
Triazole-Resistance in Environmental Aspergillus fumigatus in Latin American and African Countries

 ,
,  , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Environmental Sample Collection
2.2. Triazole Antifungals Resistance Screening
2.3. Triazole-Resistance Phenotype Determination and Cyp51A Gene Sequencing
2.4. Genotyping Samples
2.5. Fungicide Concentrations in Environmental Samples
3. Results
3.1. Triazole-Resistance in A. fumigatus Analysis
3.1.1. Mexico
3.1.2. Paraguay
3.1.3. Peru
3.1.4. Benin
3.1.5. Nigeria
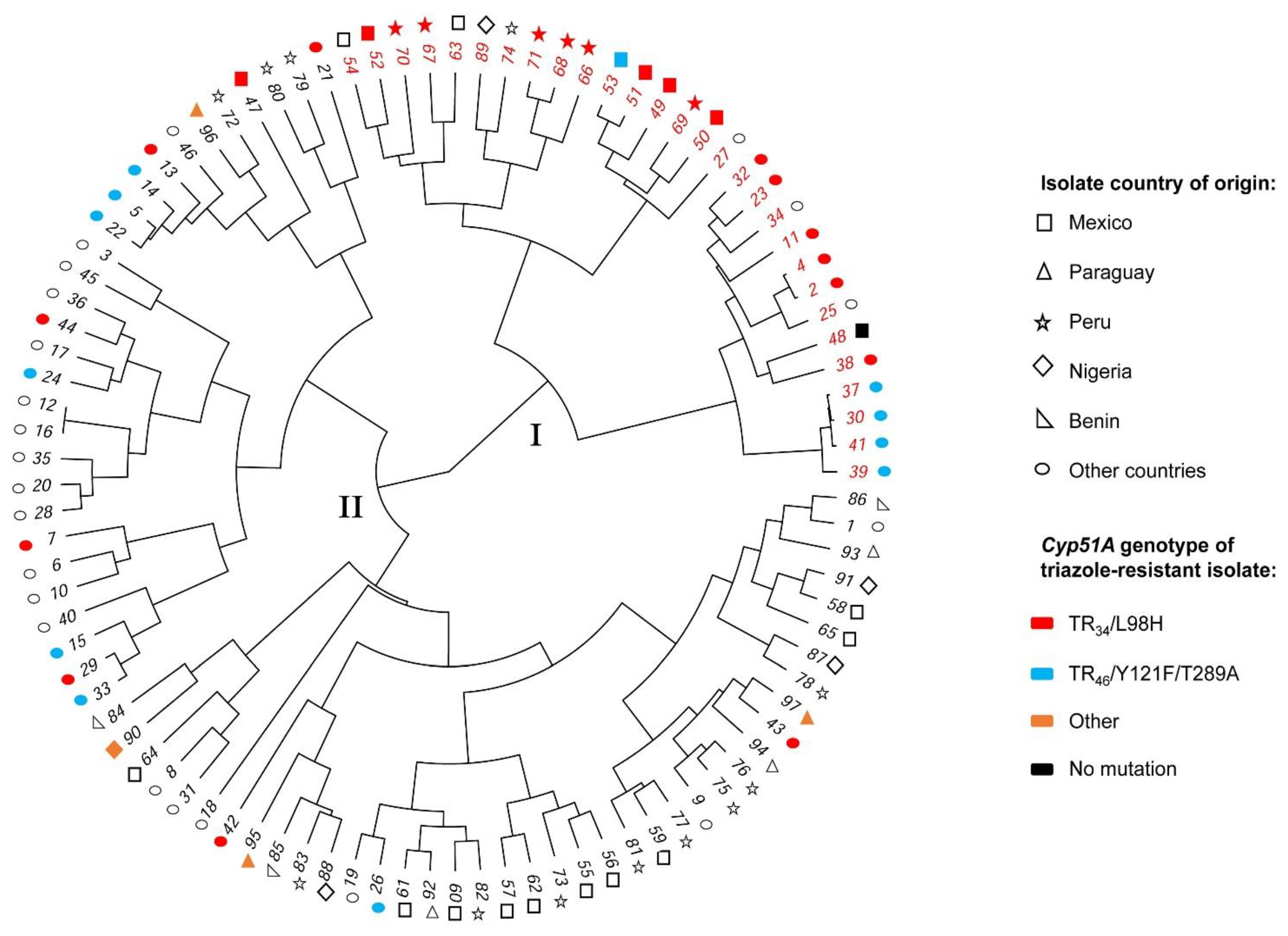
3.2. Genetic Relatedness Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kosmidis, C.; Denning, D.W. The clinical spectrum of pulmonary aspergillosis. Thorax 2014, 70, 270–277. [Google Scholar] [CrossRef]
- Schauwvlieghe, A.F.A.D.; Rijnders, B.J.A.; Philips, N.; Verwijs, R.; Vanderbeke, L.; Van Tienen, C.; Lagrou, K.; Verweij, P.E.; Van De Veerdonk, F.L.; Gommers, D.; et al. Invasive aspergillosis in patients admitted to the intensive care unit with severe influenza: A retrospective cohort study. Lancet Respir. Med. 2018, 6, 782–792. [Google Scholar] [CrossRef]
- Verweij, P.E.; Gangneux, J.-P.; Bassetti, M.; Brüggemann, R.J.M.; A Cornely, O.; Koehler, P.; Lass-Flörl, C.; van de Veerdonk, F.L.; Chakrabarti, A.; Hoenigl, M. Diagnosing COVID-19-associated pulmonary aspergillosis. Lancet Microbe 2020, 1, e53–e55. [Google Scholar] [CrossRef]
- Patterson, T.F.; Thompson, G.R.; Denning, D.W.; Fishman, J.A.; Hadley, S.; Herbrecht, R.; Kontoyiannis, D.P.; Marr, K.A.; Morrison, V.A.; Nguyen, M.H.; et al. Practice Guidelines for the Diagnosis and Management of Aspergillosis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2016, 63, e1–e60. [Google Scholar] [CrossRef]
- Ullmann, A.; Aguado, J.; Arikan-Akdagli, S.; Denning, D.; Groll, A.; Lagrou, K.; Lass-Flörl, C.; Lewis, R.; Munoz, P.; Verweij, P.; et al. Diagnosis and management of Aspergillus diseases: Executive summary of the 2017 ESCMID-ECMM-ERS guideline. Clin. Microbiol. Infect. 2018, 24, e1–e38. