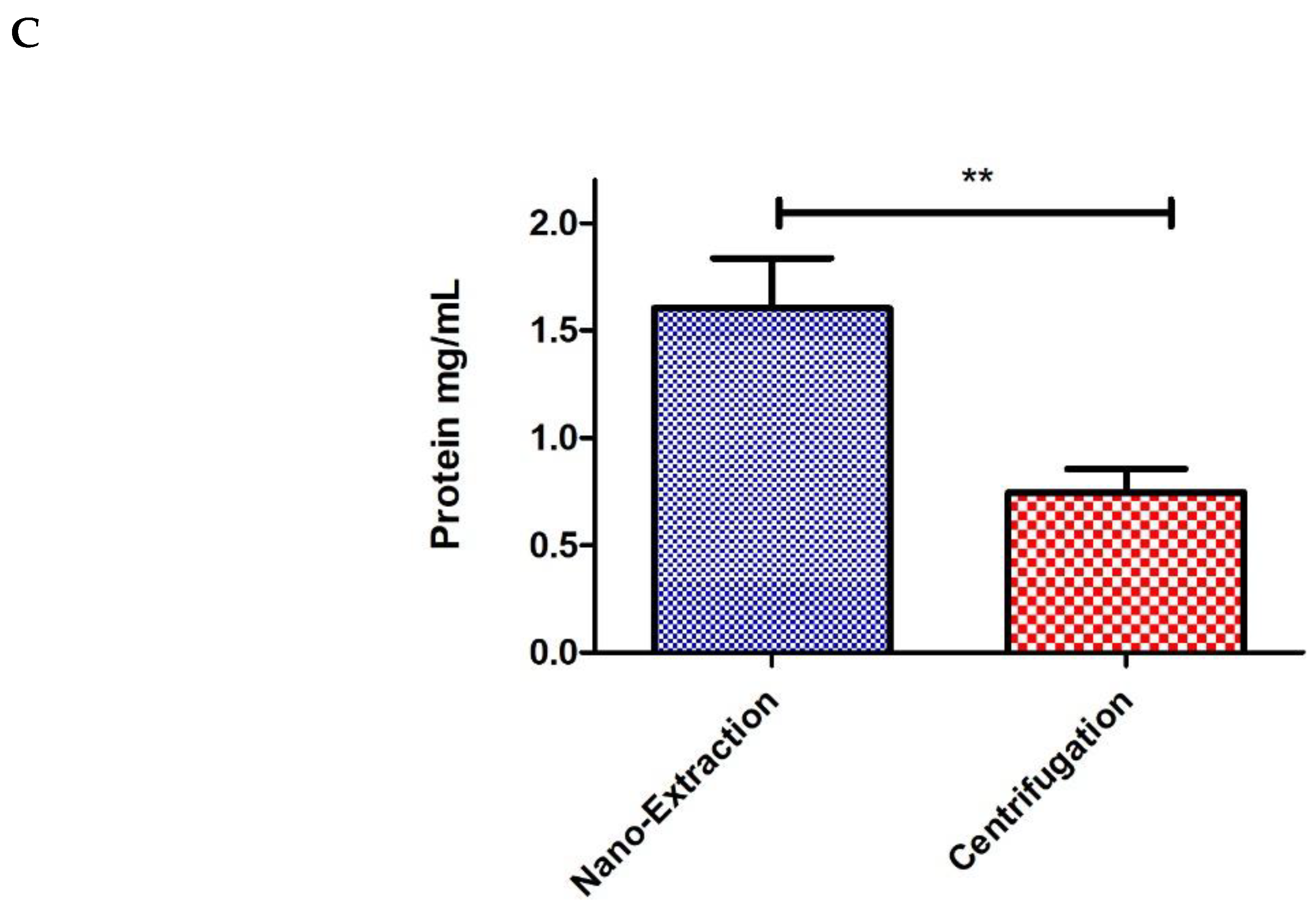
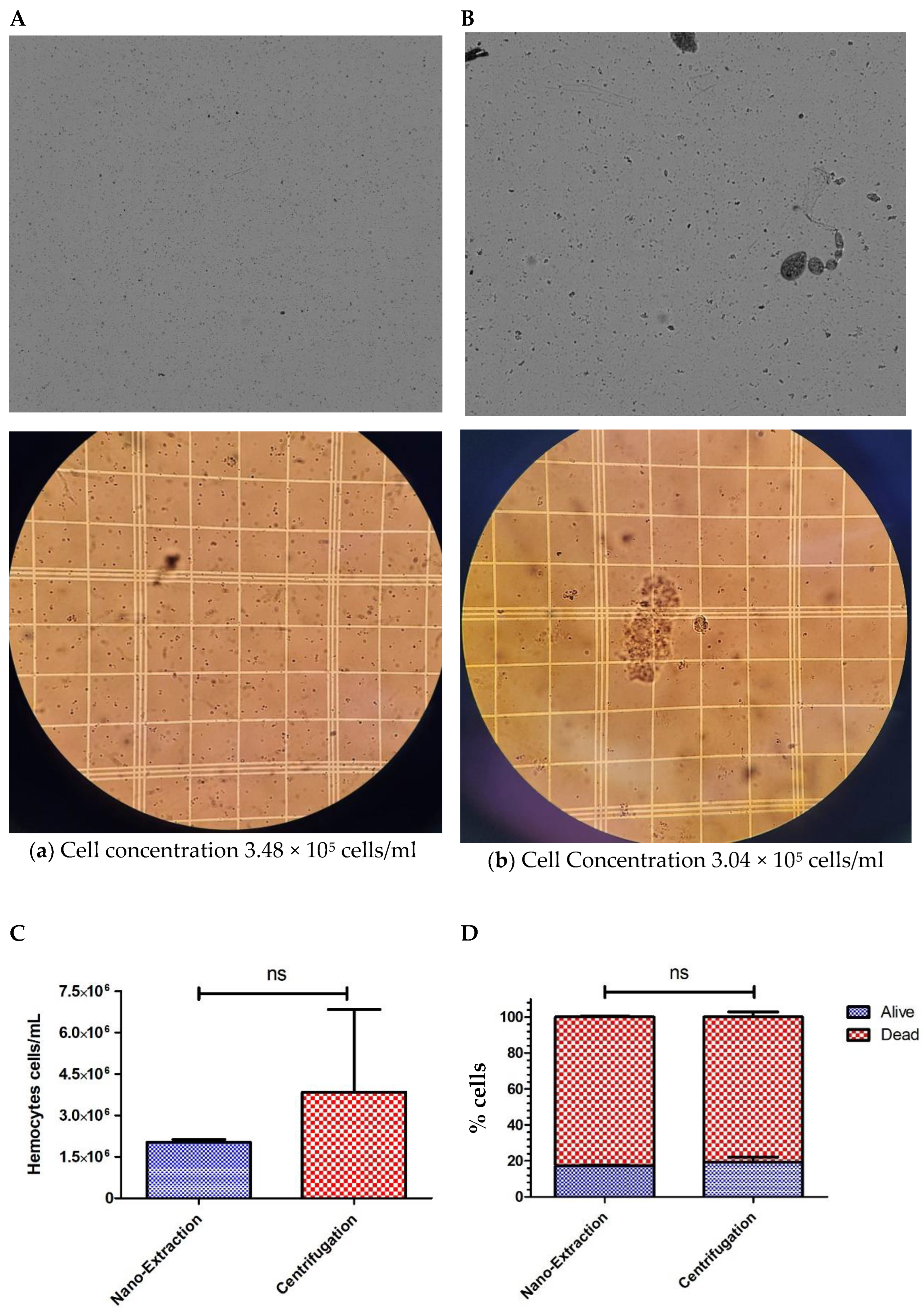
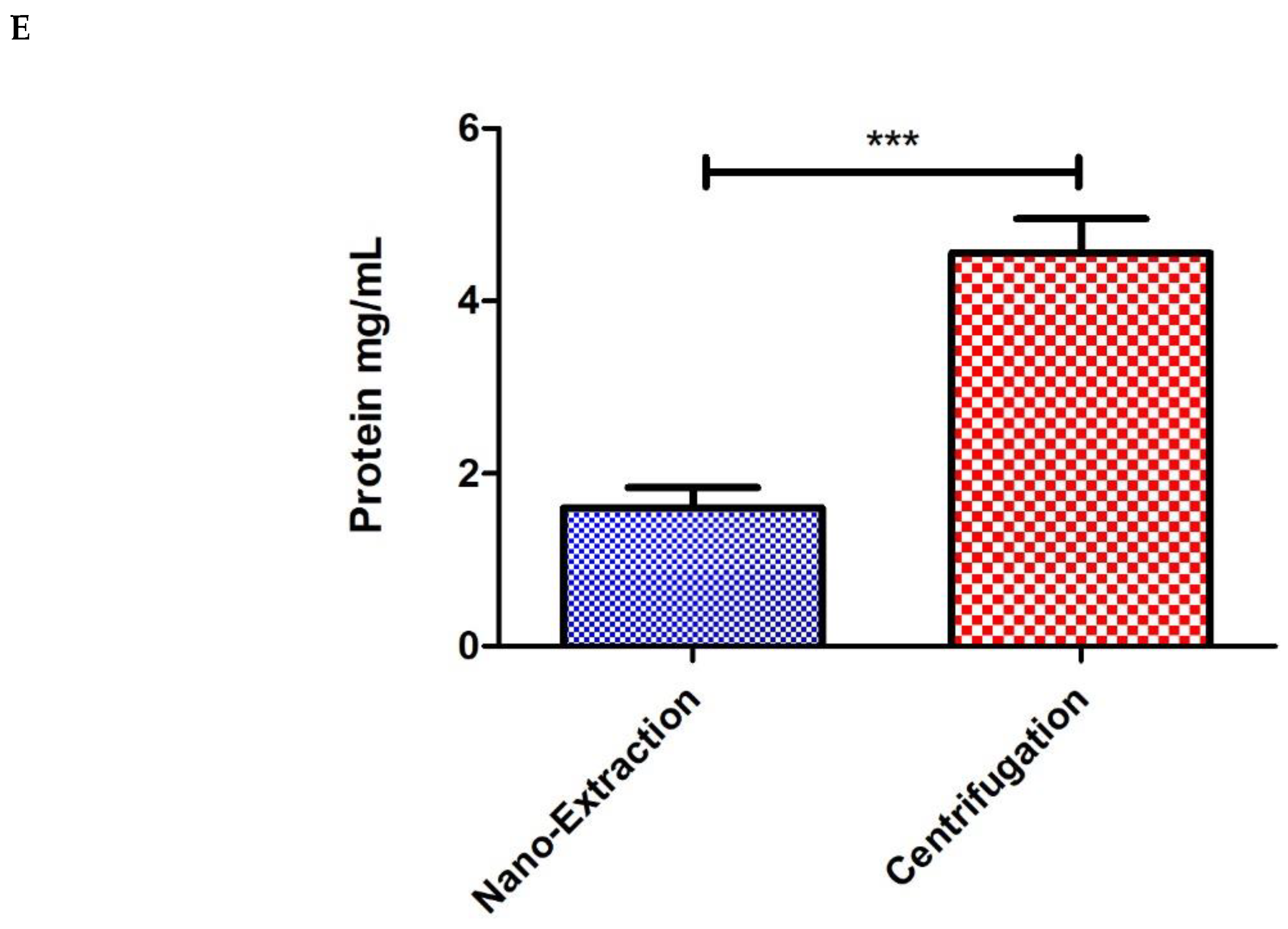
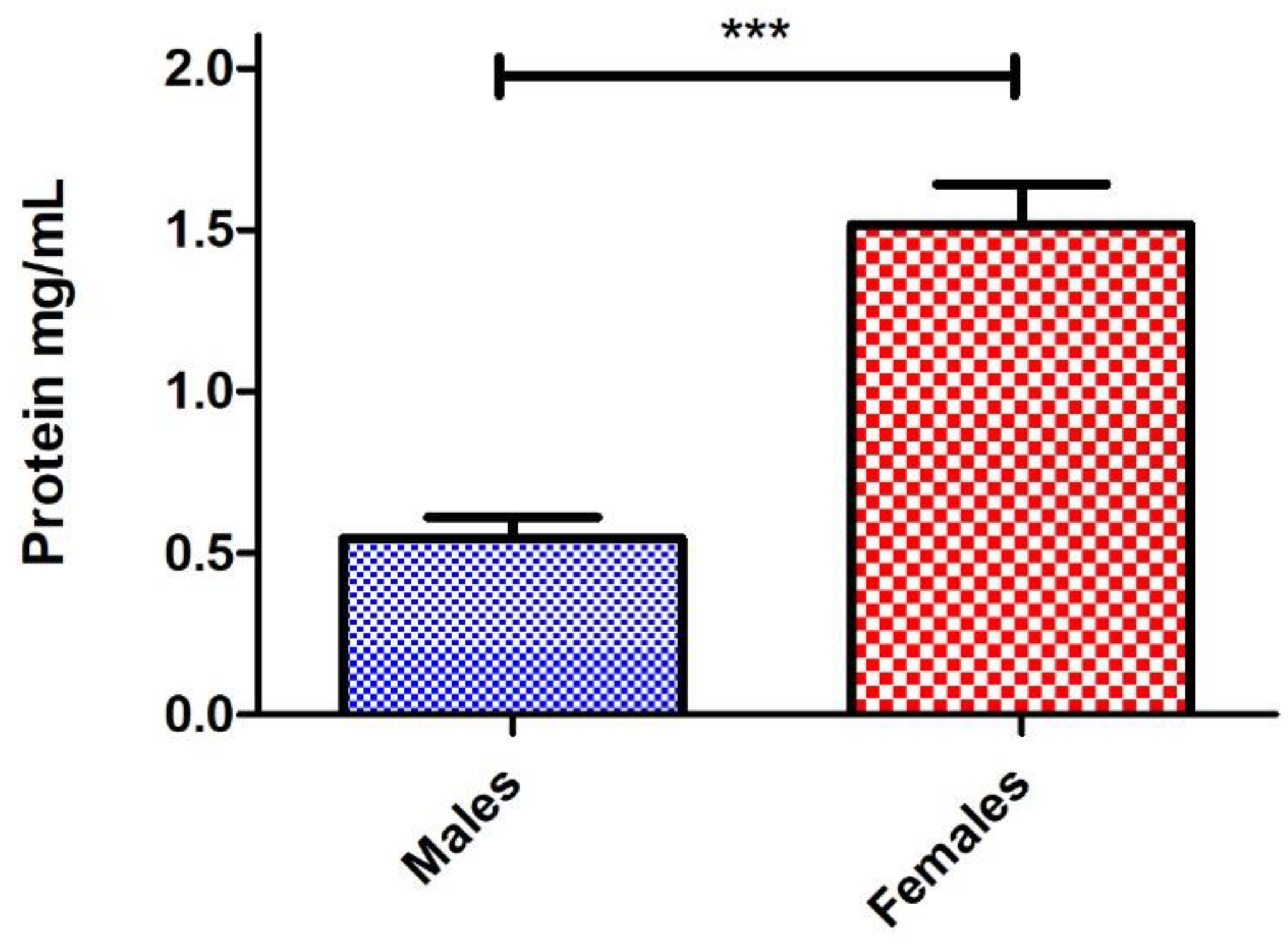
4. Results and Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adamski, Z.; Bufo, S.A.; Chowański, S.; Falabella, P.; Lubawy, J.; Marciniak, P.; Pacholska-Bogalska, J.; Salvia, R.; Scrano, L.; Slocińska, M.; et al. Beetles as Model Organisms in Physiological, Biomedical and Environmental Studies—A Review. Front. Physiol. 2019, 10, 319. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S. Using Drosophila as a model insect. Nat. Rev. Genet. 2000, 1, 218–226. [Google Scholar] [CrossRef]
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Luo, F.; Xu, Y.; Zhang, Y.; Jin, L.H. Drosophila Innate Immunity Involves Multiple Signaling Pathways and Coordinated Communication Between Different Tissues. Front. Immunol. 2022, 13, 905370. [Google Scholar] [CrossRef]
- Younes, S.; Al-Sulati, A.; Nasser, E.A.A.; Najjar, H.; Kamareddine, L. Drosophila as a Model Organism in Host-Pathogen Interaction Studies. Front. Cell Infect. Microbiol. 2020, 10, 214. [Google Scholar] [CrossRef]
