Dynamics of Saccharomyces cerevisiae Strains Isolated from Vine Bark in Vineyard: Influence of Plant Age and Strain Presence during Grape must Spontaneous Fermentations

 and
and 
Abstract

1. Introduction
2. Results and Discussion
2.1. Presence of S. cerevisiae Strains on Grape Bunches and Vine Bark Portions at Harvest Time
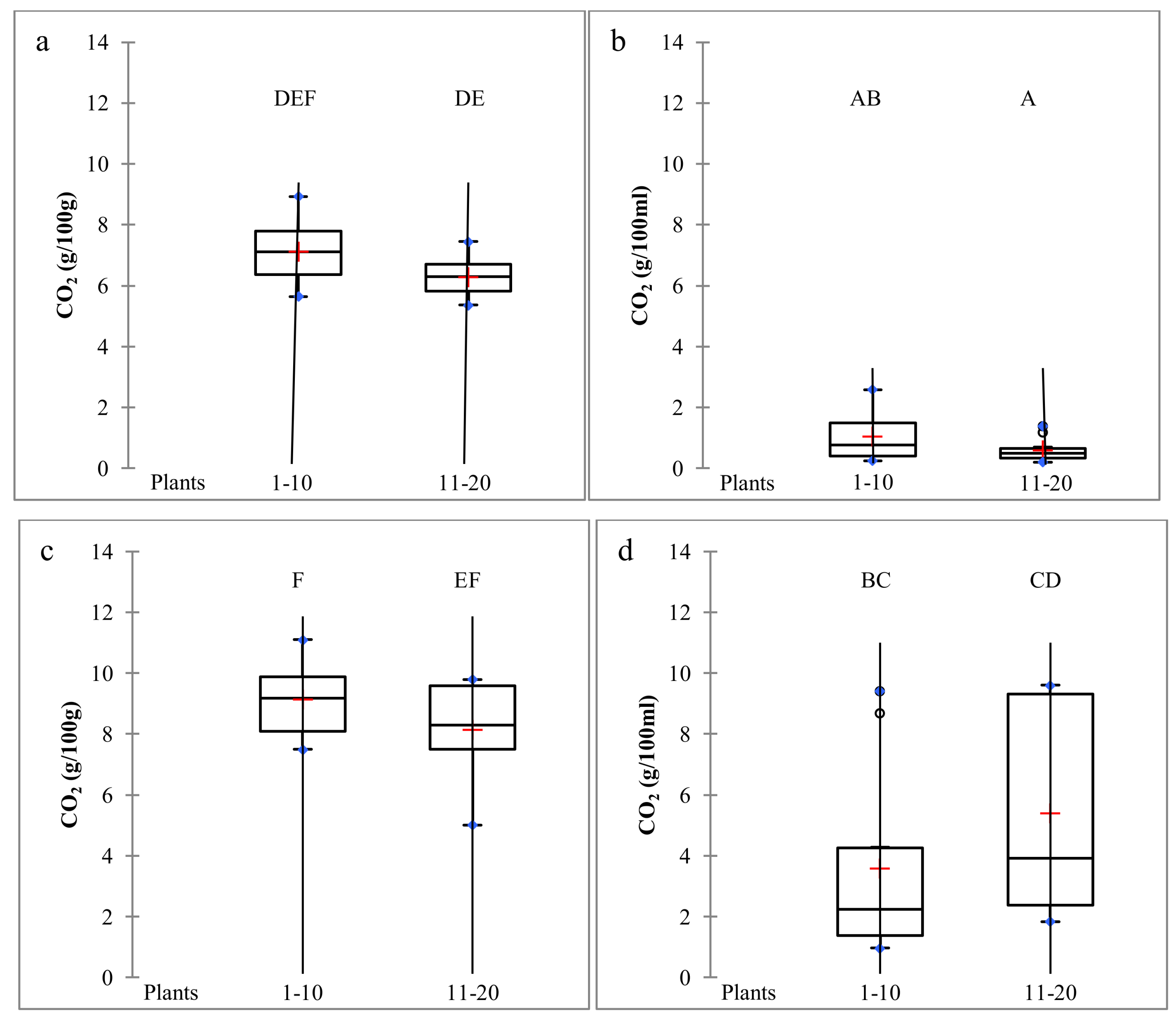
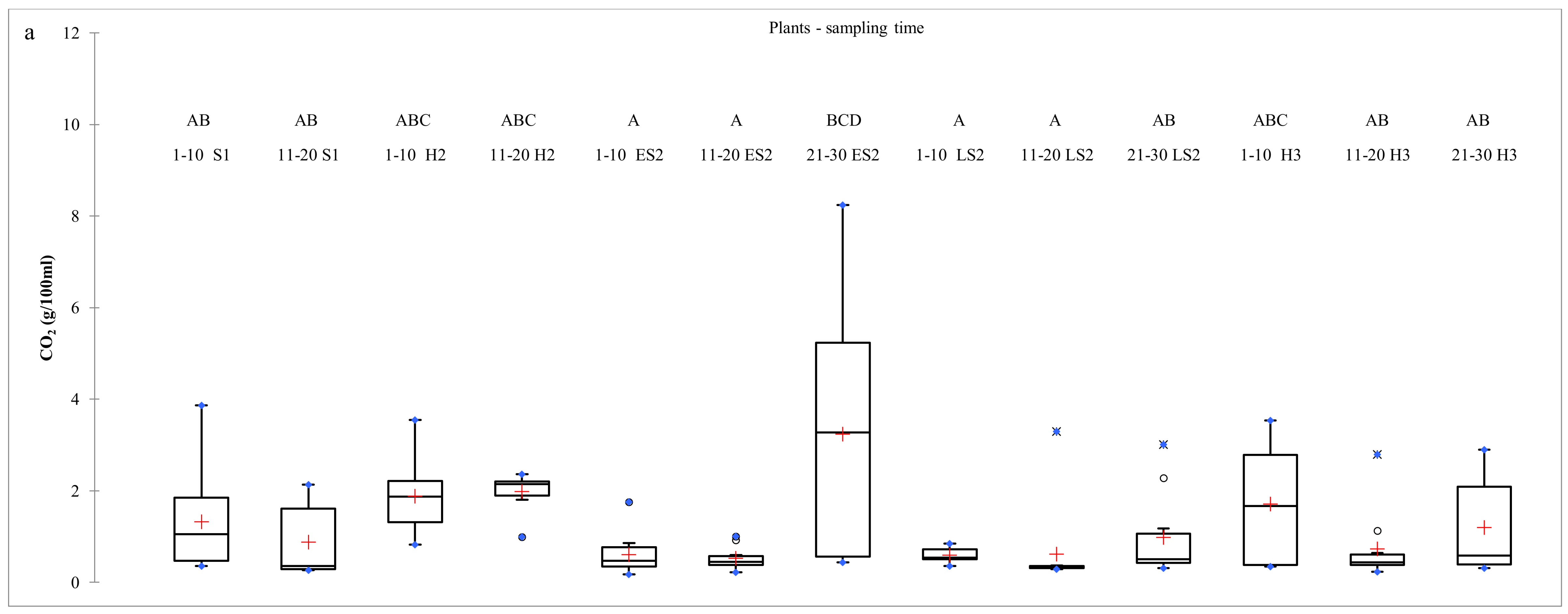
2.2. Dynamics of S. cerevisiae Strains on Vine Bark During Two Consecutive Vintages
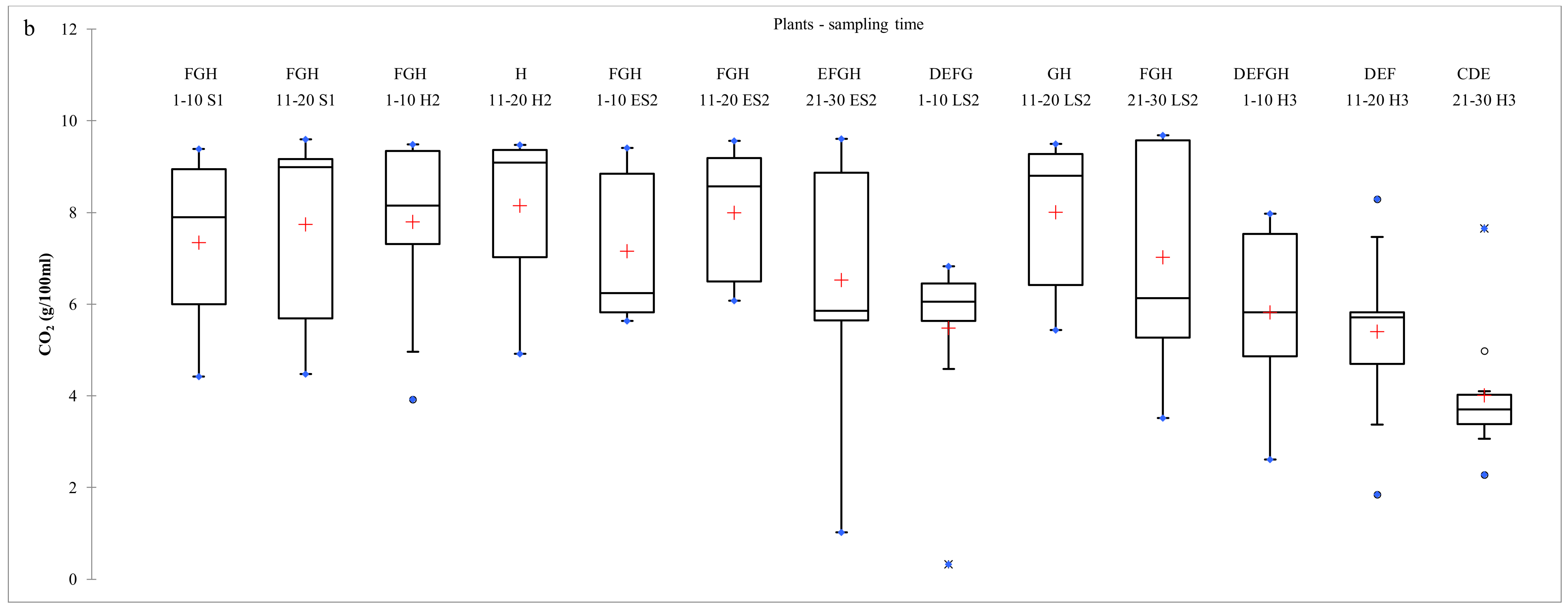
2.3. Dynamics of S. cerevisiae Strains During Pilot-Scale Vinifications
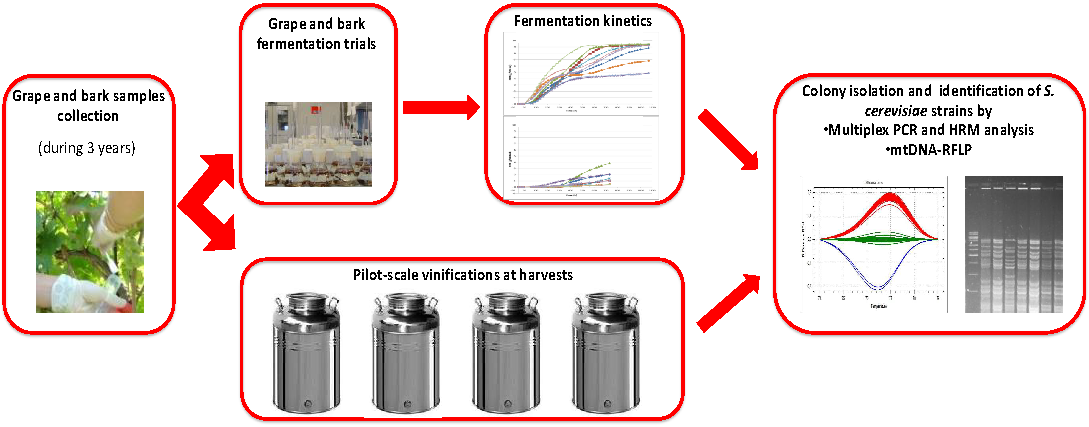
3. Materials and Methods
3.1. Wine-Making Area, Grapevine Variety, and Sampling Times
3.2. Bark and Grape Sampling and Yeast Isolation
3.3. Pilot-Scale Vinifications and Yeast Isolation
3.4. Species Identification
3.5. MtDNA RFLP Analysis
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mas, A.; Guillamón, J.M.; Beltran, G. Editorial: Non-conventional Yeast in the Wine Industry. Front. Microbiol. 2016, 7, 1494. [Google Scholar] [CrossRef] [PubMed]
- Clavijo, A.; Calderón, I.L.; Paneque, P. Diversity of Saccharomyces and non-Saccharomyces yeasts in three red grape varieties cultured in the Serrania de Ronda (Spain) vine-growing region. Int. J. Food Microbiol. 2010, 143, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Cordero-Bueso, G.; Arroyo, T.; Serrano, A.; Valero, E. Remanence and survival of commercial yeast in different ecological niches of the vineyard. FEMS Microbiol. Ecol. 2011, 77, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Crosato, G.; Carlot, M.; De Iseppi, A.; Garavaglia, J.; Pinto, L.M.N.; Ziegler, D.R.; de Souza Ramos, R.C.; Rossi, R.C.; Nadai, C.; Giacomini, A. Genetic variability and physiological traits of Saccharomyces cerevisiae strains isolated from “Vale dos Vinhedos” vineyards reflect agricultural practices and history of this Brazilian wet subtropical area. World J. Microbiol. Biotechnol. 2018, 34, 105. [Google Scholar] [CrossRef] [PubMed]