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, A.R.; Lagrou, K.; Meis, J.F.; Chowdhary, A.; Lockhart, S.R.; Verweij, E.P. On behalf of the ISHAM/ECMM Aspergillus Resistance Surveillance working group Triazole resistance surveillance in Aspergillus fumigatus. Med. Mycol. 2018, 56, S83–S92. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.; Sharma, C.; Meis, J.F. Azole-Resistant Aspergillosis: Epidemiology, Molecular Mechanisms, and Treatment. J. Infect. Dis. 2017, 216, S436–S444. [Google Scholar] [CrossRef]
- Verweij, P.E.; Lucas, J.A.; Arendrup, M.C.; Bowyer, P.; Brinkmann, A.J.; Denning, D.W.; Dyer, P.S.; Fisher, M.C.; Geenen, P.L.; Gisi, U.; et al. The one health problem of azole resistance in Aspergillus fumigatus: Current insights and future research agenda. Fungal Biol. Rev. 2020, 34, 202–214. [Google Scholar] [CrossRef]
- Schoustra, S.E.; Debets, A.J.; Rijs, A.J.; Zhang, J.; Snelders, E.; Leendertse, P.C.; Melchers, W.J.; Rietveld, A.G.; Zwaan, B.J.; Verweij, P.E. Environmental Hotspots for Azole Resistance Selection of Aspergillus fumigatus, the Netherlands. Emerg. Infect. Dis. 2019, 25, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Snelders, E.; Karawajczyk, A.; Schaftenaar, G.; Verweij, P.E.; Melchers, W.J.G. Azole Resistance Profile of Amino Acid Changes in Aspergillus fumigatus CYP51A Based on Protein Homology Modeling. Antimicrob. Agents Chemother. 2010, 54, 2425–2430. [Google Scholar] [CrossRef]
- Sewell, T.R.; Zhu, J.; Rhodes, J.; Hagen, F.; Meis, J.F.; Fisher, M.C.; Jombart, T. Nonrandom Distribution of Azole Resistance across the Global Population of Aspergillus fumigatus. mBio 2019, 10, e00392-19. [Google Scholar] [CrossRef] [PubMed]
- Resendiz-Sharpe, A.; Mercier, T.; A Lestrade, P.P.; Van Der Beek, M.T.; Borne, P.A.V.D.; Cornelissen, J.J.; De Kort, E.; A Rijnders, B.J.; Schauwvlieghe, A.F.A.D.; E Verweij, P.; et al. Prevalence of voriconazole-resistant invasive aspergillosis and its impact on mortality in haematology patients. J. Antimicrob. Chemother. 2019, 74, 2759–2766. [Google Scholar] [CrossRef]