- Kounatidis, I.; Ligoxygakis, P. Drosophila as a model system to unravel the layers of innate immunity to infection. Open Biol. 2012, 2, 120075. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.J.; Hartenstein, V.; Banerjee, U. Thicker than blood: Conserved mechanisms in Drosophila and vertebrate hematopoiesis. Dev. Cell 2003, 5, 673–690. [Google Scholar] [CrossRef] [PubMed]
- MacMillan, H.A.; Hughson, B.N. A high-throughput method of hemolymph extraction from adult Drosophila without anesthesia. J. Insect Physiol. 2014, 63, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Behrman, E.L.; Howick, V.M.; Kapun, M.; Staubach, F.; Bergland, A.O.; Petrov, D.A.; Lazzaro, B.P.; Schmidt, P.S. Rapid seasonal evolution in innate immunity of wild Drosophila melanogaster. Proc. Biol. Sci. 2018, 285, 20172599. [Google Scholar]
- Sackton, T.B.; Lazzaro, B.P.; Clark, A.G. Genotype and gene expression associations with immune function in Drosophila. PLoS Genet. 2010, 6, e1000797. [Google Scholar] [CrossRef]
- Neyen, C.; Bretscher, A.J.; Binggeli, O.; Lemaitre, B. Methods to study Drosophila immunity. Methods 2014, 68, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Mejia, M.; Heghinian, M.D.; Busch, A.; Marí, F.; Godenschwege, T.A. Paired nanoinjection and electrophysiology assay to screen for bioactivity of compounds using the Drosophila melanogaster giant fiber system. J. Vis. Exp. 2015, 62, 3597. [Google Scholar]
- Obata, F.; Tanak, S.; Kashio, S.; Tsujimura, H.; Sato, R.; Miura, M. Induction of rapid and selective cell necrosis in Drosophila using Bacillus thuringiensis Cry toxin and its silkworm receptor. BMC Biol. 2015, 13, 48. [Google Scholar] [CrossRef]
- Gong, S.; Zhang, Y.; Bao, H.; Wang, X.; Chang, C.H.; Huang, Y.C.; Deng, W.M. Tumor Allotransplantation in Drosophila melanogaster with a Programmable Auto-Nanoliter Injector. J. Vis. Exp. 2021, 168. [Google Scholar] [CrossRef]