- Valero, E.; Schuller, D.; Cambon, B.; Casal, M.; Dequin, S. Biodiversity of Saccharomyces yeast strains from grape berries of wine-producing areas using starter commercial yeasts. FEMS Yeast Res. 2007, 7, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Versavaud, A.; Courcoux, P.; Roulland, C.; Dulau, L.; Hallet, J. Genetic diversity and geographical distribution of wild Saccharomyces cerevisiae strains from the wine-producing area of Charentes, France. Appl. Environ. Microbiol. 1995, 61, 3521–3529. [Google Scholar] [PubMed]
- Viel, A.; Legras, J.L.; Nadai, C.; Carlot, M.; Lombardi, A.; Crespan, M.; Migliaro, D.; Giacomini, A.; Corich, V. The geographic distribution of Saccharomyces cerevisiae isolates within three Italian neighboring winemaking regions reveals strong differences in yeast abundance, genetic diversity and industrial strain dissemination. Front. Microbiol. 2017, 8, 1595. [Google Scholar] [CrossRef] [PubMed]
- Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. The microbial ecology of wine grape berries. Int. J. Food Microbiol. 2012, 153, 243–259. [Google Scholar] [CrossRef]
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Martini, A.; Ciani, M.; Scorzetti, G. Direct enumeration and isolation of wine yeasts from grapes surfaces. Am. J. Enol. Vitic. 1996, 47, 435–440. [Google Scholar] [CrossRef]
- Taylor, M.W.; Tsai, P.; Anfang, N.; Ross, H.A.; Goddard, M.R. Pyrosequencing reveals regional differences in fruit-associated fungal communities. Environ. Microbiol. 2014, 16, 2848–2858. [Google Scholar] [CrossRef] [PubMed]
- Robiglio, A.; Sosa, M.C.; Lutz, M.C.; Lopes, C.A.; Sangorrín, M.P. Yeast biocontrol of fungal spoilage of pears stored at low temperature. Int. J. Food Microbiol. 2011, 147, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.M.; Liu, W.Q.; Liti, G.; Wang, S.A.; Bai, F.Y. Surprisingly diverged populations of Saccharomyces cerevisiae in natural environments remote from human activity. Mol. Ecol. 2012, 21, 5404–5417. [Google Scholar] [CrossRef] [PubMed]
