Abstract
Many Calonectria species are causal agents of diseases on several forestry, agricultural and horticultural crops. Calonectria leaf blight is one of the most important diseases associated with Eucalyptus plantations and nurseries in Asia and South America. Recently, symptoms of leaf rot and leaf blight caused by Calonectria species were observed in a one-year-old Eucalyptus experimental plantation in GuangXi Province, southern China. To better understand the species diversity, mating strategy and pathogenicity of Calonectria species isolated from diseased tissues and soils, diseased leaves and soils under the trees from ten Eucalyptus urophylla hybrid genotypes were collected. Three hundred and sixty-eight Calonectria isolates were obtained from diseased Eucalyptus leaves and soils under these trees, and 245 representative isolates were selected based on the sampling substrates and Eucalyptus genotypes and identified by DNA sequence analyses based on the translation elongation factor 1-alpha (tef1), β-tubulin (tub2), calmodulin (cmdA) and histone H3 (his3) gene regions, as well as a combination of morphological characteristics. These isolates were identified as Calonectria hongkongensis (50.2%), C. pseudoreteaudii (47.4%), C. aconidialis (1.6%), C. reteaudii (0.4%) and C. auriculiformis (0.4%). This is the first report of C. reteaudii and C. auriculiformis occurrence in China. Calonectria pseudoreteaudii was isolated from both Eucalyptus diseased leaves and soils; the other four species were only obtained from soils. MAT1-1-1 and MAT1-2-1 gene amplification and mating type assignment results showed that C. pseudoreteaudii is heterothallic and an asexual cycle represents the primary reproductive mode, C. reteaudii and C. auriculiformis are likely to be heterothallic and C. hongkongensis and C. aconidialis are homothallic. Based on the genetic diversity comparisons for C. pseudoreteaudii isolates from diseased leaves and soils, we hypothesize that C. pseudoreteaudii in soils was spread from diseased leaves. Both the mycelia plug and conidia suspension inoculations indicated that all five Calonectria species were pathogenic to the two Eucalyptus genotypes tested and the tolerance of the two genotypes differed. It is necessary to understand the ecological niche and epidemiological characteristics of these Calonectria species and to select disease resistant Eucalyptus genotypes in southern China in the future.
1. Introduction
Currently, the Eucalyptus (Myrtaceae, Myrtales) plantation area has expanded to more than 5.4 million hm2 in China, accounting for nearly 2.5% of the national total forestry area. China’s Eucalyptus plantations produce more than 30 million m3 timber per year, accounting for more than 33% of China’s total domestic timber production [1]. Therefore, Eucalyptus plantations play an important role in wood supply in China. Eucalyptus plantations are mainly distributed in GuangXi, GuangDong, YunNan, FuJian, SiChuan and HaiNan Provinces in southern China. Of these, GuangXi Province has the largest area of Eucalyptus [1,2].
With the extensive development of Eucalyptus plantations over the past 30 years, pathogens and pests have rapidly emerged as a significant threat to Eucalyptus plantations in China [2,3]. Important diseases in Eucalyptus plantations include bacterial wilt caused by Ralstonia pseudosolanacearum [4,5]; stem canker/wilt caused by species of Botryosphaeriaceae [6,7,8], Cryphonectriaceae [9,10,11], Ceratocystis [12,13] and Teratosphaeria zuluensis [14,15]; and leaf spot/blight caused by Mycosphaerellaceae and Teratosphaeriaceae species [16,17], Calonectria [18,19,20] and Quambalaria [21,22]. Of these, leaf blight caused by Calonectria species is considered to be one of the most important diseases in Eucalyptus plantations in southern China [3,18,20].
The genus Calonectria includes important plant pathogens infecting more than 335 plant species, distributed by nearly 100 plant families. These plants include forestry, agricultural and horticultural crops [23,24,25]. In forestry, Calonectria species mainly attack the families Fabaceae (Acacia spp.), Myrtaceae (Eucalyptus spp.) and Pinaceae (Pinus spp.) [23,24]. In Eucalyptus, this fungus causes stem and leaf rot in nurseries, leaf and shoot blight and stem canker in plantation [5,20,23]. These diseases are mainly reported in Asia, Africa and South America [20,26,27].
To date, 23 species of Calonectria have been identified and described based on DNA sequence data in China [20,28,29]. Of these, 15 species, C. aciculata, C. crousiana, C. eucalypti, C. fujianensis, C. hawksworthii, C. pauciramosa, C. cerciana, C. pseudoreteaudii, C. aconidialis, C. asiatica, C. honghensis, C. hongkongensis, C. kyotensis, C. lateralis and C. yunnanensis have been isolated from Eucalyptus plants or the soils in southern China. The first eight of these species were isolated from infected tissues (leaves, shoots or branches) in plantations, while the latter nine species were from soils (C. cerciana and C. pseudoreteaudii were isolated from both Eucalyptus tissues and soils) [20,28,29]. Additionally, C. cerciana, C. pauciramosa and C. pseudoreteaudii were also isolated from diseased seedlings in nurseries [19,29,30] and C. pseudoreteaudii from soils in a Eucalyptus nursery [19]. Our previous research results of conidia suspension inoculations in Eucalyptus seedlings showed that C. crousiana, C. eucalypti, C. fujianensis, C. pauciramosa and C. pseudoreteaudii are all pathogenic to all the tested Eucalyptus genotypes [18,20]. All the five tested species were originally isolated from diseased Eucalyptus tissues in plantations or nurseries [18,20].
Previous research results indicated that a relatively large number of Calonectria species are distributed in Eucalyptus plantations, both in diseased tissues and soils under these trees. However, the differences of species diversity, mating strategy and pathogenicity of these fungi isolated from diseased tissues and soils remain unknown. Recently, leaf blight caused by Calonectria in one Eucalyptus experimental plantation in GuangXi Province was observed, the diseased leaves from Eucalyptus trees and soils under these trees were collected, and the Calonectria fungi were isolated. The aims of this study were to (i) identify the Calonectria fungi based on multi-gene phylogeny and morphological characteristics, (ii) compare the species diversity between isolates obtained from diseased leaves and soils, (iii) test the mating type of obtained Calonectria species, and (iv) test the pathogenicity of Calonectria species obtained from both diseased leaves and soils.
2. Materials and Methods
2.1. Disease Survey Site, Sample Collection and Fungal Isolation
The disease survey was conducted in a one-year-old Eucalyptus breeding experimental plantation in the BeiHai region, GuangXi Province, southern China (21°33′19.8756″ N, 109°42′27.0792″ E) in October, 2018. Ten Eucalyptus genotypes were planted in the experimental plantation. These included six Eucalyptus urophylla × E. grandis hybrid genotypes (CEPT1860–CEPT1865) and four E. urophylla × E. tereticornis hybrid genotypes (CEPT1866–CEPT1869). All ten Eucalyptus genotypes were naturally infected by Calonectria species (Figure 1).

Figure 1.
Disease symptoms on multiple Eucalyptus genotypes in one experimental plantation caused by species of Calonectria. (A–C): Leaf spot in three E. urophylla × E. grandis hybrid genotypes CEPT1863 (A), CEPT1861 (B), and CEPT1862 (C), the infected leaves of CEPT1861 and CEPT1862 became blighted and dried; (D–H): Leaf spot and blight in three E. urophylla × E. tereticornis hybrid genotypes, CEPT1866 (D,E), CEPT1868 (F), and CEPT1869 (G,H).
Diseased leaves with typical symptoms caused by Calonectria species were collected from 13 to 20 trees for each of the ten Eucalyptus hybrid genotypes, depending on the planted areas of each genotype. Soil samples under each sampled diseased tree were also collected. These samples of diseased leaves and soils were transported to the laboratory for isolation, morphological examination, and further molecular research.
To induce Calonectria sporulation, diseased leaves were placed in moist dishes (diameter 70 mm, height 16 mm; tissue paper moistened with sterile water) at room temperature and incubated for 1–3 days. Soil samples were baited with Medicago sativa (alfalfa) germinating seeds using the method described by Crous [23]. Fungal isolates with typical morphological characteristics of Calonectria were isolated from diseased leaves and soil samples. The conidia masses were transferred to 2% (v/v) malt extract agar (MEA) (20 g malt extract powder and 20 g agar powder per liter of water: malt extract powder was obtained from the Beijing Shuangxuan microbial culture medium products factory, Beijing, China; the agar powder was obtained from Beijing Solarbio Science & Technology Co., Ltd., Beijing, China) with a sterile needles under stereoscopic microscope and incubated for 3–5 days. To obtain pure cultures, a single hyphal tip from each culture was transferred to 2% MEA plates and incubated at room temperature for 7–10 days. The pure cultures were deposited in the culture collection (CSF) at the China Eucalypt Research Centre (CERC) of the Chinese Academy of Forestry (CAF) in ZhanJiang, GuangDong Province, China.
2.2. DNA Extraction, PCR Amplification and Sequencing
Representative isolates were selected based on the sampling substrates and Eucalyptus genotypes for DNA extraction and sequence comparisons. DNA was extracted from 10-day-old cultures and mycelia were collected using a sterilized scalpel and transferred to 2 mL Eppendorf tubes. Total genomic DNA was extracted following the CTAB protocol described by van Burik and co-authors [31]. The extracted DNA was dissolved using 30 µL TE buffer (1 M Tris-HCl and 0.5 M EDTA, pH 8.0), and 3 µL RNase (10 mg/mL) was added at 37 °C for 1 h to degrade RNA. Finally, DNA concentration was measured with a Nano-Drop 2000 spectrometer (Thermo Fisher Scientific, Waltham, MA, USA).
According to previous research results, sequences of partial gene regions of translation elongation factor 1-alpha (tef1), β-tubulin (tub2), calmodulin (cmdA), and histone H3 (his3) were used to successfully identify Calonectria species [26,28,32]. These four partial gene regions were amplified using the primer pairs EF1-728F/EF2, T1/CYLTUB1R, CAL-228F/CAL-2Rd and CYLH3F/CYLH3R, respectively, the PCR procedure was conducted as described by Liu and Chen [33], Lombard and co-authors [30] (Table 1).
To obtain accurate sequences for each sequenced isolates, all PCR products were sequenced in forward and reverse directions by the same primers used for PCR amplification by the Beijing Genomics Institute, Guangzhou, China. All sequences obtained in this study were edited using MEGA v. 6.0.5 software [34] and were deposited in GenBank (https://www.ncbi.nlm.nih.gov).

Table 1.
Primers for amplification of tef1, tub2, cmdA, his3 and mating type gene fragments.
Table 1.
Primers for amplification of tef1, tub2, cmdA, his3 and mating type gene fragments.
| Target Gene | Primer Name | Primer Sequence (5′ to 3′) | Tm (°C) | Fragment Size (bp) | Reference |
|---|---|---|---|---|---|
| translation elongation factor 1-alpha (tef1) | EF1-728F | CATCGAGAAGTTCGAGAAGG | 52 | 500 | [30,33] |
| EF2 | GGA(G/A)GTACCAGT(G/C)ATCATGTT | [30,33] | |||
| β-tubulin (tub2) | T1 | AACATGCGTGAGATTGTAAGT | 52 | 520 | [30,33] |
| CYLTUB1R | AGTTGTCGGGACGGAAGAG | [30,33] | |||
| calmodulin (cmdA) | CAL-228F | GAGTTCAAGGAGGCCTTCTCCC | 55 | 470 | [30,33] |
| CAL-2Rd | TGRTCNGCCTCDCGGATCATCTC | [30,33] | |||
| histone H3 (his3) | CYLH3F | AGGTCCACTGGTGGCAAG | 55 | 450 | [30,33] |
| CYLH3R | AGCTGGATGTCCTTGGACTG | [30,33] | |||
| MAT1-1-1 | Cal_MAT111_F | ATGCTTCCTCAGTCTTTGCT | 53 | 330 | [35] |
| Cal_MAT111_R | CTTGAAYRGGGTTGGTGG | [35] | |||
| MAT1-2-1 | Cal_MAT121_F | GCAAGGAYCGCCACCRAAT | 58 | 240 | [35] |
| Cal_MAT121_R | GACACCTCKGCGTTTCTTCTCAG | [35] |
2.3. Multi-Gene Phylogenetic Analyses
To preliminarily identify the species which the isolates obtained in this study, a standard nucleotide BLAST search was conducted using the tef1, tub2, cmdA and his3 sequences. The sequences of tef1, tub2, cmdA and his3 gene regions generated in this study were compared with sequences of type specimen strains of published Calonectria species for phylogenetic analyses. Sequences of all the published species in the relative species complexes were used for sequence comparisons and phylogenetic analyses. The datasets of Liu and co-authors [28] were used as templates for analyses.
Sequences of each of the tef1, tub2, cmdA and his3 gene regions as well as the combination of these four gene regions were aligned using the online version of MAFFT v. 7 (http://mafft.cbrc.jp/alignment/server) with the alignment strategy FFT-NS-i (Slow; interactive refinement method). After initial alignments, sequence alignments were manually edited using MEGA v. 6.0.5 software [34].
Maximum parsimony (MP) and maximum likelihood (ML) were used frequently for phylogenetic analyses of Calonectria species [28,29,36]. To test whether the analysis results between the two methods are consistent, both MP and ML were used for phylogenetic analyses for sequence datasets of each of the four genes and the combination of four gene regions. The MP and ML analyses were conducted using the methods described by Liu and Chen [33]. Phylogenetic trees were viewed using MEGA v. 6.0.5 [34]. Sequence data of two isolates of Curvicladiella cignea (CBS 109167 and CBS 109168) were used as outgroups [28].
2.4. Morphology
The representative isolates of each Calonectria species identified by DNA sequence comparisons were selected for morphological description. The size of macroconidia and width of vesicles are the most typical asexual characteristics used for morphological comparison in Calonectria [19,29,33]. The asexual structures of selected Calonectria isolates were induced in synthetic nutrient-poor agar (SNA) [37] following the method described by Liu and Chen [33]. Fifty measurements of macroconidia and vesicles were made for the selected isolates.
2.5. MAT Gene Amplification and Mating Type Assignment
To further understand the possible reproductive mode of the population of each Calonectria species identified in this study, the mating type idiomorph of each representative Calonectria isolate of an identified species was identified. Mating type primer pairs Cal_MAT111_F/Cal_MAT111_R and Cal_MAT121_F/ Cal_MAT121_R were used to amplify the MAT1-1-1 and MAT1-2-1 genes in all selected isolates using the protocol described by Li and co-authors [35] (Table 1). For the heterothallic Calonectria species based on MAT gene amplification results, the species with an adequate number of isolates as one population, the possibility that recombination had taken place was tested. A two-tailed exact binomial method [38] was used, and each population that included isolates of both mating types was tested in R version 3.6.1 to evaluate whether the MAT1-1 and MAT1-2 frequency significantly deviated from a 1:1 ratio.
2.6. Pathogenicity Tests
To determine the pathogenicity of Calonectria species obtained in this study, representative isolates of all Calonectria species isolated from diseased leaves and soils were identified by phylogenetic analyses, and morphological characteristics were selected for inoculation trials. Two Eucalyptus genotypes, E. urophylla × E. tereticornis hybrid genotype CEPT1876 and E. urophylla × E. grandis hybrid genotype CEPT1877 were selected for inoculations. The inoculated Eucalyptus seedlings were three months old and approximately 40 cm tall.
In this study, the inoculations were conducted with both mycelia plug and conidia suspension of selected Calonectria isolates. All the inoculated seedlings were in similar size. In the mycelia plug inoculations, for each Eucalyptus genotype, mycelia plugs of each isolate were inoculated on ten leaves of two to three Eucalyptus seedlings, and ten leaves of other two to three Eucalyptus seedlings treated with sterile MEA plugs were regarded as negative controls. For inoculation, mycelia plugs (5 mm diameter) from 7-day-old MEA cultures were placed upside down on the abaxial surface of the leaflets. In the conidia suspension inoculations, the conidia suspensions for each isolate were prepared using the method described in Graça and co-authors [39] and Wang and Chen [20]. The conidia suspensions prepared for each isolates were measured using a hemocytometer, being the concentration adjusted to 5 × 104 conidia/mL. For each isolate, eight seedlings of each genotype were inoculated by spraying the conidia suspension until the suspension run off the leaves. Sterile water was sprayed onto other eight seedlings as the negative control using the same treatment. To allow sufficient humidity for infection development, the Eucalyptus seedlings inoculated with mycelia plug and conidia suspension, were maintained in plastic chambers (length: 190 cm, width: 90 cm, height: 63 cm) with intermittent water nebulization for 30 s at three-hour intervals and were maintain stable climatic conditions (temperature 24–26 °C; humidity 60–70%) for three days. The experiments using both mycelia plug and conidia suspension were all repeated once using the same methodology.
The plastic chambers were removed three days after inoculation. For mycelia plug inoculations, the length of lesions produced was measured. For conidia suspension inoculations, the disease index (DI) was calculated. Leaf disease severity was assessed by estimating the percentage of lesioned area on each leaf with a scale from 0 to 5, where 0 indicated no lesions, 1 indicated that 1 to 10% area of the leaf was lesioned, 2 indicated that 11 to 25% area of the leaf was lesioned, 3 indicated that 26 to 50% area of the leaf was lesioned, 4 indicated that 51 to 75% area of the leaf was lesioned, and 5 indicated that 76 to 100% area of the leaf was lesioned. The DI was calculated according to Mishra and co-authors [40]. The percentage of lesioned area caused by Calonectria isolate on each leaf of inoculated Eucalyptus seedlings was calculated through the software “Leaf Doctor” [41].