- Tafesh-Edwards, G.; Kenney, E.; Eleftherianos, I. Drosophila melanogaster Larva Injection Protocol. J. Vis. Exp. 2021, 176, e63144. [Google Scholar] [CrossRef]
- Levy, F.; Bulet, P.; Ehret-Sabatier, L. Proteomic analysis of the systemic immune response of Drosophila. Mol. Cell Proteomics 2004, 3, 156–166. [Google Scholar] [CrossRef]
- Castillo, J.S.; Creasy, T.; Kumari, P.; Shetty, A.; Shokal, U.; Tallon, L.J.; Eleftherianos, I. Drosophila anti-nematode and antibacterial immune regulators revealed by RNA-Seq. BMC Genomics 2015, 16, 519. [Google Scholar] [CrossRef]
- Troha, K.; Buchon, N. Methods for the study of innate immunity in Drosophila melanogaster. Wiley Interdiscip. Rev. Dev. Biol. 2019, 8, e344. [Google Scholar] [CrossRef] [PubMed]
- Castillo, J.S.; Shokal, U.; Eleftherianos, I. A novel method for infecting Drosophila adult flies with insect pathogenic nematodes. Virulence 2012, 3, 339–347. [Google Scholar] [CrossRef]
- Stoepler, T.M.; Castillo, J.C.; Lill, J.T.; Eleftherianos, I. A simple protocol for extracting hemocytes from wild caterpillars. J. Vis. Exp. 2012, 69, e4173. [Google Scholar]
- Borsuk, G.; Ptaszyńska, A.A.; Olszewski, K.; Domaciuk, M.; Krutmuang, P.; Paleolog, J. A New Method for Quick and Easy Hemolymph Collection from Apidae Adults. PLoS ONE 2017, 12, e0170487. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, K.; Eleftherianos, I. A Simple Protocol for Isolating Hemolymph from Single Drosophila melanogaster Adult Flies. Methods Protoc. 2023, 6, 100. https://doi.org/10.3390/mps6050100
Jones K, Eleftherianos I. A Simple Protocol for Isolating Hemolymph from Single Drosophila melanogaster Adult Flies. Methods and Protocols. 2023; 6(5):100. https://doi.org/10.3390/mps6050100
Chicago/Turabian StyleJones, Kirah, and Ioannis Eleftherianos. 2023. "A Simple Protocol for Isolating Hemolymph from Single Drosophila melanogaster Adult Flies" Methods and Protocols 6, no. 5: 100. https://doi.org/10.3390/mps6050100
APA StyleJones, K., & Eleftherianos, I. (2023). A Simple Protocol for Isolating Hemolymph from Single Drosophila melanogaster Adult Flies. Methods and Protocols, 6(5), 100. https://doi.org/10.3390/mps6050100