Abstract
Pinus massoniana Lamb. is an important, common afforestation and timber tree species in China. Species of Pestalotiopsis are well-known pathogens of needle blight. In this study, the five representative strains were isolated from needle blight from needles of Pi. massoniana in Nanjing, Jiangsu, China. Based on multi-locus phylogenetic analyses of the three genomic loci (ITS, TEF1, and TUB2), in conjunction with morphological characteristics, a new species, namely Pestalotiopsis jiangsuensis sp. nov., was described and reported. Pathogenicity tests revealed that the five representative strains of the species described above were pathogenic to Pi. massoniana. The study revealed the diversity of pathogenic species of needle blight on Pi. massoniana. This is the first report of needle blight caused by P. jiangsuensis on Pi. massoniana in China and worldwide. This provides useful information for future research on management strategies of this disease.
1. Introduction
Pinus massoniana Lamb. is the most widely distributed timber tree species with the largest afforestation area in China [1], which provides a large amount of timber, oleoresin [2], carbon storage [3], and ecological products [4], and also has potential biomedical properties [5]. However, Pi. massoniana was found dead at the top of needles in plantations in Nanjing, Jiangsu Province with a high incidence, which seriously threatened the economic and ecological value.
Many pathogens have been reported to damage Pi. massoniana in the world; for example, its forestry and pine forests were threatened by outbreaks of pine wilt disease (PWD) caused by Bursaphelenchus xylophilus (pinewood nematode; PWN) [6]. Damping-off and root rot disease caused by Fusarium oxysporum has been found in Pi. massoniana [7,8]. Pseudofusicoccum kimberleyense and Pse. violaceum can cause dead branch disease of Pi. massoniana [9]. Pestalotiopsis funerea affected the needles of young Pi. massoniana trees and caused them to gradually dry up and fall off [10]. In addition, insect–parasitic entomopathogenic fungi such as Penicillium citrinum, Purpurecillium lilacinum, and Fusarium spp. were also confirmed to be pathogenic to Pi. massoniana [11]. However, as an important economic tree species, the host–pathogen relationship of Pi. massoniana needs more studies, and additional pathogens may be found.
Pestalotiopsis species are widely distributed in the world as endophytes, plant pathogens, or saprobes [12,13,14,15,16,17], mainly in tropical and temperate regions and have a wide range of host plants [15,18,19]. Initially, the characteristics of conidia, such as color, size, and appendages, are the key to the identification of Pestalotiopsis and related genera [20,21]. Those taxonomic groups related to the genus Pestalotiopsis are also called pestalotioid fungi. Afterwards according to the relationship between conidial morphology and multi-locus phylogeny [14,19,22,23], Pestalotiopsis sensu lato was divided into three genera by Maharachchikumbura et al. (2014) [15]—Pestalotiopsis sensu stricto, Neopestalotiopsis, and Pseudopestalotiopsis. Three genera correspond to three types of conidia, conidia with light brown or olivaceous concolorous median cells (Pestalotiopsis sensu stricto), conidia with versicolorous median cells (Neopestalotiopsis), and conidia with dark-colored concolorous median cells (Pseudopestalotiopsis) [14,19,22,24]. Pestalotioid species identification remains a major challenge because of the conidia of overlap, and the classification is complex [22,25,26].
Needle blight caused by Pestalotiopsis is a common disease in young pine forests, and the disease is widely distributed and causes serious damage. For example, Pestalotiopsis funerea can infect Pinus tabulaeformis [27], Pi. taeda [28], Pi. massoniana [10], etc. and cause needle blight. Xu et al. (2017) [29] reported that the pathogen causing the needle blight of Pi. sylvestris was P. citrina. The disease began to occur in 1974 and became popular in 1980, and it has become the main coniferous disease of trees [30,31]. Needle blight not only reduced the stock of trees but even led to the death of trees, which greatly threatened forestry production [32,33,34].
In March 2023, the needles of Pi. massoniana with the characteristics of needle blight were collected in Nanjing, Jiangsu Province. The earlier identification of Pi. massoniana needle blight in a previous study was in a different geographical area [10]; thus, the main purpose of this study was to determine the pathogen of Pi. massoniana needle blight and its pathogenicity by Koch’s postulates.
2. Materials and Methods
2.1. Field Survey and Fungal Isolation
In March 2023, needle lesions were found on Pinus massoniana in Lishui District, Nanjing, Jiangsu, China. The entire planting area of the Pi. massoniana forest was about 1800 m2. The symptoms of trees were visually observed and the needles with the symptoms were collected. Five symptomatic Pi. massoniana trees were randomly sampled. After macroscopic and microscopic observation of the collected pine needles, the pine needle fragments at the intermediate area of the diseased and healthy portions were cut off, and the surface was disinfected in 70% ethanol for 30 s, in 1% NaClO for 90 s, and then washed in sterile water for 90 s three times. Pine needle fragments were dried on sterile filter paper and incubated on potato dextrose agar (PDA) in the dark at 25 °C for 3 days. The hyphal tips of fungi emerging from tissue pieces were transferred to new PDA to obtain pure cultures. The isolates were obtained from needle blight samples of Pi. massoniana.
