Mannoprotein Content and Volatile Molecule Profiles of Trebbiano Wines Obtained by Saccharomyces cerevisiae and Saccharomyces bayanus Strains

 ,
,  , and
, and
Abstract

1. Introduction
2. Results
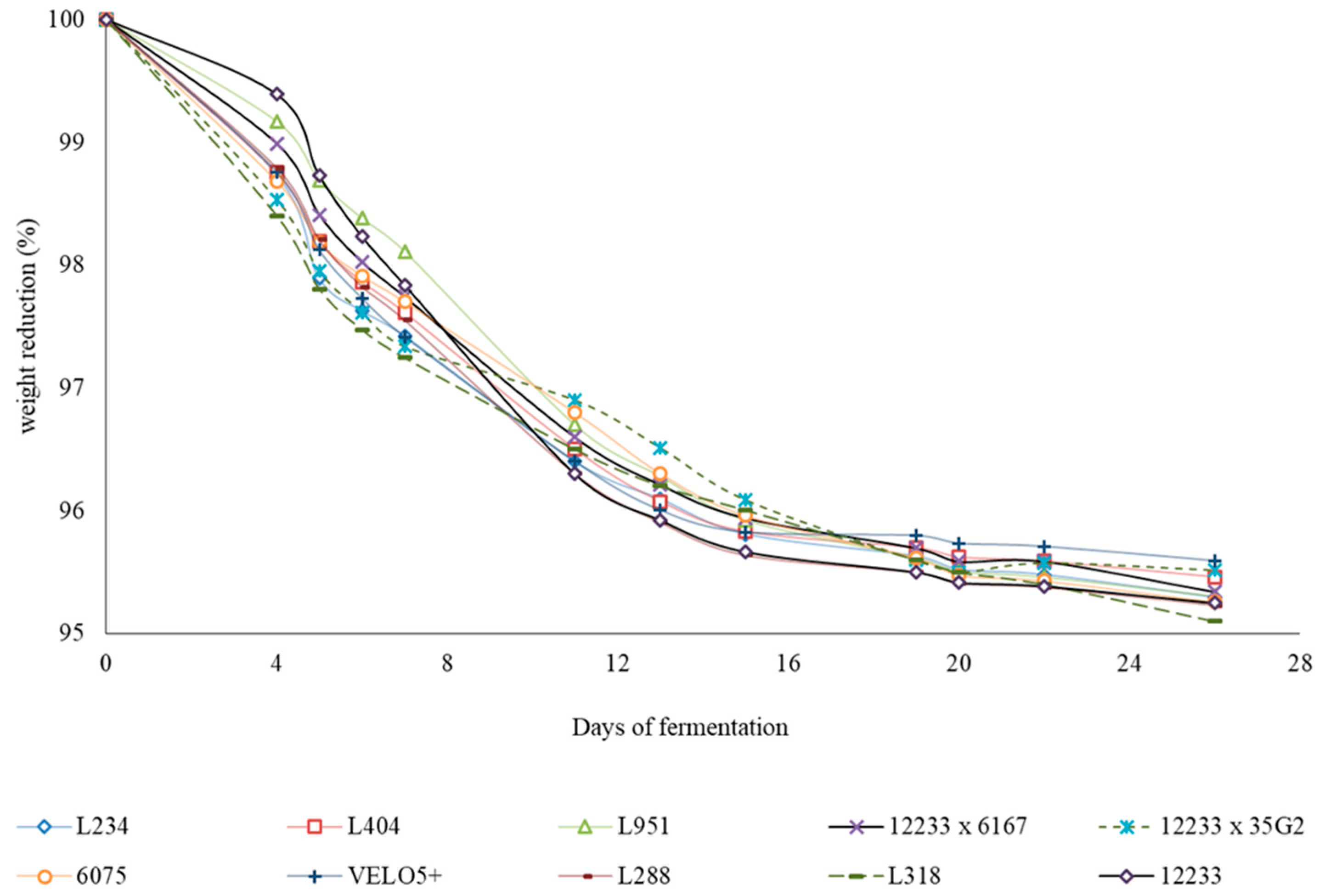
2.1. Yeast Growth and Fermentation Kinetics
2.2. Total Alcoholometric Volumic Title and Residual Sugar
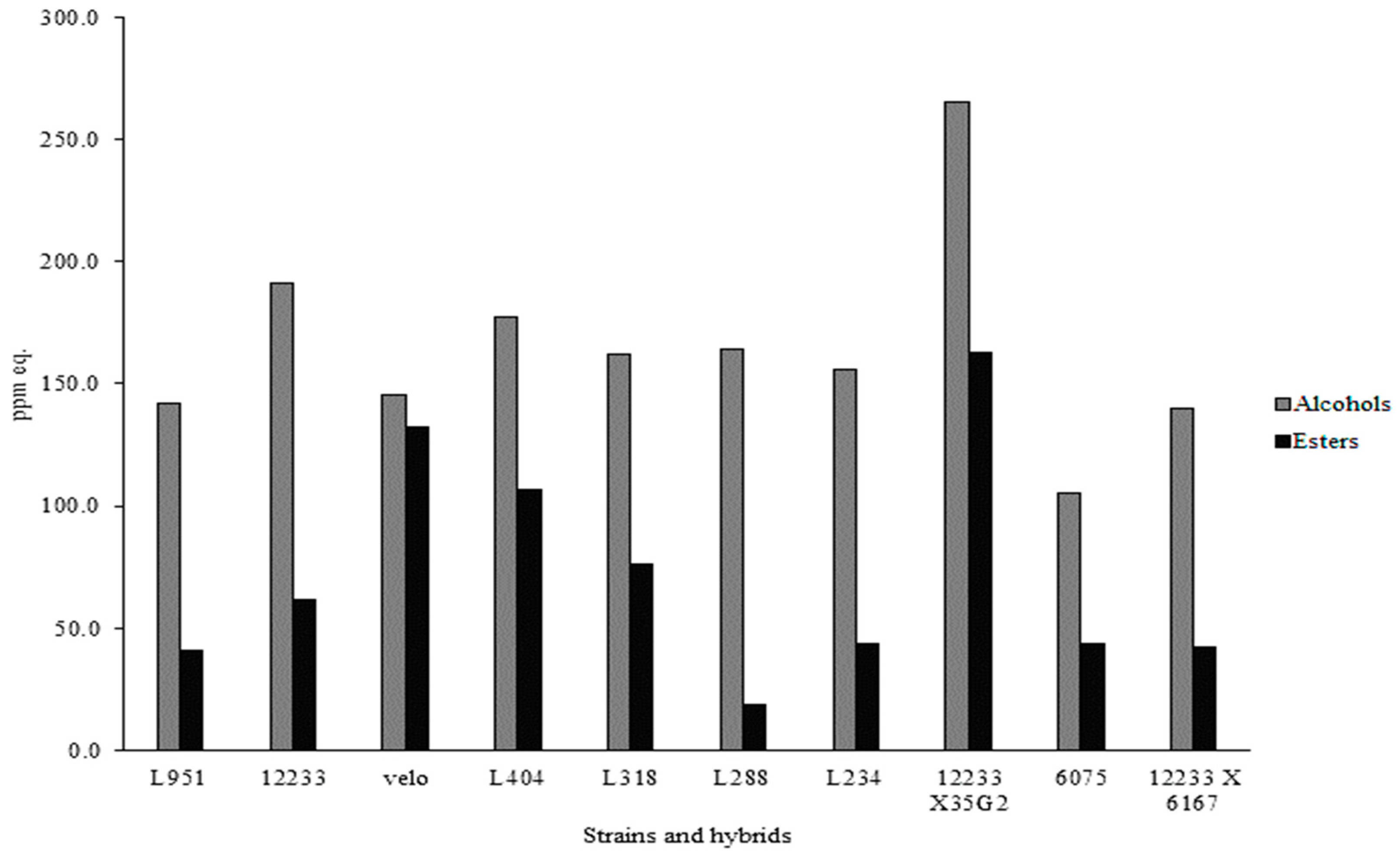
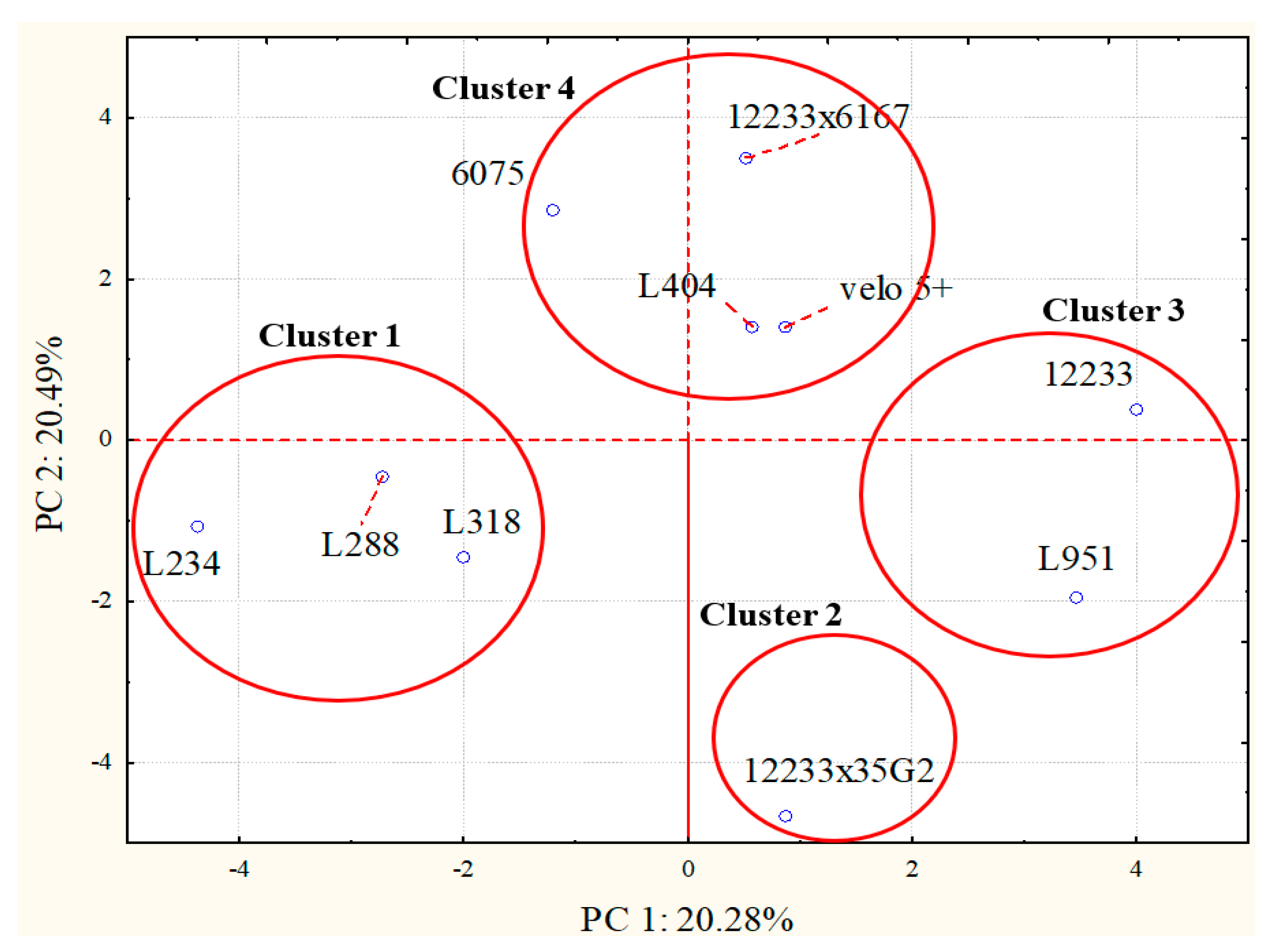
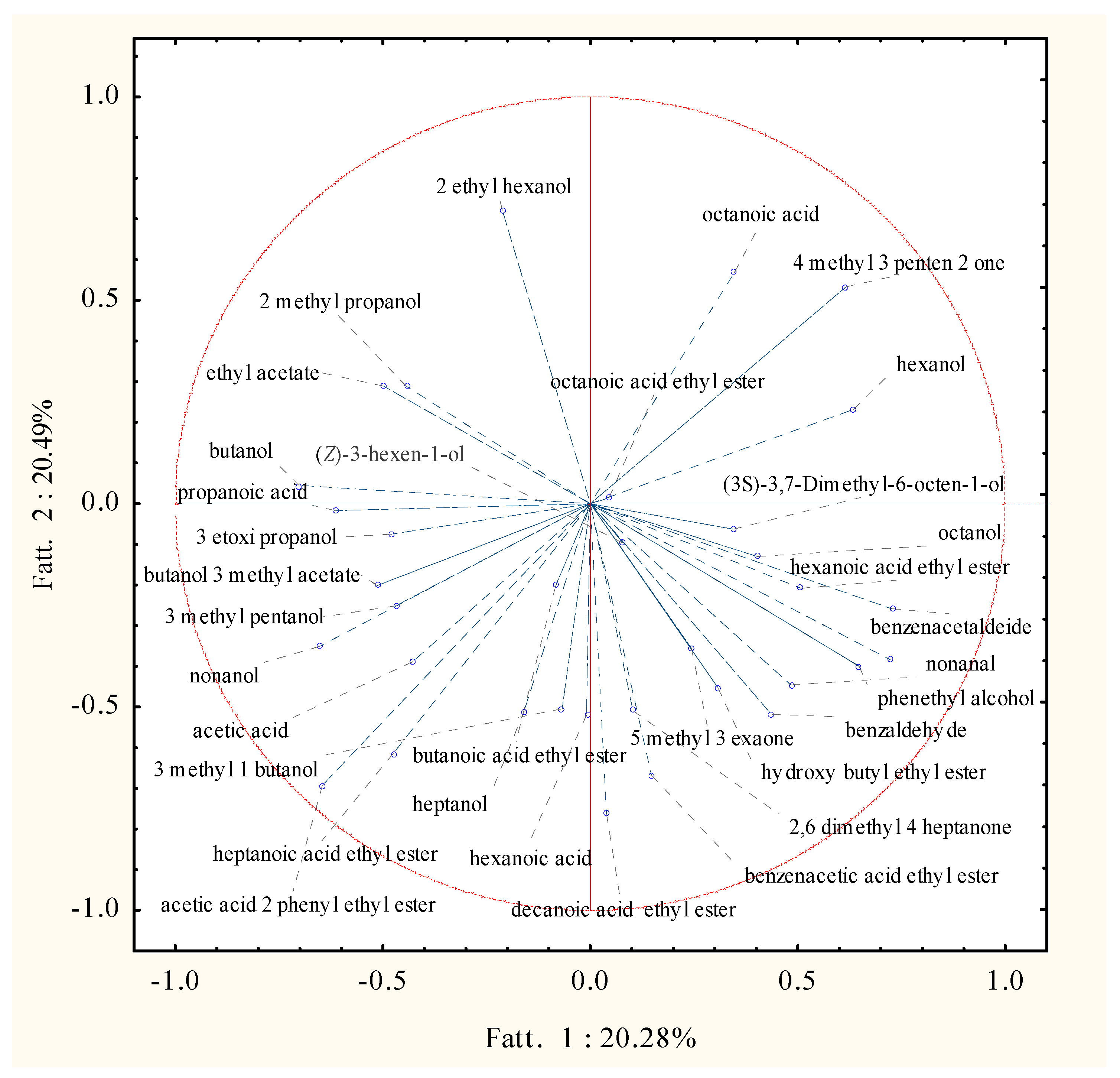
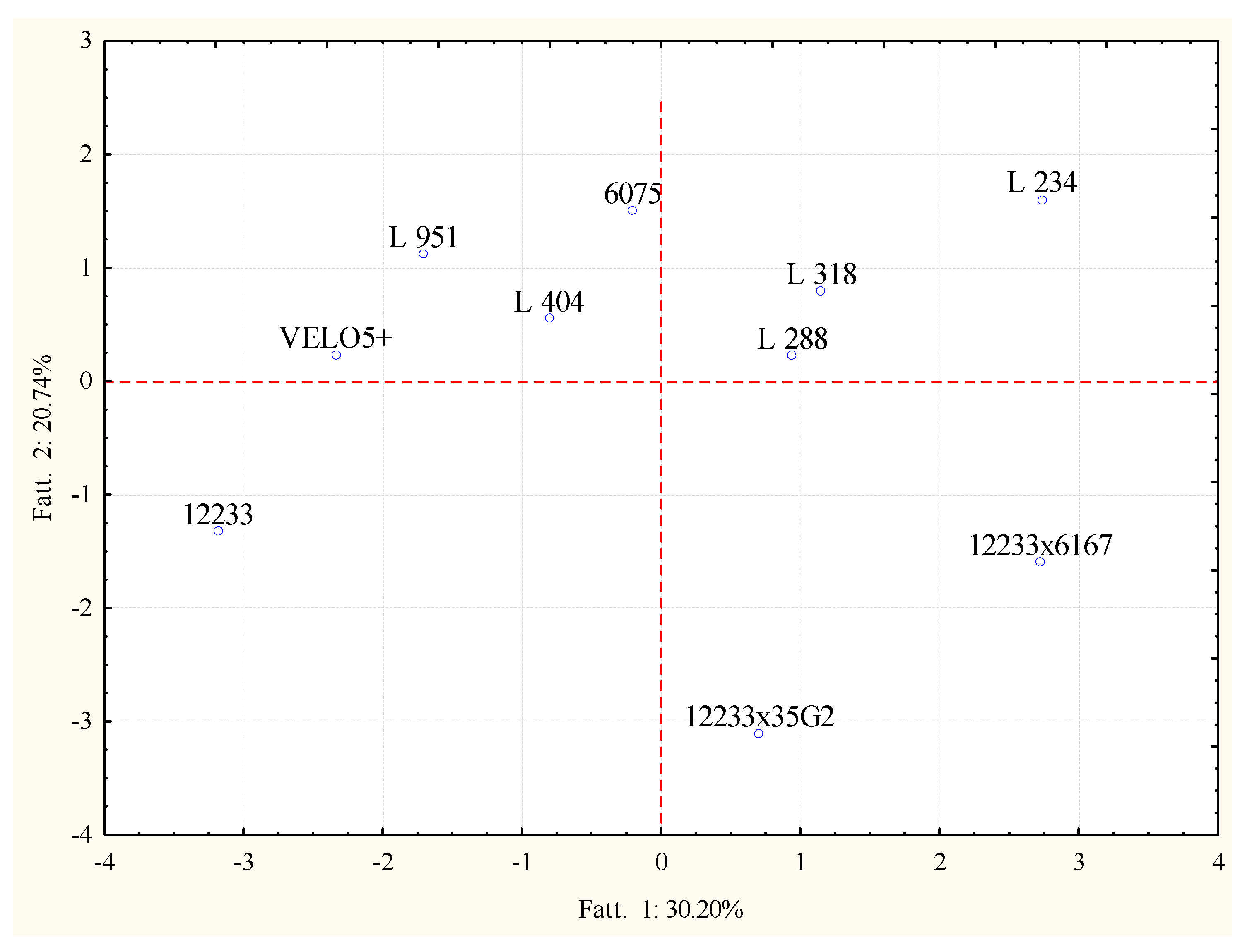
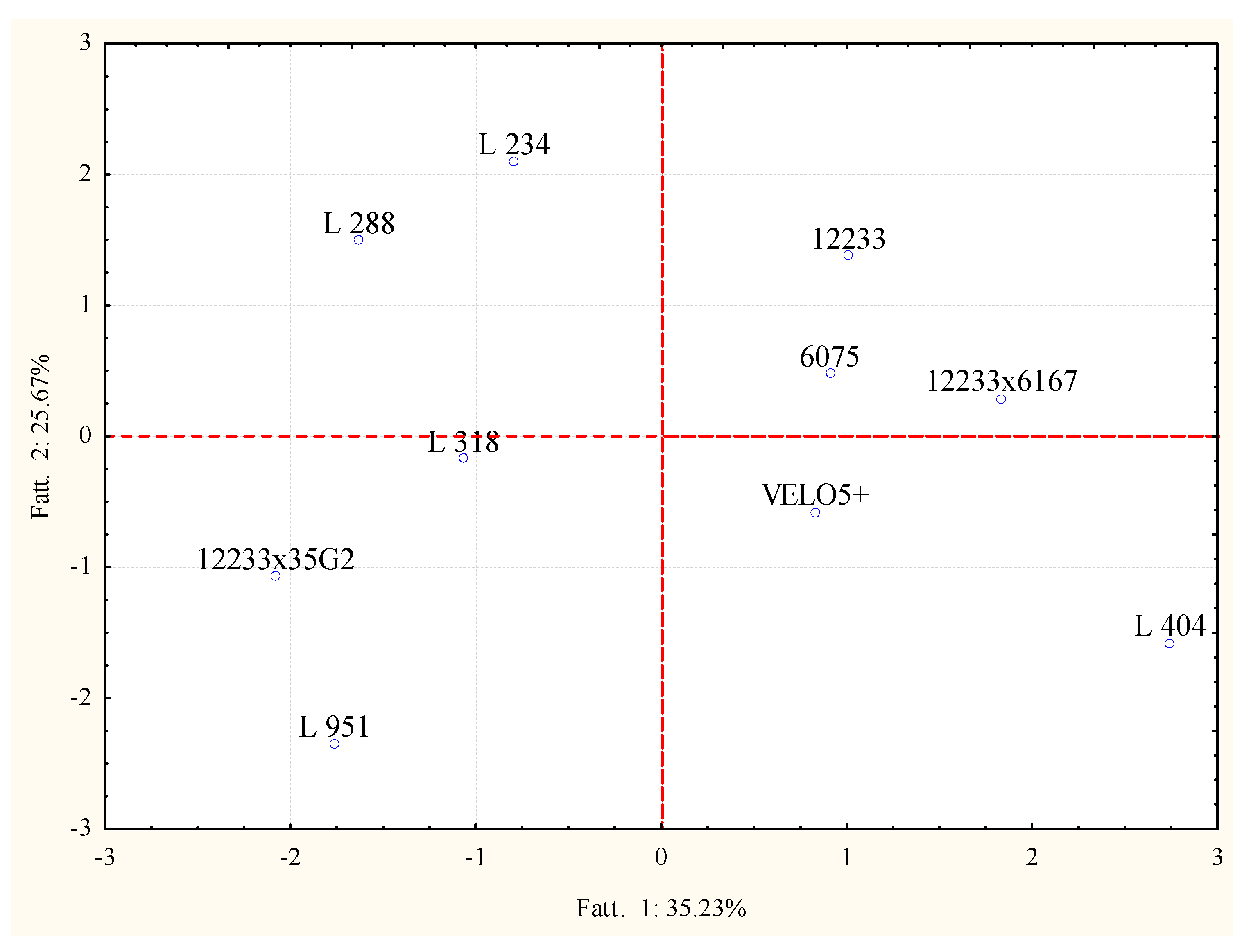
2.3. Volatile Molecular Profiles Analysis
3. Discussion
4. Material and Methods
4.1. Strains
4.2. Micro-Vinifications
4.3. Microbiological Analysis
4.4. Ethanol and WineRreducing Sugars
4.5. Mannoproteins Analysis
4.6. Volatile Molecule Profile
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Pretorius, I.S. Tailoring wine yeast for the new millennium: Novel approaches to the ancient art of winemaking. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef]
- Sagratini, G.; Maggi, F.; Caprioli, G.; Cristalli, G.; Ricciutelli, M.; Torregiani, E.; Vittori, S. Comparative study of aroma profile and phenolic content of Montepulciano monovarietal red wines from the Marches and Abruzzo regions of Italy using HS-SPME–GC–MS and HPLC–MS. Food Chem. 2012, 132, 1592–1599. [Google Scholar] [CrossRef] [PubMed]
- Patrignani, F.; Ndagijimana, M.; Vernocchi, P.; Gianotti, A.; Riponi, C.; Gardini, F.; Lanciotti, R. High-Pressure homogenization to modify yeast performance for sparkling wine production according to traditional methods. Am. J. Enol. Vitic. 2013, 64, 258–267. [Google Scholar] [CrossRef]
