
TWEAK Negatively Regulates Human Dicer

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
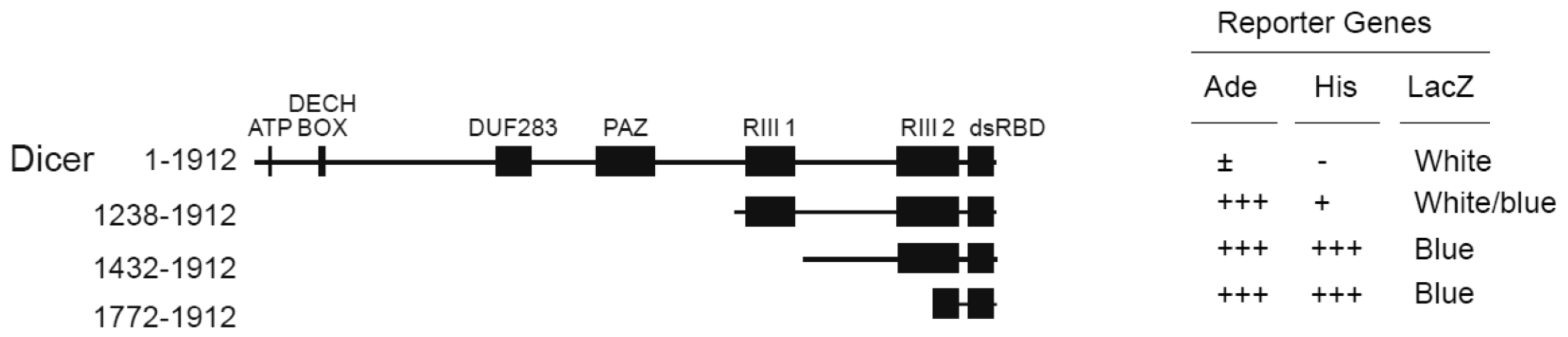
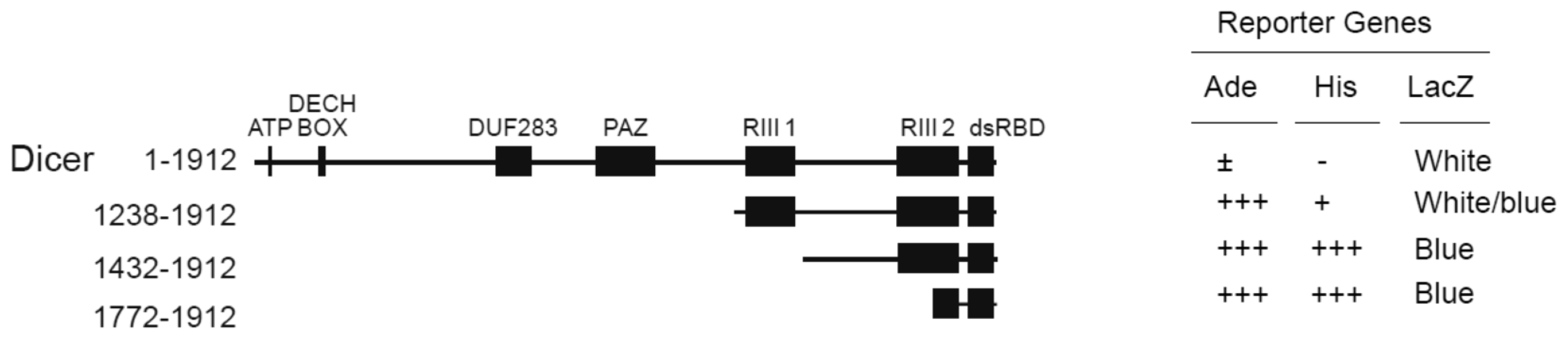
2.1. TWEAK Interacts with the C-Terminal dsRBD of Human Dicer in Yeast Two-Hybrid Assays
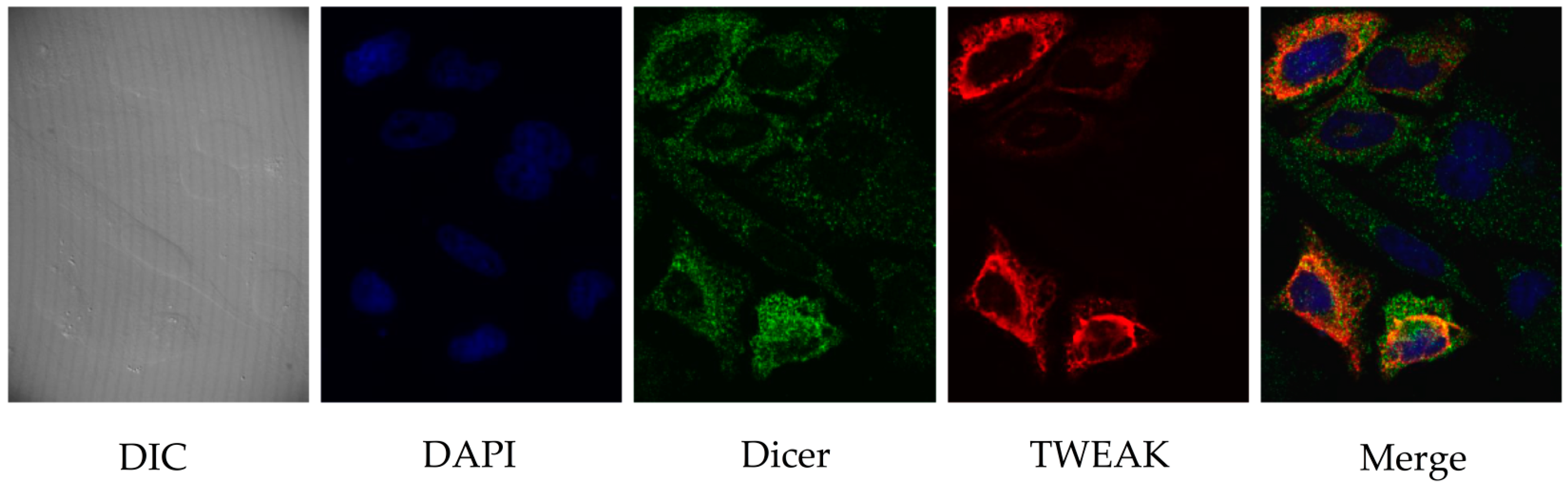
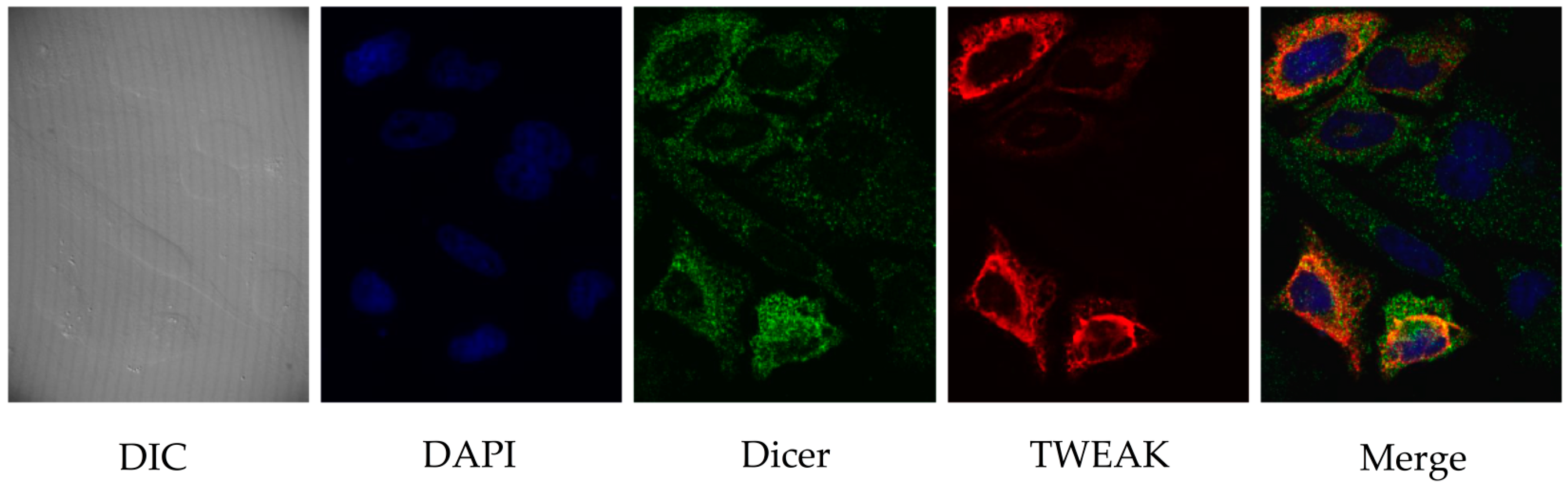
2.2. TWEAK and Dicer Proteins Colocalize at the Perinuclear Region of HeLa Cells
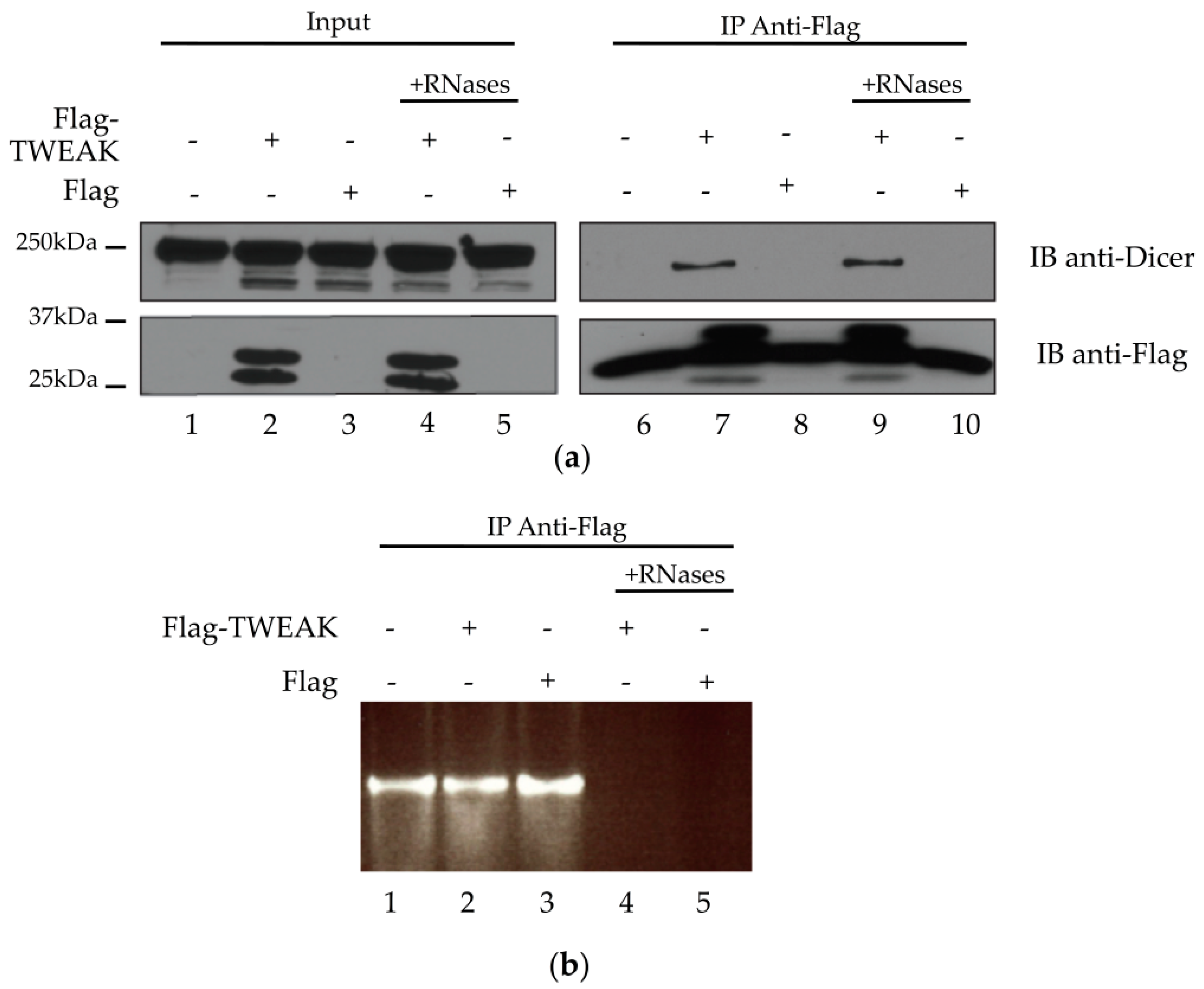
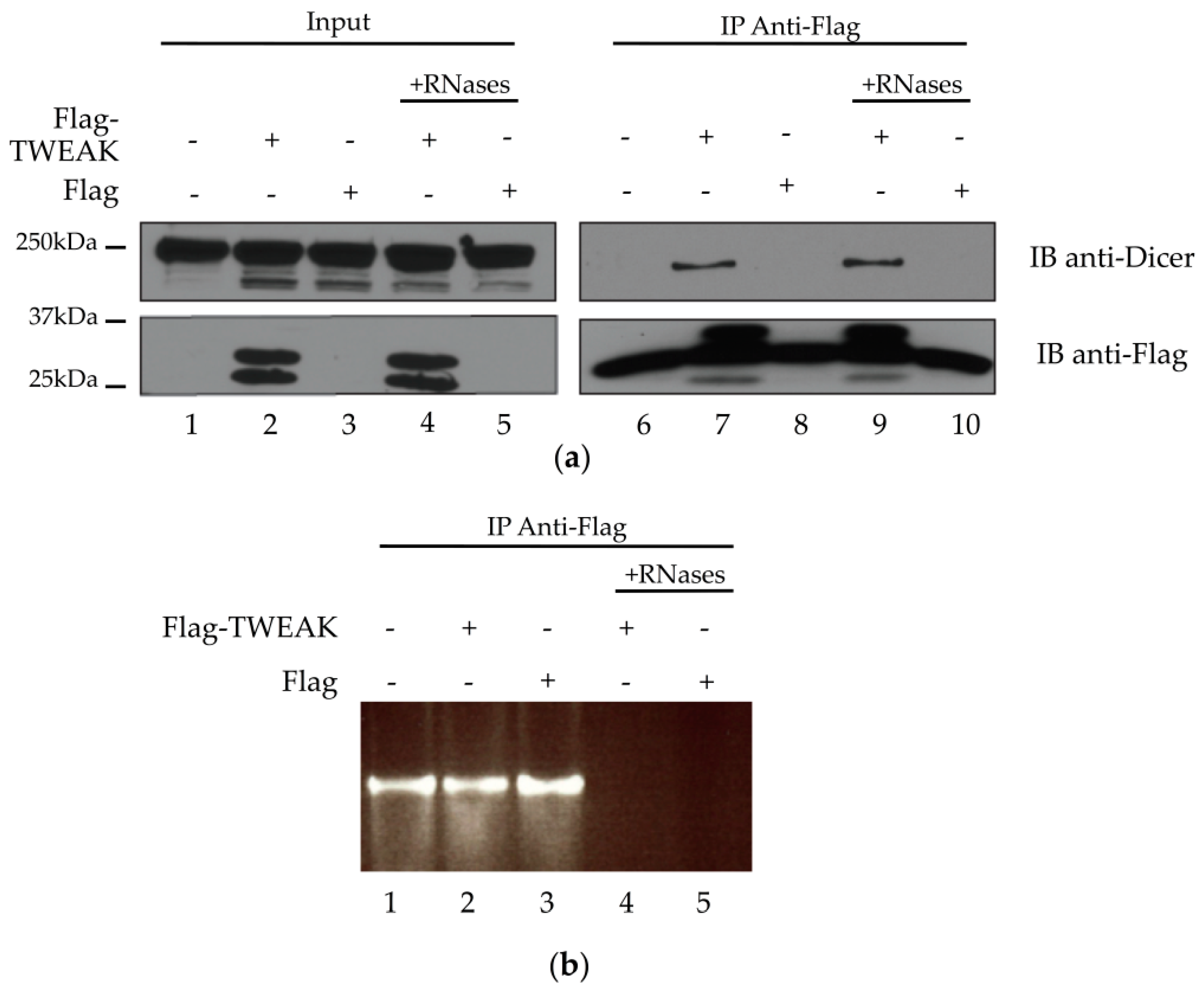
2.3. Dicer and TWEAK Proteins Form a Complex Likely Independent of RNA in HEK 293 Cells
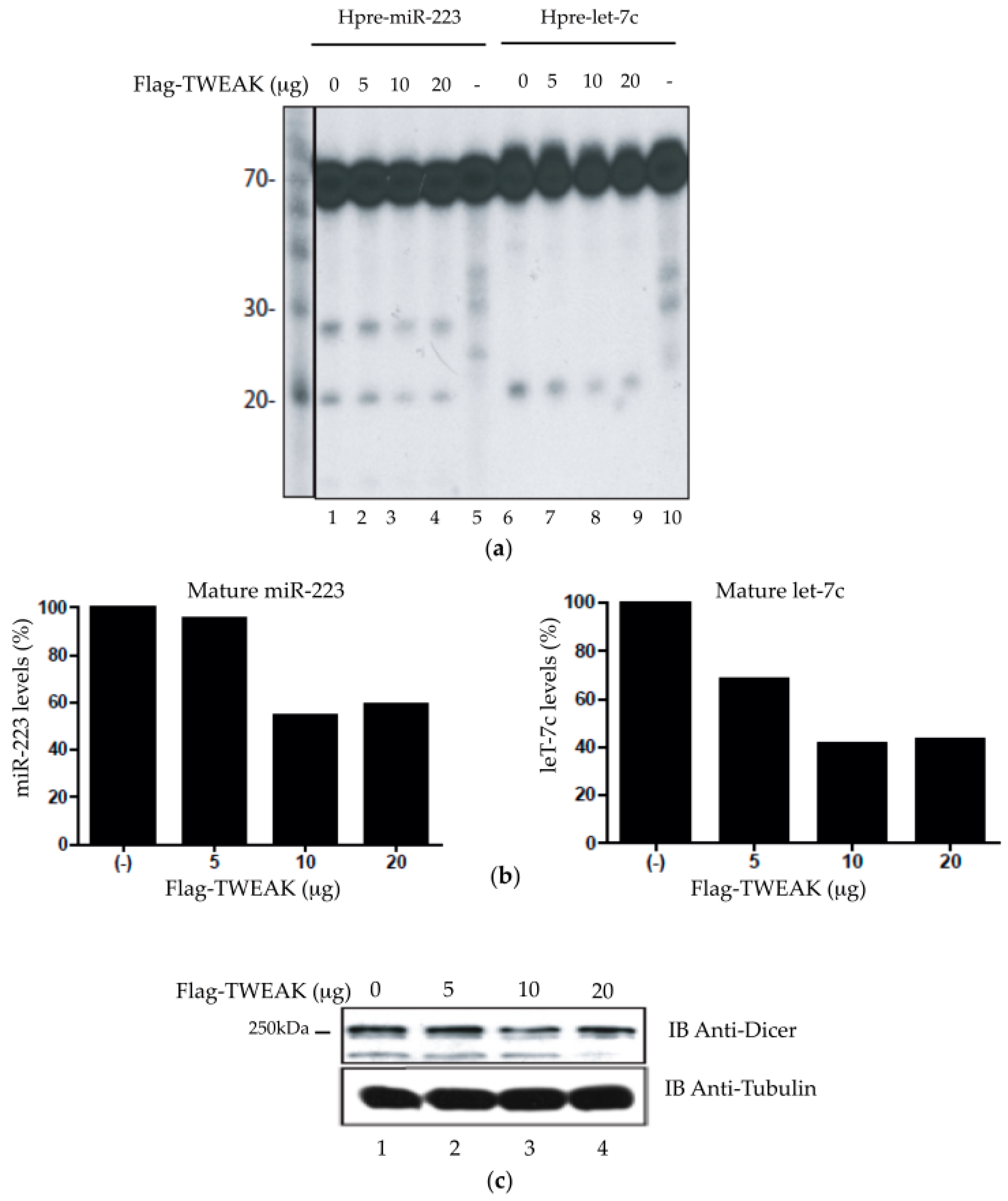
2.4. TWEAK Dose-Dependently Attenuates the Pre-MicroRNA Processing Activity of Dicer
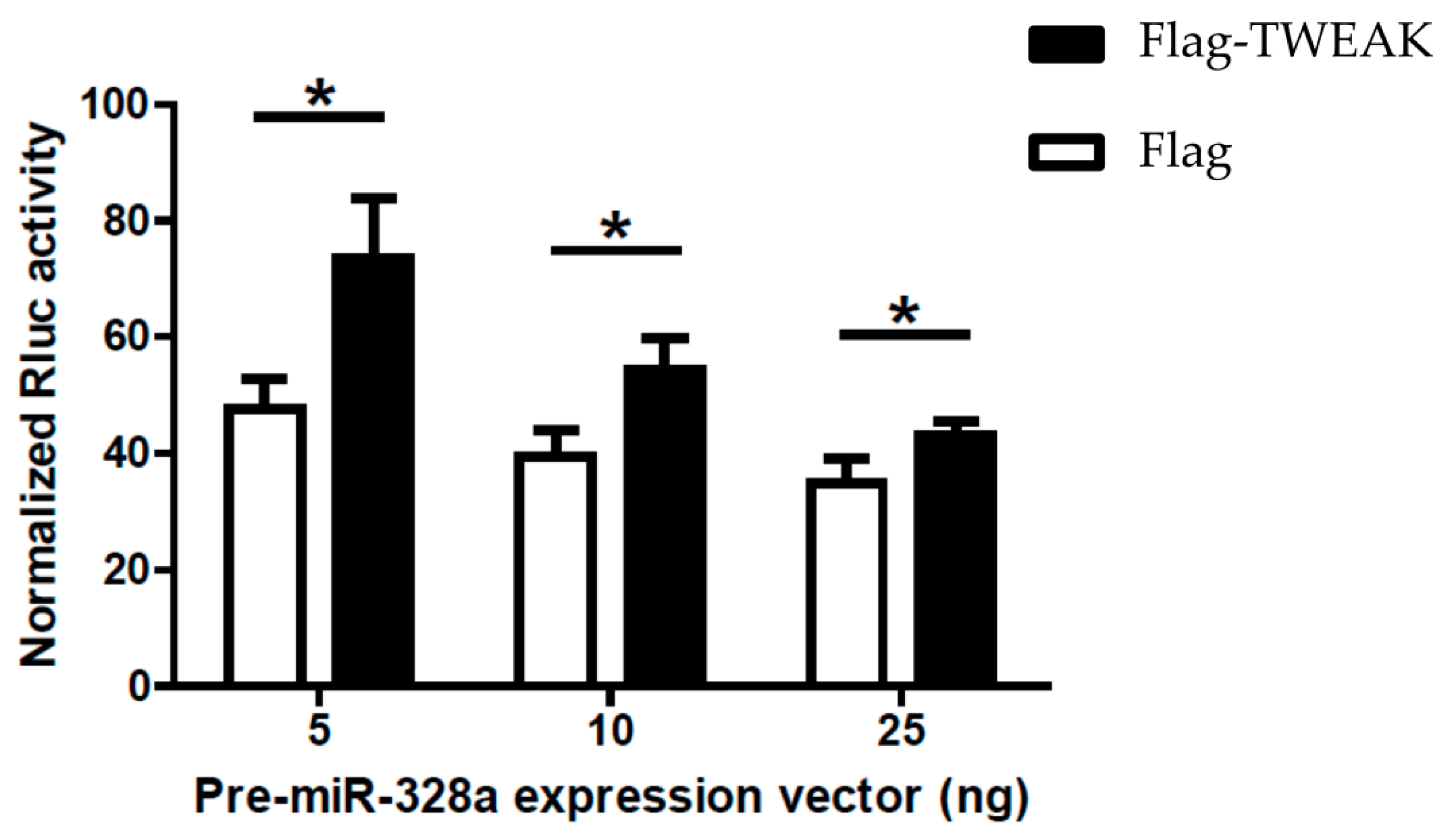
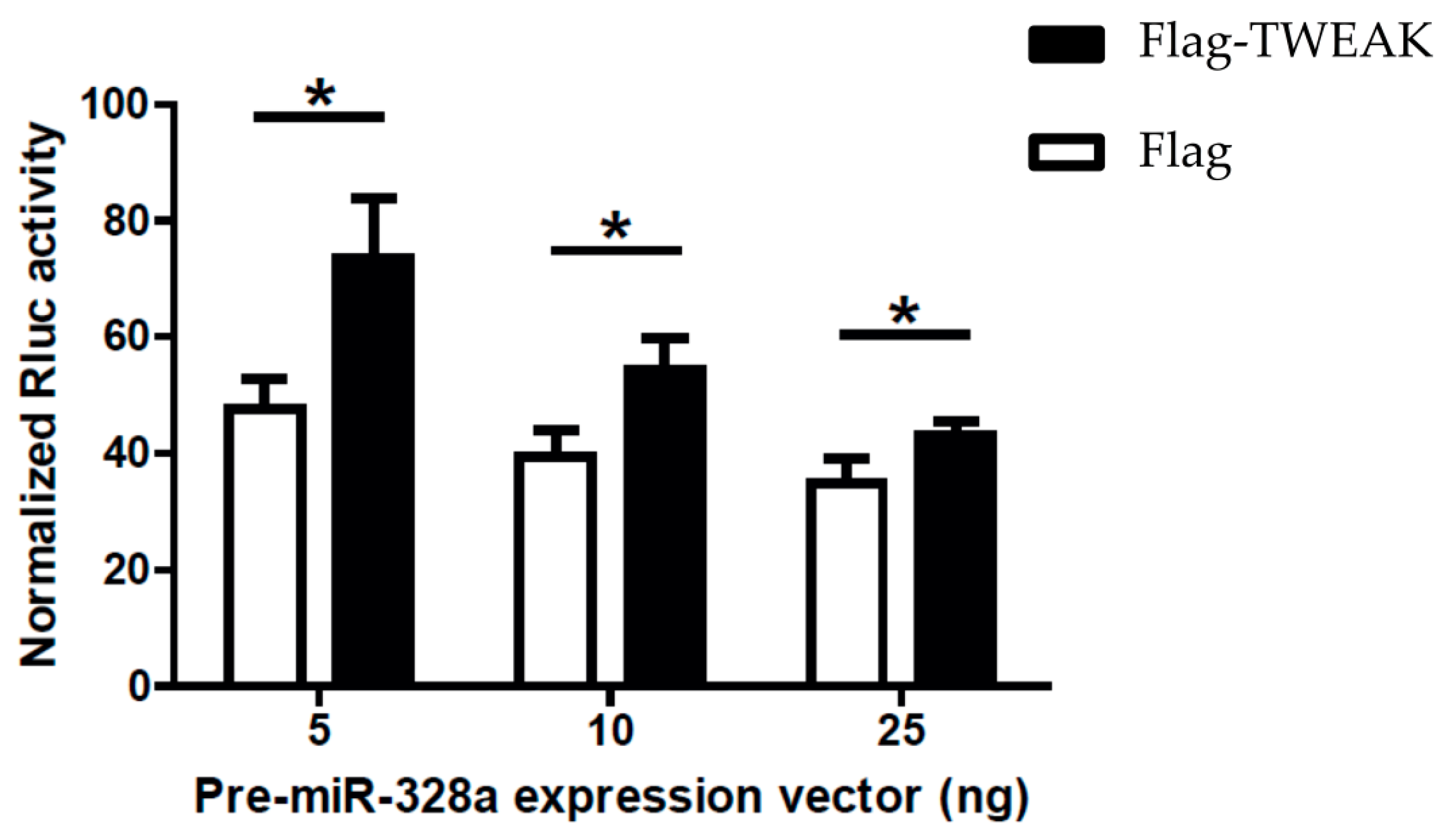
2.5. TWEAK Impairs MicroRNA-Guided RNA Silencing of a Reporter Gene Induced by a Pre-MicroRNA
3. Discussion
4. Materials and Methods
4.1. Plasmid DNA Constructs
4.2. Yeast Two-Hybrid Experiments
4.3. Confocal Immunofluorescence Microscopy
4.4. Protein Extracts and Western Blot Analysis
4.5. Immunoprecipitation Experiments
4.6. Dicer Activity Assays
4.7. Reporter Gene Activity Assay