- Stefanini, I.; Dapporto, L.; Legras, J.L.; Calabretta, A.; Di Paola, M.; De Filippo, C.; Viola, R.; Capretti, P.; Polsinelli, M.; Turillazzi, S. Role of social wasps in Saccharomyces cerevisiae ecology and evolution. Proc. Natl. Acad. Sci. USA 2012, 109, 13398–13403. [Google Scholar] [CrossRef] [PubMed]
- Naumov, G.I.; Naumova, E.S.; Sniegowski, P.D. Saccharomyces paradoxus and Saccharomyces cerevisiae are associated with exudates of North American oaks. Can. J. Microbiol. 1998, 44, 1045–1050. [Google Scholar] [CrossRef] [PubMed]
- Sniegowski, P.D.; Dombrowski, P.G.; Fingerman, E. Saccharomyces cerevisiae and Saccharomyces paradoxus coexist in a natural woodland site in North America and display different levels of reproductive isolation from European conspecifics. FEMS Yeast Res. 2002, 1, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.J.; Koufopanou, V.; Goddard, M.R.; Hetherington, R.; Schäfer, S.M.; Burt, A. Population genetics of the wild yeast Saccharomyces paradoxus. Genetics 2004, 166, 43–52. [Google Scholar] [CrossRef][Green Version]
- Sampaio, J.P.; Goncalves, P. Natural populations of Saccharomyces kudriavzevii in Portugal are associated with oak bark and are sympatric with S. cerevisiae and S. paradoxus. Appl. Environ. Microbiol. 2008, 74, 2144–2152. [Google Scholar] [CrossRef]
- Zhang, H.; Skelton, A.; Gardner, R.C.; Goddard, M.R. S. paradoxus and S. cerevisiae reside on oak trees in New Zealand: Evidence for migration from Europe and inter-species hybrids. FEMS Yeast Res. 2010, 7, 941–947. [Google Scholar] [CrossRef]
- Hyma, K.E.; Fay, J.C. Mixing of vineyard and oak-tree ecotypes of Saccharomyces cerevisiae in North American vineyards. Mol. Ecol. 2013, 22, 2917–2930. [Google Scholar] [CrossRef]