2.2. Morphological Identification
Colony morphology and pigment production on PDA was observed after 7 days at 25 °C with a 12/12 h light/dark cycle and inspected daily for fungal sporulation. Acervuli and conidial masses were observed under a Zeiss stereo microscope (SteRo Discovery v20, Oberkochen, Germany). The micromorphological characteristics of five isolates were observed by Zeiss Axio Imager A2m microscope (Carl Zeiss, Oberkochen, Germany), such as shape, color, septation, appendages, and size of conidia, conidiophores, and acervuli.
2.3. Genomic DNA Extraction, PCR, and Sequencing
Fungal genomic DNA of fungi cultured on PDA for 5 days was extracted by the cetyltrimethylammonium bromide (CTAB) method, and three distinct DNA regions were amplified by polymerase chain reactions (PCR). Three genomic loci, including the internal transcribed spacer (ITS), the partial translation elongation factor 1-alpha (TEF1), and partial β-tubulin (TUB2), were amplified with primers ITS5/ITS4 [35], EF1-728F/EF1-986R [36], and T1/Bt-2b [37,38], respectively. The protocols for amplification are shown in Table 1. Each 50 μL PCR mixture consisted of 25 μL of Premix TaqTM (Takara Biomedical Technology Company Limited, Beijing, China), 19 μL of dd H2O, 2 μL of forwarding primer, 2 μL of reverse primer, and 2 μL of DNA template. PCR purification and sequencing were performed by Sangon Biotech (Shanghai) Co., Ltd. (Nanjing, China).

Table 1.
Reaction conditions used in PCR amplification and sequencing.
2.4. Phylogenetic Analyses
Sequences with similarity of the ITS sequences generated in the present study were searched with the BLAST program on GenBank (https://blast.ncbi.nlm.nih.gov/, accessed on 3 November 2023), and the reference sequences used in this study were obtained. Concatenated multi-locus data (ITS, TEF1, and TUB2) were used for phylogenetic analyses with maximum likelihood (ML) and Bayesian Inference (BI). Neopestalotiopsis protearum (CBS 114178) was designated as an outgroup. The DNA sequences were aligned with MAFFT ver. 7.313 [39] and adjusted with BioEdit ver. 7.0.9.0 [40]. Maximum likelihood (ML) analysis was conducted on the multi-locus alignments using IQtree ver. 1.6.8 [41] with the GTR + F + I + G4 replacement model and the bootstrap method with 1000 replications to assess clade stability. RA × ML bootstrap support values were set at ML ≥ 70. Bayesian inference was analyzed using MrBayes ver. 3.2.6 with the GTR + I + G + F model (2 parallel runs, 2,000,000 generations) according to Quaedvlieg et al. (2014) [42]. Bayesian posterior probability values were set at PP ≥ 0.90. The phylogenetic trees were created in Figtree ver. 1.4.4. (http://tree.bio.ed.ac.uk/software/figtree/, accessed on 2 December 2023).
2.5. Genealogical Concordance Phylogenetic Species Recognition Analyses
The phylogenetically related ambiguous species were analyzed using the Genealogical Concordance Phylogenetic Species Recognition (GCPSR) to determine the recombination level in closely related species by performing a pairwise homoplasy index (PHI) test according to the method described by Quaedvlieg et al. (2014) [42]. A PHI result below 0.05 (Φw < 0.05) indicated significant recombination in the dataset. The relationships between closely related species were visualized in splits graphs with the LogDet transformation and splits decomposition options.
2.6. Pathogenicity Test
In this study, 12 two-year-old healthy Pi. massoniana seedlings and the three isolates representing the highest isolation frequency of Pestalotiopsis species were selected to perform the pathogenicity tests: BM 1-1, BM 1-2, BM 1-3—Pestalotiopsis jiangsuensis sp. nov. The tested plants were taken from the GuDong Green Seedling Base in Hechi, Guangxi Province, China. Healthy needles of Pi. massoniana were injured with a sterile needle. One wound was made per pine needle and conidial suspension (106 conidia·mL−1) was sprayed on the wounds. Three plants were inoculated with each isolate, and the control was treated with sterile water. Inoculated seedlings and control seedlings were placed in a tent (1.5 × 1.2 × 1.5 m) with a humidifier (300 mL/h) to maintain RH 70%. The tent was placed in a greenhouse at 25 ± 2 °C and observed continuously for 10 days. All experiments were conducted three times.
3. Results
3.1. Disease Symptoms and Fungal Isolation
In March 2023, the incidence of needle blight of Pi. massoniana found in Nanjing, Jiangsu Province was ca. 60%, and the needle disease incidence of each Pi. massoniana was as high as 80%. The early symptom was a small yellow lesion at the needle tip, which extended from the needle tip downwards, and the lesion turned gray; a dark brown band encircled the needle at the junction with the healthy part (Figure 1A–C). Eventually the lesion area expanded until all the needles were necrotic. Ninety Pestalotiopsis strains were isolated and determined, based on the colony morphologies on PDA and ITS sequence blasting, with an isolation frequency of 90% (90/100). Five representative isolates (BM 1-1, BM 1-2, BM 1-3, BM 1-4, and BM 1-5) were selected for further study and deposited at the China Forestry Culture Collection Center (CFCC).

Figure 1.