- Lestrade, P.P.; Bentvelsen, R.G.; Schauwvlieghe, A.F.A.D.; Schalekamp, S.; Van Der Velden, W.J.F.M.; Kuiper, E.J.; Van Paassen, J.; Van Der Hoven, B.; A Van Der Lee, H.; Melchers, W.J.G.; et al. Voriconazole Resistance and Mortality in Invasive Aspergillosis: A Multicenter Retrospective Cohort Study. Clin. Infect. Dis. 2019, 68, 1463–1471. [Google Scholar] [CrossRef] [PubMed]
- Verweij, P.E.; Ananda-Rajah, M.; Andes, D.; Arendrup, M.C.; Brüggemann, R.J.; Chowdhary, A.; Cornely, O.A.; Denning, D.W.; Groll, A.H.; Izumikawa, K.; et al. International expert opinion on the management of infection caused by azole-resistant Aspergillus fumigatus. Drug Resist. Updat. 2015, 21–22, 30–40. [Google Scholar] [CrossRef]
- SWAB Invasive Fungal Infections Guidelines Committee. Dutch Working Party on Antibiotic Policy. SWAB Guideline XIII—Invasive Fungal Infections 2017. 2017, pp. 1–66. Available online: https://swabid.nl/sites/default/files/SWABRichtlijnMycosen2017%28final%29_compressed.pdf (accessed on 7 May 2019).
- Rhodes, J.C. Aspergillus fumigatus: Growth and virulence. Med. Mycol. 2006, 44, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, K.L.; Mellado, E.; Lass-Flörl, C.; Rodriguez-Tudela, J.L.; Johansen, H.K.; Arendrup, M.C. Environmental Study of Azole-Resistant Aspergillus fumigatus and Other Aspergilli in Austria, Denmark, and Spain. Antimicrob. Agents Chemother. 2010, 54, 4545–4549. [Google Scholar] [CrossRef] [PubMed]
- Normand, A.C.; Becker, P.; Gabriel, F.; Cassagne, C.; Accoceberry, I.; Gari-Toussaint, M.; Hasseine, L.; De Geyter, D.; Pierard, D.; Surmont, I.; et al. Validation of a New Web Application for Identification of Fungi by Use of Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry. J. Clin. Microbiol. 2017, 55, 2661–2670. [Google Scholar] [CrossRef]
- EUCAST: The European Committee on Antimicrobial Susceptibility Testing. Method for the Determination of Broth Dilution Minimun Inhibitory Concentrations of Antifungal Agents for Conidia Forming Moulds, 2020; (EUCAST E.DEF 9.3.2 April 2020). Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/AFST/Files/EUCAST_E_Def_9.3.2_Mould_testing_definitive_revised_2020.pdf (accessed on 1 July 2020).