- Goddard, M.R.; Greig, D. Saccharomyces cerevisiae: A nomadic yeast with no niche? FEMS Yeast Res. 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Setati, M.E.; Jacobson, D.; Andong, U.C.; Bauer, F.F. The Vineyard Yeast Microbiome, a Mixed Model Microbial Map. PLoS ONE 2013, 7, e52609. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Thorngate, J.H.; Richardson, P.M.; Mills, D.A. Microbial biogeography of wine grapes is conditioned by cultivar, vintage, and climate. Proc. Natl. Acad. Sci. USA 2014, 111, E139–E148. [Google Scholar] [CrossRef] [PubMed]
- Mannazzu, I.M.; Clementi, F.; Ciani, M. Strategies and criteria for the isolation and selection of autochthonous starters. In Biodiversity and Biotechnology of Wine Yeasts; Ciani, M., Ed.; Research Signpost: Trivandrum, India, 2002; pp. 19–35. [Google Scholar]
- Pretorius, I.S. Tailoring wine yeast for the new millennium: Novel approaches to the ancient art of winemaking. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef]
- Goddard, M.R.; Anfang, N.; Tang, R.; Gardner, R.C.; Jun, C. A distinct population of Saccharomyces cerevisiae in New Zealand: Evidence for local dispersal by insects and human-aided global dispersal in oak barrels. Environ. Microbiol. 2010, 12, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Torija, M.J.; Rozes, N.; Poblet, M.; Guillamón, J.M.; Mas, A. Yeast population dynamics in spontaneous fermentations: Comparison between two different wine-producing areas over a period of three years. Antonie Leeuwenhoek 2001, 79, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Van der Westhuizen, T.J.; Augustyn, O.P.H.; Pretorius, I.S. Geographical distribution of indigenous Saccharomyces cerevisiae strains isolated from vineyards in the coastal regions of the Western Cape in South Africa. S. Afr. J. Enol. Vitic. 2000, 21, 3–9. [Google Scholar] [CrossRef][Green Version]
- Delfini, C.; Formica, J.V. Wine Microbiology: Science and Technology; Marcel Dekker: New York, NY, USA, 2001. [Google Scholar]