Symptoms of needle blight on Pinus massoniana in the field (A–C).
3.2. Phylogenetic Analyses
The concatenated sequence dataset of ITS, TEF1, and TUB2 included the five representative isolates, 120 taxa, and one outgroup taxon (Neopestalotiopsis protearum CBS 114178) with a total of 1637 base pairs (1-554 for the TEF1, 555-1163 for ITS, and 1164-1637 for TUB2) including gaps were obtained. The hosts, locations, and GenBank accession numbers of Pestalotiopsis species used for phylogenetic analyses in this study were shown in Table 2. The tree topology of the phylogenetic tree of ML and BI systems was congruent, and the bootstrap support values of RA × ML greater than 70% and the Bayesian posterior probabilities greater than 0.90 were denoted at nodes. In the phylogenetic analyses, five isolates formed a separate clade (ML/BI = 100/1), which was clustered into a big branch with four ex-type strains with a significant support (ML/BI = 98/0.92: Pestalotiopsis foliicola CFCC 54440, P. pinicola KUMCC 19-0183, P. suae CGMCC 3.23546, and P. rosea MFLUCC 12-0258. Based on the three-locus phylogenetic analyses and morphology, five strains (BM 1-1, BM 1-2, BM 1-3, BM 1-4, and BM 1-5) were identified as a new species of Pestalotiopsis (Figure 2).
Table 2.
Host, Origin, and GenBank accession numbers of strains of Pestalotiopsis species used for phylogenetic analyses.
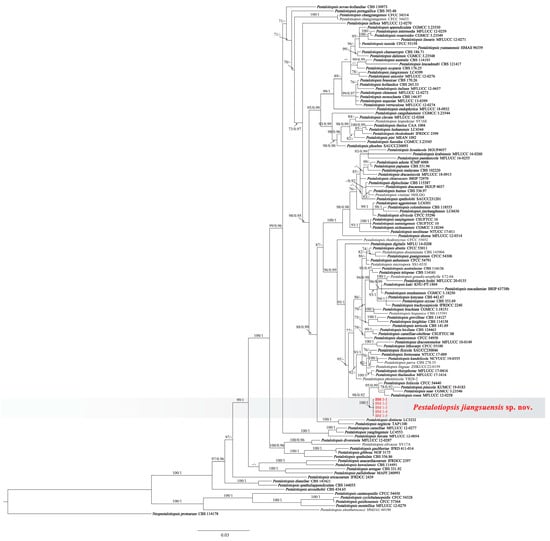
Figure 2.
Phylogenetic relationship of Pestalotiopsis jiangsuensis isolates: BM 1-1, BM 1-2, BM 1-3, BM 1-4, and BM 1-5, based on concatenated sequences of ITS, TEF1, and TUB2 genes/region. RA × ML bootstrap support values (ML ≥ 70) and Bayesian posterior probability values (PP ≥ 0.90) were shown at the nodes (ML/PP). Neopestalotiopsis protearum (CBS 114178) is used as an outgroup. Bar = 0.04 substitution per nucleotide position. The sequences from this study are in red. The ex-type strains are in bold.
Importantly, the PHI test of new species shows that no significant recombination (Φw = 0.071) events were observed between Pestalotiopsis sp. (undescribed taxon) and phylogenetically related species P. foliicola CFCC 54440, P. pinicola KUMCC 19-0183, P. suae CGMCC 3.23546, and P. rosea MFLUCC 12-0258 (Figure 3).
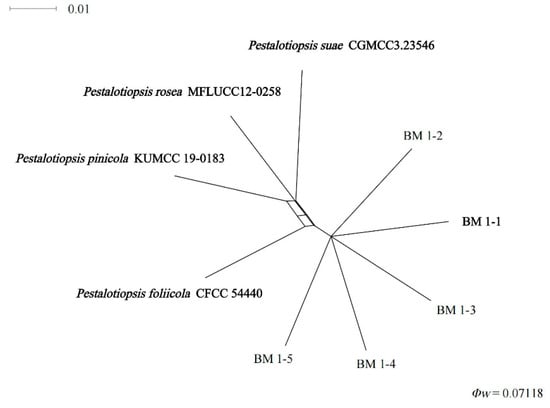
Figure 3.
Pairwise homoplasy index (PHI) test of Pestalotiopsis isolates: BM 1-1, BM 1-2, BM 1-3, BM 1-4, and BM 1-5 and closely related P. foliicola, P. pinicola, P. suae, and P. rosea using both LogDet transformation and splits decomposition. PHI test results (Φw) < 0.05 indicate significant recombination within the data set.
3.3. Taxonomy
Pestalotiopsis jiangsuensis Li-Hua Zhu, Hui Li, and D.W. Li, sp. nov. Figure 4
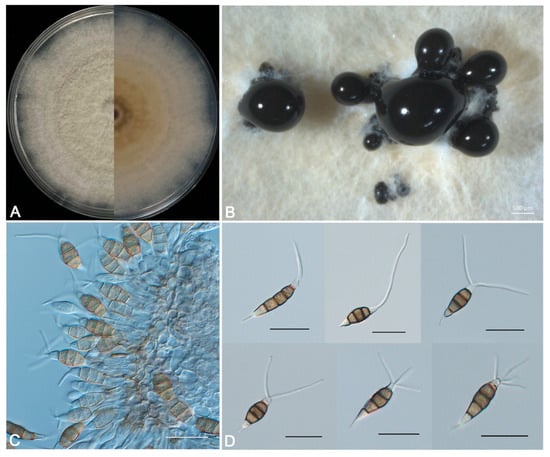
Figure 4.