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Kim, V.N.; Han, J.; Siomi, M.C. Biogenesis of small RNAs in animals. Nat. Rev. Mol. Cell. Biol. 2009, 10, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Provost, P.; Dishart, D.; Doucet, J.; Frendewey, D.; Samuelsson, B.; Radmark, O. Ribonuclease activity and RNA binding of recombinant human Dicer. EMBO J. 2002, 21, 5864–5874. [Google Scholar] [CrossRef] [PubMed]
- Lau, P.-W.; Guiley, K.Z.; De, N.; Potter, C.S.; Carragher, B.; MacRae, I.J. The molecular architecture of human Dicer. Nat. Struct. Mol. Biol. 2012, 19, 436–440. [Google Scholar] [CrossRef] [PubMed]
- Chendrimada, T.P.; Gregory, R.I.; Kumaraswamy, E.; Norman, J.; Cooch, N.; Nishikura, K.; Shiekhattar, R. TRBP recruits the dicer complex to Ago2 for microRNA processing and gene silencing. Nature 2005, 436, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Haase, A.D.; Jaskiewicz, L.; Zhang, H.; Laine, S.; Sack, R.; Gatignol, A.; Filipowicz, W. TRBP, a regulator of cellular PKR and HIV-1 virus expression, interacts with Dicer and functions in RNA silencing. EMBO Rep. 2005, 6, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Hur, I.; Park, S.Y.; Kim, Y.K.; Suh, M.R.; Kim, V.N. The role of PACT in the RNA silencing pathway. EMBO J. 2006, 25, 522–532. [Google Scholar] [CrossRef] [PubMed]
