

Identification of Exoenzymes Secreted by Entomopathogenic Fungus Beauveria pseudobassiana RGM 2184 and Their Effect on the Degradation of Cocoons and Pupae of Quarantine Pest Lobesia botrana
 ,
,
Abstract
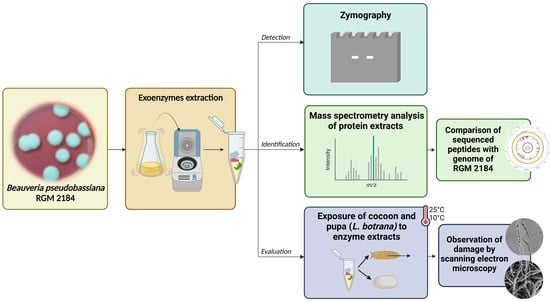
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Biological Materials and Culture Media
2.2. Extraction of Exoenzymes
2.3. Preliminary Evaluation of the Presence of Exoenzymes in Protein Extracts
2.4. Detection of Exoproteases, Exolipases, and Exochitinases
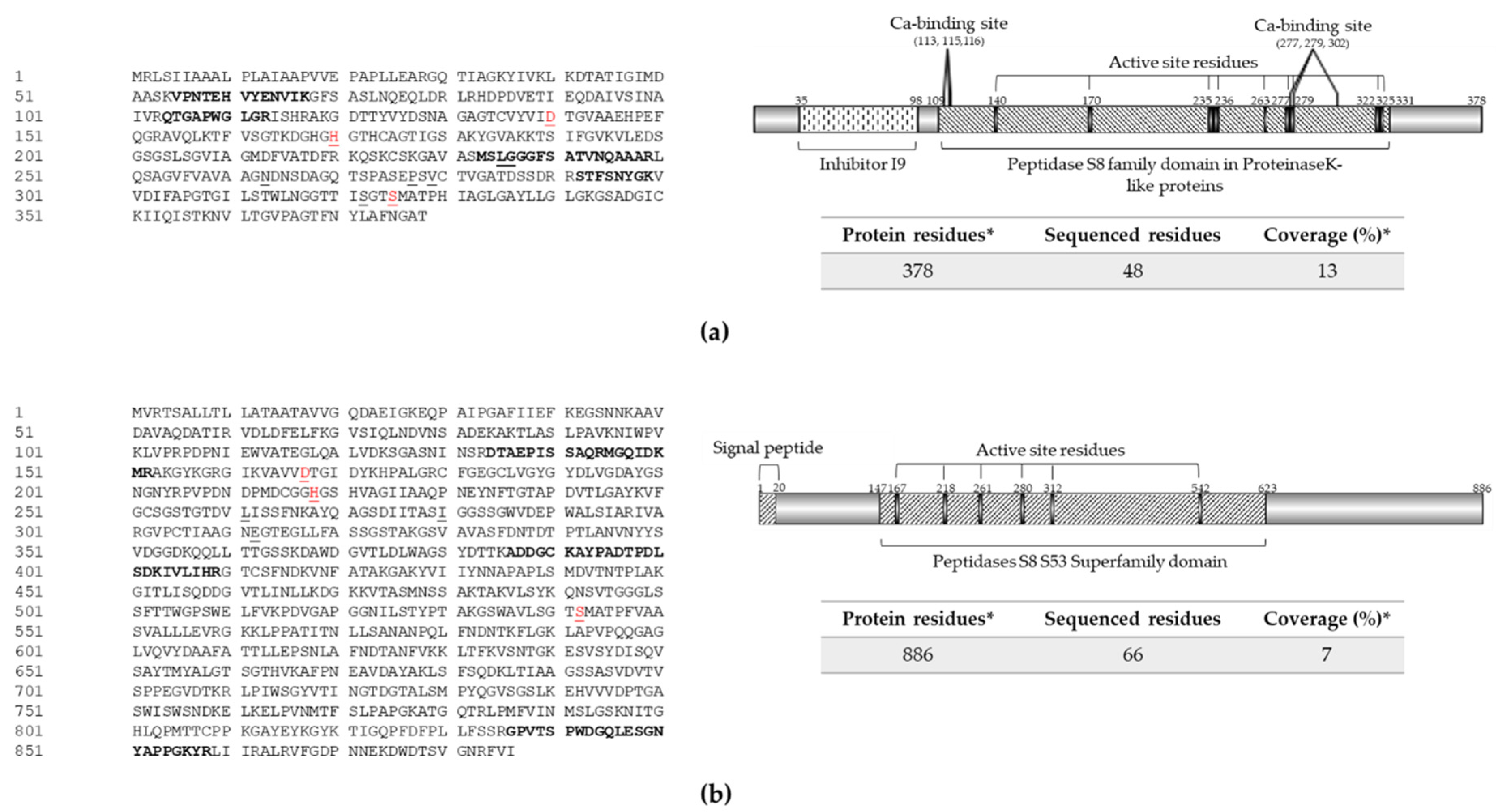
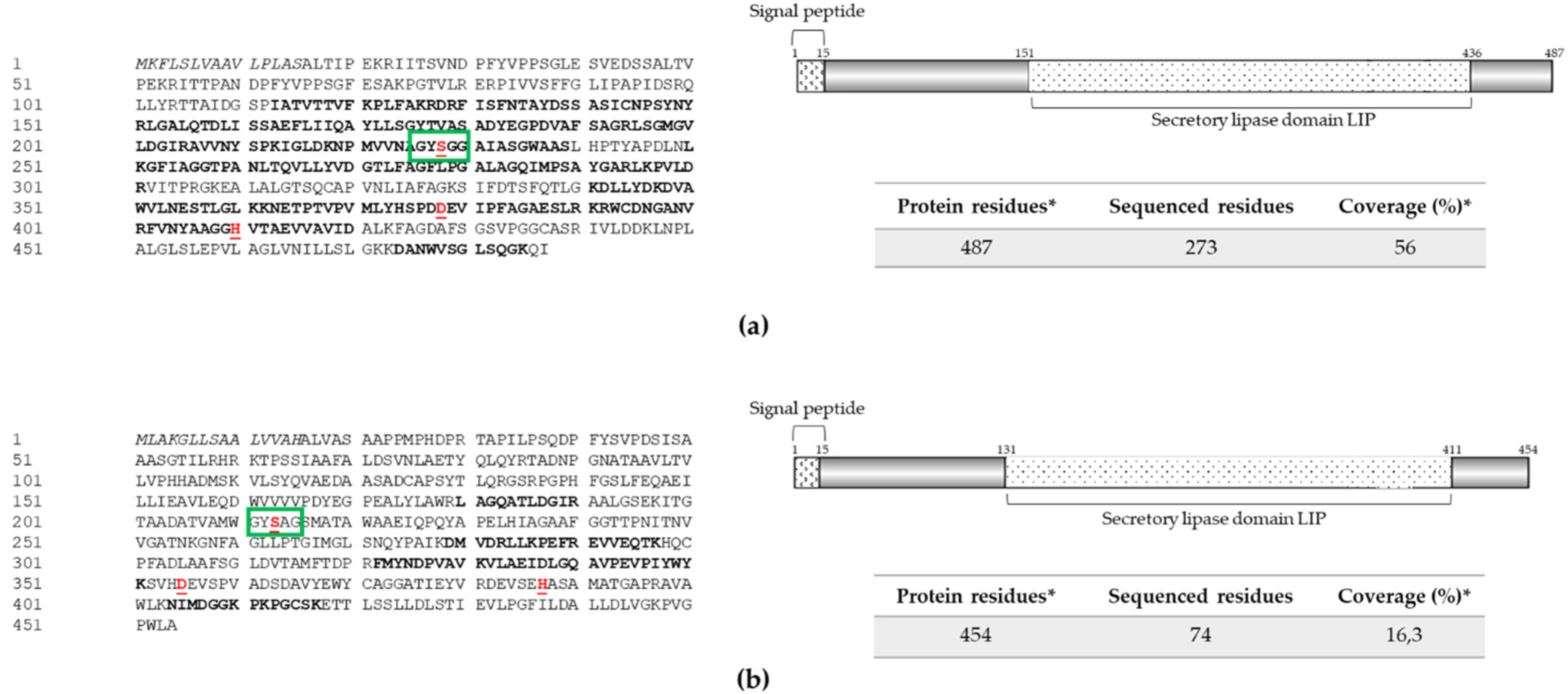
2.5. Mass Spectrometry Analysis of Protein Extracts A8 and M2
2.6. Comparative Analysis of Exoenzyme Sequences
2.7. Comparative Analysis of Amino Acid Composition
2.8. Evaluation of Exoenzymes on L. botrana Cocoon and Pupae
3. Result and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, S.; Shukla, H.; Prasad, R.; Kumar, S.; Shukla, S. Efficacy of Entomopathogenic Fungi as Green Pesticides: Current and Future Prospects. In Microorganisms for Green Revolution; Microorganisms for Sustainability; Springer: Singapore, 2017; Volume 6, ISBN 978-981-10-6241-4. [Google Scholar]
- Prabha, S.; Yadav, A.; Kumar, A.; Yadav, A.; Yadav, H.K.; Kumar, S.; Kumar, R. Biopesticides—An alternative and eco-friendly source for the control of pests in agricultural crops. Plant Arch. 2016, 16, 902–906. [Google Scholar]
- Butt, T.; Coates, C.; Dubovskiy, I.; Ratcliffe, N. Chapter Nine—Entomopathogenic Fungi: New Insights into Host–Pathogen Interactions. In Advances in Genetics; Lovett, B., St. Leger, R.J., Eds.; Academic Press: San Diego, CA, USA; London/Oxford, UK, 2016; pp. 1–58. [Google Scholar]
- Paschapur, A.; Subbanna, A.R.N.S.; Kumar, A.; Jeevan, B.; Stanley, J.; Rajashekhar, H.; Mishra, K.K. Unraveling the Importance of Metabolites from Entomopathogenic Fungi in Insect Pest Management. In Microbes for Sustainable Insect Pest Management; Khan, M.A., Ahmad, W., Eds.; Springer: Cham, Switzerland, 2021; pp. 89–120. ISBN 978-3-030-67231-7. [Google Scholar]
- Sain, S.K.; Monga, D.; Hiremani, N.S.; Nagrale, D.T.; Kranthi, S.; Kumar, R.; Kranthi, K.R.; Tuteja, O.P.; Waghmare, V.N. Evaluation of bioefficacy potential of entomopathogenic fungi against the whitefly (Bemisia tabaci Genn.) on cotton under polyhouse and field conditions. J. Invertebr. Pathol. 2021, 183, 107618. [Google Scholar] [CrossRef] [PubMed]
- Wilcken, C.F.; Dal Pogetto, M.; Lima, A.; Soliman, E.P.; Fernandes, B.V.; da Silva, I.M.; Zanuncio, A.; Barbosa, L.R.; Zanuncio, J.C. Chemical vs. entomopathogenic control of Thaumastocoris peregrinus (Hemiptera: Thaumastocoridae) via aerial application in eucalyptus plantations. Sci. Rep. 2019, 9, 9416. [Google Scholar] [CrossRef]