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs for Antifungal Agents, Version 10.0; EUCAST: Växjö, Sweden, 2020. [Google Scholar]
- Chowdhary, A.; Sharma, C.; Kathuria, S.; Hagen, F.; Meis, J.F. Azole-resistant Aspergillus fumigatus with the environmental TR46/Y121F/T289A mutation in India. J. Antimicrob. Chemother. 2013, 69, 555–557. [Google Scholar] [CrossRef]
- Vermeulen, E.; Maertens, J.; De Bel, A.; Nulens, E.; Boelens, J.; Surmont, I.; Mertens, A.; Boel, A.; Lagrou, K. Nationwide Surveillance of Azole Resistance in Aspergillus Diseases. Antimicrob. Agents Chemother. 2015, 59, 4569–4576. [Google Scholar] [CrossRef]
- De Valk, H.; Meis, J.F.G.M.; Curfs, I.M.; Muehlethaler, K.; Mouton, J.W.; Corné, H.W. Use of a Novel Panel of Nine Short Tandem Repeats for Exact and High-Resolution Fingerprinting of Aspergillus fumigatus Isolates Use of a Novel Panel of Nine Short Tandem Repeats for Exact and High-Resolution Fingerprinting of Aspergillus fumigatus Isolat. J. Clin. Microbiol. 2005, 43, 4112–4120. [Google Scholar] [CrossRef]
- Resendiz-Sharpe, A.; Hokken, M.W.J.; Mercier, T.; Merckx, R.; Verhagen, K.; Dewitte, L.; Melchers, W.J.G.; Verweij, P.E.; Maertens, J.; Lagrou, K. Hmg1 Gene Mutation Prevalence in Triazole-Resistant Aspergillus fumigatus Clinical Isolates. J. Fungi 2020, 6, 227. [Google Scholar] [CrossRef]
- R Foundation for Statistical Computing. R: A Language and Environment for Statistical Computing; R Foundation: Vienna, Austria, 2018. [Google Scholar]
- Hurst, S.F.; Berkow, E.L.; Stevenson, K.L.; Litvintseva, A.P.; Lockhart, S.R. Isolation of azole-resistant Aspergillus fumigatus from the environment in the south-eastern USA. J. Antimicrob. Chemother. 2017, 72, 2443–2446. [Google Scholar] [CrossRef] [PubMed]
- Wiederhold, N.P.; Gil, V.G.; Gutierrez, F.; Lindner, J.R.; AlBataineh, M.T.; McCarthy, D.I.; Sanders, C.; Fan, H.; Fothergill, A.W.; Sutton, D.A. First Detection of TR34 L98H and TR46 Y121F T289A Cyp51 Mutations in Aspergillus fumigatus Isolates in the United States. J. Clin. Microbiol. 2016, 54, 168–171. [Google Scholar] [CrossRef] [PubMed]
- Berkow, E.L.; Nunnally, N.S.; Bandea, A.; Kuykendall, R.; Beer, K.; Lockhart, S.R. Detection of TR 34 /L98H CYP51A Mutation through Passive Surveillance for Azole-Resistant Aspergillus fumigatus in the United States from 2015 to 2017. Antimicrob. Agents Chemother. 2018, 62, e02240-17. [Google Scholar] [CrossRef] [PubMed]
- Parent-Michaud, M.; Dufresne, P.J.; Fournier, E.; Folch, B.; Martineau, C.; Moreira, S.; Doucet, N.; De Repentigny, L.; Dufresne, S.F. Prevalence and mechanisms of azole resistance in clinical isolates of Aspergillus section Fumigati species in a Canadian tertiary care centre, 2000 to 2013. J. Antimicrob. Chemother. 2020, 75, 849–858. [Google Scholar] [CrossRef] [PubMed]
- García Hernández, J.; Leyva Morales, J.; Martínez Rodríguez, I.; Hernández Ochoa, M.; Aldana Madrid, M.; Rojas García, A.; Betancourt Lozano, M.; Perez Herrera, N.; Perera Rios, J. Estado actual de la investigación sobre plaguicidas en méxico. Rev. Int. Contam. Ambient. 2018, 34, 29–60. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). Pesticides Use Database. Last Updated 10 September 2020. Available online: http://www.fao.org/faostat/en/#data/RP (accessed on 20 January 2021).
- Fungicide Resistance Action Committee (FRAC). FRAC Code List FRAC Code List ©* 2021: Fungal Control Agents Sorted by Cross Resistance Pattern and Mode of Action (Including Coding for FRAC Groups on Product Labels); FRAC: Basel, Switzerland, 2021; Available online: https://www.frac.info/docs/default-source/publications/frac-code-list/frac-code-list-2021--final.pdf?sfvrsn=f7ec499a (accessed on 9 April 2021).