- Meister, G.; Landthaler, M.; Peters, L.; Chen, P.Y.; Urlaub, H.; Luhrmann, R.; Tuschl, T. Identification of novel Argonaute-associated proteins. Curr. Biol. 2005, 15, 2149–2155. [Google Scholar] [CrossRef] [PubMed]
- Plante, I.; Davidovic, L.; Ouellet, D.L.; Gobeil, L.A.; Tremblay, S.; Khandjian, E.W.; Provost, P. Dicer-derived microRNAs are utilized by the fragile X mental retardation protein for assembly on target rRNAs. J. Biomed. Biotechnol. 2006, 2006, 64347. [Google Scholar] [CrossRef] [PubMed]
- Pépin, G.; Perron, M.P.; Provost, P. Regulation of human Dicer by the resident ER membrane protein CLIMP-63. Nucleic Acids Res. 2012, 40, 11603–11617. [Google Scholar] [CrossRef] [PubMed]
- Chicheportiche, Y.; Bourdon, P.R.; Xu, H.; Hsu, Y.M.; Scott, H.; Hession, C.; Garcia, I.; Browning, J.L. TWEAK, a new secreted ligand in the tumor necrosis factor family that weakly induces apoptosis. J. Biol. Chem. 1997, 272, 32401–32410. [Google Scholar] [CrossRef] [PubMed]
- Dincbas-Renqvist, V.; Pepin, G.; Rakonjac, M.; Plante, I.; Ouellet, D.L.; Hermansson, A.; Goulet, I.; Doucet, J.; Samuelsson, B.; Radmark, O.; et al. Human Dicer C-terminus functions as a 5-lipoxygenase binding domain. Biochim. Biophys. Acta 2009, 1789, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Provost, P.; Samuelsson, B.; Radmark, O. Interaction of 5-lipoxygenase with cellular proteins. Proc. Natl. Acad. Sci. USA 1999, 96, 1881–1885. [Google Scholar] [CrossRef] [PubMed]