For re-isolations, small pieces of discolored leaf (approximately 0.04 cm2) from the edges of the resultant lesions were cut and placed on 2% MEA at room temperature. Re-isolations were conducted for randomly selected leaves from four randomly selected seedlings of each Eucalyptus genotype for each inoculated isolate, and the randomly selected leaves from all seedlings were inoculated as negative controls. Re-isolations were conducted for both mycelia plug and conidia suspension inoculations. The re-isolated fungi were identified and confirmed by morphological characteristics of culture, macroconidiophore and macroconidia, as well as the disease symptoms produced on the leaves with the original fungi used for inoculations. Statistical analyses were performed using SPSS Statistics 22 software (IBM Corp., Armonk, NY, USA) by one-way analysis of variance (ANOVA) for mycelia plug and conidia inoculation results, respectively. The inoculations were conducted in September, 2020 at the experimental nursery of China Eucalypt Research Centre, GuangDong Province, China.
3. Results
3.1. Disease Symptoms, Sample Collection and Fungal Isolation
Disease symptoms observed in the present study include greyish water-soaked spots on the leaves of the lower branches on the infected trees in the early stage (Figure 1). These spots subsequently form extensive necrotic areas, and leaves become dry and curly (Figure 1A,B,E,F). White masses of conidiophores with typical morphological characteristics of Calonectria species are frequently observed on the shoots and leaves of Eucalyptus trees. Different Eucalyptus genotypes infected by pathogens show different degrees of susceptibility and symptoms (Figure 1). The disease symptoms observed in this study were similar to those caused by Calonectria species in Eucalyptus trees as reported previously in China [20]. These samples of diseased leaves and soils were transported to the laboratory for isolation, morphological examination and further molecular research. Diseased leaf samples were collected from 13 to 20 trees of each of ten Eucalyptus genotypes. A total of 190 diseased leaf samples with white masses of conidiophores with typical morphological characteristics of Calonectria species were obtained from 190 diseased trees; furthermore, 190 soil samples were collected from soils under these trees (Table 2). For diseased leaf samples, Calonectria was successfully isolated from all sampled diseased trees of each of the ten Eucalyptus genotypes, with the exception of Eucalyptus genotypes CEPT1862, CEPT1863, CEPT1865 and CEPT1866. Finally, Calonectria was isolated from 184 of the 190 diseased trees (Table 2). One to two Calonectria isolates, depending on the variation of conidia morphology, from each sampled tree were isolated, and a total of 186 Calonectria isolates were obtained from sampled trees (Table 2). For soil samples, no Calonectria was isolated from soils sampled under Eucalyptus genotype CEPT1863; Calonectria isolates were obtained from two to 12 soil samples collected from the soil under each of the other nine Eucalyptus genotypes. In total, Calonectria was isolated from 47 of the 190 soil samples (Table 2). One to six Calonectria isolates were isolated from each of the soil samples in which Calonectria was induced and sporulated. In all, 182 Calonectria isolates were obtained from the 47 soil samples (Table 2). Three hundred and sixty-eight Calonectria isolates were obtained from diseased Eucalyptus trees and soils under these trees (Table 2).
Table 2.
Samples and isolates of Calonectria obtained from ten Eucalyptus genotypes in this study.
3.2. Sequencing
Sixty-three Calonectria isolates obtained from diseased trees of ten Eucalyptus genotypes (four to 12 isolates from each Eucalyptus genotype), and all 182 isolates obtained from soils were used for DNA extraction and sequence comparisons (Appendix A Table A1). The tef1 and tub2 genes were amplified for all 245 isolates. Subsequently, 73 representative isolates were selected based on tef1 and tub2 sequences so as to include all the genotypes revealed by these two loci, as well as all the sampled Eucalyptus genotypes and substrate. The cmdA and his3 loci were then also sequenced for these 73 isolates (Appendix A Table A1).
3.3. Multi-Gene Phylogenetic Analyses
The sequence fragments were approximately 500 bp for tef1, 565 bp for tub2, 685 bp for cmdA and 440 bp for his3. Based on the sequences of tef1, tub2, cmdA and his3 loci, the 73 representative isolates represented 11 genotypes. Forty isolates representing all 11 genotypes that were isolated from the diseased leaves and soils associated with all the relative Eucalyptus genotypes were selected for phylogenetic analyses (Appendix A Table A1). Results of the standard nucleotide BLAST search conducted using the tef1, tub2, cmdA and his3 sequences showed that the isolates obtained in the current study belong three species complexes of Calonectria, including C. reteaudii species complex, the C. cylindrospora species complex, and the C. kyotensis species complex. Based on the recently published results in Liu and co-authors [28], sequences of tef1, tub2, cmdA and his3 published species in the C. reteaudii species complex, C. cylindrospora species complex and C. kyotensis species complex, respectively, were used for sequence comparisons and phylogenetic analyses (Appendix A Table A2).
The partition homogeneity test (PHT) comparing the combination of tef1, tub2, cmdA and his3 gene datasets generated a p-value of 0.001, indicating the accuracy of the combined datasets did not suffer relative to the individual partitions [42], sequences of the four loci were combined for analyses. For the phylogenetic trees based on tef1, tub2, cmdA and his3 individually and the combined sequence datasets, the overall topologies were similar, but the relative position of some Calonectria species differed slightly between the MP and ML trees. The five ML trees are presented in Figure 2, Supplementary Figures S1–S4. The numbers of taxa and parsimony informative characters, statistical values for the MP analyses, and parameters for the best-fit substitution models of ML analyses are provided in Table 3.
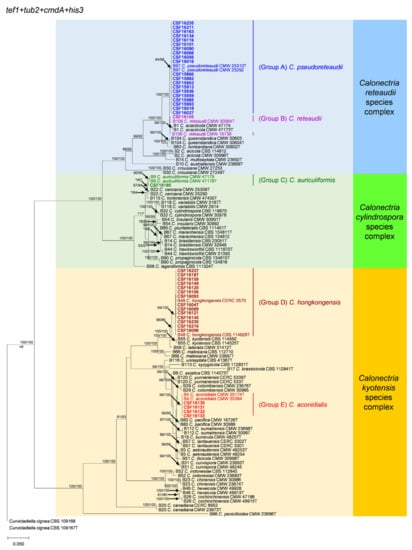
Figure 2.
Phylogenetic tree of Calonectria species based on maximum likelihood (ML) analyses of the dataset of combined tef1, tub2, cmdA and his3 gene sequences in this study. Bootstrap support values ≥70% are presented above the branches as follows: ML/MP. Bootstrap values <70% and absent are marked with “*”. Isolates highlighted in five different colors and bold were obtained in this study. Ex-type isolates are marked with “T”. The “B” species codes are consistent with the recently published results in Liu and co-authors [28]. The Curvicladiella cignea (CBS 109167 and CBS 109168) was used as outgroup taxon.
Table 3.
Statistical values of datasets for maximum parsimony and maximum likelihood analyses in this study.
Sequence data were not available for tub2 for four isolates (CSF16130, CSF16131, CSF16132 and CSF16133) obtained in this study and ex-type isolates of various published Calonectria species (Appendix A Table A1 and Table A2). The 40 Calonectria isolates were clustered in five groups (Group A, Group B, Group C, Group D and Group E) based on tef1, his3 and combined tef1/tub2/cmdA/his3 analyses (Figure 2; Supplementary Figures S1 and S4). These isolates clustered in four groups based on tub2 analyses, including Groups A, B, C, and D, where sequences of Group E were not available (Supplementary Figure S2); and four groups were based on cmdA analyses, including Groups C, D and E, where Group A clustered with Group B (Supplementary Figure S3).
The phylogenetic analyses showed that isolates in Group A and Group B belong the C. reteaudii species complex. Isolates in Group A clustered with C. pseudoreteaudii based on phylogenetic analyses of tef1, tub2 and his3 datasets (Supplementary Figures S1, S2 and S4) and clustered with C. pseudoreteaudii and C. reteaudii in the cmdA tree (Supplementary Figure S3). In the combined tef1/tub2/cmdA/his3 tree, these isolates were clustered with C. pseudoreteaudii (Figure 2). Isolates in Group A were identified as C. pseudoreteaudii. Isolate CSF16105 in Group B clustered with C. reteaudii in the tub2 and his3 trees (Supplementary Figures S2 and S4), with C. reteaudii and C. acaciicola in the tef1 tree (Supplementary Figure S1), and with C. reteaudii and C. pseudoreteaudii in the cmdA tree (Supplementary Figure S3). The isolate was clustered with C. reteaudii in the combined tef1/tub2/cmdA/his3 tree (Figure 2). The isolate in Group B was identified as C. reteaudii.
Isolate CSF16185 in Group C belongs the C. cylindrospora species complex. This isolate clustered with C. auriculiformis in the tef1 tree (Supplementary Figure S1); clustered with C. cerciana and was closely related to C. tonkinensis, C. lageniformis and C. auriculiformis in the tub2 trees (Supplementary Figure S2); clustered with C. lageniformis and was closely related to C. cerciana, C. tonkinensis and C. auriculiformis in the cmdA tree (Supplementary Figure S3) and clustered with C. auriculiformis, C. cerciana and C. tonkinensis in the his3 tree (Supplementary Figure S4). This isolate was most closely related to C. auriculiformis in the combined tef1/tub2/cmdA/his3 tree (Figure 2). The Isolate in Group C was identified as C. auriculiformis.
All the isolates in Group D and Group E belong the C. kyotensis species complex. Isolates in Group D were clustered with C. hongkongensis in each of the tef1, tub2, cmdA, his3 and the combined tef1/tub2/cmdA/his3 trees (Figure 2, Supplementary Figures S1–S4). These isolates were identified as C. hongkongensis. The tub2 sequences are not available for the four isolates in Group E; these isolates were clustered with C. aconidialis in each of the tef1, cmdA, his3 and the combined tef1/tub2/cmdA/his3 trees (Figure 2, Supplementary Figures S1, S3 and S4). Isolates in Group E were identified as C. aconidialis.
3.4. Species and Genetic Diversity Associate with Eucalyptus Genotype and Substrate
Based on the sequence comparisons of tef1, tub2, cmdA and his3 sequences, the Calonectria isolates sequenced were identified as C. hongkongensis (123), C. pseudoreteaudii (116), C. aconidialis (4), C. reteaudii (1) and C. auriculiformis (1). With the exception of C. pseudoreteaudii, which was isolated from both Eucalyptus diseased leaves and soils, the other four species were only obtained from soils under the Eucalyptus trees (Appendix A Table A1). Sixty-three isolates of C. pseudoreteaudii were isolated from diseased leaves from all the ten sampled Eucalyptus genotypes; 53 isolates of C. pseudoreteaudii and all the 123 isolates of C. hongkongensis were isolated from soils under the same nine of the ten Eucalyptus genotypes (except for CEPT1863) (Appendix A Table A1). Calonectria reteaudii, C. auriculiformis and C. aconidialis were only isolated from Eucalyptus genotypes CEPT1864, CEPT1868 and CEPT1865, respectively (Appendix A Table A1).
The genotypes of Calonectria species identified were determined by sequences of tef1, tub2, cmdA and his3; the results indicated they were generated from C. pseudoreteaudii (2), C. reteaudii (1), C. auriculiformis (1), C. hongkongensis (6) and C. aconidialis (1) (Appendix A Table A1, Table 4). For C. pseudoreteaudii, 115 of all 116 isolates presented the same genotype (Genotype 1 of C. pseudoreteaudii) (Appendix A Table A1), which was distributed in diseased leaves of all ten Eucalyptus genotypes and soils under nine Eucalyptus genotypes; only one isolate presented a different genotype (CSF16016, Genotype 2 of C. pseudoreteaudii), which was isolated from diseased leaves of CEPT1868 (Appendix A Table A1, Table 4). Ninety-eight of the 123 isolates of C. hongkongensis presented the same genotype (C. hongkongensis Genotype 1). This genotype was dominant in C. hongkongensis, and these 98 isolates were isolated from soils under eight of the ten Eucalyptus genotypes (Appendix A Table A1, Table 4).
Table 4.
Species and genetic diversity of Calonectria associated with Eucalyptus genotype and substrate.
3.5. Morphology
Based on phylogenetic analysis results, 16 isolates represented five Calonectria species (C. pseudoreteaudii: CSF15985, CSF16016, CSF16018, CSF16027, CSF16056, CSF16102 and CSF16116; C. reteaudii: CSF16105; C. auriculiformis: CSF16185; C. hongkongensis: CSF16121, CSF16145, CSF16230 and CSF16237; C. aconidialis: CSF16130, CSF16131 and CSF16133) identified in this study were selected for macroconidia and vesicle morphological comparisons (Appendix A Table A1, Table 5). These isolates can be distinguished into three groups based on the vesicle shape. Isolates of C. pseudoreteaudii and C. reteaudii produce clavate or narrowly clavate vesicles; the vesicles of C. auriculiformis are ellipsoidal to fusiform to obpyriform, and the vesicles of C. hongkongensis and C. aconidialis are pyriform to sphaeropedunculate, ovoid to sphaeropedunculate and sphaeropedunculate. With the exception of C. reteaudii and C. auriculiformis, in which only one isolate was studied for each of the two species, the average of measurements showed that significant variations exist in the size of macroconidia or width of vesicles among isolates of each species of C. pseudoreteaudii, C. hongkongensis, and C. aconidialis were observed (Table 5). For example, the macroconidia of C. pseudoreteaudii isolates CSF15985, CSF16016, CSF16018, and CSF16027 were relatively much shorter than those of the other three tested C. pseudoreteaudii isolates CSF16056, CSF16102 and CSF16116 (Table 5); the vesicles of C. hongkongensis isolates CSF16145 and CSF16237 were much wider than those of isolates CSF16121 and CSF16230 (Table 5); in C. aconidialis, the vesicles of isolate CSF16130 were much longer than those of isolate CSF16131 (Table 5). The average of measurements further showed that no significant variations also exist in the size of macroconidia or width of vesicles among all isolates of each species of C. pseudoreteaudii, C. hongkongensis, or C. aconidialis were observed (Table 5). For example, the width of vesicles among seven isolates of C. pseudoreteaudii were similar, and there were no major differences in macroconidia size among isolates of C. hongkongensis and C. aconidialis (Table 5).
For each of the five Calonectria species found in this study, the shape of vesicle and septate number of macroconidia among isolates obtained during the current study and the originally described strains were consistent (Table 5). The measurements showed that the macroconidia size and vesicles width of isolates obtained in the current study and the originally described strains of the same Calonectria species were not always similar; for example, the macroconidia lengths of C. pseudoreteaudii isolates obtained in this study were much shorter than the originally described strains of C. pseudoreteaudii [30], and the macroconidia length of C. hongkongensis isolates obtained in the current study was shorter than the originally described C. hongkongensis strains [43] (Table 5). For each species of C. reteaudii and C. auriculiformis, the macroconidia size of isolates obtained in the current study were similar to the originally described strains of relative species [36,44] (Table 5). The vesicle measurements showed that the vesicle widths of C. hongkongensis isolates obtained in the current study were much shorter than those of the originally described strains of C. hongkongensis [43], while for C. pseudoreteaudii, C. reteaudii and C. auriculiformis, the vesicle widths of isolates obtained in the current study were similar to the originally described strains of relative species [30,36,44] (Table 5).
Table 5.
Morphological comparisons of Calonectria isolates and species obtained in the current study.
Table 5.