- Di Gianvito, P.; Perpetuini, G.; Tittarelli, F.; Schirone, M.; Arfelli, G.; Piva, A.; Patrignani, F.; Lanciotti, R.; Olivastri, L.; Suzzi, G.; et al. Impact of Saccharomyces cerevisiae strains on traditional sparkling wines production. Food Res. Int. 2018, 109, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Guadarrama, A.; Fernandez, J.; Iniguez, M.; Souto, J.; De Saja, J. Discrimination of wine aroma using an array of conducting polymer sensors in conjunction with solid-phase micro-extraction (SPME) technique. Sens. Actuators B Chem. 2001, 77, 401–408. [Google Scholar] [CrossRef]
- Lanciotti, R.; Patrignani, F.; Iucci, L.; Saracino, P.; Guerzoni, M.E. Potential of high pressure homogenization in the control and enhancement of proteolytic and fermentative activities of some Lactobacillus Species. Food Chem. 2007, 102, 542–550. [Google Scholar] [CrossRef]
- Henschke, P. Wine Yeast. In Yeast Sugar Metabolism; Zimmermann, F.K., Entian, K.D., Eds.; Technomic Publishing: Lancaster, PA, USA, 1997. [Google Scholar]
- Jolly, N.; Augustyn, O.; Pretorius, I. The role and use of non-Saccharomyces yeasts in wine production. S. Afri. J. Enol. Vitic. 2006, 27, 15–39. [Google Scholar] [CrossRef]
- Fleet, G. In the Yeasts: Yeast Organelles; Rose, A.H., Harrison, J.S., Eds.; Academic Press: London, UK, 1991. [Google Scholar]
- Klis, F.M.; Boorsma, A.; De Groot, P.W. Cell wall construction in Saccharomyces cerevisiae. Yeast 2006, 23, 185–202. [Google Scholar] [CrossRef]
- Ribéreau-Gayon, P.; Dubourdieu, D.; Donèche, B.; Lonvaud, A. Handbook of Enology, Volume 1: The Microbiology of Wine and Vinifications; John Wiley and Sons: Hoboken, NJ, USA, 2006; Volume 1. [Google Scholar]
- Riou, V.; Vernhet, A.; Doco, T.; Moutounet, M. Aggregation of grape seed tannins in model wine—Effect of wine polysaccharides. Food Hydrocoll. 2002, 16, 17–23. [Google Scholar] [CrossRef]
- Rodrigues, A.; Ricardo-da-Silva, J.M.; Lucas, C.; Laureano, O. Effect of commercial mannoproteins on wine colour and tannins stability. Food Chem. 2012, 131, 907–914. [Google Scholar] [CrossRef]