- Provost, P.; Doucet, J.; Hammarberg, T.; Gerisch, G.; Samuelsson, B.; Radmark, O. 5-Lipoxygenase interacts with coactosin-like protein. J. Biol. Chem. 2001, 276, 16520–16527. [Google Scholar] [CrossRef] [PubMed]
- Boissonneault, V.; Plante, I.; Rivest, S.; Provost, P. MicroRNA-298 and microRNA-328 regulate expression of mouse beta-amyloid precursor protein-converting enzyme 1. J. Biol. Chem. 2009, 284, 1971–1981. [Google Scholar] [CrossRef] [PubMed]
- Boissonneault, V.; St-Gelais, N.; Plante, I.; Provost, P. A polymerase chain reaction-based cloning strategy applicable to functional microRNA studies. Anal. Biochem. 2008, 381, 166–168. [Google Scholar] [CrossRef] [PubMed]
- James, P.; Halladay, J.; Craig, E.A. Genomic libraries and a host strain designed for highly efficient two-hybrid selection in yeast. Genetics 1996, 144, 1425–1436. [Google Scholar] [PubMed]
- Ma, E.; MacRae, I.J.; Kirsch, J.F.; Doudna, J.A. Autoinhibition of human Dicer by its internal helicase domain. J. Mol. Biol. 2008, 380, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Zhou, K.; Smith, A.M.; Noland, C.L.; Doudna, J.A. Differential roles of human Dicer-binding proteins TRBP and PACT in small RNA processing. Nucleic Acids Res. 2013, 41, 6568–6576. [Google Scholar] [CrossRef] [PubMed]