Morphological comparisons of Calonectria isolates and species obtained in the current study.
| Species | Isolate/Species | Macroconidia (L × W) 1,2,3 | Macroconidia Average (L × W) 1,2 | Macroconidia Septation | Vesicle Width 1,2,3 | Vesicle Width Average 1 |
|---|---|---|---|---|---|---|
| C. pseudoreteaudii | Isolate CSF15985 (this study) | (69–)75.5–85(–88) × (6.5–)7–8(–8.5) | 80.5 × 7.5 | (3–)5(–6) | (2–)2.5–3.5(–4) | 3 |
| Isolate CSF16016 (this study) | (71–)76–88.5(–98) × (6–)6.5–7.5(–8) | 82.5 × 7 | (3–)5 | (2–)2–3.5(–4) | 3 | |
| Isolate CSF16018 (this study) | (75.5–)79–87.5(–94.5) × (6.5–)7–8(–9) | 83 × 7.5 | 5 | (2–)2–3.5(–4) | 2.5 | |
| Isolate CSF16027 (this study) | (71–)78–89.5(–95) × (6–)6.5–8(–9) | 84 × 7.5 | 5 | (2–)2.5–3.5(–4.5) | 3 | |
| Isolate CSF16056 (this study) | (77.5–)87–104.5(–112.5) × (6–)7–8(–9) | 96 × 7.5 | 5 | (3–)3.5–4.5(–5.5) | 4 | |
| Isolate CSF16102 (this study) | (84–)87.5–97.5(–105) × (6–)6.5–8(–10) | 92.5 × 7.5 | 5(–7) | (2–)2.5–3.5(–4) | 3 | |
| Isolate CSF16116 (this study) | (76–)84.5–96.5(–104) × (5.5–)7.5–8.5(–10) | 90.5 × 8 | 5 | (2–)2.5–4(–4.5) | 3 | |
| Species (this study) | (69–)79–95(–112.5) × (5.5–)7–8(–10) | 87 × 7.5 | (3–)5(–7) | (2–)2.5–4(–5.5) | 3 | |
| Species [30] | (88–)96–112(–119) × 7–9(–10) | 104 × 8 | 5(–8) | 3–5 | N/A 4 | |
| C. reteaudii | Isolate CSF16105 (this study) | (71.5–)77.5–87.5(–92.5) × (6–)6.5–7.5(–8) | 82.5 × 7 | 5 | (3.5–)4–5(–6) | 4.5 |
| Species [44] | (50–)75–95(–120) × (5–)6–7 | 84 × 6.5 | (1–)5(–6) | (3–)5(–6) | N/A | |
| C. auriculiformis | Isolate CSF16185 (this study) | (34–)36.5–43.5(–47) × (3.5–)4–5(–5.5) | 40 × 4.5 | 1 | (4.5–)6–9.5(–12) | 7.5 |
| Species [36] | (40–)41–45(–47) × (3–)4–5 | 43 × 4 | 1 | 6–12 | N/A | |
| C. hongkongensis | Isolate CSF16121 (this study) | (34.5–)36.5–41(–46) × (3.5–)3.5–4.5(–4.5) | 39 × 4 | 1 | (3–)3–6(–12) | 4.5 |
| Isolate CSF16145 (this study) | (36–)37–40(–42.5) × (4–)4–4.5(–5) | 38.5 × 4.5 | 1 | (5–)5.5–8.5(–10.5) | 7 | |
| Isolate CSF16230 (this study) | (35–)37–40.5(–43.5) × (4–)4–4.5(–5) | 38.5 × 4.5 | 1 | (3–)3.5–5(–5.5) | 4 | |
| Isolate CSF16237 (this study) | (35–)37–41.5(–43) × (3.5–)4–4.5(–5) | 39 × 4 | 1 | (3.5–)4.5–8(–11) | 6.5 | |
| Species (this study) | (34.5–)36.5–41(–46) × (3.5–)4–4.5(–5) | 40 × 4 | 1 | (3–)3.5–6.5(–12) | 5 | |
| Species [43] | (38–)45–48(–53) × 4(–4.5) | 46.5 × 4 | 1 | 8–14 | N/A | |
| C. aconidialis | Isolate CSF16130 (this study) | (35–)40–46.5(–50.5) × (4–)4–5(–5.5) | 43 × 4.5 | 1 | (5–)5.5–9.5(–13) | 7.5 |
| Isolate CSF16131 (this study) | (37–)42–48.5(–53) × (4–)5–5.5(–6) | 45.5 × 5.5 | 1 | (3–)2.5–6(–12) | 4.5 | |
| Isolate CSF16133 (this study) | (37–)41.5–47(–51) × (3.5–)4–5(–5.5) | 44.5 × 4.5 | 1 | (3.5–)4.5–7.5(–10) | 6 | |
| Species (this study) | (35–)41–47.5(–53) × (3.5–)4–5.5(–6) | 44 × 5 | 1 | (3–)4–8(–13) | 6 | |
| Species [29] | N/A | N/A | N/A | N/A | N/A |
1 All measurements are in µm. 2 L × W = length × width. 3 Measurements are presented in the format [(minimum–) (average – standard deviation) – (average + standard deviation) (–maximum)]. 4 N/A represents data that is not available.
3.6. MAT Gene Amplification and Mating Type Assignment
Only C. pseudoreteaudii was isolated from both diseased leaves and soils under the sampled trees, to understand the differences of mating type assignment between isoaltes from diseased leaves and soils, all the C. pseudoreteaudii isolates were selected for MAT gene amplification. All the isolates identified as C. reteaudii, C. auriculiformis and C. aconidialis were used for MAT gene amplification, since only six isolates were identified as these species. Few isoaltes of C. hongkongensis were selected for amplification, since both the MAT1-1-1 and MAT1-2-1 genes were successfully amplified for all the selected isolate during the the preliminary MAT gene amplification. One hundred and thirty-four Calonectria isolates were selected to amplify the MAT1-1-1 and MAT1-2-1 genes. These included all 116 isolates of C. pseudoreteaudii sequenced for multiple-gene sequence identification (63 isolates from diseased leaves of all ten sampled Eucalyptus genotypes, 53 isolates from soils under nine Eucalyptus genotypes), 12 random selected isolates of C. hongkongensis, and all the six isolates of C. reteaudii (CSF16105), C. auriculiformis (CSF16185) and C. aconidialis (CSF16130, CSF16131, CSF16132 and CSF16133) obtained in the current study (Appendix A Table A1, Figure 3). The mating type idiomorphs were successfully amplified in all 134 Calonectria isolates (Appendix A Table A1). Each isolate was identified by positive amplification of a 350 bp fragment MAT1-1-1 and/or a 270 bp MAT1-2-1 product. Only the MAT1-1-1 or MAT1-2-1 gene was successfully amplified for each isolate of C. pseudoreteaudii confirming the heterothallic nature of the species (Figure 3). Calonectria reteaudii isolate CSF16105 had only the MAT1-1 mating type, and C. auriculiformis isolate CSF16185 had only the MAT1-2 mating type (Figure 3), suggesting that both C. reteaudii and C. auriculiformis are likely to be heterothallic species. All the amplified isolates of C. hongkongensis and C. aconidialis had both the MAT1-1 and MAT1-2 mating types, confirming that they are homothallic species (Figure 3).
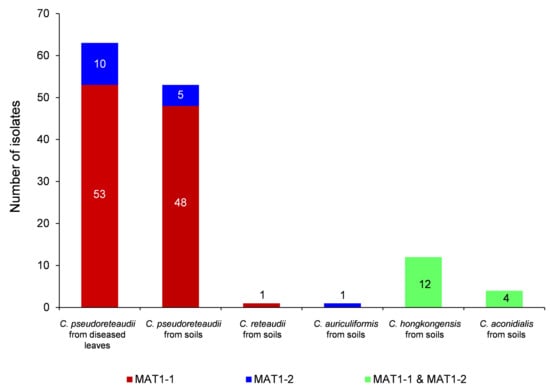
Figure 3.
Bar graph showing the distribution of mating type idiomorphs (MAT1-1, MAT1-2, and MAT1-1 and MAT1-2) in five species of Calonectria.
When considering the mating gene diversity of each Calonectria species, only one genotype of MAT1-1-1 gene was successfully amplified for 101 (53 from diseased leaves, 48 from soils) of the 116 C. pseudoreteaudii isolates, two genotypes of MAT1-2-1 gene for ten isolates from diseased leaves, and one genotype of MAT1-2-1 gene for five isolates from soil (this genotype is the same as one of the two genotypes from diseased leaves) (Appendix A Table A1, Figure 3). Two genotypes for each of MAT1-1-1 and MAT1-2-1 genes were amplified for 12 C. hongkongensis isolates from soils (Appendix A Table A1, Figure 3). For the four C. aconidialis isolates, only one genotype for each of the MAT1-1-1 and MAT1-2-1 genes was amplified (Appendix A Table A1, Figure 3).
For the three heterothallic or putative heterothallic species, C. pseudoreteaudii, C. reteaudii and C. auriculiformis, only C. pseudoreteaudii had an adequate number of isolates as a population to test whether recombination took place. The mating type frequencies of MAT1-1 and MAT1-2 for the C. pseudoreteaudii isolates from diseased leaves (63 isolates), soils under Eucalyptus trees (53 isolates), and from diseased leaves and soils (116 isolates) were 5.3, 9.6 and 6.7, respectively, which all significantly deviated from a 1:1 ratio (p < 0.01), implying that the asexual cycle represents the primary reproductive mode in C. pseudoreteaudii in the sampled area in the current study (Appendix A Table A1, Figure 3).
3.7. Pathogenicity Tests
Twelve isolates representing five Calonectria species (C. pseudoreteaudii: CSF15985, CSF16016, CSF16027, CSF16056 and CSF16116; C. reteaudii: CSF16105; C. auriculiformis: CSF16185; C. hongkongensis: CSF16145, CSF16230 and CSF16237; and C. aconidialis: CSF16130 and CSF16133) were selected to inoculate on seedlings of two Eucalyptus genotypes CEPT1876 and CEPT1877 using mycelia plugs (Appendix A Table A1, Figure 4).
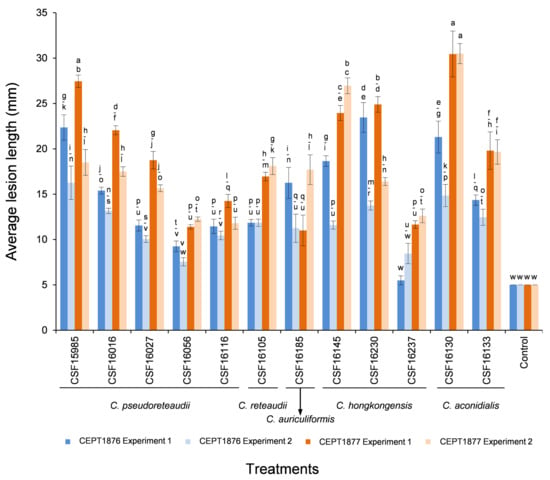
Figure 4.
Column chart indicating the average lesion length (mm) on leaves resulting from mycelia plug inoculation trials of two Eucalyptus hybrid genotypes inoculated with five Calonectria species and the controls; two experiments were conducted. Vertical bars represent standard error of means. Bars topped with different letters indicate treatment means that are significantly different (p = 0.05).
For conidia suspension inoculations, eight isolates representing five Calonectria species (C. pseudoreteaudii: CSF15985, CSF16027, CSF16056 and CSF16116; C. reteaudii: CSF16105; C. auriculiformis: CSF16185; C. hongkongensis: CSF16230; and C. aconidialis: CSF16130) that produced abundant masses of conidiophores and macroconidia were selected to inoculate on seedlings of the two Eucalyptus genotypes (Appendix A Table A1, Figure 5).
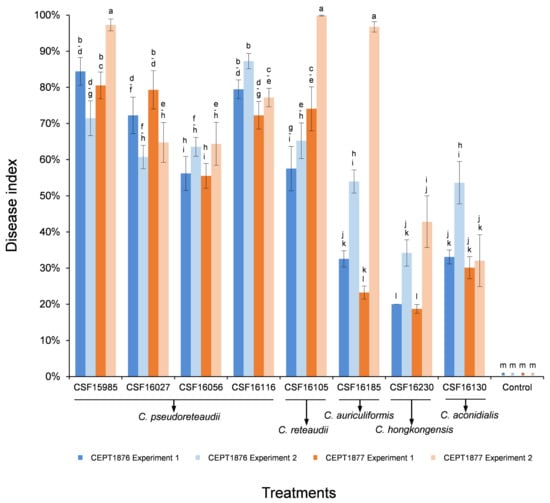
Figure 5.
Column chart indicating the disease index (%) resulting from conidia suspension inoculation trials of two Eucalyptus hybrid genotypes inoculated with five Calonectria species and the controls, two experiments were conducted. Vertical bars represent standard error of means. Bars topped with different letters indicate that treatment means are significantly different (p = 0.05). The “*” indicates that the disease indexes of negative controls are zero.
The mycelia plugs of all 12 tested isolates produced leaf spot/lesion symptoms on leaves (Figure 6A,B), while no lesions were observed on the negative control inoculations (Figure 6C,D). All seedlings of the two Eucalyptus genotypes inoculated with the conidia suspension of each of the eight Calonectria isolates developed leaf spot and shoot blight symptoms (Figure 6M,N), while no disease symptoms were observed on the leaves and shoots of the negative control seedlings (Figure 6O,P). The Calonectria species with the same morphological characteristics as the originally inoculated fungi were successfully re-isolated from diseased tissues on the inoculated leaves, but never from the negative control seedlings, thus fulfilling the requirements of Koch’s postulates.

Figure 6.
Symptoms on seedlings of E. urophylla × E. tereticornis hybrid genotype CEPT1876 and E. urophylla × E. grandis hybrid genotype CEPT1877 inoculated by Calonectria mycelia plugs/MEA plugs (A–L) and conidia suspensions/sterile water (M–T) of five Calonectria isolates. (A): CEPT1876 inoculated by isolate CSF16130 (C. aconidialis); (B): CEPT1877 inoculated by isolate CSF16016 (C. pseudoreteaudii); (C,D): No disease symptoms were observed on leaves of CEPT1876 (C) and CEPT1877 (D) inoculated by sterile MEA plugs (negative controls); (E): Isolate CSF16130 (C. aconidialis) produced the longest lesions on CEPT1877; (F): Long lesions produced by isolate CSF16145 (C. hongkongensis) on CEPT1877; (G,H): Isolate CSF16230 (C. hongkongensis) produced long lesions on CEPT1876 (G) and CEPT1877 (H); (I,J): Isolate CSF15985 (C. pseudoreteaudii) produced significantly larger lesions on CEPT1877 than that of isolate CSF16056 (C. pseudoreteaudii); (K,L): The inoculation results of isolate CSF16016 (C. pseudoreteaudii) indicated that CEPT1876 was significantly more tolerant than CEPT1877; (M): Lesions on leaves of CEPT1877 inoculated by isolate CSF16105 (C. reteaudii); (N): Genotype CEPT1877 rotted after inoculations by isolate CSF16130 (C. aconidialis), abundant white mass of conidiophores were observed; (O,P): No disease symptoms on CEPT1876 (O) and CEPT1877 (P) inoculated by sterile water (negative controls); (Q,R): All leaves of CEPT1877 blighted and the seedling died after infection by isolates CSF15985 (Q) (C. pseudoreteaudii) and CSF16185 (R) (C. auriculiformis); (S,T): Lesions on CEPT1876 (S) and CEPT1877 (T) inoculated by isolate CSF16130 (C. aconidialis). (A–L,M–P,S,T) are in the first experiment; (Q,R) are in the second experiment.
The data of lesion length and disease index resulting from mycelia plug and conidia suspension were not normally distributed based on a Kolmogorov–Smirnov normality test (p < 0.05) in SPSS v. 22.0. Thus, all the data were transformed (Kolmogorov–Smirnov normality test, p = 0.2) by conducting a Rank transformation using the statistical package SPSS v. 22.0. There were significant differences (p < 0.05) between the results of the two experiments of mycelia plug inoculations. The results of two experiments of conidia suspension inoculations also significant differences (p < 0.05). This may be due to inconsistent climatic and seedling conditions during the two experiments for each inoculation of mycelia plugs and conidia suspensions. Therefore, the data of each experiment were separately analyzed.
For the mycelia plug inoculations, the two experiments consistently showed that the lesions produced by most isolates in both experiments were significantly larger than the mycelia plug diameter (p < 0.05), with the exception of isolate CSF16056 (C. pseudoreteaudii) on Eucalyptus genotype CEPT1876 in Experiment Two, and isolate CSF16237 (C. aconidialis) on CEPT1876 in both experiments (Figure 4). The analyses of average lesion length showed that the longest lesions were produced by isolate CSF16130 (C. aconidialis) on Eucalyptus genotype CEPT1877 in both experiments (Figure 4 and Figure 6). Overall, isolates CSF15985 (C. pseudoreteaudii), CSF16145, CSF16230 (C. hongkongensis), and CSF16130 (C. aconidialis) produced relatively larger lesions than other isolates on the two tested Eucalyptus in the two experiments (Figure 4 and Figure 6E–I). The results showed that the average lesion length caused by some isolates of the same Calonectria species was significantly different; for example, on each of the two Eucalyptus genotypes in both experiments, the lesions produced by isolate CSF15985 were significantly larger than those of isolates CSF16056 and CSF16116 (C. pseudoreteaudii) (p < 0.05) (Figure 6I,J), and isolate CSF16145 produced significantly larger lesions than isolate CSF16237 (C. hongkongensis) (p < 0.05) (Figure 4). The analysis results showed that there were differences in the susceptibility of the two Eucalyptus genotypes to the tested isolates. In the two experiments, the average lesion length caused by all 12 Calonectria isolates on Eucalyptus genotype CEPT1877 was larger than those on genotype CEPT1876, with the exception of isolate CSF16185 in Experiment One (Figure 4 and Figure 6K,L). The results suggested that Eucalyptus genotype CEPT1876 is relatively more tolerant than CEPT1877 to the Calonectria species tested in this study.
For the conidia suspension inoculations, the two experiments consistently showed that more than 20% of the leaf area of both Eucalyptus genotypes were lesioned or rotted after infection by eight inoculated Calonectria isolates, with the exception of isolate CSF16230 in Experiment One (Figure 5). The analyses of average disease index showed that isolates CSF15985 (C. pseudoreteaudii), CSF16105 (C. reteaudii), and CSF16185 (C. auriculiformis) caused relatively severe disease in Eucalyptus genotype CEPT1877 in Experiment Two, with lesions covering nearly the entire leaf area (Figure 5 and Figure 6Q,R). Overall, isolates of C. pseudoreteaudii and C. reteaudii were more pathogenic than C. hongkongensis and C. aconidialis in both experiments (Figure 5). In the two experiments, the average disease indexes generated from eight Calonectria isolates on Eucalyptus genotype CEPT1877 were higher than those on genotype CEPT1876, with the exception of isolates CSF15985, CSF16056 (C. pseudoreteaudii), CSF16185 (C. auriculiformis), and CSF16230 (C. hongkongensis) in Experiment One, and isolates CSF16116 (C. pseudoreteaudii) and CSF16130 (C. aconidialis) in both experiments (Figure 5 and Figure 6S,T).
Judging from both the mycelia plug and conidia suspension inoculations, all the tested isolates of five Calonectria species produced diseased spots/lesions on the leaves of two inoculated Eucalyptus genotypes within three days; pathogenicity differences existed among isolates of the same Calonectria species; and Eucalyptus genotype CEPT1876 was relatively more tolerant than CEPT1877 to the majority of Calonectria isolates tested in this study (Figure 4 and Figure 5). The results further showed that the relative pathogenicity among five Calonectria species were not consistent between the inoculations of mycelia plug and conidia suspension. Overall, the pathogenicities among the five Calonectria species in mycelia plug inoculations were similar, while C. hongkongensis and C. aconidialis were less pathogenic than C. pseudoreteaudii and C. reteaudii in conidia suspension inoculation (Figure 4 and Figure 5).