- Snelders, E.; Camps, S.M.T.; Karawajczyk, A.; Schaftenaar, G.; Kema, G.H.J.; Van Der Lee, H.A.; Klaassen, C.H.; Melchers, W.J.G.; Verweij, P.E. Triazole Fungicides Can Induce Cross-Resistance to Medical Triazoles in Aspergillus fumigatus. PLoS ONE 2012, 7, e31801. [Google Scholar] [CrossRef] [PubMed]
- Sagatova, A.; Keniya, M.V.; Wilson, R.K.; Sabherwal, M.; Tyndall, J.D.A.; Monk, B.C. Triazole resistance mediated by mutations of a conserved active site tyrosine in fungal lanosterol 14α-demethylase. Sci. Rep. 2016, 6, 26213. [Google Scholar] [CrossRef]
- Caldas, E.D.; Food and Agriculture Organization of the United Nations (FAO). Tebuconazole Physical and Chemical Properties, Metabolims, Use and Residues; FAO: Rome, Italy, 2012. [Google Scholar]
- Ambrus, Á.; van der VeldeKoerts, T.; Ossendorp, B.C. PROPICONAZOLE (160). 2005. Available online: http://www.fao.org/fileadmin/templates/agphome/documents/Pests_Pesticides/JMPR/Evaluation07/Propiconazole.pdf (accessed on 9 April 2021).
- Gonzalez-Lara, M.F.; Roman-Montes, C.M.; Diaz-Lomeli, P.; Rangel-Cordero, A.; O Valenzuela, M.; Ponce-De-Leon, A.; Sifuentes-Osornio, J.; Ostrosky-Zeichner, L.; Martinez-Gamboa, A. Azole resistance and cyp51A mutation screening in Aspergillus fumigatus in Mexico. J. Antimicrob. Chemother. 2019, 74, 2047–2050. [Google Scholar] [CrossRef]
- Pontes, L.; Beraquet, C.A.G.; Arai, T.; Pigolli, G.L.; Lyra, L.; Watanabe, A.; Moretti, M.L.; Schreiber, A.Z. Aspergillus fumigatus Clinical Isolates Carrying CYP51A with TR34/L98H/S297T/F495I Substitutions Detected after Four-Year Retrospective Azole Resistance Screening in Brazil. Antimicrob. Agents Chemother. 2019, 64. [Google Scholar] [CrossRef]
- Alvarez-Moreno, C.; Lavergne, R.-A.; Hagen, F.; Morio, F.; Meis, J.F.; Le Pape, P. Azole-resistant Aspergillus fumigatus harboring TR34/L98H, TR46/Y121F/T289A and TR53 mutations related to flower fields in Colombia. Sci. Rep. 2017, 7, srep45631. [Google Scholar] [CrossRef]
- Alvarez-Moreno, C.; Lavergne, R.-A.; Hagen, F.; Morio, F.; Meis, J.F.; Le Pape, P. Fungicide-driven alterations in azole-resistant Aspergillus fumigatus are related to vegetable crops in Colombia, South America. Mycologia 2019, 111, 217–224. [Google Scholar] [CrossRef]
- Redagricola. Redagrícola Una Conversación Técnica Sobre Agricultura. 2013. Available online: https://www.redagricola.com/pe/assets/uploads/2017/08/raperu09.pdf (accessed on 9 April 2021).
- Servicio Nacional de Sanidad Agraria del Peru (SENASA). Lista De Plaguicidas Agricolas Con Registro Vigente Del Peru. Anexo 1. Available online: https://www.senasa.gob.pe/senasa/descargasarchivos/jer/SUB_SEC_NOR/ANEXO 1.PDF (accessed on 9 April 2021).
- (SENAVE) SN de C y SV y de S. Prod_Fitosanitarios del Paraguay. Available online: http://secure.senave.gov.py:8443/registros/servlet/prod_agro (accessed on 9 April 2021).