- Daniels, S.M.; Melendez-Peña, C.E.; Scarborough, R.J.; Daher, A.; Christensen, H.S.; El Far, M.; Purcell, D.F.J.; Lainé, S.; Gatignol, A. Characterization of the TRBP domain required for Dicer interaction and function in RNA interference. BMC Mol. Biol. 2009, 65, 1373–1378. [Google Scholar] [CrossRef] [PubMed]
- Wostenberg, C.; Lary, J.W.; Sahu, D.; Acevedo, R.; Quarles, K.A.; Cole, J.L.; Showalter, S.A. The role of human Dicer-dsRBD in processing small regulatory RNAs. PLoS ONE 2012, 7, e51829. [Google Scholar] [CrossRef] [PubMed]
- Guil, S.; Caceres, J.F. The multifunctional RNA-binding protein hnRNP A1 is required for processing of miR-18a. Nat. Struct. Mol. Biol. 2007, 14, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Trabucchi, M.; Briata, P.; Filipowicz, W.; Ramos, A.; Gherzi, R.; Rosenfeld, M.G. KSRP promotes the maturation of a group of miRNA precursors. Adv. Exp. Med. Biol. 2010, 700, 1493–1500. [Google Scholar]
- Panguluri, S.K.; Bhatnagar, S.; Kumar, A.; McCarthy, J.J.; Srivastava, A.K.; Cooper, N.G.; Lundy, R.F.; Kumar, A. Genomic profiling of messenger RNAs and microRNAs reveals potential mechanisms of TWEAK-induced skeletal muscle wasting in mice. PLoS ONE 2010, 5, e8760. [Google Scholar] [CrossRef] [PubMed]