- Gonzalez-Ramos, D.; Cebollero, E.; Gonzalez, R. A recombinant Saccharomyces cerevisiae strain overproducing mannoproteins stabilizes wine against protein haze. Appl. Environ. Microbiol. 2008, 74, 5533–5540. [Google Scholar] [CrossRef]
- Da Silva Araujo, V.; de Melo, A.N.F.; Costa, A.G.; Castro-Gomez, R.H.; Madruga, M.S.; de Souza, E.L.; Magnani, M. Followed extraction of β-glucan and mannoprotein from spent brewer’s yeast (Saccharomyces uvarum) and application of the obtained mannoprotein as a stabilizer in mayonnaise. Innov. Food Sci. Emerg. Technol. 2014, 23, 164–170. [Google Scholar] [CrossRef]
- Vincenzi, S.; Crapisi, A.; Curioni, A. Foamability of Prosecco wine: Cooperative effects of high molecular weight glycocompounds and wine PR-proteins. Food Hydrocoll. 2014, 34, 202–207. [Google Scholar] [CrossRef]
- Núñez, Y.P.; Carrascosa, A.V.; González, R.; Polo, M.C.; Martínez-Rodríguez, A. Isolation and characterization of a thermally extracted yeast cell wall fraction potentially useful for improving the foaming properties of sparkling Wines. J. Agric. Food Chem. 2006, 54, 7898–7903. [Google Scholar] [CrossRef]
- Escot, S.; Feuillat, M.; Dulau, L.; Charpentier, C. Release of polysaccharides by yeasts and the influence of released polysaccharides on colour stability and wine astringency. Aust. J. Grape Wine Res. 2001, 7, 153–159. [Google Scholar] [CrossRef]
- Alcalde-Eon, C.; Pérez-Mestre, C.; Ferreras-Charro, R.; Rivero, F.J.; Heredia, F.J.; Escribano-Bailon, M.T. The addition of mannoproteins and/or seeds during winemaking and their effects on pigment composition and color stability. J. Agric. Food Chem. 2019, 67, 4031–4042. [Google Scholar] [CrossRef]
- Moruno, E.G.; Sanlorenzo, C.; Boccaccino, B.; Di Stefano, R. Treatment with yeast to reduce the concentration of ochratoxin A in red wine. Am. J. Enol. Vitic. 2005, 56, 73–76. [Google Scholar]