- Miles, M.R.; Levy, C.; Morel, W.; Mueller, T.; Steinlage, T.; Van Rij, N.; Frederick, R.D.; Hartman, G.L. International Fungicide Efficacy Trials for the Management of Soybean Rust. Plant Dis. 2007, 91, 1450–1458. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, B.; Illescas, L.R.; Posadas, A.; E Campos, P. Azole resistance among clinical isolates of Aspergillus fumigatus in Lima-Peru. Med. Mycol. 2019, 58, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Joseph, L.; Hagen, F.; Meis, J.F.; Khan, Z. Concomitant occurrence of itraconazole-resistant and -susceptible strains of Aspergillus fumigatus in routine cultures. J. Antimicrob. Chemother. 2015, 70, 412–415. [Google Scholar] [CrossRef] [PubMed]
- Yerbanga, I.; Resendiz-Sharpe, A.; Bamba, S.; Lagrou, K.; Diallo, S.N.; Rodriguez-Villalobos, H.; Denis, O.; Montesinos, I. First Investigative Study of Azole-Resistant Aspergillus fumigatus in the Environment in Burkina Faso. Int. J. Environ. Res. Public Health 2021, 18, 2250. [Google Scholar] [CrossRef] [PubMed]
- Sharma, C.; Hagen, F.; Moroti, R.; Meis, J.F.; Chowdhary, A. Triazole-resistant Aspergillus fumigatus harbouring G54 mutation: Is it de novo or environmentally acquired? J. Glob. Antimicrob. Resist. 2015, 3, 69–74. [Google Scholar] [CrossRef]
- Kemoi, E.K.; Nyerere, A.; Bii, C.C. Triazole-Resistant Aspergillus fumigatus from Fungicide-Experienced Soils in Naivasha Subcounty and Nairobi County, Kenya. Int. J. Microbiol. 2018, 2018, 7147938. [Google Scholar] [CrossRef] [PubMed]
- Federal Operation Coordinating Unit, National Social Safety Nets Coordinating Office (NASSCO). Federal Republic of Nigeria Youth Employment and Social Support Operation (YESSO). INTEGRATED PEST MANAGEMENT PLAN (IPMP). 2016. Available online: http://documents1.worldbank.org/curated/en/115291468099839070/pdf/SFG2220-EA-P157899-Box396258B-PUBLIC-Disclosed-5-20-2016.pdf (accessed on 21 January 2021).
- Agbohessi, P.T.; Toko, I.I.; Ouédraogo, A.; Jauniaux, T.; Mandiki, S.; Kestemont, P. Assessment of the health status of wild fish inhabiting a cotton basin heavily impacted by pesticides in Benin (West Africa). Sci. Total Environ. 2015, 506–507, 567–584. [Google Scholar] [CrossRef]
- Ouédraogo, J.B.; Ouédraogo, R.; Ilboudo, S.; Bayili, B.; Paré, T.; Kékélé, A.; Sawadogo, B. Use of Agricultural Pesticides in Three Regions in the West of Burkina Faso and Evaluation of Their Impact on Health and the Environment: The Case of the Mouhoun Boucle, Cascades and Hauts-Bassins Regions. 2016. Available online: http://www.pic.int/Portals/5/download.aspx?d=UNEP-FAO-RC-Workshop-BurkinaFaso-Report-201212.Fr.pdf (accessed on 26 February 2021). (In French).