- Altimira, F.; Arias-Aravena, M.; Jian, L.; Real, N.; Correa, P.; González, C.; Godoy, S.; Castro, J.F.; Zamora, O.; Vergara, C.; et al. Genomic and Experimental Analysis of the Insecticidal Factors Secreted by the Entomopathogenic Fungus Beauveria pseudobassiana RGM 2184. J. Fungi 2022, 8, 253. [Google Scholar] [CrossRef] [PubMed]
- Lopes, R.B.; Martins, I.; Souza, D.A.; Faria, M. Influence of some parameters on the germination assessment of mycopesticides. J. Invertebr. Pathol. 2013, 112, 236–242. [Google Scholar] [CrossRef] [PubMed]
- de Carolina Sánchez-Pérez, L.; Barranco-Florido, J.E.; Rodríguez-Navarro, S.; Cervantes-Mayagoitia, J.F. Enzymes of Entomopathogenic Fungi, Advances and Insights. In Advances in Enzyme Research; Scientific Research Publishing: Wuhan, China, 2014; Volume 2. [Google Scholar]
- Petrisor, C.; Stoian, G. The role of hydrolytic enzymes produced by entomopathogenic fungi in pathogenesis of insects mini review. Rom. J. Plant Prot. 2017, 10, 66–72. [Google Scholar]
- Litwin, A.; Nowak, M.; Różalska, S. Entomopathogenic fungi: Unconventional applications. Rev. Environ. Sci. Biotechnol. 2020, 19, 23–42. [Google Scholar] [CrossRef]
- Mondal, S.; Baksi, S.; Koris, A.; Vatai, G. Journey of enzymes in entomopathogenic fungi. Pac. Sci. Rev. 2016, 18, 85–99. [Google Scholar] [CrossRef]
- Altimira, F.; De La Barra, N.; Godoy, P.; Roa, J.; Godoy, S.; Vitta, N.; Tapia, E. Lobesia botrana: A Biological Control Approach with a Biopesticide Based on Entomopathogenic Fungi in the Winter Season in Chile. Insects 2021, 13, 8. [Google Scholar] [CrossRef]
- Cheong, P.C.; Glare, T.R.; Rostás, M.; Haines, S.; Brookes, J.J.; Ford, S. Lack of involvement of chitinase in direct toxicity of Beauveria bassiana cultures to the aphid Myzus persicae. J. Invertebr. Pathol. 2020, 169, 107276. [Google Scholar] [CrossRef]
- Safavi, S.A.; Shah, F.A.; Pakdel, A.K.; Reza, G.; Bandani, A.R.; Butt, T.M. Effect of nutrition on growth and virulence of the entomopathogenic fungus Beauveria bassiana. FEMS Microbiol. Lett. 2007, 270, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Lohse, R.; Jakobs-Schönwandt, D.; Patel, A.V. Screening of liquid media and fermentation of an endophytic Beauveria bassiana strain in a bioreactor. AMB Express 2014, 4, 47. [Google Scholar] [CrossRef] [PubMed]
- Secades, P.; Guijarro, J.A. Purification and characterization of an extracellular protease from the fish pathogen Yersinia ruckeri and effect of culture conditions on production. Appl. Environ. Microbiol. 1999, 65, 3969–3975. [Google Scholar] [CrossRef] [PubMed]
- Izrael-Zivkovic, L.T.; Gojgic-Cvijovic, G.Đ.; Gopcevic, K.R.; Vrvic, M.M.; Karadzic, I.M. Enzymatic characterization of 30 kDa lipase from Pseudomonas aeruginosa ATCC 27853. J. Basic Microbiol. 2009, 49, 452–462. [Google Scholar] [CrossRef]