- Caridi, A. New perspectives in safety and quality enhancement of wine through selection of yeasts based on the parietal adsorption activity. Int. J. Food Microbiol. 2007, 120, 167–172. [Google Scholar] [CrossRef]
- Cecchini, F.; Morassut, M.; Moruno, E.G.; Di Stefano, R. Influence of yeast strain on ochratoxin A content during fermentation of white and red must. Food Microbiol. 2006, 23, 411–417. [Google Scholar] [CrossRef]
- Jamal, Z.; Miot-Sertier, C.; Thibau, F.; Dutilh, L.; Lonvaud-Funel, A.; Ballestra, P.; Le Marrec, C.; Dols-Lafargue, M. Distribution and functions of phosphotransferase system genes in the genome of the lactic acid bacterium Oenococcus oeni. Appl. Environ. Microbiol. 2013, 79, 3371–3379. [Google Scholar] [CrossRef]
- Diez, L.; Guadalupe, Z.; Ayestarán, B.N.; Ruiz-Larrea, F. Effect of yeast mannoproteins and grape polysaccharides on the growth of wine lactic acid and acetic acid bacteria. J. Agric. Food Chem. 2010, 58, 7731–7739. [Google Scholar] [CrossRef]
- Legras, J.-L.; Moreno-Garcia, J.; Zara, S.; Zara, G.; Garcia-Martinez, T.; Mauricio, J.C.; Mannazzu, I.; Coi, A.L.; Bou Zeidan, M.; Dequin, S. Flor yeast: New perspectives beyond wine aging. Front. Microbiol. 2016, 7, 503. [Google Scholar] [CrossRef]
- Alexandre, H.; Blanchet, S.; Charpentier, C. Identification of a 49-KDa hydrophobic cell wall mannoprotein present in velum yeast which may be implicated in velum formation. FEMS Microbiol. Lett. 2000, 185, 147–150. [Google Scholar] [CrossRef][Green Version]
- Del Barrio-Galán, R.; Pérez-Magariño, S.; Ortega-Heras, M.; Guadalupe, Z.; Ayestarán, B. Polysaccharide characterization of commercial dry yeast preparations and their effect on white and red wine composition. LWT Food Sci. Technol. 2012, 48, 215–223. [Google Scholar] [CrossRef]