- Nardi, T.; Carlot, M.; De Bortoli, E.; Corich, V.; Giacomini, A. A rapid method for differentiating Saccharomyces sensu stricto strains from other yeast species in an enological environment. FEMS Microbiol. Lett. 2006, 264, 168–173. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nadai, C.; Bovo, B.; Giacomini, A.; Corich, V. New rapid PCR protocol based on high-resolution melting analysis to identify Saccharomyces cerevisiae and other species within its genus. J. Appl. Microbiol. 2018, 124, 1232–1242. [Google Scholar] [CrossRef]
- Querol, A.; Ramon, D. The application of molecular techniques in wine microbiology. Trends Food Sci. Technol. 1996, 7, 73–78. [Google Scholar] [CrossRef]
- Bovo, B.; Giacomini, A.; Corich, V. Effects of grape marcs acidification treatment on the evolution of indigenous yeast populations during the production of grappa. J. Appl. Microbiol. 2011, 111, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, A.R.; Santamaria, P.; Epifanio, S.; Garijo, P.; López, R.L. Ecology of spontaneous fermentation in one winery during 5 consecutive years. Lett. Appl. Microbiol. 1999, 29, 411–415. [Google Scholar] [CrossRef]
- Settanni, L.; Sannino, C.; Francesca, N.; Guarcello, R.; Moschetti, G. Yeast ecology of vineyards within Marsala wine area (western Sicily) in two consecutive vintages and selection of autochthonous Saccharomyces cerevisiae strains. J. Biosci. Bioeng. 2012, 114, 606–614. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vine Plant | H1 | S1 | H2 | ES2 | LS2 | H3 |
|---|---|---|---|---|---|---|
| 1 | − | Y3.1 (100%) | Y3.1 (100%) | − | − | − |
| 2 | − | DV10 (100%) | − | − | − | − |
| 3 | − | T10 (100%) | T10 (100%) | T10 (100%) | − | T10 (100%) |
| 4 | − | − | Y3.1 (100%) | − | − | − |
| 5 | − | Y3.1 (100%) | T13 (100%) | − | − | − |
| 6 | C6.1 (100%) | − | − | Y3.1 (100%) | − | − |
| 7 | − | − | − | − | T17 (100%) | − |
| 8 | − | Y3.1 (100%) | T10 (100%) | Y3.1 (91%) T10 (9%) | − | Y3.1 (100%) |
| 9 | − | T14 (100%) | Y3.1 (100%) | − | − | − |
| 10 | C10.1 (100%) | Y3.1 (100%) | VIN13 (63%) T15 (37%) | T10 (100%) | − | Y3.1 (100%) |
| Total strain number | 2 | 4 | 5 | 2 | 1 | 2 |