4. Discussion
In this study, leaf disease with the typical symptoms caused by Calonectria species was observed in one Eucalyptus experimental plantation in GuangXi Province in southern China. A relatively large number of Calonectria isolates were isolated from diseased leaves of ten Eucalyptus genotypes and soils under the sampled trees in the plantation. Two hundred and forty-five isolates were identified based on DNA sequence comparisons and combined with the morphological characteristics of representative isolates. These fungi were identified as C. pseudoreteaudii, C. reteaudii, C. auriculiformis, C. hongkongensis and C. aconidialis. Calonectria pseudoreteaudii and C. hongkongensis were the dominant species, and this is the first report of C. reteaudii and C. auriculiformis isolated in China. With the exception of C. pseudoreteaudii, which was isolated from both diseased Eucalyptus leaves and soils, the other four species were only isolated from soils. For the isolates obtained from soils, C. hongkongensis (68% of the isolates from soils) was the dominant species, followed by C. pseudoreteaudii (29% of the isolates from soils). The MAT genes amplification and mating type frequency test results showed that C. pseudoreteaudii is a heterothallic species, and the asexual cycle represents the primary reproductive mode, C. reteaudii and C. auriculiformis are likely to be heterothallic species, and C. hongkongensis and C. aconidialis are homothallic species. Inoculations indicated that the five Calonectria species were all pathogenic to the two tested Eucalyptus genotypes.
The isolates obtained in this study were identified mainly based on DNA sequence comparisons of tef1, tub2, cmdA and his3 gene regions. The sequences of the four genes have been widely used to clearly distinguish between intra- and inter-specific divergence of the Calonectria genus [26,32,36]. Recently, Liu and co-authors [28] conducted a comprehensive phylogenetic analyses of the Calonectria genus based on DNA sequences of eight gene regions; the results showed that tef1 and tub2 sequences had the strongest ability to correctly identify species, followed by cmdA, his3, rpb2 and act gene regions, and that these six gene regions are effective DNA barcodes for Calonectria. The phylogenetic analyses indicated that Calonectria species could be recognized based on the tef1, tub2, cmdA and his3 gene region phylogeny [28]. The phylogenetic analysis results in this study showed that C. pseudoreteaudii and C. reteaudii belong the C. reteaudii species complex, C. auriculiformis in the C. cylindrospora species complex, and C. hongkongensis and C. aconidialis in the C. kyotensis species complex. The morphological comparisons in the current study indicated that the vesicle shape and septate number of macroconidia are relatively stable among isolates of the same species, while the morphological overlaps about shape of vesicle and septate number of macroconidia exist between Calonectria species in the same species complex (C. pseudoreteaudii and C. reteaudii; C. hongkongensis and C. aconidialis). The morphological results further showed that significant variations in macroconidia size or vesicle width exist among Calonectria isolates of the same species, which is consistent with the results of previous studies [20]. Results in this study confirmed the importance of multi-gene sequence phylogeny in species clarification and identification in Calonectria.
Calonectria pseudoreteaudii and C. hongkongensis were the two dominant species obtained in this study. Calonectria pseudoreteaudii was isolated from diseased leaves of all ten sampled Eucalyptus genotypes. Both C. pseudoreteaudii and C. hongkongensis were isolated from soils under nine of the ten Eucalyptus genotypes. Calonectria pseudoreteaudii was first isolated from diseased Eucalyptus cuttings in one nursery in GuangDong Province in southern China [30]; the fungus was further isolated from diseased Eucalyptus leaves in FuJian, GuangDong, GuangXi and HaiNan Provinces [19,28,45], and from soils under plantation Eucalyptus trees in HaiNan Province [28,29] and soils under the Eucalyptus seedlings in a nursery in GuangDong Province [19,28]. Recently, this species were widely obtained from ten Eucalyptus species and a number of E. grandis, E. tereticornis and E. urophylla hybrid genotypes in Eucalyptus plantations at 13 sites and one nursery in Leizhou Peninsula in southern China [20,28], and also from Macadamia sp. in YunNan Province [46]. The results of previous and current studies indicated that C. pseudoreteaudii may be distributed in more geographic regions and more plant hosts, both on plant tissues and in soils. Calonectria hongkongensis was first isolated from soil in Hong Kong [43]; currently, this fungus has been isolated from soils under the Eucalyptus trees in GuangXi and GuangDong and HaiNan Provinces [19,28,29], and from soils under natural forests in FuJian Province and Hong Kong [19,28]. No C. hongkongensis has been isolated from diseased tissues till now, which indicated that this species may be widely distributed in soils in different geographic regions in southern China.
Only one to four isolates were obtained for each species of C. reteaudii, C. auriculiformis and C. aconidialis in this study. Calonectria reteaudii has been reported from diseased leaves of multiple Eucalyptus species in Vietnam [23,28,36]. Calonectria auriculiformis was first reported from soils in an Acacia auriculiformis plantation in Vietnam [28,36]. Both C. reteaudii and C. auriculiformis have never been reported in China until now. Calonectria aconidialis was first isolated and described from soils under Eucalyptus plantations in GuangDong, GuangXi and HaiNan Provinces in southern China [29], and was later reported from other regions in GuangXi Province [19]. Combined with the results of previous research and current studies, C. reteaudii may cause disease in Eucalyptus trees in China, and the three species are likely distributed in soils in other un-sampled areas in southern China.
Previous research results showed that C. pseudoreteaudii [30,35] and C. reteaudii [23,44] are heterothallic species, C. auriculiformis is probably to be heterothallic [35,36], and both C. hongkongensis [35,43] and C. aconidialis [29] are homothallic species. Based on the results of MAT1-1-1 and MAT1-2-1 gene amplifications, the sexual thallism in Calonectria species was successfully detected in Li and co-authors [35]. The detection of the sexual thallism based on MAT gene amplifications in five Calonectria species in this study is supported by the results of previous studies [30,35,43].
Calonectria pseudoreteaudii was frequently isolated from diseased tissues of Eucalyptus trees and seedlings widely distributed in southern China [19,20,28], and occasionally in the soils under Eucalyptus trees and seedlings [19,29]. The results of the current study indicate that C. pseudoreteaudii is widely distributed on both diseased Eucalyptus leaves and soils in the plantation. Based on the sequences of tef1, tub2, cmdA and his3 genes as DNA barcodes and MAT1-1-1 and MAT1-2-1 for mating type determination, the genotype diversity of C. pseudoreteaudii isolates from diseased leaves is higher than the isolates from soils, and the genotypes generated from diseased leaves contain those from soils. Combined with the research results of the current and previous studies, C. pseudoreteaudii is probably propagated from diseased leaves to the soil.
Calonectria hongkongensis is the most dominant species isolated in soils in this study. This species was also frequently isolated from soils in other areas in southern China [19,29,43]. For the C. hongkongensis isolates obtained in the current study, multiple genotypes were generated based on DNA barcodes genes (six genotypes) and MAT1-1-1 (two genotypes) and MAT1-2-1 (two genotypes) genes. No C. hongkongensis was identified in diseased Eucalyptus leaves both in this and previous studies. Therefore, probably C. hongkongensis is a soil-borne species that exclusively inhabits the soil.
The results of pathogenicity tests based on mycelia plug and conidia suspension inoculations in this study showed that all tested isolates of five Calonectria species are pathogenic to the two tested Eucalyptus genotypes. This was unsurprising for C. pseudoreteaudii and C. reteaudii, since inoculations in previous studies indicated that C. pseudoreteaudii is highly pathogenic to tested Eucalyptus genotypes [20], and C. reteaudii is considered to be an important pathogen in Eucalyptus, as well as many other plants [23]. This study conducted the first pathogenicity test for C. auriculiformis, C. hongkongensis and C. aconidialis. All three species are pathogenic to the Eucalyptus genotypes tested, which is cause for concern due to their potential threat to Eucalyptus.
Conidia suspensions have been widely used to test the pathogenicities of Calonectria species [20,47,48]. However, it is difficult to induce enough conidia to use for inoculations for some isolates/species of Calonectria. The pathogenicity test results in the current study showed that relative pathogenicities among five Calonectria species are not consistent between the inoculations of mycelia plug and conidia suspension. The conidia need to germinate to develop appressoria and mycelia that penetrate plant cells; the disease symptoms of inoculations using conidia took longer to be observed, which is one of the potential reasons the results of two inoculations were not consistent.
This study expanded our understanding of the species diversity, morphological characteristics, host/substrate range, mating strategy, mating type assignment, genetic diversity and pathogenicity of Calonectria species in diseased leaves and soils in the same Eucalyptus plantation. Results indicated that there were differences in diversity and host/substrate range among species from diseased leaves and soils, differences in genetic diversity between isolates of the same species from diseased leaves and soils, and some pathogenic species were only isolated from soils but never from diseased leaves. Further studies are necessary to increase the knowledge on fungi ecological niche, the propagation pathway for these species, and the pathogenesis of these species. The inoculation results further indicated that the tolerance of different Eucalyptus genotypes are different, which highlights the importance of selecting disease resistant Eucalyptus genotypes in the future.
Supplementary Materials
The following are available online at https://www.mdpi.com/2309-608X/7/2/73/s1, Figure S1: Phylogenetic tree of Calonectria species based on maximum likelihood (ML) analyses of the tef1 gene sequences, Figure S2: Phylogenetic tree of Calonectria species based on ML analyses of the tub2 gene sequences, Figure S3: Phylogenetic tree of Calonectria species based on ML analyses of the cmdA gene sequences, Figure S4: Phylogenetic tree of Calonectria species based on ML analyses of the his3 gene sequences.
Author Contributions
S.C. conceived and designed the experiments. S.C. collected the samples. W.W. performed the laboratory work and pathogenicity tests. All authors analyzed the data. All authors wrote and revised the paper. All authors contributed to the research and approved the final version of the manuscript. All authors agree to be accountable for all aspects of the work. All authors have read and agreed to the published version of the manuscript.
Funding
This study was initiated through the bilateral agreement between the Governments of South Africa and China and supported by The National Key R&D Program of China (China-South Africa Forestry Joint Research Centre Project; project No. 2018YFE0120900), the Fundamental Research Funds for the Central Non-Profit Research Institution of CAF (Project No. CAFYBB2018QC003), the National Ten-thousand Talents Program (Project No. W03070115) and the GuangDong Top Young Talents Program (Project No. 20171172).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article and Supplementary Materials.
Acknowledgments
We thank GuoQing Li and QuanChao Wang for their assistance in collecting samples. We thank QuanChao Wang, WenWen Li, FeiFei Liu, LanSen Sun and JiaLong Han for their assistance in conducting inoculations. We thank QianLi Liu, QuanChao Wang, JieQiong Li and GuoQing Li for their assistance in analyzing the data. We thank LetPub (www.letpub.com) for providing linguistic assistance during the preparation of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Table A1.
Isolates sequenced and used for phylogenetic analyses, mating studies, morphological studies and pathogenicity tests in this study.
Table A1.
Isolates sequenced and used for phylogenetic analyses, mating studies, morphological studies and pathogenicity tests in this study.
| Identity | Genotype 1 | Isolate No. 2 | Sample No. | Host/Substrate | Collectors | Mating Gene Genotype 3 | Thallism 4 | Mating Type | GenBank Accession No. 5 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MAT1-1-1 | MAT1-2-1 | MAT1-1-1 | MAT1-2-1 | tef1 | tub2 | cmdA | his3 | ||||||||
| C. pseudoreteaudii | AA-- | CSF15865 6 | 20181021-1-(9) | Eucalyptus genotype CEPT1860 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No 7 | HE | MAT1-1 | MW290683 | No | MW285158 | MW285398 | – 8 | – |
| C. pseudoreteaudii | AAAA | CSF15866 6,9 | 20181021-1-(11) | Eucalyptus genotype CEPT1860 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290684 | No | MW285159 | MW285399 | MW290541 | MW290611 |
| C. pseudoreteaudii | AA-- | CSF15877 6 | 20181021-1-(33) | Eucalyptus genotype CEPT1860 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290685 | No | MW285160 | MW285400 | – | – |
| C. pseudoreteaudii | AA-- | CSF15879 6 | 20181021-1-(37) | Eucalyptus genotype CEPT1860 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290686 | No | MW285161 | MW285401 | – | – |
| C. pseudoreteaudii | AA-- | CSF15881 6 | 20181021-1-(41) | Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290687 | No | MW285162 | MW285402 | – | – |
| C. pseudoreteaudii | AAAA | CSF15882 6,9 | 20181021-1-(43) | Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290688 | No | MW285163 | MW285403 | MW290542 | MW290612 |
| C. pseudoreteaudii | AA-- | CSF15886 6 | 20181021-1-(51) | Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | No | MAT1-2-1_GT1 | HE | MAT1-2 | No | MW319732 | MW285164 | MW285404 | – | – |
| C. pseudoreteaudii | AA-- | CSF15887 6 | 20181021-1-(53) | Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290689 | No | MW285165 | MW285405 | – | – |
| C. pseudoreteaudii | AA-- | CSF15888 6 | 20181021-1-(55) | Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290690 | No | MW285166 | MW285406 | – | – |
| C. pseudoreteaudii | AA-- | CSF15892 6 | 20181021-1-(63) | Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290691 | No | MW285167 | MW285407 | – | – |
| C. pseudoreteaudii | AA-- | CSF15901 6 | 20181021-1-(81) | Eucalyptus genotype CEPT1862 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290692 | No | MW285168 | MW285408 | – | – |
| C. pseudoreteaudii | AAAA | CSF15903 6,9 | 20181021-1-(85) | Eucalyptus genotype CEPT1862 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290693 | No | MW285169 | MW285409 | MW290543 | MW290613 |
| C. pseudoreteaudii | AA-- | CSF15906 6 | 20181021-1-(91) | Eucalyptus genotype CEPT1862 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290694 | No | MW285170 | MW285410 | – | – |
| C. pseudoreteaudii | AA-- | CSF15908 6 | 20181021-1-(95) | Eucalyptus genotype CEPT1862 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290695 | No | MW285171 | MW285411 | – | – |
| C. pseudoreteaudii | AA-- | CSF15912 6 | 20181021-1-(105) | Eucalyptus genotype CEPT1862 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290696 | No | MW285172 | MW285412 | – | – |
| C. pseudoreteaudii | AAAA | CSF15913 6,9 | 20181021-1-(107) | Eucalyptus genotype CEPT1863 | S.F. Chen, G.Q. Li & Q.C. Wang | No | MAT1-2-1_GT1 | HE | MAT1-2 | No | MW319733 | MW285173 | MW285413 | MW290544 | MW290614 |
| C. pseudoreteaudii | AA-- | CSF15914 6 | 20181021-1-(109) | Eucalyptus genotype CEPT1863 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290697 | No | MW285174 | MW285414 | – | – |
| C. pseudoreteaudii | AA-- | CSF15916 6 | 20181021-1-(113) | Eucalyptus genotype CEPT1863 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290698 | No | MW285175 | MW285415 | – | – |
| C. pseudoreteaudii | -A-- | CSF15919 6 | 20181021-1-(119) | Eucalyptus genotype CEPT1863 | S.F. Chen, G.Q. Li & Q.C. Wang | No | MAT1-2-1_GT1 | HE | MAT1-2 | No | MW319734 | No | MW285416 | – | – |
| C. pseudoreteaudii | AA-- | CSF15922 6 | 20181021-1-(129) | Eucalyptus genotype CEPT1863 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290699 | No | MW285176 | MW285417 | – | – |
| C. pseudoreteaudii | AA-- | CSF15925 6 | 20181021-1-(137) | Eucalyptus genotype CEPT1863 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290700 | No | MW285177 | MW285418 | – | – |
| C. pseudoreteaudii | AAAA | CSF15927 6 | 20181021-1-(141) | Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290701 | No | MW285178 | MW285419 | MW290545 | MW290615 |
| C. pseudoreteaudii | AAAA | CSF15933 6 | 20181021-1-(153) | Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290702 | No | MW285179 | MW285420 | MW290546 | MW290616 |
| C. pseudoreteaudii | AAAA | CSF15936 6,9 | 20181021-1-(159) | Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290703 | No | MW285180 | MW285421 | MW290547 | MW290617 |
| C. pseudoreteaudii | AA-- | CSF15939 6 | 20181021-1-(165) | Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290704 | No | MW285181 | MW285422 | – | – |
| C. pseudoreteaudii | AA-- | CSF15942 6 | 20181021-1-(171) | Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290705 | No | MW285182 | MW285423 | – | – |
| C. pseudoreteaudii | AA-- | CSF15947 6 | 20181021-1-(181) | Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290706 | No | MW285183 | MW285424 | – | – |
| C. pseudoreteaudii | AAAA | CSF15955 6 | 20181021-1-(195) | Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290707 | No | MW285184 | MW285425 | MW290548 | MW290618 |