- Clawson, G.A.; Matters, G.L.; Xin, P.; Imamura-Kawasawa, Y.; Du, Z.; Thiboutot, D.M.; Helm, K.F.; Neves, R.I.; Abraham, T. Macrophage-tumor cell fusions from peripheral blood of melanoma patients. PLoS ONE 2015, 10, e0134320. [Google Scholar] [CrossRef] [PubMed]
- De Ketelaere, A.; Vermeulen, L.; Vialard, J.; van de Weyer, I.; van Wauwe, J.; Haegeman, G.; Moelans, I. Involvement of GSK-3β in TWEAK-mediated NF-κb activation. FEBS Lett. 2004, 566, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Roos, C.; Wicovsky, A.; Müller, N.; Salzmann, S.; Rosenthal, T.; Kalthoff, H.; Trauzold, A.; Seher, A.; Henkler, F.; Kneitz, C.; et al. Soluble and transmembrane TNF-like weak inducer of apoptosis differentially activate the classical and noncanonical NF-κB pathway. J. Immunol. 2010, 185, 1593–1605. [Google Scholar] [CrossRef] [PubMed]
- Berindan-Neagoe, I.; Calin, G.A. Molecular pathways: MicroRNAs, cancer cells, and microenvironment. Clin. Cancer Res. 2014, 20, 6247–6253. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.A.; Muñoz-García, B.; Martín-Ventura, J.L.; Madrigal-Matute, J.; Orbe, J.; Páramo, J.A.; Ortega, L.; Egido, J.; Blanco-Colio, L.M. The CD163-expressing macrophages recognize and