- Guadalupe, Z.; Palacios, A.; Ayestarán, B. Maceration enzymes and mannoproteins: A possible strategy to increase colloidal stability and color extraction in red wines. J. Agric. Food Chem. 2007, 55, 4854–4862. [Google Scholar] [CrossRef]
- White, M.A.; Diffenbaugh, N.; Jones, G.V.; Pal, J.; Giorgi, F. Extreme heat reduces and shifts United States premium wine production in the 21st century. Proc. Natl. Acad. Sci. USA 2006, 103, 11217–11222. [Google Scholar] [CrossRef]
- Quijada-Morín, N.; Williams, P.; Rivas-Gonzalo, J.C.; Doco, T.; Escribano-Bailón, M.T. Polyphenolic, polysaccharide and oligosaccharide composition of Tempranillo red wines and their relationship with the perceived astringency. Food Chem. 2014, 154, 44–51. [Google Scholar] [CrossRef]
- Rinaldi, A.; Coppola, M.; Moio, L. Aging of Aglianico and Sangiovese wine on mannoproteins: Effect on astringency and colour. LWT 2019, 105, 233–241. [Google Scholar] [CrossRef]
- Chalier, P.; Angot, B.; Delteil, D.; Doco, T.; Gunata, Z. Interactions between aroma compounds and whole mannoprotein isolated from Saccharomyces cerevisiae strains. Food Chem. 2007, 100, 22–30. [Google Scholar] [CrossRef]
- Costa, G.P.; Nicolli, K.P.; Welke, J.E.; Manfroi, V.; Zini, C.A. Volatile Profile of sparkling wines produced with the addition of mannoproteins or lees before second fermentation performed with free and immobilized yeasts. J. Braz. Chem. Soc. 2018, 29, 1866–1875. [Google Scholar] [CrossRef]
- Mateo, J.; Jiménez, M. Monoterpenes in grape juice and wines. J. Chromatogr. A 2000, 881, 557–567. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F. Chapter 4—Use of Non-Saccharomyces Yeasts in Red Winemaking. In Red Wine Technology; Morata, A., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 51–68. [Google Scholar] [CrossRef]