Morphological characteristics of Pestalotiopsis jiangsuensis sp. nov. BM 1-1. (A) Fungal colony on PDA, 5 d growth from above (L) and below (R). (B) Conidiomata and conidial masses. (C) Conidiophores, conidiogenous cells, and conidia. (D) Conidia. Scale bars: (B) = 500 μm, (C,D) = 20 μm.
Index Fungorum No: IF 900494
Etymology: the epithet referring to the province where the holotype was collected.
Description: Sporadic black and gregarious conidiomata produced on PDA after 7 days under light at 25 °C, globose, semi-immersed, dark brown to black, up to 400 μm diam (Figure 4B); conidiophores indistinct and reduced to conidiogenous cells. Conidiogenous cells (4.5-) 7.0–12.8 (−15.3) × (2.4-) 3.3 –5.6 (−6.5) µm (11.4 ± 2.5 × 4.4 ± 0.9 µm, n = 30), hyaline, ampulliform or cylindrical, and sometimes slightly wide at the base (Figure 4C). Conidia phragmospores, (20.3-) 22.1–25.5 (−27.3) × (6.2-) 6.7–8.2 (−8.7) µm (23.4 ± 1.8 × 7.5 ± 0.5 µm, n = 30), fusoid, ellipsoid, straight to slightly curved, 4-septate (Figure 4D); basal cell hyaline, obconic, thin-walled, 3.5–5.9 μm long; three median cells (12.7-) 13.7–15.5 (−16.5) × (6.2-) 6.7–7.4 (−7.9) µm (14.2 ± 1.0 × 7.2 ± 0.5 µm, n = 30), doliiform, wall rugose, concolorous, brown, septa darker than the rest of the cell (second cell from the base 4.2–5.9 μm long; third cell 4.8–5.7 μm long; fourth cell 4.0–5.4 μm long); apical cell hyaline, smooth-walled, conic or trapezoid, tapering toward the apex, 2.6–4.4 μm long, with 1–4 tubular apical appendages (mostly 2 and very few 4), arising from the apical crest, unbranched, filiform, 8.7–23.4 μm long; basal appendage single, tubular, unbranched, centric, 1.4–6.3 μm long.
Culture characteristics: Colonies on PDA flat with sparse aerial mycelia on the surface after 7 d at 25 °C, edge undulate, pale honey-colored, and reverse pale brown in the center and pale luteous margin (Figure 4A).
Holotype: China, Jiangsu province, Nanjing city, Lishui district, Baima National Agricultural Science and Technology Park, 119°10′44″ N, 31°36′28″ E (DMS), isolated from needles of Pinus massoniana, 1 March 2023, Hui Li, holotype CFCC 59538. Holotype is a living specimen being maintained via lyophilization at the China Forestry Culture Collection Center (CFCC), Chinese Academy of Forestry, Beijing, China, and ex-type BM 1-1 is stored at Forest Pathology Laboratory, Nanjing Forestry University.
Habitat and host: On needles of Pinus massoniana with needle blight.
Known distribution: Nanjing, Jiangsu Province, China.
Additional specimens examined: China, Jiangsu province, Nanjing city, Lishui district, Baima National Agricultural Science and Technology Park, 119°10′44″ N, 31°36′28″ E (DMS), isolated from needles of Pinus massoniana, 1 March 2023, Hui Li, cultures: CFCC 59539 (=BM 1-2), CFCC 59540 (=BM 1-3), CFCC 59541 (=BM 1-4), and CFCC 59542 (=BM 1-5).
Notes: Pestalotiopsis jiangsuensis is a species often having one to four tubular apical appendages, which are phylogenetically and morphologically well distinguished from P. foliicola CFCC 54440, P. pinicola KUMCC 19-0183, P. suae CGMCC 3.23546, and P. rosea MFLUCC 12-0258. Although the five strains studied are a sister clade of P. foliicola CFCC 54440, P. pinicola KUMCC 19-0183, P. suae CGMCC 3.23546, and P. rosea MFLUCC 12-0258, the number of apical appendages is quite different. Pestalotiopsis folicola, P. pinicola and P. suae have two to three apical appendages; P. rosea has one to three tubular apical appendages, and some appendages are branched. The strains in this study have one to four apical appendages, and the appendages are unbranched.
Pestalotiopsis funerea has two to four apical appendages, and Pestalotiopsis lawsoniae has two apical appendages. They also have differences with P. jiangsuensis. In addition, P. funerea has a longer basal appendage than that of P. jiangsuensis (5–7) µm vs. (1.4–6.3) µm [43,44].
