
MAT Loci Play Crucial Roles in Sexual Development but Are Dispensable for Asexual Reproduction and Pathogenicity in Rice Blast Fungus Magnaporthe oryzae


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture
2.2. Sexual Reproduction
2.3. MAT Loci Deletion and Mutant Recovery
2.4. MAT Gene Deletion and Mutant Recovery
2.5. RNA Extraction and qRT-PCR
2.6. Microscopic Analysis
2.7. Subcellular Distribution
2.8. Assays for Asexual Reproduction, Carbon Utilization, Chemical Resistance and Pathogenicity-Related Developments
3. Results
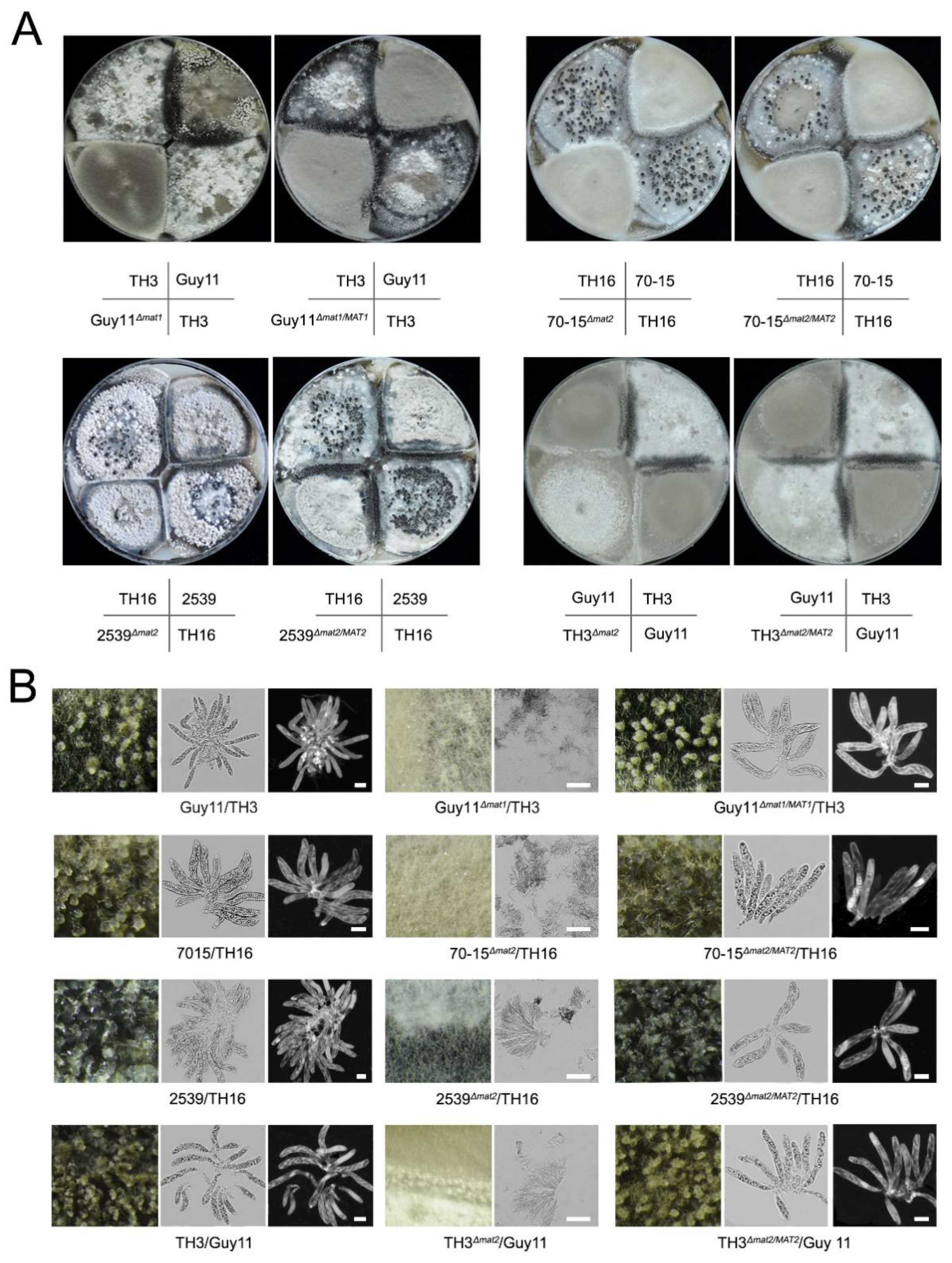