- Thongekkaew, J.; Fujii, T.; Masaki, K.; Koyama, K. Evaluation of Candida easanensis JK8 β-glucosidase with potentially hydrolyse non-volatile glycosides of wine aroma precursors. Nat. Prod. Res. 2018, 1–5. [Google Scholar] [CrossRef]
- Maturano, Y.P.; Assof, M.; Fabani, M.P.; Nally, M.C.; Jofré, V.; Assaf, L.A.R.; Toro, M.E.; De Figueroa, L.I.C.; Vazquez, F. Enzymatic activities produced by mixed Saccharomyces and non-Saccharomyces cultures: Relationship with wine volatile composition. Antonie Van Leeuwenhoek 2015, 108, 1239–1256. [Google Scholar] [CrossRef]
- Rodríguez, M.; Lopes, C.; Van Broock, M.; Valles, S.; Ramón, D.; Caballero, A. Screening and typing of Patagonian wine yeasts for glycosidase activities. J. Appl. Microbiol. 2004, 96, 84–95. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and future of non-Saccharomyces yeasts: From spoilage microorganisms to biotechnological tools for improving wine aroma complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef]
- Doco, T.; Vuchot, P.; Cheynier, V.; Moutounet, M. Structural modification of wine arabinogalactans during aging on lees. Am. J. Enol. Vitic. 2003, 54, 150–157. [Google Scholar]
- Vidal, S.; Williams, P.; Doco, T.; Moutounet, M.; Pellerin, P. The polysaccharides of red wine: Total fractionation and characterization. Carbohydr. Polym. 2003, 54, 439–447. [Google Scholar] [CrossRef]
- Pérez-Través, L.; Querol, A.; Pérez-Torrado, R. Increased mannoprotein content in wines produced by Saccharomyces kudriavzevii× Saccharomyces cerevisiae hybrids. Int. J. Food Microbiol. 2016, 237, 35–38. [Google Scholar] [CrossRef]
- Vernocchi, P.; Ndagijimana, M.; Serrazanetti, D.I.; López, C.C.; Fabiani, A.; Gardini, F.; Guerzoni, M.E.; Lanciotti, R. Use of Saccharomyces cerevisiae strains endowed with β-glucosidase activity for the production of Sangiovese wine. World J. Microbiol. Biotechnol. 2011, 27, 1423–1433. [Google Scholar] [CrossRef]