- Kim, J.S.; Roh, J.Y.; Choi, J.Y.; Wang, Y.; Shim, H.J.; Je, Y.H. Correlation of the aphicidal activity of Beauveria bassiana SFB-205 supernatant with enzymes. Fungal Biol. 2010, 114, 120–128. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Choi, N.S.; Kim, B.H.; Park, C.S.; Han, Y.J.; Lee, H.W.; Choi, J.H.; Lee, S.G.; Song, J.J. Multiple-layer substrate zymography for detection of several enzymes in a single sodium dodecyl sulfate gel. Anal. Biochem 2009, 386, 121–122. [Google Scholar] [CrossRef]
- Canchignia, H.; Altimira, F.; Montes, C.; Sánchez, E.; Tapia, E.; Miccono, M.; Espinoza, D.; Aguirre, C.; Seeger, M.; Prieto, H. Candidate nematicidal proteins in a new Pseudomonas veronii isolate identified by its antagonistic properties against Xiphinema index. J. Gen. Appl. Microbiol. 2017, 63, 11–21. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform 2004, 5, 113. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Sahl, J.W.; Caporaso, J.G.; Rasko, D.A.; Keim, P. The large-scale blast score ratio (LS-BSR) pipeline: A method to rapidly compare genetic content between bacterial genomes. PeerJ 2014, 2, e332. [Google Scholar] [CrossRef]
- Benjamini, Y.; Krieger, A.M.; Yekutieli, D. Adaptive linear step-up procedures that control the false discovery rate. Biometrika 2006, 93, 491–507. [Google Scholar] [CrossRef]
- Rangi, A.; Jajpura, L. The biopolymer sericin: Extraction and applications. J. Text Sci. Eng. 2015, 5, 1–5. [Google Scholar] [CrossRef]
- Balabanidou, V.; Grigoraki, L.; Vontas, J. Insect cuticle: A critical determinant of insecticide resistance. Curr. Opin. Insect Sci. 2018, 27, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.J.; Mou, Y.N.; Tong, S.M.; Ying, S.H.; Feng, M.G. Subtilisin-like Pr1 proteases marking the evolution of pathogenicity in a wide-spectrum insect-pathogenic fungus. Virulence 2020, 11, 365–380. [Google Scholar] [CrossRef]
- Andreis, F.C.; Schrank, A.; Thompson, C.E. Molecular evolution of Pr1 proteases depicts ongoing diversification in Metarhizium spp. Mol. Genet. Genomics. 2019, 294, 901–917. [Google Scholar] [CrossRef]
- Betzel, C.; Teplyakov, A.V.; Harutyunyan, E.H.; Saenger, W.; Wilson, K.S. Thermitase and proteinase K: A comparison of the refined three-dimensional structures of the native enzymes. Protein Eng. Des. Sel. 1990, 3, 161–172. [Google Scholar] [CrossRef]
- Siezen, R.J.; Leunissen, J.A. Subtilases: The superfamily of subtilisin-like serine proteases. Protein Sci. 1997, 6, 501–523. [Google Scholar] [CrossRef]
- Arnórsdóttir, J.; Kristjánsson, M.M.; Ficner, R. Crystal structure of a subtilisin-like serine proteinase from a psychrotrophic Vibrio species reveals structural aspects of cold adaptation. FEBS J. 2005, 272, 832–845. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Ying, S.H.; Zheng, P.; Wang, Z.L.; Zhang, S.; Xie, X.Q.; Shang, Y.; Leger, R.J.; Zhao, G.-P.; Wang, C.; et al. Genomic perspectives on the evolution of fungal entomopathogenicity in Beauveria bassiana. Sci. Rep. 2012, 2, 483. [Google Scholar] [CrossRef] [PubMed]