3.4. Pathogenicity Test
In the experiment of Koch’s postulates, the three representative isolates were pathogenic to Pi. massoniana needles. The development of disease symptoms was observed during a 10-day period. At 5 d, all the Pestalotiopsis jiangsuensis isolates developed gray to gray-brown lesions on wounded needles of Pi. massoniana (Figure 5B–D). At 10 d, the lesion expanded, and in severe cases, the whole needle was necrotic (Figure 5F–H). No symptoms developed on the needles of the control (Figure 5A,E). In this study, the pathogenicity of Pestalotiopsis jiangsuensis is strong; for example, the lesions spread almost to the whole needle after 10 days. It may also relate to its high isolation rate. Pestalotiopsis jiangsuensis was successfully re-isolated from 100% of the inoculated plants and identified based on morphological features and phylogenetic analysis of ITS. Thus, Koch’s postulates had been fulfilled.

Figure 5.
Pathogenicity of representative isolates of Pestalotiopsis jiangsuensis sp. nov. (BM 1-1, BM 1-2, and BM 1-3) on Pinus massoniana. (A) No symptoms were observed on control pine needles treated with sterile water after 5 days. (B–D) Symptoms on pine needles inoculated with conidial suspensions of BM 1-1, BM 1-2, and BM 1-3 after 5 days, respectively. (E) No symptoms observed on control pine needles treated with sterile water after 10 days. (F–H) Symptoms on pine needles inoculated with conidial suspensions of BM 1-1, BM 1-2, and BM 1-3 after 10 days.
4. Discussion
Pestalotiopsis was established by Steyeart (1949) [45] and typified with Pestalotiopsis guepinii Steyaert. Pestalotiopsis sensu lato was classified based on conidia with five-celled, the middle three intermediate colored cells, and hyaline end cells. After that, its taxonomic characteristics gradually changed into conidia spindle-shaped, with five-celled, with colorless or nearly colorless cells at both ends, dark cells in the middle, and one or more branched or unbranched apical appendages arising from the apical cell, with or without basal stalk [20,21,46,47]. The excessive overlap of conidia makes it difficult to identify Pestalotioid species only by morphological characteristics [19]. Although some additional taxonomic features can also be used as the basis for the identification of Pestalotiopsis—such as the pigmentation of median cells, which is an important character to distinguish Pestalotiopsis funerea and P. triseta [23,48]—there are still great limitations [17,22,49]. However, the application of molecular data in the identification of Pestalotiopsis species has greatly improved the accuracy and credibility [22,23,26,50,51]. Pestalotiopsis sensu lato was segregated into three genera by Maharachchikumbura et al. (2014) [15] as Pestalotiopsis sensu stricto, Neopestalotiopsis, and Pseudopestalotiopsis, based on both morphological characteristics and phylogenetic analyses. Gu et al. (2022) [17] identified six new Pestalotiopsis species from Rhododendron, based on phylogenetic analyses of combined ITS, TEF1, and TUB2 genes/region along with morphological characteristics. Maharachchikumbura et al. (2012) [14] identified 23 species of Pestalotiopsis from different host plants in China, including 14 new species, based on phylogenetic analysis of ITS, TEF1, and TUB2 genes/region and morphology. More importantly, concatenating ITS, TUB2, and TEF1 sequences can provide better identification information for Pestalotiopsis [14,52].
The Global Biodiversity Information Facility (https://www.gbif.org/, accessed on 24 November 2023) displays 9320 records of Pestalotiopsis from all over the world, including years and coordinates [53]. The data show that most of them are distributed in Australia, Brazil, China, and the United States. Pestalotiopsis as a plant pathogen has a wide range of symptoms on the hosts, such as withering or chlorosis of leaves, dead shoots or tips, and canker [15]. In Pinus spp., it may be characterized by shoot blight, trunk necrosis, needle blight, and pinecone decay [54]. It is not uncommon that a species of Pestalotiopsis was successfully isolated from needles of Pinus species [34]. For example, Pestalotiopsis neglecta and P. citrina isolated from Pi. sylvestris can cause the needles to turn yellow partially or completely and even cause death of the trees [29,34]. Pestalotiopsis bessey isolated from Pi. halenpesis can cause the entire needles to turn dark gray-brown and eventually cause the death of the trees [55,56]. Pestalotiopsis pini isolated from Pi. Pinea can cause the needles and branches to wither, trunk necrosis, and pinecone rot [54]. Pestalotiopsis is also an endophytic fungus of some Pinus spp., such as P. funerea, and it was isolated from the healthy needles of Pi. pinaster [57].
Interestingly, the pathogen of Pi. massoniana needle blight isolated in a previous study was P. funerea [58], but the pathogen obtained in this study was Pestalotiopsis jiangsuensis, which indicated that the pathogens of the same genus on the same host were diverse. Silva et al. [54] isolated P. disseminata and P. pini from Pi. Pinea, and their results also confirmed this view. Similarly, the same species of Pestalotiopsis can be found on different plant hosts, such as P. funereal, which was isolated from Pi. tabulaeformis, Pi. taeda, and Pi. massoniana [10,27,28]. Pestalotiopsis chamaeropis was isolated from Quercus sp., Castanopsis sp., and Camellia sp. [15,49,59]. However, in the current study the samples were only collected from one site. In future research, the investigation areas should be expanded to study fungal diversity on Pinus spp. and related ecological functions.