| C. pseudoreteaudii | AAAA | CSF15956 6 | 20181021-1-(197) | Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290708 | No | MW285185 | MW285426 | MW290549 | MW290619 |
| C. pseudoreteaudii | AAAA | CSF15959 6,9 | 20181021-1-(205) | Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290709 | No | MW285186 | MW285427 | MW290550 | MW290620 |
| C. pseudoreteaudii | AA-- | CSF15964 6 | 20181021-1-(213) | Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290710 | No | MW285187 | MW285428 | – | – |
| C. pseudoreteaudii | AAAA | CSF15965 6 | 20181021-1-(215) | Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290711 | No | MW285188 | MW285429 | MW290551 | MW290621 |
| C. pseudoreteaudii | AAAA | CSF15968 6 | 20181021-1-(221) | Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290712 | No | MW285189 | MW285430 | MW290552 | MW290622 |
| C. pseudoreteaudii | -A-- | CSF15971 6 | 20181021-1-(227) | Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290713 | No | No | MW285431 | – | – |
| C. pseudoreteaudii | AA-- | CSF15972 6 | 20181021-1-(229) | Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290714 | No | MW285190 | MW285432 | – | – |
| C. pseudoreteaudii | AAAA | CSF15985 6,9,10,11 | 20181021-1-(257) | Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290715 | No | MW285191 | MW285433 | MW290553 | MW290623 |
| C. pseudoreteaudii | AA-- | CSF15986 6 | 20181021-1-(259) | Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290716 | No | MW285192 | MW285434 | – | – |
| C. pseudoreteaudii | AA-- | CSF15987 6 | 20181021-1-(261) | Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290717 | No | MW285193 | MW285435 | – | – |
| C. pseudoreteaudii | AA-- | CSF15991 6 | 20181021-1-(269) | Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290718 | No | MW285194 | MW285436 | – | – |
| C. pseudoreteaudii | AAAA | CSF15993 6,9 | 20181021-1-(273) | Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290719 | No | MW285195 | MW285437 | MW290554 | MW290624 |
| C. pseudoreteaudii | AAAA | CSF15995 6 | 20181021-1-(277) | Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290720 | No | MW285196 | MW285438 | MW290555 | MW290625 |
| C. pseudoreteaudii | AA-- | CSF15996 6 | 20181021-1-(279) | Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290721 | No | MW285197 | MW285439 | – | – |
| C. pseudoreteaudii | AA-- | CSF15998 6 | 20181021-1-(283) | Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290722 | No | MW285198 | MW285440 | – | – |
| C. pseudoreteaudii | AA-- | CSF15999 6 | 20181021-1-(285) | Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290723 | No | MW285199 | MW285441 | – | – |
| C. pseudoreteaudii | AA-- | CSF16001 6 | 20181021-1-(289) | Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | No | MAT1-2-1_GT1 | HE | MAT1-2 | No | MW319735 | MW285200 | MW285442 | – | – |
| C. pseudoreteaudii | AAAA | CSF16007 6 | 20181021-1-(301) | Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | No | MAT1-2-1_GT1 | HE | MAT1-2 | No | MW319736 | MW285201 | MW285443 | MW290556 | MW290626 |
| C. pseudoreteaudii | AA-- | CSF16008 6 | 20181021-1-(303) | Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | No | MAT1-2-1_GT2 | HE | MAT1-2 | No | MW319737 | MW285202 | MW285444 | – | – |
| C. pseudoreteaudii | AA-- | CSF16009 6 | 20181021-1-(305) | Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | No | MAT1-2-1_GT2 | HE | MAT1-2 | No | MW319738 | MW285203 | MW285445 | – | – |
| C. pseudoreteaudii | AAAA | CSF16010 6 | 20181021-1-(307) | Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290724 | No | MW285204 | MW285446 | MW290557 | MW290627 |
| C. pseudoreteaudii | AA-- | CSF16013 6 | 20181021-1-(313) | Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290725 | No | MW285205 | MW285447 | – | – |
| C. pseudoreteaudii | AA-- | CSF16017 6 | 20181021-1-(321) | Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290726 | No | MW285207 | MW285449 | – | – |
| C. pseudoreteaudii | AAAA | CSF16018 6,9,10 | 20181021-1-(323) | Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290727 | No | MW285208 | MW285450 | MW290559 | MW290629 |
| C. pseudoreteaudii | AA-- | CSF16019 6 | 20181021-1-(325) | Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | No | MAT1-2-1_GT1 | HE | MAT1-2 | No | MW319740 | MW285209 | MW285451 | – | – |
| C. pseudoreteaudii | AA-- | CSF16021 6 | 20181021-1-(329) | Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290728 | No | MW285210 | MW285452 | – | – |
| C. pseudoreteaudii | AAAA | CSF16023 6 | 20181021-1-(333) | Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290729 | No | MW285211 | MW285453 | MW290560 | MW290630 |
| C. pseudoreteaudii | AA-- | CSF16024 6 | 20181021-1-(335) | Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | No | MAT1-2-1_GT2 | HE | MAT1-2 | No | MW319741 | MW285212 | MW285454 | – | – |
| C. pseudoreteaudii | AAAA | CSF16027 6,9,10,11 | 20181021-1-(341) | Eucalyptus genotype CEPT1869 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290730 | No | MW285213 | MW285455 | MW290561 | MW290631 |
| C. pseudoreteaudii | AAAA | CSF16031 6 | 20181021-1-(349) | Eucalyptus genotype CEPT1869 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290731 | No | MW285214 | MW285456 | MW290562 | MW290632 |
| C. pseudoreteaudii | AA-- | CSF16035 6 | 20181021-1-(357) | Eucalyptus genotype CEPT1869 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290732 | No | MW285215 | MW285457 | – | – |
| C. pseudoreteaudii | AA-- | CSF16039 6 | 20181021-1-(365) | Eucalyptus genotype CEPT1869 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290733 | No | MW285216 | MW285458 | – | – |
| C. pseudoreteaudii | AA-- | CSF16042 6 | 20181021-1-(371) | Eucalyptus genotype CEPT1869 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290734 | No | MW285217 | MW285459 | – | – |
| C. pseudoreteaudii | AA-- | CSF16045 6 | 20181021-1-(377) | Eucalyptus genotype CEPT1869 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290735 | No | MW285218 | MW285460 | – | – |
| C. pseudoreteaudii | AAAA | CSF16053 6 | 20181021-1-(12) | soil under Eucalyptus genotype CEPT 1860 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290737 | No | MW285225 | MW285467 | MW290564 | MW290634 |
| C. pseudoreteaudii | AA-- | CSF16054 6 | 20181021-1-(12) | soil under Eucalyptus genotype CEPT 1860 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290738 | No | MW285226 | MW285468 | – | – |
| C. pseudoreteaudii | AA-- | CSF16055 6 | 20181021-1-(12) | soil under Eucalyptus genotype CEPT 1860 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290739 | No | MW285227 | MW285469 | – | – |
| C. pseudoreteaudii | AAAA | CSF16056 6,9,10,11 | 20181021-1-(12) | soil under Eucalyptus genotype CEPT 1860 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290740 | No | MW285228 | MW285470 | MW290565 | MW290635 |
| C. pseudoreteaudii | AA-- | CSF16057 6 | 20181021-1-(12) | soil under Eucalyptus genotype CEPT 1860 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290741 | No | MW285229 | MW285471 | – | – |
| C. pseudoreteaudii | AA-- | CSF16062 6 | 20181021-1-(38) | soil under Eucalyptus genotype CEPT 1860 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290742 | No | MW285232 | MW285474 | – | – |
| C. pseudoreteaudii | AA-- | CSF16063 6 | 20181021-1-(38) | soil under Eucalyptus genotype CEPT 1860 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290743 | No | MW285233 | MW285475 | – | – |
| C. pseudoreteaudii | AA-- | CSF16064 6 | 20181021-1-(38) | soil under Eucalyptus genotype CEPT 1860 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290744 | No | MW285234 | MW285476 | – | – |
| C. pseudoreteaudii | AAAA | CSF16066 6,9 | 20181021-1-(44) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290745 | No | MW285236 | MW285478 | MW290566 | MW290636 |
| C. pseudoreteaudii | AA-- | CSF16067 6 | 20181021-1-(44) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290746 | No | MW285237 | MW285479 | – | – |
| C. pseudoreteaudii | AA-- | CSF16068 6 | 20181021-1-(44) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290747 | No | MW285238 | MW285480 | – | – |
| C. pseudoreteaudii | AA-- | CSF16072 6 | 20181021-1-(52) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290748 | No | MW285242 | MW285484 | – | – |
| C. pseudoreteaudii | AA-- | CSF16076 6 | 20181021-1-(54) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290749 | No | MW285246 | MW285488 | – | – |
| C. pseudoreteaudii | AA-- | CSF16077 6 | 20181021-1-(54) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290750 | No | MW285247 | MW285489 | – | – |
| C. pseudoreteaudii | AA-- | CSF16078 6 | 20181021-1-(54) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290751 | No | MW285248 | MW285490 | – | – |
| C. pseudoreteaudii | AA-- | CSF16079 6 | 20181021-1-(54) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290752 | No | MW285249 | MW285491 | – | – |
| C. pseudoreteaudii | AA-- | CSF16080 6 | 20181021-1-(56) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290753 | No | MW285250 | MW285492 | – | – |
| C. pseudoreteaudii | AA-- | CSF16082 6 | 20181021-1-(56) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290754 | No | MW285251 | MW285493 | – | – |
| C. pseudoreteaudii | AA-- | CSF16083 6 | 20181021-1-(56) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290755 | No | MW285252 | MW285494 | – | – |
| C. pseudoreteaudii | AA-- | CSF16084 6 | 20181021-1-(56) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290756 | No | MW285253 | MW285495 | – | – |
| C. pseudoreteaudii | AA-- | CSF16085 6 | 20181021-1-(64) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290757 | No | MW285254 | MW285496 | – | – |
| C. pseudoreteaudii | AA-- | CSF16086 6 | 20181021-1-(64) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290758 | No | MW285255 | MW285497 | – | – |
| C. pseudoreteaudii | AA-- | CSF16087 6 | 20181021-1-(64) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290759 | No | MW285256 | MW285498 | – | – |
| C. pseudoreteaudii | AA-- | CSF16089 6 | 20181021-1-(64) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290760 | No | MW285257 | MW285499 | – | – |
| C. pseudoreteaudii | AAAA | CSF16090 6,9 | 20181021-1-(86) | soil under Eucalyptus genotype CEPT1862 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290761 | No | MW285258 | MW285500 | MW290568 | MW290638 |
| C. pseudoreteaudii | AA-- | CSF16094 6 | 20181021-1-(106) | soil under Eucalyptus genotype CEPT1862 | S.F. Chen, G.Q. Li & Q.C. Wang | No | MAT1-2-1_GT1 | HE | MAT1-2 | No | MW319745 | MW285262 | MW285504 | – | – |
| C. pseudoreteaudii | AA-- | CSF16095 6 | 20181021-1-(106) | soil under Eucalyptus genotype CEPT1862 | S.F. Chen, G.Q. Li & Q.C. Wang | No | MAT1-2-1_GT1 | HE | MAT1-2 | No | MW319746 | MW285263 | MW285505 | – | – |
| C. pseudoreteaudii | AAAA | CSF16101 6,9 | 20181021-1-(154) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | No | MAT1-2-1_GT1 | HE | MAT1-2 | No | MW319748 | MW285269 | MW285511 | MW290577 | MW290647 |
| C. pseudoreteaudii | AA-- | CSF16102 6,10 | 20181021-1-(154) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | No | MAT1-2-1_GT1 | HE | MAT1-2 | No | MW319749 | MW285270 | MW285512 | – | – |
| C. pseudoreteaudii | AA-- | CSF16103 6 | 20181021-1-(154) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | No | MAT1-2-1_GT1 | HE | MAT1-2 | No | MW319750 | MW285271 | MW285513 | – | – |
| C. pseudoreteaudii | AA-- | CSF16114 6 | 20181021-1-(172) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290766 | No | MW285281 | MW285523 | – | – |
| C. pseudoreteaudii | AAAA | CSF16116 6,9,10,11 | 20181021-1-(196) | soil under Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290767 | No | MW285283 | MW285525 | MW290580 | MW290650 |
| C. pseudoreteaudii | AA-- | CSF16117 6 | 20181021-1-(196) | soil under Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290768 | No | MW285284 | MW285526 | – | – |
| C. pseudoreteaudii | AA-- | CSF16118 6 | 20181021-1-(196) | soil under Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290769 | No | MW285285 | MW285527 | – | – |
| C. pseudoreteaudii | AA-- | CSF16119 6 | 20181021-1-(196) | soil under Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290770 | No | MW285286 | MW285528 | – | – |
| C. pseudoreteaudii | AAAA | CSF16134 6,9 | 20181021-1-(222) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290777 | No | MW285301 | MW285539 | MW290590 | MW290660 |
| C. pseudoreteaudii | AA-- | CSF16135 6 | 20181021-1-(222) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290778 | No | MW285302 | MW285540 | – | – |
| C. pseudoreteaudii | AA-- | CSF16136 6 | 20181021-1-(222) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290779 | No | MW285303 | MW285541 | – | – |
| C. pseudoreteaudii | AA-- | CSF16137 6 | 20181021-1-(222) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290780 | No | MW285304 | MW285542 | – | – |
| C. pseudoreteaudii | AA-- | CSF16138 6 | 20181021-1-(228) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290781 | No | MW285305 | MW285543 | – | – |
| C. pseudoreteaudii | AA-- | CSF16139 6 | 20181021-1-(228) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290782 | No | MW285306 | MW285544 | – | – |
| C. pseudoreteaudii | AA-- | CSF16140 6 | 20181021-1-(228) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290783 | No | MW285307 | MW285545 | – | – |
| C. pseudoreteaudii | AA-- | CSF16141 6 | 20181021-1-(230) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290784 | No | MW285308 | MW285546 | – | – |
| C. pseudoreteaudii | AA-- | CSF16142 6 | 20181021-1-(230) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290785 | No | MW285309 | MW285547 | – | – |
| C. pseudoreteaudii | AA-- | CSF16143 6 | 20181021-1-(230) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290786 | No | MW285310 | MW285548 | – | – |
| C. pseudoreteaudii | AA-- | CSF16144 6 | 20181021-1-(230) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290787 | No | MW285311 | MW285549 | – | – |
| C. pseudoreteaudii | AAAA | CSF16163 6,9 | 20181021-1-(278) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290790 | No | MW285328 | MW285566 | MW290597 | MW290667 |
| C. pseudoreteaudii | AA-- | CSF16164 6 | 20181021-1-(278) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290791 | No | MW285329 | MW285567 | – | – |
| C. pseudoreteaudii | AA-- | CSF16165 6 | 20181021-1-(278) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290792 | No | MW285330 | MW285568 | – | – |
| C. pseudoreteaudii | AA-A | CSF16209 6 | 20181021-1-(320) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290793 | No | MW285369 | MW285610 | No | MW290673 |
| C. pseudoreteaudii | AAAA | CSF16211 6,9 | 20181021-1-(320) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290794 | No | MW285371 | MW285612 | MW290603 | MW290674 |
| C. pseudoreteaudii | AAAA | CSF16235 6,9 | 20181021-1-(342) | soil under Eucalyptus genotype CEPT1869 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290798 | No | MW285392 | MW285632 | MW290609 | MW290681 |
| C. pseudoreteaudii | AA-- | CSF16236 6 | 20181021-1-(342) | soil under Eucalyptus genotype CEPT1869 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | HE | MAT1-1 | MW290799 | No | MW285393 | MW285633 | – | – |
| C. pseudoreteaudii | AABA | CSF16016 6,9,10,11 | 20181021-1-(319) | Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | No | MAT1-2-1_GT2 | HE | MAT1-2 | No | MW319739 | MW285206 | MW285448 | MW290558 | MW290628 |
| C. reteaudii | AAAA | CSF16105 6,9,10,11 | 20181021-1-(154) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | No | P_HE | MAT1-1 | MW290765 | No | MW285272 | MW285514 | MW290578 | MW290648 |
| C. auriculiformis | AAAA | CSF16185 6,9,10,11 | 20181021-1-(302) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | No | MAT1-2-1_GT1 | P_HE | MAT1-2 | No | MW319759 | MW285349 | MW285587 | MW290598 | MW290669 |
| C. hongkongensis | AAAA | CSF16047 6,9 | 20181021-1-(2) | soil under Eucalyptus genotype CEPT1860 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | MAT1-2-1_GT1 | HO | homothallic | MW290736 | MW319742 | MW285219 | MW285461 | MW290563 | MW290633 |
| C. hongkongensis | AA-- | CSF16048 | 20181021-1-(2) | soil under Eucalyptus genotype CEPT1860 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285220 | MW285462 | – | – |
| C. hongkongensis | AA-- | CSF16049 | 20181021-1-(10) | soil under Eucalyptus genotype CEPT1860 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285221 | MW285463 | – | – |
| C. hongkongensis | AA-- | CSF16050 | 20181021-1-(10) | soil under Eucalyptus genotype CEPT1860 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285222 | MW285464 | – | – |
| C. hongkongensis | AA-- | CSF16051 | 20181021-1-(10) | soil under Eucalyptus genotype CEPT1860 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285223 | MW285465 | – | – |
| C. hongkongensis | AA-- | CSF16052 | 20181021-1-(10) | soil under Eucalyptus genotype CEPT1860 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285224 | MW285466 | – | – |
| C. hongkongensis | AA-- | CSF16058 | 20181021-1-(34) | soil under Eucalyptus genotype CEPT1860 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285230 | MW285472 | – | – |
| C. hongkongensis | AA-- | CSF16059 | 20181021-1-(34) | soil under Eucalyptus genotype CEPT1860 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285231 | MW285473 | – | – |
| C. hongkongensis | AA-- | CSF16065 | 20181021-1-(38) | soil under Eucalyptus genotype CEPT1860 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285235 | MW285477 | – | – |
| C. hongkongensis | AAAA | CSF16069 9 | 20181021-1-(44) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285239 | MW285481 | MW290567 | MW290637 |
| C. hongkongensis | AA-- | CSF16070 | 20181021-1-(44) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285240 | MW285482 | – | – |
| C. hongkongensis | AA-- | CSF16071 | 20181021-1-(52) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285241 | MW285483 | – | – |
| C. hongkongensis | AA-- | CSF16073 | 20181021-1-(52) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285243 | MW285485 | – | – |
| C. hongkongensis | AA-- | CSF16074 | 20181021-1-(52) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285244 | MW285486 | – | – |
| C. hongkongensis | AA-- | CSF16075 | 20181021-1-(52) | soil under Eucalyptus genotype CEPT1861 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285245 | MW285487 | – | – |
| C. hongkongensis | AAAA | CSF161069 | 20181021-1-(160) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285273 | MW285515 | MW290579 | MW290649 |