- Ruan, B.; London, V.; Fisher, K.E.; Gallagher, D.T.; Bryan, P.N. Engineering substrate preference in subtilisin: Structural and kinetic analysis of a specificity mutant. Biochemistry 2008, 47, 6628–6636. [Google Scholar] [CrossRef]
- Borgi, I.; Gargouri, A. Investigations on a hyper-proteolytic mutant of Beauveria bassiana: Broad substrate specificity and high biotechnological potential of a serine protease. FEMS Microbiol. Lett. 2014, 351, 23–31. [Google Scholar] [CrossRef]
- Gonçalves, E.C.S.; Perez, M.M.; Vici, A.C.; Salgado, J.C.S.; de Souza Rocha, M.; de Almeida, P.Z.; da Conceição, J.; de Almeida, S.; Coutinho, R.; Tironi, A.; et al. Potential biodiesel production from Brazilian plant oils and spent coffee grounds by Beauveria bassiana lipase 1 expressed in Aspergillus nidulans A773 using different agroindustry inputs. J. Clean. Prod. 2020, 256, 120513. [Google Scholar] [CrossRef]
- Hube, B.; Stehr, F.; Bossenz, M.; Mazur, A.; Kretschmar, M.; Schäfer, W. Secreted lipases of Candida albicans: Cloning, characterisation and expression analysis of a new gene family with at least ten members. Arch. Microbiol. 2000, 174, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Leng, B.; Xiao, Y.; Jin, K.; Ma, J.; Fan, Y.; Feng, J.; Yang, X.; Zhang, Y.; Pei, Y. Cloning of Beauveria bassiana chitinase gene Bbchit1 and its application to improve fungal strain virulence. Appl. Environ. Microbiol. 2005, 71, 363–370. [Google Scholar] [CrossRef]
- Joshi, S.; Satyanarayana, T. Biotechnology of cold-active proteases. Biology 2013, 2, 755–783. [Google Scholar] [CrossRef]
- Kuan, C.S.; Tien, C.F.; See, T.W.C.; Few, L.L. Sequence analysis and homology modeling of glyceraldehyde-3-phosphate dehydrogenase of an antarctic psychrophilic Pseudomonas sp. Afr. J. Microbiol. Res. 2012, 6, 881–891. [Google Scholar] [CrossRef]
- Vici, A.C.; da Cruz, A.F.; Facchini, F.D.A.; de Carvalho, C.C.; Pereira, M.G.; Fonseca-Maldonado, R.; Ward, R.J.; Pessela, B.C.; Fernandez-Lorente, G.; Torres, F.A.G.; et al. Beauveria bassiana Lipase A expressed in Komagataella (Pichia) pastoris with potential for biodiesel catalysis. Front. Microbiol. 2015, 6, 1083. [Google Scholar] [CrossRef]
- Lovera, A.; Belaich, M.; Villamizar, L.; Patarroyo, M.A.; Barrera, G. Enhanced virulence of Beauveria bassiana against Diatraea saccharalis using a soluble recombinant enzyme with endo-and exochitinase activity. Biol. Control 2020, 144, 104211. [Google Scholar] [CrossRef]
- Fan, Y.; Fang, W.; Guo, S.; Pei, X.; Zhang, Y.; Xiao, Y.; Li, D.; Jin, K.; Bidochka, M.J.; Pei, Y. Increased insect virulence in Beauveria bassiana strains overexpressing an engineered chitinase. Appl. Environ. Microbiol. 2007, 73, 295–302. [Google Scholar] [CrossRef]
- Mustafha, S.M.; Kamaruddin, S.; Mahadi, N.M.; Murad, A.M.A.; Abu Bakar, F.D. Gene isolation and prediction of the corresponding three-dimensional structure of subtilisin from the psychrophilic yeast, Glaciozyma antarctica PI12. Malays. J. Microbiol. 2018, 14, 452–461. [Google Scholar] [CrossRef]