- Lahr, J.; Buij, R.; Katagira, F.; Van Der Valk, H. Pesticides in the Southern Agricultural Growth Corridor of Tanzania (SAGCOT): A Scoping Study of Current and Future Use, Associated Risks and Identification of Actions for Risk Mitigation; Wageningen Environmental Research: Wageningen, The Netherlands, 2016. [Google Scholar] [CrossRef]
- Isla, G.; Leonardelli, F.; Tiraboschi, I.N.; Refojo, N.; Hevia, A.; Vivot, W.; Szusz, W.; Córdoba, S.B.; García-Effron, G. First Clinical Isolation of an Azole-Resistant Aspergillus fumigatus Isolate Harboring a TR46 Y121F T289A Mutation in South America. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef]
- Snelders, E.; Camps, S.M.; Karawajczyk, A.; Rijs, A.J.; Zoll, J.; Verweij, P.E.; Melchers, W.J. Genotype–phenotype complexity of the TR46/Y121F/T289A cyp51A azole resistance mechanism in Aspergillus fumigatus. Fungal Genet. Biol. 2015, 82, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Fraczek, M.G.; Bromley, M.; Bowyer, P. An Improved Model of the Aspergillus fumigatus CYP51A Protein. Antimicrob. Agents Chemother. 2011, 55, 2483–2486. [Google Scholar] [CrossRef]
- . Escribano, P.; Recio, S.; Peláez, T.; Bouza, E.; Guinea, J. Aspergillus fumigatus Strains with Mutations in thecyp51AGene Do Not Always Show Phenotypic Resistance to Itraconazole, Voriconazole, or Posaconazole. Antimicrob. Agents Chemother. 2011, 55, 2460–2462. [Google Scholar] [CrossRef] [PubMed]
- Alanio, A.; Cabaret, O.; Sitterlé, E.; Costa, J.-M.; Brisse, S.; Cordonnier, C.; Bretagne, S. Azole Preexposure Affects the Aspergillus fumigatus Population in Patients. Antimicrob. Agents Chemother. 2012, 56, 4948–4950. [Google Scholar] [CrossRef][Green Version]
- Buil, J.B.; Van Der Lee, H.A.L.; Rijs, A.J.M.M.; Zoll, J.; Hovestadt, J.A.M.F.; Melchers, W.J.G.; Verweij, P.E. Single-Center Evaluation of an Agar-Based Screening for Azole Resistance in Aspergillus fumigatus by Using VIPcheck. Antimicrob. Agents Chemother. 2017, 61, e01250-17. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Country | Samples (No.) | % of Total Samples | % of A. fumigatus Positive Culture Samples |
|---|---|---|---|
| Mexico | |||
| Total samples | 198 | ||
| Growth any fungi | 172 | 87.0 | |
| A. fumigatus species | 102 | 51.5 | |
| Triazole-resistant A. fumigatus | 7 | 3.5 | 6.9 |
| Cyp51A gene mutation | 6 | 3.0 | 5.9 |
| No mutation cyp51A gene mutation | 1 | 0.5 | 1.0 |
| Paraguay | |||
| Total samples | 85 | ||
| Growth any fungi | 62 | 73 | |
| A. fumigatus species | 36 | 42.4 | |
| Triazole-resistant A. fumigatus | 3 | 3.5 | 8.3 |
| Cyp51A gene mutation | 2 | 2.4 | 5.6 |
| No mutation cyp51A gene mutation | 1 | 1.2 | 2.8 |
| Peru | |||
| Total samples | 106 | ||
| Growth any fungi | 87 | 82.1 | |
| A. fumigatus species | 61 | 57.5 | |
| Triazole-resistant A. fumigatus | 6 | 5.7 | 9.8 |
| Cyp51A gene mutation | 6 | 5.7 | 9.8 |
| No mutation cyp51A gene mutation | 0 | 0.0 | 0.0 |
| Benin | |||
| Total samples | 95 | ||
| Growth any fungi | 86 | 90.5 | |
| A. fumigatus species | 25 | 26.3 | |
| Triazole-resistant A. fumigatus | 0 | 0.0 | 0.0 |
| Cyp51A gene mutation | 0 | 0.0 | 0.0 |
| No mutation cyp51A gene mutation | 0 | 0.0 | 0.0 |
| Nigeria | |||