- OIV. Resolution OENO 26/2004; OIV: Paris, France, 2004. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Days of Fermentation | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 4 | 11 | 19 | 26 | ||||||||||||
| Saccharomyces cerevisiae | L 234 | 6.1 | ± | 0.3 | 6.6 | ± | 0.3 | 7.0 | ± | 0.3 | 7.4 | ± | 0.3 | 6.9 | ± | 0.2 |
| 12233 | 4.9 | ± | 0.4 | 5.5 | ± | 0.2 | 7.0 | ± | 0.3 | 7.8 | ± | 0.5 | 7.0 | ± | 0.3 | |
| L 288 | 5.9 | ± | 0.1 | 6.1 | ± | 0.5 | 6.9 | ± | 0.2 | 7.4 | ± | 0.3 | 6.9 | ± | 0.3 | |
| L 404 | 5.9 | ± | 0.3 | 6.5 | ± | 0.4 | 7.1 | ± | 0.4 | 7.8 | ± | 0.3 | 7.2 | ± | 0.4 | |
| L 318 | 6.3 | ± | 0.3 | 7.1 | ± | 0.4 | 7.4 | ± | 0.35 | 7.8 | ± | 0.4 | 7.0 | ± | 0.3 | |
| VELO5+ | 5.5 | ± | 0.5 | 6.0 | ± | 0.4 | 7.1 | ± | 0.3 | 7.7 | ± | 0.5 | 7.2 | ± | 0.2 | |
| 6075 | 6.1 | ± | 0.4 | 6.5 | ± | 0.4 | 6.6 | ± | 0.26 | 7.0 | ± | 0.3 | 6.5 | ± | 0.3 | |
| S. cerevisiae hybrids | 12233x35G2 | 5.2 | ± | 0.4 | 6.5 | ± | 0.5 | 7.5 | ± | 0.42 | 7.9 | ± | 0.2 | 7.2 | ± | 0.5 |
| 12233x6167 | 6.3 | ± | 0.6 | 7.0 | ± | 0.3 | 7.2 | ± | 0.44 | 7.7 | ± | 0.3 | 7.1 | ± | 0.4 | |
| Saccharomyces bayanus | L 951 | 5.4 | ± | 0.4 | 5.9 | ± | 0.4 | 6.2 | ± | 0.3 | 6.2 | ± | 0.4 | 6.0 | ± | 0.4 |
| Strain | TAV | Residual Sugar (g/l) | |||||
|---|---|---|---|---|---|---|---|
| S. cerevisiae | L 234 | 10.10 a,* | ± | 0.12 | 4.00 a,* | ± | 0.20 |
| L12233 | 11.12 b | ± | 0.11 | 1.50 b,d | ± | 0.30 | |
| L 288 | 11.50 c | ± | 0.15 | 1.00 b | ± | 0.50 | |
| L 404 | 10.80 d | ± | 0.10 | 4.00 a | ± | 0.30 | |
| VELO 5+ | 11.20 b | ± | 0.09 | 2.00 d | ± | 0.30 | |
| 6075 | 11.00 b | ± | 0.17 | 4.00 a | ± | 0.31 | |
| L 318 | 11.40 c | ± | 0.11 | 2.00 d | ± | 0.30 | |
| S.cerevisiae hybrids | 12233x35G2 | 10.80 d | ± | 0.28 | 5.00 c | ± | 0.34 |