5. Conclusions
In this study, we examined five strains, all of which were pathogenic to Pi. massoniana. Combined with morphology, multi-locus phylogenetic analyses, and GCPSR principle, these five strains were identified to be a new species to science, Pestalotiopsis jiangsuensis. This is the first report of needle blight caused by P. jiangsuensis on Pi. massoniana in China and worldwide, and it will provide useful information for future studies on all the phytopathological perspectives of this fungus and the management strategies of this newly emerged disease.
Author Contributions
Conceptualization, L.-H.Z.; methodology, H.L., J.-Y.X. and Y.-Q.B. software, H.L.; validation, H.L.; formal analysis, H.L.; investigation, H.L., J.-Y.X. and Y.-Q.B.; resources, L.-H.Z.; data curation, H.L.; writing—original draft preparation, H.L.; writing—review and editing, D.-W.L.; visualization, H.L. and B.-Y.P.; supervision, D.-W.L.; project administration, L.-H.Z.; funding acquisition, L.-H.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by National Key R & D Program of China (2022YFD1401005), and the National Natural Science Foundation of China (grant number 31971659).
Institutional Review Board Statement
Not applicable for studies not involving humans or animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors would like to thank those who provided assistance and advice for this study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Lu, J.Y.; Chen, H.; Yang, Z.Q.; Sun, S.; Luo, Q.F.; Xie, J.K.; Tan, J.H. Physiological and molecular mechanisms of the response of roots of Pinus massoniana Lamb. to low-temperature stress. Front. Plant Sci. 2022, 13, 954324. [Google Scholar] [CrossRef]
- Liu, Q.H.; Zhou, Z.C.; Wei, Y.C.; Shen, D.Y.; Feng, Z.P.; Hong, S.P. Genome-wide identification of differentially expressed genes associated with the high yielding of oleoresin in secondary xylem of masson pine (Pinus massoniana Lamb.) by transcriptomic analysis. PLoS ONE 2015, 10, e0132624. [Google Scholar] [CrossRef]
- Kang, B.; Liu, S.R.; Zhang, G.G.; Chang, J.G.; Wen, Y.G.; Ma, J.M.; Hao, W.F. Carbon accumulation and distribution in Pinus massoniana and Cunninghamia lanceolata mixed forest ecosystem in Daqingshan, Guangxi, China. Acta Ecol. Sin. 2006, 26, 1320–1327. [Google Scholar] [CrossRef]
- Yang, Z.; Xia, H.; Tan, J.; Feng, Y.; Huang, Y. Selection of superior families of Pinus massoniana in southern China for large-diameter construction timber. J. For. Res. 2020, 31, 475–484. [Google Scholar] [CrossRef]
- He, L.; Zhao, C.; Yan, M.; Zhang, L.Y.; Xia, Y.Z. Inhibition of P-glycoprotein function by procyanidine on blood-brain barrier. Phytother. Res. 2009, 23, 933–937. [Google Scholar] [CrossRef]
- Xie, Y.; Liu, B.; Gao, K.; Zhao, Y.; Li, W.; Deng, L.; Zhou, Z.; Liu, Q. Comprehensive analysis and functional verification of the Pinus massoniana NBS-LRR gene family involved in the resistance to Bursaphelenchus xylophilus. Int. J. Mol. Sci. 2023, 24, 1812. [Google Scholar] [CrossRef]
- Luo, X.; Yu, C. First report of damping-off disease caused by Fusarium oxysporum in Pinus massoniana in China. J. Plant Dis. Prot. 2020, 127, 401–409. [Google Scholar] [CrossRef]
- Yu, C.; Luo, X. Trichoderma koningiopsis controls Fusarium oxysporum causing damping-off in Pinus massoniana seedlings by regulating active oxygen metabolism, osmotic potential, and the rhizosphere microbiome. Biol. Control 2020, 150, 104352. [Google Scholar] [CrossRef]
- Li, G.Q.; Wu, W.X.; Lu, L.Q.; Chen, B.Y.; Chen, S.F. Characterization of Pseudofusicoccum species from diseased plantation-grown Acacia mangium, Eucalyptus spp., and Pinus massoniana in Southern China. Pathogens 2023, 12, 574. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.X.; Pan, F.Y.; Li, D.X. Regularity of outbreak and control techniques of Pinus massoniana cercospora needle blight. J. Zhejiang For. Sci. Technol. 2002, 4, 64–65+84. [Google Scholar]
- Wu, S.X.; Wu, J.; Wang, Y.; Qu, Y.F.; He, Y.; Wang, J.Y.; Cheng, J.H.; Zhang, L.Q.; Cheng, C.H. Discovery of entomopathogenic fungi across geographical regions in southern China on pine sawyer beetle Monochamus alternatus and implication for multi-pathogen vectoring potential of this beetle. Front. Plant Sci. 2022, 13, 1061520. [Google Scholar] [CrossRef]