- Aldossari, N.; Ishii, S. Isolation of cold-adapted nitrate-reducing fungi that have potential to increase nitrate removal in woodchip bioreactors. J. Appl. Microbiol. 2021, 131, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Alias, N.; Mazian, A.; Salleh, A.B.; Basri, M.; Rahman, R.N.Z.R.A. Molecular cloning and optimization for high level expression of cold-adapted serine protease from Antarctic yeast Glaciozyma antarctica PI12. Enzym. Res 2014, 2014, 197938. [Google Scholar] [CrossRef] [PubMed]
- Arnorsdottir, J.; Smáradóttir, R.B.; Magnusson, O.T.; Thorbjarnardóttir, S.H.; Eggertsson, G.; Kristjánsson, M.M. Characterization of a cloned subtilisin-like serine proteinase from a psychrotrophic Vibrio species. Eur. J. Biochem. 2002, 269, 5536–5546. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Wang, P.; Zeng, X.; Bartlett, D.H.; Wang, F. Shewanella psychrophila sp. nov. and Shewanella piezotolerans sp. nov., isolated from west Pacific deep-sea sediment. Int. J. Syst. Evol. Microbiol. 2007, 57, 60–65. [Google Scholar] [CrossRef]
- Techtmann, S.M.; Fitzgerald, K.S.; Stelling, S.C.; Joyner, D.C.; Uttukar, S.M.; Harris, A.P.; Alshibli, N.K.; Brown, S.D.; Hazen, T.C. Colwellia psychrerythraea strains from distant deep sea basins show adaptation to local conditions. Front. Environ. Sci. 2016, 4, 33. [Google Scholar] [CrossRef]
- Florczak, T.; Daroch, M.; Wilkinson, M.C.; Białkowska, A.; Bates, A.D.; Turkiewicz, M.; Iwanejko, L.A. Purification, characterisation and expression in Saccharomyces cerevisiae of LipG7 an enantioselective, cold-adapted lipase from the Antarctic filamentous fungus Geomyces sp. P7 with unusual thermostability characteristics. Enzyme Microb. Technol. 2013, 53, 18–24. [Google Scholar] [CrossRef]
- Mayordomo, I.; Randez-Gil, F.; Prieto, J.A. Isolation, purification, and characterization of a cold-active lipase from Aspergillus nidulans. J. Agric. Food Chem. 2000, 48, 105–109. [Google Scholar] [CrossRef]
- Luo, Y.; Zheng, Y.; Jiang, Z.; Ma, Y.; Wei, D. A novel psychrophilic lipase from Pseudomonas fluorescens with unique property in chiral resolution and biodiesel production via transesterification. Appl. Microbiol. Biotechnol. 2006, 73, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Litantra, R.; Lobionda, S.; Yim, J.H.; Kim, H.K. Expression and biochemical characterization of cold-adapted lipases from Antarctic Bacillus pumilus strains. J. Microbiol. Biotechnol. 2013, 23, 1221–1228. [Google Scholar] [CrossRef]
- Ramli, A.N.; Mahadi, N.M.; Rabu, A.; Murad, A.M.; Bakar, F.D.; Illias, R.M. Molecular cloning, expression and biochemical characterisation of a cold-adapted novel recombinant chitinase from Glaciozyma antarctica PI12. Microb. Cell Factories 2011, 10, 94. [Google Scholar] [CrossRef] [PubMed]