| Total samples | 100 | ||
| Growth any fungi | 93 | 93 | |
| A. fumigatus species | 46 | 46 | |
| Triazole-resistant A. fumigatus | 1 | 1.0 | 2.2 |
| Cyp51A gene mutation | 1 | 1.0 | 2.2 |
| No mutation cyp51A gene mutation | 0 | 0.0 | 0.0 |
| Country—Regions: | Sample ID Number | EUCAST MIC (mg/L) 1 | cyp51A Gene Mutation | Location 3 | ||||
|---|---|---|---|---|---|---|---|---|
| AMB | VCZ | POS | ITC | TEB 2 | ||||
| Mexico | ||||||||
| Guanajuato | 47 | 1 | 4 | 0.5 | >8 | 8 | TR34/L98H | Flower bed (U) |
| Guanajuato | 48 | 1 | 4 | 0.5 | 16 | 0.5 | No mutation | Park (U) |
| Guanajuato | 49 | 0.5 | 2 | 1 | >16 | 8 | TR34/L98H | Commercial compost (U) |
| Guanajuato | 50 | 1 | 4 | 0.5 | >16 | 8 | TR34/L98H/S297T/F495I | Commercial compost (U) |
| Guanajuato | 51 | 1 | 16 | 2 | >16 | 32 | TR34/L98H | Flower bed (U) |
| Mexico City | 52 | 0.5 | 4 | 0.5 | >16 | 16 | TR34/L98H | Greenhouse (U) |
| Mexico City | 53 | 0.5 | >16 | 0.5 | 1 | 16 | TR46/Y121F/T289A | Flower bed (U) |
| Paraguay | ||||||||
| Asunción | 95 | 0.5 | 2 | 0.25 | 1 | 4 | F46Y, M172V, E427K | Flower bed (U) |
| Asunción | 96 | 1 | 4 | 0.25 | 1 | 2 | No mutation | Flower bed (U) |
| Asunción | 97 | 1 | 2 | 0.25 | 1 | 4 | F46Y, M172V, N248T, D255E, E427K | Flower bed (U) |
| Peru | ||||||||
| Lima City | 66 | 1 | 2 | 1 | >16 | 8 | TR34/L98H/S297T/F495I | Agricultural field (R) |
| Lima City | 67 | 1 | 8 | 1 | >16 | 16 | TR34/L98H | Agricultural field (R) |
| Lima City | 68 | 0.25 | 8 | 1 | >16 | 16 | TR34/L98H | Park (U) |
| Lima City | 69 | 0.5 | 8 | 1 | >16 | 16 | TR34/L98H | Flower bed (U) |
| Lima City | 70 | 0.5 | 4 | 0.5 | >16 | 32 | TR34/L98H | Agricultural field (R) |
| Lima City | 71 | 0.5 | 8 | 1 | >16 | 64 | TR34/L98H | Commercial compost (U) |
| Nigeria | ||||||||
| Lagos Ibadan | 90 | 1 | 2 | 0.25 | 4 | 4 | M172V | Flower bed (U) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Resendiz-Sharpe, A.; Dewaele, K.; Merckx, R.; Bustamante, B.; Vega-Gomez, M.C.; Rolon, M.; Jacobs, J.; Verweij, P.E.; Maertens, J.; Lagrou, K. Triazole-Resistance in Environmental Aspergillus fumigatus in Latin American and African Countries. J. Fungi 2021, 7, 292. https://doi.org/10.3390/jof7040292
Resendiz-Sharpe A, Dewaele K, Merckx R, Bustamante B, Vega-Gomez MC, Rolon M, Jacobs J, Verweij PE, Maertens J, Lagrou K. Triazole-Resistance in Environmental Aspergillus fumigatus in Latin American and African Countries. Journal of Fungi. 2021; 7(4):292. https://doi.org/10.3390/jof7040292
Chicago/Turabian StyleResendiz-Sharpe, Agustin, Klaas Dewaele, Rita Merckx, Beatriz Bustamante, Maria Celeste Vega-Gomez, Miriam Rolon, Jan Jacobs, Paul E. Verweij, Johan Maertens, and Katrien Lagrou. 2021. "Triazole-Resistance in Environmental Aspergillus fumigatus in Latin American and African Countries" Journal of Fungi 7, no. 4: 292. https://doi.org/10.3390/jof7040292
APA StyleResendiz-Sharpe, A., Dewaele, K., Merckx, R., Bustamante, B., Vega-Gomez, M. C., Rolon, M., Jacobs, J., Verweij, P. E., Maertens, J., & Lagrou, K. (2021). Triazole-Resistance in Environmental Aspergillus fumigatus in Latin American and African Countries. Journal of Fungi, 7(4), 292. https://doi.org/10.3390/jof7040292