| 11 | Y1.1 (10%) Y1.2 (90%) | C6.1 (100%) | Premium Blanc (33%) T4 (67%) | − | Y3.1 (100%) | − |
| 12 | − | DV10 (100%) | Premium Blanc (90%) C6.1 (10%) | − | − | − |
| 13 | Y3.1 (100%) | T1 (100%) | Y3.1 (100%) | Y3.1 (100%) | − | − |
| 14 | Y4.1 (20%) Y4.2 (80%) | − | Y3.1 (100%) | − | T10 (100%) | − |
| 15 | − | − | T5 (100%) | C6.1 (100%) | − | − |
| 16 | − | T2 (100%) | − | T9 (100%) | − | − |
| 17 | − | Premium Blanc (100%) | T6 (75%) T7 (25%) | Y3.1 (100%) | T16 (100%) | − |
| 18 | − | T3 (100%) | T6 (15%) T8 (85%) | T10 (100%) | Y3.1 (100%) | − |
| 19 | − | Y1.1 (100%) | − | Y3.1 (100%) | − | Y3.1 (100%) |
| 20 | − | − | DV10 (100%) | Y3.1 (100%) | Y3.1 (100%) | − |
| Total strain number | 5 | 7 | 9 | 4 | 3 | 1 |
| 21 | − | − | − | |||
| 22 | − | T18 (100%) | − | |||
| 23 | − | T18 (100%) | − | |||
| 24 | T11 (100%) | − | − | |||
| 25 | Y3.1 (100%) | − | − | |||
| 26 | T12 (100%) | T18 (100%) | − | |||
| 27 | − | T18 (100%) | − | |||
| 28 | Y3.1 (100%) | − | Y3.1 (100%) | |||
| 29 | − | − | − | |||
| 30 | − | − | − | |||
| Total strain number | 3 | 1 | 1 |
| 6% EtOH | End Fermentation | |||
|---|---|---|---|---|
| Strain | YV | OV | YV | OV |
| Aromatic K | 0 | 3 | 0 | 3 |
| D254 | 4 | 3 | 4 | 5 |
| ES181 | 11 | 0 | 7 | 1 |
| Fruity flavor | 17 | 21 | 18 | 15 |
| Premium Blanc | 0 | 1 | 0 | 0 |
| VL1 | 7 | 1 | 3 | 3 |
| VL3 | 1 | 1 | 0 | 0 |
| WAM | 3 | 1 | 4 | 1 |
| F2 | 0 | 0 | 1 | 0 |
| F4 | 0 | 0 | 1 | 0 |
| F5 | 0 | 0 | 1 | 0 |
| F7 | 0 | 0 | 1 | 0 |
| F8 | 1 | 0 | 0 | 0 |
| F9 | 2 | 0 | 6 | 5 |
| F12 | 0 | 0 | 0 | 1 |
| F13 | 0 | 0 | 0 | 2 |
| F15 | 0 | 0 | 0 | 1 |
| F17 | 1 | 1 | 0 | 0 |
| F18 | 4 | 0 | 5 | 0 |
| F20 | 0 | 3 | 0 | 0 |
| F21 | 0 | 3 | 0 | 3 |
| F23 | 0 | 6 | 0 | 8 |
| F24 | 0 | 1 | 0 | 0 |
| F25 | 0 | 0 | 0 | 2 |
| F26 | 0 | 0 | 0 | 1 |
| F27 | 0 | 1 | 0 | 0 |
| F28 | 0 | 1 | 0 | 1 |
| F29 | 0 | 1 | 0 | 0 |
| F30 | 0 | 1 | 0 | 0 |
| F31 | 0 | 1 | 0 | 0 |
| F32 | 0 | 0 | 1 | 0 |
| Total colonies | 51 | 50 | 52 | 52 |
| – SO2 | + SO2 | |||
|---|---|---|---|---|
| Strain | YV | OV | YV | OV |