- Aldossari, N.; Ishii, S. Genome Sequence of Linnemannia hyalina Strain SCG-10, a Cold-Adapted and Nitrate-Reducing Fungus Isolated from Cornfield Soil in Minnesota, USA. Microbiol. Resour. Announc. 2021, 10, e00692-21. [Google Scholar] [CrossRef]
- Bendt, A.; Hüller, H.; Kammel, U.; Helmke, E.; Schweder, T. Cloning, expression, and characterization of a chitinase gene from the Antarctic psychrotolerant bacterium Vibrio sp. strain Fi: 7. Extremophiles 2001, 5, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, Y.; Tan, H.; Chi, N.; Zhang, Q.; Du, Y.; Yin, H. Characterisation of a chitinase from Pseudoalteromonas sp. DL-6, a marine psychrophilic bacterium. Int. J. Biol. Macromol. 2014, 70, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Leplat, J.; François, A.; Bousta, F. Parengyodontium album, a frequently reported fungal species in the cultural heritage environment. Fungal Biol. Rev. 2020, 34, 126–135. [Google Scholar] [CrossRef]
- Brunner-Mendoza, C.; Reyes-Montes, M.D.R.; Moonjely, S.; Bidochka, M.J.; Toriello, C. A review on the genus Metarhizium as an entomopathogenic microbial biocontrol agent with emphasis on its use and utility in Mexico. Biocontrol. Sci. Technol. 2019, 29, 83–102. [Google Scholar] [CrossRef]
- Veno, J.; Ahmad, A.N.H.; Mohamad, A.M.S.; Masomian, M.; Abd, R.; Rahman, R.N.Z. Directed evolution of recombinant C-terminal truncated Staphylococcus epidermidis lipase AT2 for the enhancement of thermostability. Int. J. Mol. Sci. 2017, 18, 2202. [Google Scholar] [CrossRef]
- Panoff, J.M.; Corroler, D.; Thammavongs, B.; Boutibonnes, P. Differentiation between cold shock proteins and cold acclimation proteins in a mesophilic gram-positive bacterium, Enterococcus faecalis JH2-2. J. Bacteriol. 1997, 179, 4451–4454. [Google Scholar] [CrossRef]
- Li, L.L.; Taghavi, S.; Izquierdo, J.A.; van der Lelie, D. Complete genome sequence of Clostridium sp. strain BNL1100, a cellulolytic mesophile isolated from corn stover. J. Bacteriol. 2012, 194, 6982–6983. [Google Scholar] [CrossRef] [PubMed]
- Cavazzini, D.; Grossi, G.; Levati, E.; Vallese, F.; Montanini, B.; Bolchi, A.; Zanotti, G.; Ottonello, S. A family of archaea-like carboxylesterases preferentially expressed in the symbiotic phase of the mycorrhizal fungus Tuber melanosporum. Sci. Rep. 2017, 7, 7628. [Google Scholar] [CrossRef] [PubMed]
- Takii, Y.; Ikeda, K.; Sato, C.; Yano, M.; Sato, T.; Konno, H. Production and characterization of β-glucosidase from Rhizopus oryzae MIBA348. J. Biol. Macromol. 2005, 5, 11–16. [Google Scholar]
- Itaya, M.; Tsuge, K.; Koizumi, M.; Fujita, K. Combining two genomes in one cell: Stable cloning of the Synechocystis PCC6803 genome in the Bacillus subtilis 168 genome. Proc. Natl. Acad. Sci. USA. 2005, 102, 15971–15976. [Google Scholar] [CrossRef]
- Jaiswal, A.; Preet, M.; Tripti, B. Production and optimization of lipase enzyme from mesophiles and thermophiles. J. Microb. Biochem. Technol. 2017, 9, 126–131. [Google Scholar] [CrossRef]