- Sun, H.T.; Cao, R.B. Identification of Pestalotiopsis parasitized on fruit crops. Acta Agric. Zhejiangensis 1990, 16, 179–185. (In Chinese) [Google Scholar]
- Xu, L.; Kusakari, S.; Hosomi, A. Postharvest disease of grape caused by Pestalotiopsis species. Jpn. J. Phytopathol. 1999, 65, 305–311. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Guo, L.D.; Cai, L.; Chukeatirote, E.; Wu, W.P.; Sun, X.; Hyde, K.D. A multi-locus backbone tree for Pestalotiopsis, with a polyphasic characterization of 14 new species. Fungal Divers. 2012, 56, 95–129. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Groenewald, J.Z.; Xu, J.; Crous, P.W. Pestalotiopsis revisited. Stud. Mycol. 2014, 79, 121–186. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Zhang, W.; Liu, M.; Maharachchikumbura, S.S.N.; Zhou, Y.; Huang, J.B.; Nilthong, S.; Wang, Z.Y.; Li, X.H.; Yan, J.Y.; et al. Identification and characterization of Pestalotiopsis-like fungi related to grapevine diseases in China. Fungal Biol. 2015, 119, 348–361. [Google Scholar] [CrossRef] [PubMed]
- Gu, R.; Bao, D.F.; Shen, H.W.; Su, X.J.; Li, Y.X.; Luo, Z.L. Endophytic Pestalotiopsis species associated with Rhododendron in Cangshan Mountain, Yunnan Province, China. Front. Microbiol. 2022, 13, 1016782. [Google Scholar] [CrossRef]
- Bate-Smith, E.C.; Metcalfe, C.R. Leucanthocyanins. 3. The nature and systematic distribution of tannin in dicotyledonous plants. Bot. J. Linn. Soc. 1957, 55, 669–705. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Guo, L.D.; Chukeatirote, E.; Ekachai, C.; Bahkali, A.H.; Hyde, K.D. Pestalotiopsis-morphology, phylogeny, biochemistry and diversity. Fungal Divers. 2011, 50, 167–187. [Google Scholar] [CrossRef]
- Guba, E.F. Monograph of Pestalotia and Monochaetia; Harvard University Press: Cambridge, MA, USA, 1961. [Google Scholar]
- Sutton, B.C. The Coelomycetes. Fungi Imperfecti with Pycnidia, Acervuli and Stromata; Commonwealth Mycological Institute: Surrey, UK, 1980. [Google Scholar] [CrossRef]
- Jeewon, R.; Liew, E.C.Y.; Simpson, J.A.; Hodgkiss, I.J.; Hyde, K.D. Phylogenetic significance of morphological characters in the taxonomy of Pestalotiopsis species. Mol. Phylogenet. Evol. 2003, 27, 372–383. [Google Scholar] [CrossRef]
- Hu, H.L.; Jeewon, R.; Zhou, D.Q.; Zhou, T.X.; Hyde, K.D. Phylogenetic diversity of endophytic Pestalotiopsis species in Pinus armandii and Ribes spp.: Evidence from rDNA and β-tubulin gene phylogenies. Fungal Divers. 2007, 24, 1–22. [Google Scholar]
- Liu, A.R.; Chen, S.C.; Wu, S.Y.; Xu, T.; Guo, L.D.; Jeewon, R.; Wei, J.G. Cultural studies coupled with DNA based sequence analyses and its implication on pigmentation as a phylogenetic marker in Pestalotiopsis taxonomy. Mol. Phylogenet. Evol. 2010, 57, 528–535. [Google Scholar] [CrossRef]
- Jeewon, R.; Liew, E.C.Y.; Hyde, K.D. Phylogenetic relationships of Pestalotiopsis and allied genera inferred from ribosomal DNA sequences and morphological characters. Mol. Phylogenet. Evol. 2002, 25, 378–392. [Google Scholar] [CrossRef]
- Tejesvi, M.V.; Tamhankar, S.A.; Kini, K.R.; Rao, V.S.; Prakash, H.S. Phylogenetic analysis of endophytic Pestalotiopsis species from ethnopharmaceutically important medicinal trees. Fungal Divers. 2009, 38, 167–183. [Google Scholar]
- Huang, Q.; He, J.S. Identification and biological characteristics of pathogen of Pinus tabulaeformis blight. J. Sichuan For. Sci. Technol. 2000, 21, 28–30. [Google Scholar] [CrossRef]
- Huang, G.F. The best method of comprehensive control of loblolly pine blight and defoliation by exploring suspected pine wood nematode disease. Flowers 2018, 12, 364–368. [Google Scholar]
- Xu, Y.; Ren, H.T.; Wang, P.; Zhao, H.X.; Song, Y.Q.; Yu, Q.F.; Liu, X.F. Pathogenic fungi of Pinus sylvestris var. mongolica red blight. J. West China For. Sci. 2017, 46, 91–95. [Google Scholar] [CrossRef]
- Chen, S.C. Research progress and prospect of forest diseases in China. For. Pest Dis. 1997, 4, 38–40. [Google Scholar]
- Bednářová, M.; Dvořák, M.; Janoušek, J.; Jankovský, L. Other foliar diseases of coniferous trees. In Infectious Forest Diseases; Gonthier, P., Nicolotti, G., Eds.; CABI: Wallingford, UK, 2013; pp. 458–487. [Google Scholar]
- Orlikowski, L.B.; Ptaszek, M.; Warabieda, W. Occurrence and harmfulness of Pestalotiopsis funerea to ornamental coniferous plants. Prog. Plant Prot. 2014, 54, 25–30. [Google Scholar]