| CRU 211 | 7 | 26 | 0 | 1 |
| D254 | 0 | 1 | 0 | 0 |
| DV10 | 7 | 2 | 7 | 3 |
| ES181 | 5 | 12 | 1 | 9 |
| Fruity flavor | 2 | 6 | 0 | 0 |
| Premium Blanc | 1 | 0 | 0 | 0 |
| VL1 | 21 | 2 | 22 | 8 |
| VL3 | 0 | 0 | 2 | 3 |
| WAM | 3 | 0 | 11 | 25 |
| P2 | 0 | 1 | 0 | 0 |
| P3 | 0 | 0 | 0 | 1 |
| P4 | 2 | 0 | 0 | 6 |
| P5 | 0 | 0 | 0 | 2 |
| P6 | 1 | 0 | 0 | 0 |
| P7 | 1 | 0 | 1 | 0 |
| P8 | 0 | 0 | 1 | 0 |
| P9 | 0 | 0 | 2 | 0 |
| P10 | 0 | 0 | 1 | 0 |
| P11 | 0 | 8 | 0 | 0 |
| P12 | 0 | 0 | 1 | 0 |
| P13 | 0 | 0 | 1 | 0 |
| P14 | 10 | 2 | 10 | 2 |
| Total colonies | 60 | 60 | 60 | 60 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nadai, C.; Vendramini, C.; Carlot, M.; Andrighetto, C.; Giacomini, A.; Corich, V. Dynamics of Saccharomyces cerevisiae Strains Isolated from Vine Bark in Vineyard: Influence of Plant Age and Strain Presence during Grape must Spontaneous Fermentations. Fermentation 2019, 5, 62. https://doi.org/10.3390/fermentation5030062
Nadai C, Vendramini C, Carlot M, Andrighetto C, Giacomini A, Corich V. Dynamics of Saccharomyces cerevisiae Strains Isolated from Vine Bark in Vineyard: Influence of Plant Age and Strain Presence during Grape must Spontaneous Fermentations. Fermentation. 2019; 5(3):62. https://doi.org/10.3390/fermentation5030062
Chicago/Turabian StyleNadai, Chiara, Chiara Vendramini, Milena Carlot, Christian Andrighetto, Alessio Giacomini, and Viviana Corich. 2019. "Dynamics of Saccharomyces cerevisiae Strains Isolated from Vine Bark in Vineyard: Influence of Plant Age and Strain Presence during Grape must Spontaneous Fermentations" Fermentation 5, no. 3: 62. https://doi.org/10.3390/fermentation5030062
APA StyleNadai, C., Vendramini, C., Carlot, M., Andrighetto, C., Giacomini, A., & Corich, V. (2019). Dynamics of Saccharomyces cerevisiae Strains Isolated from Vine Bark in Vineyard: Influence of Plant Age and Strain Presence during Grape must Spontaneous Fermentations. Fermentation, 5(3), 62. https://doi.org/10.3390/fermentation5030062