| C. hongkongensis | AA-- | CSF16107 | 20181021-1-(160) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285274 | MW285516 | – | – |
| C. hongkongensis | AA-- | CSF16108 | 20181021-1-(160) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285275 | MW285517 | – | – |
| C. hongkongensis | AA-- | CSF16109 | 20181021-1-(160) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285276 | MW285518 | – | – |
| C. hongkongensis | AA-- | CSF16110 | 20181021-1-(172) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285277 | MW285519 | – | – |
| C. hongkongensis | AA-- | CSF16111 | 20181021-1-(172) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285278 | MW285520 | – | – |
| C. hongkongensis | AA-- | CSF16112 | 20181021-1-(172) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285279 | MW285521 | – | – |
| C. hongkongensis | AA-- | CSF16113 | 20181021-1-(172) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285280 | MW285522 | – | – |
| C. hongkongensis | AA-- | CSF16115 | 20181021-1-(172) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285282 | MW285524 | – | – |
| C. hongkongensis | AAAA | CSF16120 9 | 20181021-1-(198) | soil under Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285287 | MW285529 | MW290581 | MW290651 |
| C. hongkongensis | AA-- | CSF16125 | 20181021-1-(214) | soil under Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285292 | MW285534 | – | – |
| C. hongkongensis | AA-- | CSF16126 | 20181021-1-(214) | soil under Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285293 | MW285535 | – | – |
| C. hongkongensis | AA-- | CSF16127 | 20181021-1-(214) | soil under Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285294 | MW285536 | – | – |
| C. hongkongensis | AA-- | CSF16128 | 20181021-1-(214) | soil under Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285295 | MW285537 | – | – |
| C. hongkongensis | AA-- | CSF16129 | 20181021-1-(214) | soil under Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285296 | MW285538 | – | – |
| C. hongkongensis | AAAA | CSF16149 9 | 20181021-1-(258) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285316 | MW285554 | MW290595 | MW290665 |
| C. hongkongensis | AA-- | CSF16150 | 20181021-1-(258) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285317 | MW285555 | – | – |
| C. hongkongensis | AA-- | CSF16151 | 20181021-1-(258) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285318 | MW285556 | – | – |
| C. hongkongensis | AA-- | CSF16152 | 20181021-1-(258) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285319 | MW285557 | – | – |
| C. hongkongensis | AA-- | CSF16153 | 20181021-1-(260) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285320 | MW285558 | – | – |
| C. hongkongensis | AA-- | CSF16154 | 20181021-1-(260) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285321 | MW285559 | – | – |
| C. hongkongensis | AA-- | CSF16155 | 20181021-1-(260) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285322 | MW285560 | – | – |
| C. hongkongensis | AA-- | CSF16156 | 20181021-1-(260) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285323 | MW285561 | – | – |
| C. hongkongensis | AAAA | CSF16159 9 | 20181021-1-(274) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285324 | MW285562 | MW290596 | MW290666 |
| C. hongkongensis | AA-- | CSF16160 | 20181021-1-(274) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285325 | MW285563 | – | – |
| C. hongkongensis | AA-- | CSF16161 | 20181021-1-(274) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285326 | MW285564 | – | – |
| C. hongkongensis | AA-- | CSF16162 | 20181021-1-(274) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285327 | MW285565 | – | – |
| C. hongkongensis | AA-- | CSF16166 | 20181021-1-(278) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285331 | MW285569 | – | – |
| C. hongkongensis | AA-- | CSF16167 | 20181021-1-(280) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285332 | MW285570 | – | – |
| C. hongkongensis | AA-- | CSF16169 | 20181021-1-(280) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285333 | MW285571 | – | – |
| C. hongkongensis | AA-- | CSF16170 | 20181021-1-(280) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285334 | MW285572 | – | – |
| C. hongkongensis | AA-- | CSF16171 | 20181021-1-(284) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285335 | MW285573 | – | – |
| C. hongkongensis | AA-- | CSF16172 | 20181021-1-(284) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285336 | MW285574 | – | – |
| C. hongkongensis | AA-- | CSF16173 | 20181021-1-(284) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285337 | MW285575 | – | – |
| C. hongkongensis | AA-- | CSF16174 | 20181021-1-(284) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285338 | MW285576 | – | – |
| C. hongkongensis | AA-- | CSF16175 | 20181021-1-(286) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285339 | MW285577 | – | – |
| C. hongkongensis | AA-- | CSF16176 | 20181021-1-(286) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285340 | MW285578 | – | – |
| C. hongkongensis | AA-- | CSF16177 | 20181021-1-(286) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285341 | MW285579 | – | – |
| C. hongkongensis | AA-- | CSF16178 | 20181021-1-(286) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285342 | MW285580 | – | – |
| C. hongkongensis | AA-- | CSF16179 | 20181021-1-(290) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285343 | MW285581 | – | – |
| C. hongkongensis | AA-- | CSF16180 | 20181021-1-(290) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285344 | MW285582 | – | – |
| C. hongkongensis | AA-- | CSF16181 | 20181021-1-(290) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285345 | MW285583 | – | – |
| C. hongkongensis | AA-- | CSF16182 | 20181021-1-(290) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285346 | MW285584 | – | – |
| C. hongkongensis | AA-- | CSF16183 | 20181021-1-(290) | soil under Eucalyptus genotype CEPT1867 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285347 | MW285585 | – | – |
| C. hongkongensis | AA-A | CSF16184 | 20181021-1-(302) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285348 | MW285586 | No | MW290668 |
| C. hongkongensis | AA-- | CSF16186 | 20181021-1-(302) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285350 | MW285588 | – | – |
| C. hongkongensis | AAAA | CSF16187 9 | 20181021-1-(302) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285351 | MW285589 | MW290599 | MW290670 |
| C. hongkongensis | AAAA | CSF16188 | 20181021-1-(304) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285352 | MW285590 | MW290600 | MW290671 |
| C. hongkongensis | AA-- | CSF16189 | 20181021-1-(304) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285353 | MW285591 | – | – |
| C. hongkongensis | AA-- | CSF16190 | 20181021-1-(304) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285354 | MW285592 | – | – |
| C. hongkongensis | -A-- | CSF16191 | 20181021-1-(304) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | No | MW285593 | – | – |
| C. hongkongensis | AA-- | CSF16192 | 20181021-1-(306) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285355 | MW285594 | – | – |
| C. hongkongensis | AA-- | CSF16193 | 20181021-1-(306) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285356 | MW285595 | – | – |
| C. hongkongensis | AA-- | CSF16194 | 20181021-1-(306) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285357 | MW285596 | – | – |
| C. hongkongensis | AA-- | CSF16195 | 20181021-1-(306) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285358 | MW285597 | – | – |
| C. hongkongensis | -A-- | CSF16196 | 20181021-1-(306) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | No | MW285598 | – | – |
| C. hongkongensis | AA-- | CSF16197 | 20181021-1-(306) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285359 | MW285599 | – | – |
| C. hongkongensis | -A-- | CSF16199 | 20181021-1-(308) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | No | MW285601 | – | – |
| C. hongkongensis | AA-- | CSF16202 | 20181021-1-(314) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285363 | MW285604 | – | – |
| C. hongkongensis | AA-- | CSF16203 | 20181021-1-(314) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285364 | MW285605 | – | – |
| C. hongkongensis | AA-- | CSF16204 | 20181021-1-(314) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285365 | MW285606 | – | – |
| C. hongkongensis | AA-- | CSF16205 | 20181021-1-(314) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285366 | MW285607 | – | – |
| C. hongkongensis | AA-- | CSF16207 | 20181021-1-(320) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285367 | MW285608 | – | – |
| C. hongkongensis | AA-- | CSF16208 | 20181021-1-(320) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285368 | MW285609 | – | – |
| C. hongkongensis | AA-- | CSF16210 | 20181021-1-(320) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285370 | MW285611 | – | – |
| C. hongkongensis | AA-- | CSF16212 | 20181021-1-(322) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285372 | MW285613 | – | – |
| C. hongkongensis | AA-- | CSF16213 | 20181021-1-(322) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285373 | MW285614 | – | – |
| C. hongkongensis | AA-- | CSF16214 | 20181021-1-(322) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285374 | MW285615 | – | – |
| C. hongkongensis | AA-- | CSF16215 | 20181021-1-(322) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285375 | MW285616 | – | – |
| C. hongkongensis | AA-- | CSF16218 | 20181021-1-(324) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285378 | MW285619 | – | – |
| C. hongkongensis | AA-- | CSF16219 | 20181021-1-(324) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285379 | MW285620 | – | – |
| C. hongkongensis | AA-- | CSF16221 | 20181021-1-(326) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285380 | MW285621 | – | – |
| C. hongkongensis | AA-- | CSF16222 | 20181021-1-(326) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285381 | MW285622 | – | – |
| C. hongkongensis | AA-- | CSF16223 | 20181021-1-(330) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285382 | MW285623 | – | – |
| C. hongkongensis | AA-- | CSF16224 | 20181021-1-(330) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285383 | MW285624 | – | – |
| C. hongkongensis | AA-- | CSF16225 | 20181021-1-(330) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285384 | MW285625 | – | – |
| C. hongkongensis | AA-- | CSF16226 | 20181021-1-(330) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285385 | MW285626 | – | – |
| C. hongkongensis | A--- | CSF16231 | 20181021-1-(336) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285390 | No | – | – |
| C. hongkongensis | AA-- | CSF16234 | 20181021-1-(336) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285391 | MW285631 | – | – |
| C. hongkongensis | AAAA | CSF16237 6,9,10,11 | 20181021-1-(350) | soil under Eucalyptus genotype CEPT1869 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | MAT1-2-1_GT2 | HO | homothallic | MW290800 | MW319763 | MW285394 | MW285634 | MW290610 | MW290682 |
| C. hongkongensis | AA-- | CSF16238 | 20181021-1-(350) | soil under Eucalyptus genotype CEPT1869 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285395 | MW285635 | – | – |
| C. hongkongensis | AA-- | CSF16239 | 20181021-1-(350) | soil under Eucalyptus genotype CEPT1869 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285396 | MW285636 | – | – |
| C. hongkongensis | AA-- | CSF16240 | 20181021-1-(350) | soil under Eucalyptus genotype CEPT1869 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285397 | MW285637 | – | – |
| C. hongkongensis | ABAA | CSF16096 6,9 | 20181021-1-(142) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | MAT1-2-1_GT2 | HO | homothallic | MW290764 | MW319747 | MW285264 | MW285506 | MW290572 | MW290642 |
| C. hongkongensis | ABAA | CSF16097 | 20181021-1-(142) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285265 | MW285507 | MW290573 | MW290643 |
| C. hongkongensis | ABAA | CSF16098 | 20181021-1-(142) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285266 | MW285508 | MW290574 | MW290644 |
| C. hongkongensis | ABAA | CSF16099 | 20181021-1-(142) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285267 | MW285509 | MW290575 | MW290645 |
| C. hongkongensis | ABAA | CSF16100 | 20181021-1-(142) | soil under Eucalyptus genotype CEPT1864 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285268 | MW285510 | MW290576 | MW290646 |
| C. hongkongensis | ABAA | CSF16216 6,9 | 20181021-1-(324) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT2 | MAT1-2-1_GT2 | HO | homothallic | MW290795 | MW319760 | MW285376 | MW285617 | MW290604 | MW290675 |
| C. hongkongensis | AB-A | CSF16217 | 20181021-1-(324) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285377 | MW285618 | No | MW290676 |
| C. hongkongensis | ACAA | CSF16121 6,9,10 | 20181021-1-(206) | soil under Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT2 | MAT1-2-1_GT2 | HO | homothallic | MW290771 | MW319751 | MW285288 | MW285530 | MW290582 | MW290652 |
| C. hongkongensis | ACAA | CSF16122 | 20181021-1-(206) | soil under Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285289 | MW285531 | MW290583 | MW290653 |
| C. hongkongensis | ACAA | CSF16123 | 20181021-1-(206) | soil under Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285290 | MW285532 | MW290584 | MW290654 |
| C. hongkongensis | ACAA | CSF16124 6 | 20181021-1-(206) | soil under Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT2 | MAT1-2-1_GT2 | HO | homothallic | MW290772 | MW319752 | MW285291 | MW285533 | MW290585 | MW290655 |
| C. hongkongensis | ADA- | CSF16198 | 20181021-1-(308) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285360 | MW285600 | MW290601 | No |
| C. hongkongensis | AD-- | CSF16200 | 20181021-1-(308) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285361 | MW285602 | No | No |
| C. hongkongensis | ADAA | CSF16201 | 20181021-1-(308) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285362 | MW285603 | MW290602 | MW290672 |
| C. hongkongensis | ADAA | CSF16227 6 | 20181021-1-(334) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | MAT1-2-1_GT2 | HO | homothallic | MW290796 | MW319761 | MW285386 | MW285627 | MW290605 | MW290677 |
| C. hongkongensis | ADAA | CSF16228 | 20181021-1-(334) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285387 | MW285628 | MW290606 | MW290678 |
| C. hongkongensis | ADAA | CSF16229 | 20181021-1-(334) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285388 | MW285629 | MW290607 | MW290679 |
| C. hongkongensis | ADAA | CSF16230 6,9,10,11 | 20181021-1-(334) | soil under Eucalyptus genotype CEPT1868 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | MAT1-2-1_GT2 | HO | homothallic | MW290797 | MW319762 | MW285389 | MW285630 | MW290608 | MW290680 |
| C. hongkongensis | BAAA | CSF16091 6 | 20181021-1-(86) | soil under Eucalyptus genotype CEPT1862 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | MAT1-2-1_GT2 | HO | homothallic | MW290762 | MW319743 | MW285259 | MW285501 | MW290569 | MW290639 |
| C. hongkongensis | BAAA | CSF16092 | 20181021-1-(86) | soil under Eucalyptus genotype CEPT1862 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285260 | MW285502 | MW290570 | MW290640 |
| C. hongkongensis | BAAA | CSF16093 6,9 | 20181021-1-(86) | soil under Eucalyptus genotype CEPT1862 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | MAT1-2-1_GT2 | HO | homothallic | MW290763 | MW319744 | MW285261 | MW285503 | MW290571 | MW290641 |
| C. hongkongensis | CAAA | CSF16145 6,9,10,11 | 20181021-1-(256) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | MAT1-2-1_GT2 | HO | homothallic | MW290788 | MW319757 | MW285312 | MW285550 | MW290591 | MW290661 |
| C. hongkongensis | CAAA | CSF16146 | 20181021-1-(256) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285313 | MW285551 | MW290592 | MW290662 |
| C. hongkongensis | CAAA | CSF16147 | 20181021-1-(256) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | – | – | – | – | – | – | MW285314 | MW285552 | MW290593 | MW290663 |
| C. hongkongensis | CAAA | CSF16148 6 | 20181021-1-(256) | soil under Eucalyptus genotype CEPT1866 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | MAT1-2-1_GT2 | HO | homothallic | MW290789 | MW319758 | MW285315 | MW285553 | MW290594 | MW290664 |
| C. aconidialis | A-AA | CSF16130 6,9,10,11 | 20181021-1-(216) | soil under Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | MAT1-2-1-GT1 | HO | homothallic | MW290773 | MW319753 | MW285297 | No | MW290586 | MW290656 |
| C. aconidialis | A-AA | CSF16131 6,9,10 | 20181021-1-(216) | soil under Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | MAT1-2-1-GT1 | HO | homothallic | MW290774 | MW319754 | MW285298 | No | MW290587 | MW290657 |
| C. aconidialis | A-AA | CSF16132 6,9 | 20181021-1-(216) | soil under Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | MAT1-2-1-GT1 | HO | homothallic | MW290775 | MW319755 | MW285299 | No | MW290588 | MW290658 |
| C. aconidialis | A-AA | CSF16133 6,9,10,11 | 20181021-1-(216) | soil under Eucalyptus genotype CEPT1865 | S.F. Chen, G.Q. Li & Q.C. Wang | MAT1-1-1_GT1 | MAT1-2-1-GT1 | HO | homothallic | MW290776 | MW319756 | MW285300 | No | MW290589 | MW290659 |
1 Genotype within each Calonectria species, determined by sequences of the tef1, tub2, cmdA and his3 regions; “-” means not available. 2 CSF: Culture Collection located at China Eucalypt Research Centre (CERC), Chinese Academy of Forestry, ZhanJiang, GuangDong Province, China. 3 Genotype of each mating gene within each Calonectria species, determined by sequences of Mat1-1-1 or Mat1-2-1; GT1, GT2 mean genotype 1 and genotype 2, respectively. 4 HE = Heterothallic; HO = Homothallic; P_HE = Putative heterothallic. 5 tef1 = translation elongation factor 1-alpha; tub2 = β-tubulin; cmdA = calmodulin; his3 = histone H3. 6 Isolates used for mating studies. 7 “No” represents the relative locus was not successfully amplified in the current study. 8 “–” represents the relative locus was not amplified in the current study. 9 Isolates used for phylogenetic analyses. 10 Isolates used for morphological studies. 11 Isolates used for pathogenicity tests.
Table A2.
Isolates from other studies and used in the phylogenetic analyses for this study.
Table A2.