- Muroi, F.; Tachibana, Y.; Soulenthone, P.; Yamamoto, K.; Mizuno, T.; Sakurai, T.; Kobayashi, Y.; Kasuya, K.I. Characterization of a poly (butylene adipate-co-terephthalate) hydrolase from the aerobic mesophilic bacterium Bacillus pumilus. Polym. Degrad. Stab. 2017, 137, 11–22. [Google Scholar] [CrossRef]
- Chilli, W.O.N.; Assam, I. Molecular characterizations of microbial antagonists and development of bioformulations for management of bacterial. J. Exp. Biol. 2015, 3, 2. [Google Scholar] [CrossRef]
- Wu, X.; Tian, Z.; Jiang, X.; Zhang, Q.; Wang, L. Enhancement in catalytic activity of Aspergillus niger XynB by selective site-directed mutagenesis of active site amino acids. Appl. Microbiol. Biotechnol. 2018, 102, 249–260. [Google Scholar] [CrossRef]
- Gaur, A.C.; Sadasivam, K.V.; Mathur, R.S.; Magu, S.P. Role of mesophilic fungi in composting. Agric. Wastes 1982, 4, 453–460. [Google Scholar] [CrossRef]
- Watanabe, T.; Kobori, K.; Miyashita, K.; Fujii, T.; Sakai, H.; Uchida, M.; Tanaka, H. Identification of glutamic acid 204 and aspartic acid 200 in chitinase A1 of Bacillus circulans WL-12 as essential residues for chitinase activity. J. Biol. Chem. 1993, 268, 18567–18572. [Google Scholar] [CrossRef]
- Xu, P.; Ni, Z.F.; Zong, M.H.; Ou, X.Y.; Yang, J.G.; Lou, W.Y. Improving the thermostability and activity of Paenibacillus pasadenensis chitinase through semi-rational design. Int. J. Biol. Macromol. 2020, 150, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Oshima, T.; Imahori, K. Description of Thermus thermophilus (Yoshida and Oshima) comb. nov., a nonsporulating thermophilic bacterium from a Japanese thermal spa. Int. J. Syst. Evol. Microbiol. 1974, 24, 102–112. [Google Scholar] [CrossRef]
- Li, A.N.; Xie, C.; Zhang, J.; Zhang, J.; Li, D.C. Cloning, expression, and characterization of serine protease from thermophilic fungus Thermoascus aurantiacus var. levisporus. J. Microbiol. 2011, 49, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Anderson, I.; Göker, M.; Nolan, M.; Lucas, S.; Hammon, N.; Deshpande, S.; Cheng, J.-F.; Tapia, R.; Han, C.; Goodwin, L.; et al. Complete genome sequence of the hyperthermophilic chemolithoautotroph Pyrolobus fumarii type strain (1AT). Stand. Genom. Sci. 2011, 4, 381–392. [Google Scholar] [CrossRef][Green Version]
- Atomi, H.; Fukui, T.; Kanai, T.; Morikawa, M.; Imanaka, T. Description of Thermococcus kodakaraensis sp. nov., a well studied hyperthermophilic archaeon previously reported as Pyrococcus sp. KOD1. Archaea 2004, 1, 263–267. [Google Scholar] [CrossRef]
- Nguyen, Q.D.; Rezessy-Szabó, J.M.; Claeyssens, M.; Stals, I.; Hoschke, Á. Purification and characterisation of amylolytic enzymes from thermophilic fungus Thermomyces lanuginosus strain ATCC 34626. Enzym. Microb. Technol. 2002, 31, 345–352. [Google Scholar] [CrossRef]