- Hu, R.R.; Liang, J.; Xie, X.; Zhang, Y.J.; Zhang, X.Y. Incidence of pine needle blight and its relationship with site factors of Japanese red pine forests in the Kunyushan Mountains, East China. Glob. Ecol. Conserv. 2020, 22, e00922. [Google Scholar] [CrossRef]
- Monteiro, P.; Gonçalves, M.F.M.; Pinto, G.; Silva, B.; Martín-García, J.; Diez, J.J.; Alves, A. Three novel species of fungi associated with pine species showing needle blight-like disease symptoms. Eur. J. Plant Pathol. 2022, 162, 183–202. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Quaedvlieg, W.; Binder, M.; Groenewald, J.Z.; Summerell, B.A.; Carnegie, A.J.; Burgess, T.I.; Crous, P.W. Introducing the consolidated species concept to resolve species in the Teratosphaeriaceae. Persoonia 2014, 33, 1–40. [Google Scholar] [CrossRef]
- Mundkur, B.B.; Kheswalla, K.F. Indian and Burman species of the d=genera Pestalotia and Monochaetia. Mycologia 1942, 34, 308–317. [Google Scholar] [CrossRef]
- Yuan, C. Study on Molecular Detection Technologies for Two Forest Fungus Diseases Pathogen in Sichuan. Master’s Thesis, Sichuan Agricultural University, Ya’an, China, 2012. [Google Scholar]
- Steyaert, R.L. Contribution à l’étude monographique de Pestalotia de Not. et Monochaetia Sacc. (Truncatella gen. nov. et Pestalotiopsis gen. nov.). Bull. Du Jard. Bot. De L’état A Brux. 1949, 19, 285–347. [Google Scholar] [CrossRef]
- Nag Raj, T.R. Coelomycetous Anamorphs with Appendage-Bearing Conidia; Mycologue Publications: Waterloo, ON, Canada, 1993. [Google Scholar]
- Ge, Q.X.; Chen, Y.X.; Xu, T. Flora Fungorum Sinicorum, V. 38 Pestalotiopsis; Science Press: Beijing, China, 2009. [Google Scholar]
- Griffiths, D.A.; Swart, H.J. Conidial structure in two species of Pestalotiopsis. Trans. Br. Mycol. Soc. 1974, 62, 295. [Google Scholar] [CrossRef]
- Liu, F.; Hou, L.; Raza, M.; Cai, L. Pestalotiopsis and allied genera from Camellia, with description of 11 new species from China. Sci. Rep. 2017, 7, 866. [Google Scholar] [CrossRef]
- Jeewon, R.; Liew, E.C.Y.; Hyde, K. Phylogenetic evaluation of species nomenclature of Pestalotiopsis in relation to host association. Fungal Divers. 2004, 17, 39–55. [Google Scholar]
- Liu, A.R.; Xu, T.; Guo, L.D. Molecular and morphological description of Pestalotiopsis hainanensis sp. nov., a new endophyte from a tropical region of China. Fungal Divers. 2007, 24, 23–36. [Google Scholar]
- Li, L.L.; Yang, Q.; Li, H. Morphology, phylogeny, and pathogenicity of Pestalotioid species on Camellia oleifera in China. J. Fungi 2021, 7, 1080. [Google Scholar] [CrossRef]
- Zhang, Z.X.; Liu, X.Y.; Zhang, X.G.; Meng, Z. Morphological and phylogenetic analyses reveal two new species and a new record of Phyllosticta (Botryosphaeriales, Phyllostictaceae) from Hainan, China. MycoKeys 2022, 91, 1–23. [Google Scholar] [CrossRef]
- Silva, A.C.; Diogo, E.; Henriques, J.; Ramos, A.P.; Sandoval-Denis, M.; Crous, P.W.; Bragança, H. Pestalotiopsis pini sp. nov., an emerging pathogen on stone pine (Pinus pinea L.). Forests 2020, 11, 805. [Google Scholar] [CrossRef]
- Bezos, D.; Martínez-Álvarez, P.; Sanz-ros, A.V.; Martín-García, J.; Fernandez, M.M.; Diez, J.J. Fungal communities associated with bark beetles in Pinus radiata plantations in Northern Spain affected by pine pitch canker, with special focus on Fusarium species. Forests 2018, 9, 698. [Google Scholar] [CrossRef]
- Chen, J.; Hao, X.; Liu, X.; Ma, L. First report of Pestalotiopsis neglecta causing black spot needle blight of Pinus sylvestris var. mongolica in China. Plant Dis. 2020, 104, 1545. [Google Scholar] [CrossRef]
- Martínez-Álvarez, P.; Martín-García, J.; Rodríguez-Ceinós, S.; Diez, J.J. Monitoring endophyte populations in pine plantations and native oak forests in Northern Spain. For. Syst. 2012, 21, 373–382. [Google Scholar] [CrossRef]
- Qiu, D.X.; Tan, S.B.; Wu, J.C. Preliminary study on of Pinus massoniana. Sci. Silvae Sin. 1980, 3, 203–207. [Google Scholar]
- Jiang, N.; Voglmayr, H.; Xue, H.; Piao, C.G.; Li, Y. Morphology and phylogeny of Pestalotiopsis (Sporocadaceae, Amphisphaeriales) from Fagaceae leaves in China. Microbiol. Spectr. 2022, 10, e03272-22. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).