Isolates from other studies and used in the phylogenetic analyses for this study.
| Species Code 1 | Species | Isolate No. 2,3 | Other Collection Number 3 | Hosts | Area of Occurrence | Collector | GenBank Accession Numbers 4 | References or Source of Data | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| tef1 | tub2 | cmdA | his3 | ||||||||
| B1 | Calonectria acaciicola | CMW 47173T | CBS 143557 | Soil (Acacia auriculiformis plantation) | Do Luong, Nghe An, Vietnam | N.Q. Pham and T.Q. Pham | MT412690 | MT412930 | MT335160 | MT335399 | [28,36] |
| CMW 47174 | CBS 143558 | Soil (A. auriculiformis plantation) | Do Luong, Nghe An, Vietnam | N.Q. Pham and T.Q. Pham | MT412691 | MT412931 | MT335161 | MT335400 | [28,36] | ||
| B2 | C. acicola | CMW 30996T | – | Phoenix canariensis | Northland, New Zealand | H. Pearson | MT412692 | MT412932 | MT335162 | MT335401 | [28,49,50] |
| CBS 114812 | CMW 51216 | P. canariensis | Northland, New Zealand | H. Pearson | MT412693 | MT412933 | MT335163 | MT335402 | [28,49,50] | ||
| B4 | C. aconidialis | CMW 35174T | CBS 136086; CERC 1850 | Soil (Eucalyptus plantation) | HaiNan, China | X. Mou and S.F. Chen | MT412695 | N/A 5 | MT335165 | MT335404 | [28,29] |
| CMW 35384 | CBS 136091; CERC 1886 | Soil (Eucalyptus plantation) | HaiNan, China | X. Mou and S.F. Chen | MT412696 | N/A | MT335166 | MT335405 | [28,29] | ||
| B5 | C. aeknauliensis | CMW 48253T | CBS 143559 | Soil (Eucalyptus plantation) | Aek Nauli, North Sumatra, Indonesia | M.J. Wingfield | MT412710 | N/A | MT335180 | MT335419 | [28,36] |
| CMW 48254 | CBS 143560 | Soil (Eucalyptus plantation) | Aek Nauli, North Sumatra, Indonesia | M.J. Wingfield | MT412711 | N/A | MT335181 | MT335420 | [28,36] | ||
| B8 | C. asiatica | CBS 114073T | CMW 23782; CPC 3900 | Debris (leaf litter) | Prathet Thai, Thailand | N.L. Hywel-Jones | AY725705 | AY725616 | AY725741 | AY725658 | [43,50] |
| B9 | C. auriculiformis | CMW 47178T | CBS 143561 | Soil (A. auriculiformis plantation) | Hau Loc, Thanh Hoa, Vietnam | N.Q. Pham and T.Q. Pham | MT412721 | MT412944 | MT335190 | MT335430 | [28,36] |
| CMW 47179 | CBS 143562 | Soil (A. auriculiformis plantation) | Hau Loc, Thanh Hoa, Vietnam | N.Q. Pham and T.Q. Pham | MT412722 | MT412945 | MT335191 | MT335431 | [28,36] | ||
| B10 | C. australiensis | CMW 23669T | CBS 112954; CPC 4714 | Ficus pleurocarpa | Queensland, Australia | C. Pearce and B. Paulus | MT412723 | MT412946 | MT335192 | MT335432 | [28,50,51] |
| B14 | C. brasiliensis | CBS 230.51T | IMI 299576 | Eucalyptus sp. | Ceara state, Brazil | T.R. Ciferri | MT412731 | MT412953 | MT335200 | MT335440 | [23,28,52] |
| CMW 32949 | CBS 114257; CPC 1944 | Eucalyptus sp. | Aracruz, Brazil | A.C. Alfenas | MT412732 | MT412954 | MT335201 | MT335441 | [28,50] | ||
| B17 | C. brassicicola | CBS 112841T | CMW 51206; CPC 4552 | Soil (Brassica sp.) | Indonesia | M.J. Wingfield | KX784689 | KX784619 | KX784561 | N/A | [32] |
| B19 | C. bumicola | CMW 48257T | CBS 143575 | Soil (Eucalyptus plantation) | Aek Nauli, North Sumatra, Indonesia | M.J. Wingfield | MT412736 | N/A | MT335205 | MT335445 | [28,36] |
| B20 | C. canadiana | CMW 23673T | CBS 110817; STE-U 499 | Picea sp. | Canada | S. Greifenhagen | MT412737 | MT412958 | MT335206 | MT335446 | [23,28,53,54] |
| CERC 8952 | – | Soil | HeNan, China | S.F. Chen | MT412821 | MT413035 | MT335290 | MT335530 | [28,33] | ||
| B22 | C. cerciana | CMW 25309T | CBS 123693 | E. urophylla × E. grandis hybrid cutting | CERC nursery, GuangDong, China | M.J. Wingfield and X.D. Zhou | MT412742 | MT412963 | MT335211 | MT335451 | [28,30] |
| CMW 25290 | CBS 123695 | E. urophylla × E. grandis hybrid cutting | CERC nursery, GuangDong, China | M.J. Wingfield and X.D. Zhou | MT412743 | MT412964 | MT335212 | MT335452 | [28,30] | ||
| B23 | C. chinensis | CMW 23674T | CBS 114827; CPC 4101 | Soil | Hong Kong, China | E.C.Y. Liew | MT412751 | MT412972 | MT335220 | MT335460 | [28,43,50] |
| CMW 30986 | CBS 112744; CPC 4104 | Soil | Hong Kong, China | E.C.Y. Liew | MT412752 | MT412973 | MT335221 | MT335461 | [28,43,50] | ||
| B26 | C. cochinchinensis | CMW 49915T | CBS 143567 | Soil (Hevea brasiliensis plantation) | Duong Minh Chau, Tay Ninh, Vietnam | N.Q. Pham, Q.N. Dang and T.Q. Pham | MT412756 | MT412977 | MT335225 | MT335465 | [28,36] |
| CMW 47186 | CBS 143568 | Soil (A. auriculiformis plantation) | Song May, Dong Nai, Vietnam | N.Q. Pham and T.Q. Pham | MT412757 | MT412978 | MT335226 | MT335466 | [28,36] | ||
| B29 | C. colombiensis | CMW 23676T | CBS 112220; CPC 723 | Soil (E. grandis trees) | La Selva, Colombia | M.J. Wingfield | MT412759 | MT412980 | MT335228 | MT335468 | [28,43] |
| CMW 30985 | CBS 112221; CPC 724 | Soil (E. grandis trees) | La Selva, Colombia | M.J. Wingfield | MT412760 | MT412981 | MT335229 | MT335469 | [28,43] | ||
| B30 | C. crousiana | CMW 27249T | CBS 127198 | E. grandis | FuJian, China | M.J. Wingfield | MT412761 | MT412982 | MT335230 | MT335470 | [18,28] |
| CMW 27253 | CBS 127199 | E. grandis | FuJian, China | M.J. Wingfield | MT412762 | MT412983 | MT335231 | MT335471 | [18,28] | ||
| B31 | C. curvispora | CMW 23693T | CBS 116159; CPC 765 | Soil | Tamatave, Madagascar | P.W. Crous | MT412763 | N/A | MT335232 | MT335472 | [23,28,29,50,55] |
| CMW 48245 | CBS 143565 | Soil (Eucalyptus plantation) | Aek Nauli, North Sumatra, Indonesia | M.J. Wingfield | MT412764 | N/A | MT335233 | MT335473 | [28,36] | ||
| B32 | C. cylindrospora | CBS 119670 | CMW 51310; CPC 12766 | Pistacia lentiscus | Italy | N/A | MT412767 | MT412985 | MT335236 | MT335476 | [28,29,32,56] |
| CMW 30978 | CBS 110666; P90.1479; STE-U 497 | Ilex vomitoria | Florida, USA | N.E. El-Gholl | MT412768 | MT412986 | MT335237 | MT335477 | [23,28,50,56] | ||
| B44 | C. hawksworthii | CBS 111870T | CMW 51194; CPC 2405 | Nelumbo nucifera | Pamplemousses garden, Mauritius | A. Peerally | MT412785 | MT413003 | MT335254 | MT335494 | [23,28] |
| CMW 31393 | CBS 136641 | E. urophylla × E. grandis | GuangXi, China | X. Zhou and G. Zhao | MT412778 | MT412996 | MT335247 | MT335487 | [28,29] | ||
| B46 | C. heveicola | CMW 49913T | CBS 143570 | Soil (Hevea brasiliensis plantation) | Bau Bang, Binh Duong, Vietnam | N.Q. Pham, Q.N. Dang and T.Q. Pham | MT412786 | MT413004 | MT335255 | MT335495 | [28,36] |
| CMW 49928 | CBS 143571 | Soil | Bu Gia Map National Park, Binh Phuoc, Vietnam | N.Q. Pham, Q.N. Dang and T.Q. Pham | MT412811 | MT413025 | MT335280 | MT335520 | [28,36] | ||
| B48 | C. hongkongensis | CBS 114828T | CMW 51217; CPC 4670 | Soil | Hong Kong, China | M.J. Wingfield | MT412789 | MT413007 | MT335258 | MT335498 | [28,43] |
| CERC 3570 | CMW 47271 | Soil (Eucalyptus plantation) | BeiHai, GuangXi, China | S.F. Chen, J.Q. Li and G.Q. Li | MT412791 | MT413009 | MT335260 | MT335500 | [19,28] | ||
| B51 | C. ilicicola | CMW 30998T | CBS 190.50; IMI 299389; STE-U 2482 | Solanum tuberosum | Bogor, Java, Indonesia | K.B. Boedijn and J. Reitsma | MT412797 | N/A | MT335266 | MT335506 | [23,28,50] |
| B52 | C. indonesiae | CMW 23683T | CBS 112823; CPC 4508 | Syzygium aromaticum | Warambunga, Indonesia | M.J. Wingfield | MT412798 | MT413015 | MT335267 | MT335507 | [28,43] |
| CBS 112840 | CMW 51205; CPC 4554 | S. aromaticum | Warambunga, Indonesia | M.J. Wingfield | MT412799 | MT413016 | MT335268 | MT335508 | [28,43] | ||
| B54 | C. insularis | CMW 30991T | CBS 114558; CPC 768 | Soil | Tamatave, Madagascar | P.W. Crous | MT412800 | MT413017 | MT335269 | MT335509 | [28,32,50,57] |
| CMW 30992 | CBS 114559; CPC 954 | Soil | Conejos, Veracruz, Mexico | M.J. Wingfield | MT412801 | MT413018 | MT335270 | MT335510 | [28,32,50] | ||
| B55 | C. kyotensis | CBS 114525T | ATCC 18834; CMW 51824; CPC 2367 | Robinia pseudoacacia | Japan | T. Terashita | MT412802 | MT413019 | MT335271 | MT335511 | [23,28,32] |
| CBS 114550 | CMW 51825; CPC 2351 | Soil | China | M.J. Wingfield | MT412777 | MT412995 | MT335246 | MT335486 | [28,32] | ||
| B56 | C. lageniformis | CBS 111324T | CMW 51177; CPC 1473 | Leaf of Eucalyptus sp. | Rivière Noire, Mauritius | H. Smith | KX784702 | KX784632 | KX784574 | N/A | [32,58] |
| B57 | C. lantauensis | CERC 3302T | CBS 142888; CMW 47252 | Soil | LiDao, Hong Kong, China | M.J. Wingfield and S.F. Chen | MT412803 | N/A | MT335272 | MT335512 | [19,28] |
| CERC 3301 | CBS 142887; CMW 47251 | Soil | LiDao, Hong Kong, China | M.J. Wingfield and S.F. Chen | MT412804 | N/A | MT335273 | MT335513 | [19,28] | ||
| B58 | C. lateralis | CMW 31412T | CBS 136629 | Soil (Eucalyptus plantation) | GuangXi, China | X. Zhou, G. Zhao and F. Han | MT412805 | MT413020 | MT335274 | MT335514 | [28,29] |
| B63 | C. lombardiana | CMW 30602T | CBS 112634; CPC 4233; Lynfield 417 | Xanthorrhoea australis | Victoria, Australia | T. Baigent | MT412926 | MT413133 | MT335395 | MT335635 | [23,30,51] |
| B66 | C. malesiana | CMW 23687T | CBS 112752; CPC 4223 | Soil | Northern Sumatra, Indonesia | M.J. Wingfield | MT412817 | MT413031 | MT335286 | MT335526 | [28,43] |
| CBS 112710 | CMW 51199; CPC 3899 | Leaf litter | Prathet, Thailand | N.L. Hywel-Jones | MT412818 | MT413032 | MT335287 | MT335527 | [28,43] | ||
| B67 | C. maranhensis | CBS 134811T | LPF142 | Eucalyptus sp. (leaf) | Açailandia, Maranhao, Brazil | A.C. Alfenas | KM395861 | KM395948 | KM396035 | KM396118 | [26] |
| CBS 134812 | LPF143 | Eucalyptus sp. (leaf) | Açailandia, Maranhao, Brazil | A.C. Alfenas | KM395862 | KM395949 | KM396036 | KM396119 | [26] | ||
| B74 | C. multiseptata | CMW 23692T | CBS 112682; CPC 1589 | E. grandis | North Sumatra, Indonesia | M.J. Wingfield | MT412830 | MT413044 | MT335299 | MT335539 | [23,28,51,59] |
| B80 | C. pacifica | CMW 16726T | A1568; CBS 109063;IMI 354528;STE-U 2534 | Araucaria heterophylla | Hawaii, USA | M. Aragaki | MT412842 | N/A | MT335311 | MT335551 | [23,28,43,53] |
| CMW 30988 | CBS 114038 | Ipomoea aquatica | Auckland, New Zealand | C.F. Hill | MT412843 | N/A | MT335312 | MT335552 | [23,28,43,50] | ||
| B86 | C. penicilloides | CMW 23696T | CBS 174. 55; STE-U 2388 | Prunus sp. | Hatizyo Island, Japan | M. Ookubu | MT412869 | MT413081 | MT335338 | MT335578 | [23,28] |
| B89 | C. plurilateralis | CBS 111401T | CMW 51178; CPC 1637 | Soil | Ecuador | M.J. Wingfield | MT412871 | MT413083 | MT335340 | MT335580 | [28,32] |
| B90 | C. propaginicola | CBS 134815T | LPF220 | Eucalyptus sp. (seedling) | Santana, Pará, Brazil | A.C. Alfenas | KM395866 | KM395953 | KM396040 | KM396123 | [26] |
| CBS 134816 | LPF222 | Eucalyptus sp. (seedling) | Santana, Pará, Brazil | A.C. Alfenas | KM395867 | KM395954 | KM396041 | KM396124 | [26] | ||
| B97 | C. pseudoreteaudii | CMW 25310T | CBS 123694 | E. urophylla × E. grandis | GuangDong, China | M.J. Wingfield and X.D. Zhou | MT412885 | MT413096 | MT335354 | MT335594 | [28,30] |
| CMW 25292 | CBS 123696 | E. urophylla × E. grandis | GuangDong, China | M.J. Wingfield and X.D. Zhou | MT412886 | MT413097 | MT335355 | MT335595 | [28,30] | ||
| B104 | C. queenslandica | CMW 30604T | CBS 112146; CPC 3213 | E. urophylla | Lannercost, Queensland, Australia | B. Brown | MT412898 | MT413108 | MT335367 | MT335607 | [28,30,44] |
| CMW 30603 | CBS 112155; CPC 3210 | E. pellita | Lannercost, Queensland, Australia | P.Q Thu and K.M. Old | MT412899 | MT413109 | MT335368 | MT335608 | [28,30,44] | ||
| B106 | C. reteaudii | CMW 30984T | CBS 112144; CPC 3201 | E. camaldulensis | Chon Thanh, Binh Phuoc, Vietnam | M.J. Dudzinski and P.Q. Thu | MT412901 | MT413111 | MT335370 | MT335610 | [23,28,44,51] |
| CMW 16738 | CBS 112143; CPC 3200 | Eucalyptus leaves | Binh Phuoc, Vietnam | M.J. Dudzinski and P.Q. Thu | MT412902 | MT413112 | MT335371 | MT335611 | [23,28,44,51] | ||
| B112 | C. sumatrensis | CMW 23698T | CBS 112829; CPC 4518 | Soil | Northern Sumatra, Indonesia | M.J. Wingfield | MT412913 | N/A | MT335382 | MT335622 | [28,43] |
| CMW 30987 | CBS 112934; CPC 4516 | Soil | Northern Sumatra, Indonesia | M.J. Wingfield | MT412914 | N/A | MT335383 | MT335623 | [28,43] | ||
| B113 | C. syzygiicola | CBS 112831T | CMW 51204; CPC 4511 | Syzygium aromaticum | Sumatra, Indonesia | M.J. Wingfield | KX784736 | KX784663 | N/A | N/A | [32] |
| B115 | C. tonkinensis | CMW 47430T | CBS 143576 | Soil (Eucalyptus plantation) | Bavi, Hanoi, Vietnam | N.Q. Pham and T.Q. Pham | MT412915 | MT413122 | MT335384 | MT335624 | [28,36] |
| B116 | C. uniseptata | CBS 413.67T | CMW 23678; CPC 2391; IMI 299577 | Paphiopedilum callosum | Celle, Germany | W. Gerlach | GQ267307 | GQ267208 | GQ267379 | GQ267248 | [32] |
| B118 | C. variabilis | CMW 3187T | AR2675; CBS 114677; CPC 2436 | Schefflera morototoni | Pará, Brazil | F.C. de Albuquerque | MT412923 | MT413130 | MT335392 | MT335632 | [23,28,32,50,60] |
| CMW 2914 | CBS 112691; CPC 2506 | Theobroma grandiflorum | Pará, Brazil | F. Carneiro | MT412924 | MT413131 | MT335393 | MT335633 | [23,28,32,50,60] | ||
| B120 | C. yunnanensis | CERC 5339T | CBS 142897; CMW 47644 | Soil (Eucalyptus plantation) | YunNan, China | S.F. Chen and J.Q. Li | MT412927 | MT413134 | MT335396 | MT335636 | [19,28] |
| CERC 5337 | CBS 142895; CMW 47642 | Soil (Eucalyptus plantation) | YunNan, China | S.F. Chen and J.Q. Li | MT412928 | MT413135 | MT335397 | MT335637 | [19,28] | ||
| Curvicladiella cignea | CBS 109167T | CPC 1595; MUCL 40269 | Decaying leaf | French Guiana | C. Decock | KM231867 | KM232002 | KM231287 | KM231461 | [51,56,61] | |
| CBS 109168 | CPC 1594; MUCL 40268 | Decaying seed | French Guiana | C. Decock | KM231868 | KM232003 | KM231286 | KM231460 | [51,56,61] | ||
1 Codes (B1 to B120) of the 120 accepted Calonectria species resulting from Liu and co-authors [28]. 2 T: ex-type isolates of the species. 3 AR: Amy Y. Rossman working collection; ATCC: American Type Culture Collection, Virginia, USA; CBS: Westerdijk Fungal Biodiversity Institute, Utrecht, The Netherlands; CERC: China Eucalypt Research Centre, ZhanJiang, GuangDong Province, China; CMW: Culture collection of the Forestry and Agricultural Biotechnology Institute (FABI), University of Pretoria, Pretoria, South Africa; CPC: Pedro Crous working collection housed at Westerdijk Fungal Biodiversity Institute; IMI: International Mycological Institute, CABI Bioscience, Egham, Bakeham Lane, UK; MUCL: Mycotheque, Laboratoire de Mycologie Systematique st Appliqee, I’Universite, Louvian-la-Neuve, Belgium; STE-U: Department of Plant Pathology, University of Stellenbosch, South Africa; no other collection number. 4 tef1: translation elongation factor 1-alpha; tub2: β-tubulin; cmdA: calmodulin; his3: histone H3. 5 N/A represents data that is not available.