internalize TWEAK: Potential consequences in atherosclerosis. Atherosclerosis 2009, 207, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Bover, L.C.; Cardó-Vila, M.; Kuniyasu, A.; Sun, J.; Rangel, R.; Takeya, M.; Aggarwal, B.B.; Arap, W.; Pasqualini, R. A previously unrecognized protein-protein interaction between TWEAK and CD163: Potential biological implications. J. Immunol. 2007, 178, 8183–8194. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Kuroi, M.; Harashima, H. Influence of endosomal escape and degradation of α-galactosylceramide loaded liposomes on CD1d antigen presentation. Mol. Pharm. 2015, 12, 2791–2799. [Google Scholar] [CrossRef] [PubMed]
- Haselmann, V.; Kurz, A.; Bertsch, U.; Hübner, S.; Olempska-Müller, M.; Fritsch, J.; Häsler, R.; Pickl, A.; Fritsche, H.; Annewanter, F.; et al. Nuclear death receptor TRAIL-R2 inhibits maturation of let-7 and promotes proliferation of pancreatic and other tumor cells. Gastroenterology 2014, 146, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, M.J.; Lewis, E.E.; Shelden, E.A.; Somers, E.; Pavlic, R.; McCune, W.J.; Richardson, B.C. The apoptotic ligands TRAIL, TWEAK, and Fas ligand mediate monocyte death induced by autologous lupus T cells. J. Immunol. 2002, 169, 6020–6029. [Google Scholar] [CrossRef] [PubMed]