| 12233x6167 | 10.70 d | ± | 0.33 | 3.00 e | ± | 0.29 | |
| S. bayanus | L 951 | 10.70 d | ± | 0.11 | 5.00 c | ± | 0.31 |
| Strain | Mannoproteins (mg/L) | |||
|---|---|---|---|---|
| S. cerevisiae | L 234 | 57.0 a,* | ± | 0.3 |
| 12233 | 87.0 b | ± | 0.2 | |
| L 288 | 72.0 c | ± | 0.2 | |
| L 404 | 69.0 d | ± | 0.3 | |
| 6075 | 56.0 i | ± | 0.1 | |
| L 318 | 67.0 l | ± | 0.1 | |
| VELO 5+ | 58.0 f | ± | 0.1 | |
| S. cerevisiae hybrids | 12233x35G2 | 83.0 g | ± | 0.1 |
| 12233x6167 | 104.0 h | ± | 0.3 | |
| S. bayanus | L 951 | 92.0 e | ± | 0.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braschi, G.; Ricci, A.; Grazia, L.; Versari, A.; Patrignani, F.; Lanciotti, R. Mannoprotein Content and Volatile Molecule Profiles of Trebbiano Wines Obtained by Saccharomyces cerevisiae and Saccharomyces bayanus Strains. Fermentation 2019, 5, 66. https://doi.org/10.3390/fermentation5030066
Braschi G, Ricci A, Grazia L, Versari A, Patrignani F, Lanciotti R. Mannoprotein Content and Volatile Molecule Profiles of Trebbiano Wines Obtained by Saccharomyces cerevisiae and Saccharomyces bayanus Strains. Fermentation. 2019; 5(3):66. https://doi.org/10.3390/fermentation5030066
Chicago/Turabian StyleBraschi, Giacomo, Arianna Ricci, Luigi Grazia, Andrea Versari, Francesca Patrignani, and Rosalba Lanciotti. 2019. "Mannoprotein Content and Volatile Molecule Profiles of Trebbiano Wines Obtained by Saccharomyces cerevisiae and Saccharomyces bayanus Strains" Fermentation 5, no. 3: 66. https://doi.org/10.3390/fermentation5030066
APA StyleBraschi, G., Ricci, A., Grazia, L., Versari, A., Patrignani, F., & Lanciotti, R. (2019). Mannoprotein Content and Volatile Molecule Profiles of Trebbiano Wines Obtained by Saccharomyces cerevisiae and Saccharomyces bayanus Strains. Fermentation, 5(3), 66. https://doi.org/10.3390/fermentation5030066