References
- Xie, Y.J.; Du, A.P. Eucalypts in Southern China; CCP Central Party School Press: Beijing, China, 2019. [Google Scholar]
- Xie, Y.J.; Arnold, R.J.; Wu, Z.H.; Chen, S.F.; Du, A.P.; Luo, J.Z. Advances in eucalypt research in China. Front. Agric. Sci. Eng. 2017, 4, 380–390. [Google Scholar] [CrossRef]
- Zhou, X.D.; Wingfield, M.J. Eucalypt diseases and their management in China. Australas. Plant Pathol. 2011, 40, 339–345. [Google Scholar] [CrossRef]
- Carstensen, G.D.; Venter, S.N.; Wingfield, M.J.; Coutinho, T.A. Two Ralstonia species associated with bacterial wilt of Eucalyptus. Plant Pathol. 2017, 66, 393–403. [Google Scholar] [CrossRef]
- Old, K.M.; Wingfield, M.J.; Yuan, Z.Q. A Manual of Diseases of Eucalypts in South-East Asia; Center for International Forestry Research: Jakarta, Indonesia, 2003. [Google Scholar]
- Chen, S.F.; Pavlic, D.; Roux, J.; Xie, Y.J.; Wingfield, M.J.; Zhou, X.D. Characterization of Botryosphaeriaceae from plantation–grown Eucalyptus species in South China. Plant Pathol. 2010, 60, 739–751. [Google Scholar] [CrossRef]
- Li, G.Q.; Liu, F.F.; Li, J.Q.; Liu, Q.L.; Chen, S.F. Botryosphaeriaceae from Eucalyptus plantations and adjacent plants in China. Persoonia 2018, 40, 63–95. [Google Scholar] [CrossRef]
- Li, G.Q.; Slippers, B.; Wingfield, M.J.; Chen, S.F. Variation in Botryosphaeriaceae from Eucalyptus plantations in YunNan Province in southwestern China across a climatic gradient. IMA Fungus 2020, 11, 22. [Google Scholar] [CrossRef]
- Chen, S.F.; Gryzenhout, M.; Roux, J.; Xie, Y.J.; Wingfield, M.J.; Zhou, X.D. Identification and pathogenicity of Chrysoporthe cubensis on Eucalyptus and Syzygium spp. in South China. Plant Dis. 2010, 94, 1143–1150. [Google Scholar] [CrossRef]
- Wang, W.; Liu, Q.L.; Li, G.Q.; Liu, F.F.; Chen, S.F. Phylogeny and Pathogenicity of Celoporthe species from Plantation Eucalyptus in Southern China. Plant Dis. 2018, 102, 1915–1927. [Google Scholar] [CrossRef]
- Wang, W.; Li, G.Q.; Liu, Q.L.; Chen, S.F. Cryphonectriaceae on Myrtales in China: Phylogeny, host range, and pathogenicity. Persoonia 2020, 45, 101–131. [Google Scholar] [CrossRef]
- Chen, S.F.; Van Wyk, M.; Roux, J.; Wingfield, M.J.; Xie, Y.J.; Zhou, X.D. Taxonomy and pathogenicity of Ceratocystis species on Eucalyptus trees in South China, including C. chinaeucensis sp. nov. Fungal Divers. 2013, 58, 267–279. [Google Scholar] [CrossRef]
- Liu, F.F.; Mbenoun, M.; Barnes, I.; Roux, J.; Wingfield, M.J.; Li, G.Q.; Li, J.Q.; Chen, S.F. New Ceratocystis species from Eucalyptus and Cunninghamia in South China. Antonie Van Leeuwenhoek 2015, 107, 1451–1473. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.F.; Barnes, I.; Chungu, D.; Wingfield, M.J.; Xie, Y.J.; Zhou, X.D. High population diversity and increasing importance of the Eucalyptus stem canker pathogen, Teratosphaeria zuluensis, in South China. Australas. Plant Pathol. 2011, 40, 407–415. [Google Scholar] [CrossRef]
- Cortinas, M.N.; Burgess, T.B.; Xu, D.; Crous, P.W.; Wingfield, B.D.; Wingfield, M.J. First record of Colletogloeopsis zuluense comb. nov. causing a stem canker of Eucalyptus in China. Mycol. Res. 2006, 110, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Burgess, T.I.; Andjic, V.; Hardy, G.E.S.J.; Dell, B.; Xu, D. First report of Phaeophleospora destructans in China. J. Trop. For. Sci. 2006, 18, 144–146. [Google Scholar]
- Burgess, T.I.; Barber, P.A.; Sufaati, S.; Xu, D.; Hardy, G.E.S.J.; Dell, B. Mycosphaerella spp. on Eucalyptus in Asia: New species, new hosts and new records. Fungal Divers. 2007, 24, 135–157. [Google Scholar]
- Chen, S.F.; Lombard, L.; Roux, J.; Xie, Y.J.; Wingfield, M.J.; Zhou, X.D. Novel species of Calonectria associated with Eucalyptus leaf blight in Southeast China. Persoonia 2011, 26, 1–12. [Google Scholar] [CrossRef]
- Li, J.Q.; Wingfield, M.J.; Liu, Q.L.; Barnes, I.; Roux, J.; Lombard, L.; Crous, P.W.; Chen, S.F. Calonectria species isolated from Eucalyptus plantations and nurseries in South China. IMA Fungus 2017, 8, 259–286. [Google Scholar] [CrossRef]
- Wang, Q.C.; Chen, S.F. Calonectria pentaseptata causes severe leaf disease on cultivated Eucalyptus in Leizhou Peninsula of southern China. Plant Dis. 2020, 104, 493–509. [Google Scholar] [CrossRef]
- Chen, S.F.; Liu, Q.L.; Li, G.Q.; Wingfield, M.J. Quambalaria species associated with eucalypt diseases in southern China. Front. Agric. Sci. Eng. 2017, 4, 433–447. [Google Scholar] [CrossRef]
- Zhou, X.D.; De Beer, Z.W.; Xie, Y.J.; Pegg, G.S.; Wingfield, M.J. DNA–based identification of Quambalaria pitereka causing severe leaf blight of Corymbia citriodora in China. Fungal Divers. 2007, 25, 245–254. [Google Scholar]
- Crous, P.W. Taxonomy and Pathology of Cylindrocladium (Calonectria) and Allied Genera; APS Press: St. Paul, MN, USA, 2002. [Google Scholar]
- Lombard, L.; Crous, P.W.; Wingfield, B.D.; Wingfield, M.J. Species concepts in Calonectria (Cylindrocladium). Stud. Mycol. 2010, 66, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Vitale, A.; Crous, P.W.; Lombard, L.; Polizzi, G. Calonectria diseases on ornamental plants in Europe and the Mediterranean basi: An overview. J. Plant Pathol. 2013, 95, 463–476. [Google Scholar]
- Alfenas, R.F.; Lombard, L.; Pereira, O.L.; Alfenas, A.C.; Crous, P.W. Diversity and potential impact of Calonectria species in Eucalyptus plantations in Brazil. Stud. Mycol. 2015, 80, 89–130. [Google Scholar] [CrossRef] [PubMed]
- Rodas, C.A.; Lombard, L.; Gryzenhout, M.; Slippers, B.; Wingfield, M.J. Cylindrocladium blight of Eucalyptus grandis in Colombia. Australas. Plant Pathol. 2005, 34, 143–149. [Google Scholar] [CrossRef]
- Liu, Q.L.; Li, J.Q.; Wingfield, M.J.; Duong, T.A.; Wingfield, B.D.; Crous, P.W.; Chen, S.F. Reconsideration of species boundaries and proposed DNA barcodes for Calonectria. Stud. Mycol. 2020. [Google Scholar] [CrossRef]
- Lombard, L.; Chen, S.F.; Mou, X.; Zhou, X.D.; Crous, P.W.; Wingfield, M.J. New species, hyper–diversity and potential importance of Calonectria spp. from Eucalyptus in South China. Stud. Mycol. 2015, 80, 151–188. [Google Scholar] [CrossRef] [PubMed]
- Lombard, L.; Zhou, X.D.; Crous, P.W.; Wingfield, B.D.; Wingfield, M.J. Calonectria species associated with cutting rot of Eucalyptus. Persoonia 2010, 24, 1–11. [Google Scholar] [CrossRef]
- Van Burik, J.A.H.; Schreckhise, R.W.; White, T.C.; Bowden, R.A.; Myerson, D. Comparison of six extraction techniques for isolation of DNA from filamentous fungi. Med Mycol. 1998, 36, 299–303. [Google Scholar] [CrossRef]
- Lombard, L.; Wingfield, M.J.; Alfenas, A.C.; Crous, P.W. The forgotten Calonectria collection: Pouring old wine into new bags. Stud. Mycol. 2016, 85, 159–198. [Google Scholar] [CrossRef]
- Liu, Q.L.; Chen, S.F. Two novel species of Calonectria isolated from soil in a natural forest in China. MycoKeys 2017, 26, 25–60. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis v. 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Q.; Wingfield, B.D.; Wingfield, M.J.; Barnes, I.; Fourie, A.; Crous, P.W.; Chen, S.F. Mating genes in Calonectria and evidence for a heterothallic ancestral state. Persoonia 2020, 45, 163–176. [Google Scholar] [CrossRef]
- Pham, N.; Barnes, I.; Chen, S.F.; Liu, F.F.; Dang, Q.; Pham, T.; Lombard, L.; Crous, P.W.; Wingfield, M.J. Ten new species of Calonectria from Indonesia and Vietnam. Mycologia 2019, 111, 78–102. [Google Scholar] [CrossRef] [PubMed]
- Nirenberg, H.I. A simplified method for identifying Fusarium spp. occurring on wheat. Can. J. Bot. 1981, 59, 1599–1609. [Google Scholar] [CrossRef]
- Hollander, M.; Wolfe, D.A.; Chicken, E. Nonparametric Statistical Methods; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Graça, R.N.; Alfenas, A.C.; Maffia, L.A.; Titon, M.; Alfenas, R.F.; Lau, D.; Rocabado, J.M.A. Factors influencing infection of Eucalypts by Cylindrocladium pteridis. Plant Pathol. 2009, 58, 971–981. [Google Scholar] [CrossRef]
- Mishra, K.K.; Kolte, S.J.; Nashaat, N.I.; Awasthi, R.P. Pathological and biochemical changes in Brassica juncea (mustard) infected with Albugo candida (white rust). Plant Pathol. 2009, 58, 80–86. [Google Scholar] [CrossRef]
- Pethybridge, S.J.; Nelson, S.C. Leaf Doctor: A new portable application for quantifying plant disease severity. Plant Dis. 2015, 99, 1310–1316. [Google Scholar] [CrossRef]
- Cunningham, C.W. Can three incongruence tests predict when data should be combined? Mol. Biol. Evol. 1997, 14, 733–740. [Google Scholar] [CrossRef]
- Crous, P.W.; Groenewald, J.Z.; Risede, J.M.; Simoneau, P.; Hywel-Jones, N.L. Calonectria species and their Cylindrocladium anamorphs: Species with sphaeropedunculate vesicles. Stud. Mycol. 2004, 50, 415–430. [Google Scholar]
- Kang, J.C.; Crous, P.W.; Old, K.M.; Dudzinski, M.J. Non–conspecificity of Cylindrocladium quinqueseptatum and Calonectria quinqueseptata based on a β–tubulin gene phylogeny and morphology. Can. J. Bot. 2001, 79, 1241–1247. [Google Scholar] [CrossRef]
- Chen, Q.Z.; Guo, W.S.; Ye, X.Z.; Huang, X.P.; Wu, Y.Z. Identification of Calonectria associated with Eucalyptus leaf blight in Fujian Province. J. Fujian Coll. For. 2013, 33, 176–182. [Google Scholar]
- Jiang, G.Z.; Gao, F.; Yue, H.; He, X.Y. First Report of Fruit Spot of Macadamia sp. Caused by Calonectria pentaseptata in China. Plant Dis. 2020, 104, 575. [Google Scholar] [CrossRef]
- Alfenas, R.F.; Pereira, O.L.; Freitas, R.G.; Freitas, C.S.; Dita, M.A.D.; Alfenas, A.C. Mass spore production and inoculation of Calonectria pteridis on Eucalyptus spp. under different environmental conditions. Trop. Plant Pathol. 2013, 38, 406–413. [Google Scholar] [CrossRef]
- Freitas, R.G.; Alfenas, R.F.; Guimarães, L.M.S.; Badel, J.L.; Alfenas, A.C. Genetic diversity and aggressiveness of Calonectria pteridis in Eucalyptus spp. Plant Pathol. 2019, 68, 869–877. [Google Scholar] [CrossRef]
- Gadgil, P.; Dick, M. Fungi silvicolae novazelandiae: 10. N. Z. J. For. Sci. 2004, 44, 1–7. [Google Scholar] [CrossRef]
- Lombard, L.; Crous, P.W.; Wingfield, B.D.; Wingfield, M.J. Phylogeny and systematics of the genus Calonectria. Stud. Mycol. 2010, 66, 31–69. [Google Scholar] [CrossRef]
- Crous, P.W.; Groenewald, J.Z.; Risede, J.M.; Simoneau, P.; Hyde, K.D. Calonectria species and their Cylindrocladium anamorphs: Species with clavate vesicles. Stud. Mycol. 2006, 55, 213–226. [Google Scholar] [CrossRef]
- Lombard, L.; Crous, P.W.; Wingfield, B.D.; Wingfield, M.J. Multigene phylogeny and mating tests reveal three cryptic species related to Calonectria pauciramosa. Stud. Mycol. 2010, 66, 15–30. [Google Scholar] [CrossRef]
- Kang, J.C.; Crous, P.W.; Schoch, C.L. Species concepts in the Cylindrocladium floridanum and Cy. spathiphylli complexes (Hypocreaceae) based on multi-allelic sequence data, sexual compatibility and morphology. Syst. Appl. Microbiol. 2001, 24, 206–217. [Google Scholar] [CrossRef]
- Lechat, C.; Crous, P.W.; Groenewald, J.Z. The enigma of Calonectria species occurring on leaves of Ilex aquifolium in Europe. IMA Fungus 2010, 1, 101–108. [Google Scholar] [CrossRef]
- Victor, D.; Crous, P.W.; Janse, B.J.H.; Wingfield, M.J. Genetic variation in Cylindrocladium floridanum and other morphologically similar Cylindrocladium species. Syst. Appl. Microbiol. 1997, 20, 268–285. [Google Scholar] [CrossRef]
- Lombard, L.; van der Merwe, N.A.; Groenewald, J.Z.; Crous, P.W. Generic concepts in Nectriaceae. Stud. Mycol. 2015, 80, 189–245. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Crous, P.W.; Wingfield, B.D.; Wingfield, M.J. The Cylindrocladium candelabrum species complex includes four distinct mating populations. Mycologia 1999, 91, 286–298. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Marincowitz, S.; Barnes, I.; Bensch, K.; Braun, U.; Camporesi, E.; Damm, U.; et al. Genera of phytopathogenic fungi: GOPHY 1. Stud. Mycol. 2017, 86, 99–216. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Mohammed, C.; Yuan, Z.Q. New foliar pathogens of Eucalyptus from Australia and Indonesia. Mycol. Res. 1998, 102, 527–532. [Google Scholar] [CrossRef]
- Crous, P.W.; Janse, B.J.H.; Victor, D.; Marais, G.F.; Alfenas, A.C. Characterization of some Cylindrocladium species with three-septate conidia using morphology, isozyme banding patterns and DNA polymorphisms. Syst. Appl. Microbiol. 1993, 16, 266–273. [Google Scholar] [CrossRef]
- Decock, C.; Crous, P.W. Curvicladium gen. nov., a new hyphomycete genus from French Guiana. Mycologia 1998, 90, 276–281. [Google Scholar] [CrossRef][Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).