- Gao, Y.L.; Zhang, M.L.; Wang, X.; Zhang, H.D.; Huang, J.Z.; Li, L. Isolation and characterization of a new cytotoxic polyketide–amino acid hybrid from Thermothelomyces thermophilus ATCC 42464. Nat. Prod. Res. 2021, 35, 1792–1798. [Google Scholar] [CrossRef]
- McMullan, G.; Christie, J.M.; Rahman, T.J.; Banat, I.M.; Ternan, N.G.; Marchant, R. Habitat, applications and genomics of the aerobic, thermophilic genus Geobacillus. Biochem. Soc. Trans. 2004, 32, 214–217. [Google Scholar] [CrossRef]
- Amlacher, S.; Sarges, P.; Flemming, D.; van Noort, V.; Kunze, R.; Devos, D.P.; Arumugam, M.; Bork, P.; Hurt, E. Insight into structure and assembly of the nuclear pore complex by utilizing the genome of a eukaryotic thermophile. Cell 2011, 146, 277–289. [Google Scholar] [CrossRef]
- Brienzo, M.; Arantes, V.; Milagres, A.M. Enzymology of the thermophilic ascomycetous fungus Thermoascus aurantiacus. Fungal Biol. Rev. 2008, 22, 120–130. [Google Scholar] [CrossRef]
- Zverlov, V.V.; Fuchs, K.P.; Schwarz, W.H. Chi18A, the endochitinase in the cellulosome of the thermophilic, cellulolytic bacterium Clostridium thermocellum. Appl. Environ. Microbiol. 2002, 68, 3176–3179. [Google Scholar] [CrossRef] [PubMed]
- Altimira, F.; De La Barra, N.; Rebufel, P.; Soto, S.; Soto, R.; Estay, P.; Vitta, N.; Tapia, E. Potential biological control of the pupal stage of the European grapevine moth Lobesia botrana by the entomopathogenic fungus Beauveria pseudobassiana in the winter season in Chile. BMC Res. Notes 2019, 12, 548. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arias-Aravena, M.; Altimira, F.; Gutiérrez, D.; Ling, J.; Tapia, E. Identification of Exoenzymes Secreted by Entomopathogenic Fungus Beauveria pseudobassiana RGM 2184 and Their Effect on the Degradation of Cocoons and Pupae of Quarantine Pest Lobesia botrana. J. Fungi 2022, 8, 1083. https://doi.org/10.3390/jof8101083
Arias-Aravena M, Altimira F, Gutiérrez D, Ling J, Tapia E. Identification of Exoenzymes Secreted by Entomopathogenic Fungus Beauveria pseudobassiana RGM 2184 and Their Effect on the Degradation of Cocoons and Pupae of Quarantine Pest Lobesia botrana. Journal of Fungi. 2022; 8(10):1083. https://doi.org/10.3390/jof8101083
Chicago/Turabian StyleArias-Aravena, Matias, Fabiola Altimira, Daniela Gutiérrez, Jian Ling, and Eduardo Tapia. 2022. "Identification of Exoenzymes Secreted by Entomopathogenic Fungus Beauveria pseudobassiana RGM 2184 and Their Effect on the Degradation of Cocoons and Pupae of Quarantine Pest Lobesia botrana" Journal of Fungi 8, no. 10: 1083. https://doi.org/10.3390/jof8101083
APA StyleArias-Aravena, M., Altimira, F., Gutiérrez, D., Ling, J., & Tapia, E. (2022). Identification of Exoenzymes Secreted by Entomopathogenic Fungus Beauveria pseudobassiana RGM 2184 and Their Effect on the Degradation of Cocoons and Pupae of Quarantine Pest Lobesia botrana. Journal of Fungi, 8(10), 1083. https://doi.org/10.3390/jof8101083