- Burkly, L.C.; Michaelson, J.S.; Zheng, T.S. TWEAK/Fn14 pathway: An immunological switch for shaping tissue responses. Immunol. Rev. 2011, 244, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Perron, M.P.; Landry, P.; Plante, I.; Provost, P. Detection of human Dicer and Argonaute 2 catalytic activity. Methods Mol. Biol. 2011, 725, 121–141. [Google Scholar] [PubMed]

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lambert, M.; Pépin, G.; Peralta-Zaragoza, O.; Matusiak, R.; Ly, S.; Landry, P.; Provost, P. TWEAK Negatively Regulates Human Dicer. Non-Coding RNA 2016, 2, 12. https://doi.org/10.3390/ncrna2040012
Lambert M, Pépin G, Peralta-Zaragoza O, Matusiak R, Ly S, Landry P, Provost P. TWEAK Negatively Regulates Human Dicer. Non-Coding RNA. 2016; 2(4):12. https://doi.org/10.3390/ncrna2040012
Chicago/Turabian StyleLambert, Marine, Geneviève Pépin, Oscar Peralta-Zaragoza, Raphaël Matusiak, Sophia Ly, Patricia Landry, and Patrick Provost. 2016. "TWEAK Negatively Regulates Human Dicer" Non-Coding RNA 2, no. 4: 12. https://doi.org/10.3390/ncrna2040012
APA StyleLambert, M., Pépin, G., Peralta-Zaragoza, O., Matusiak, R., Ly, S., Landry, P., & Provost, P. (2016). TWEAK Negatively Regulates Human Dicer. Non-Coding RNA, 2(4), 12. https://doi.org/10.3390/ncrna2040012