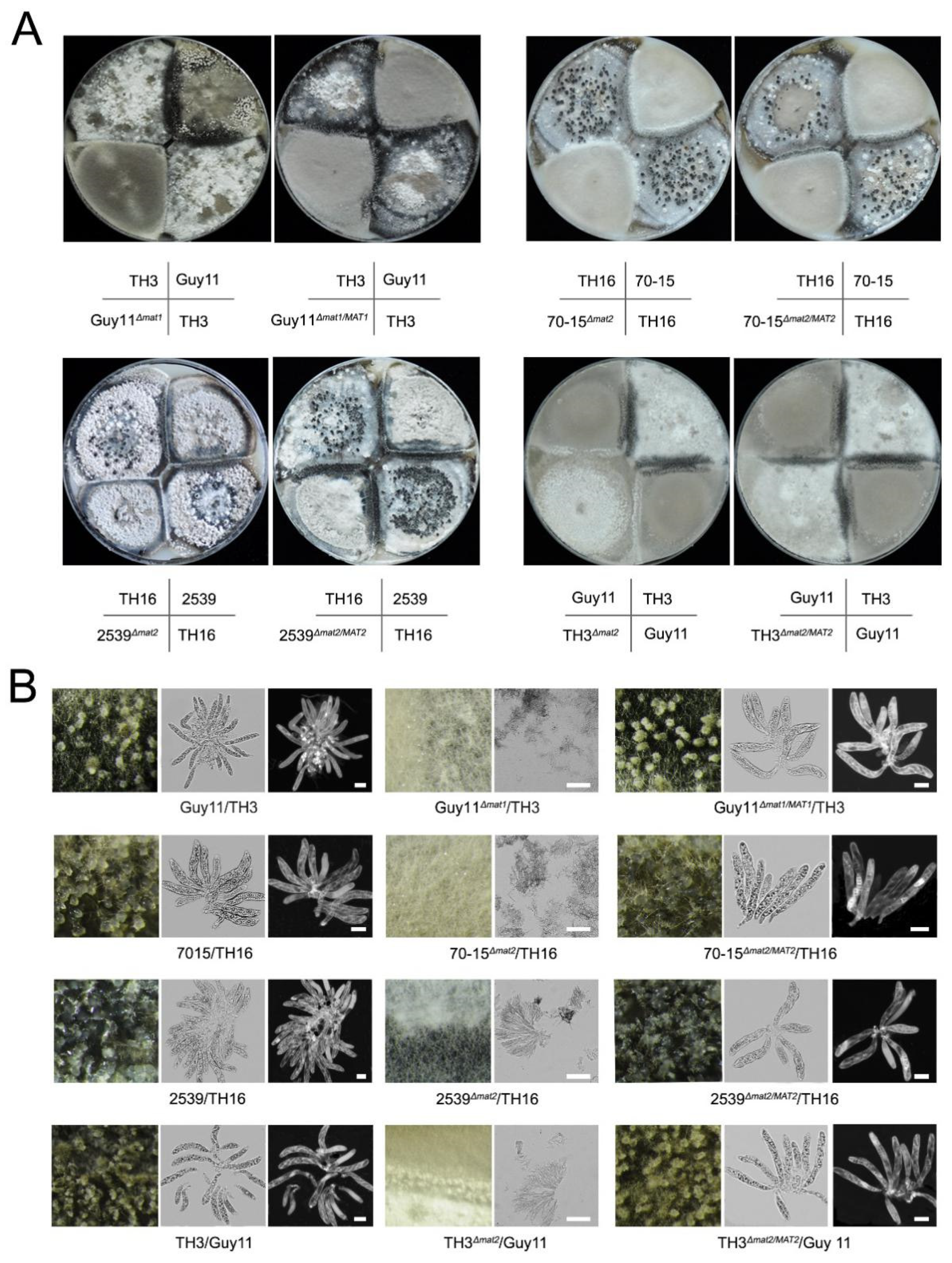
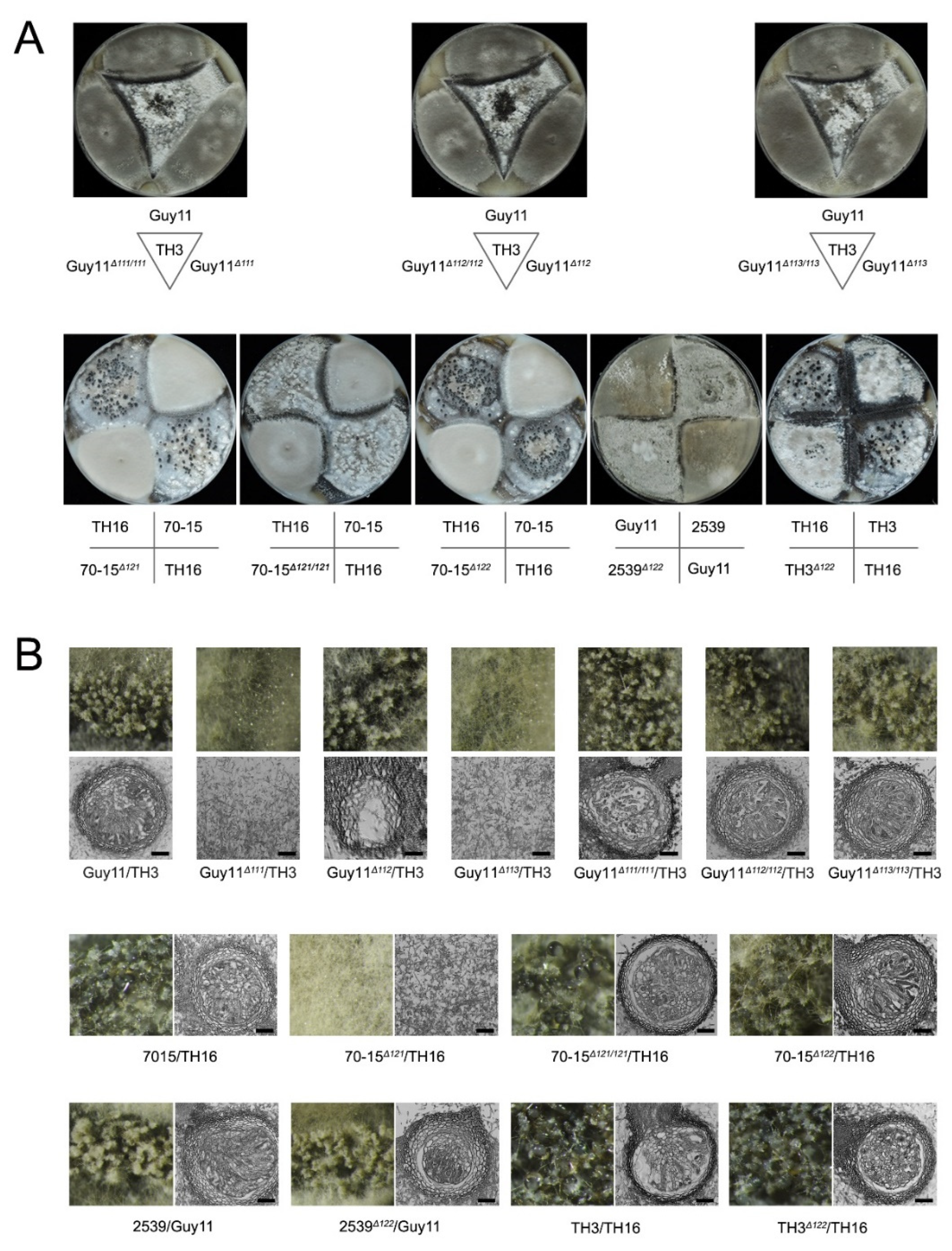
3.1. Both of the MAT Loci Are Indispensable for Sexual Reproduction
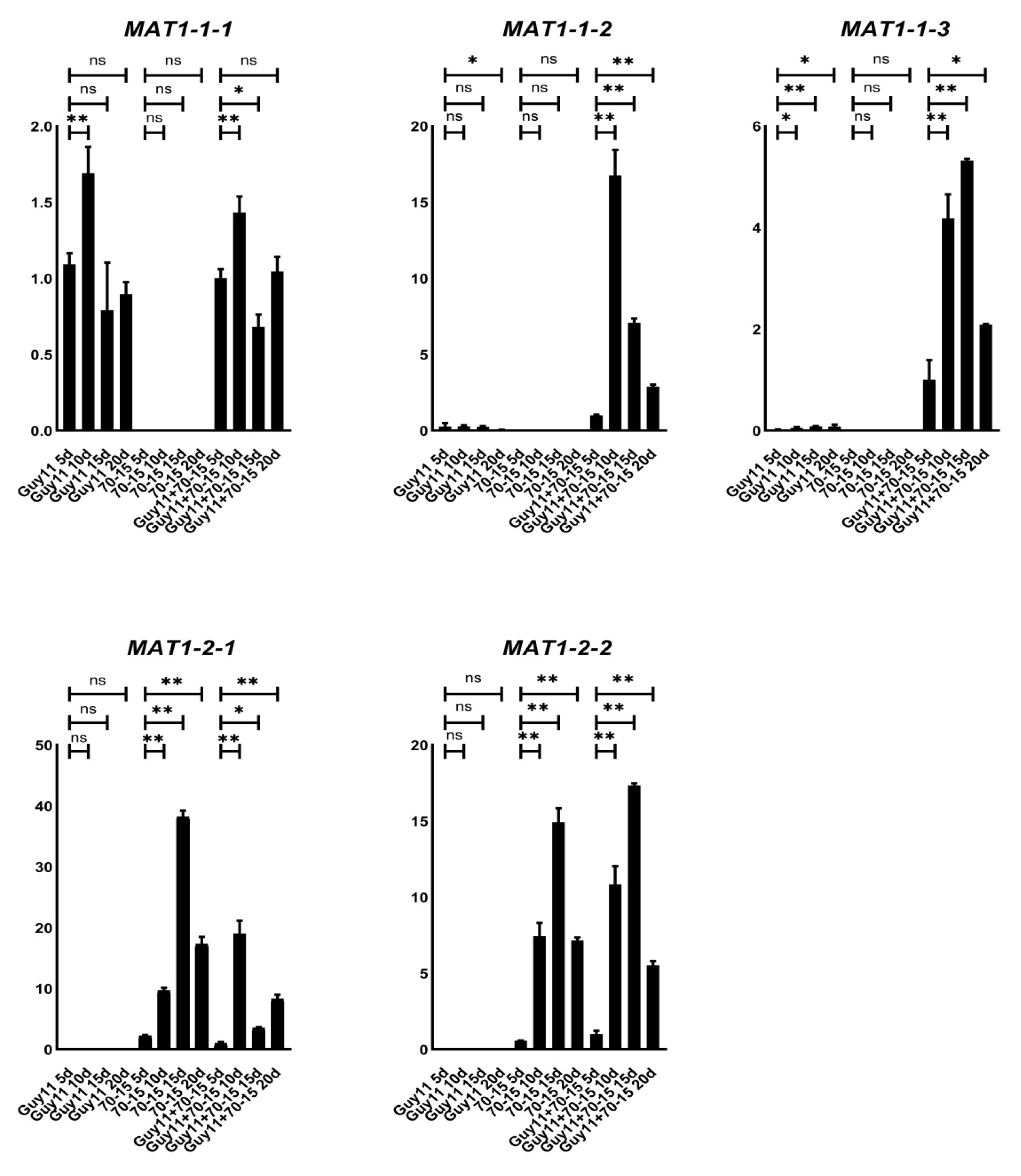
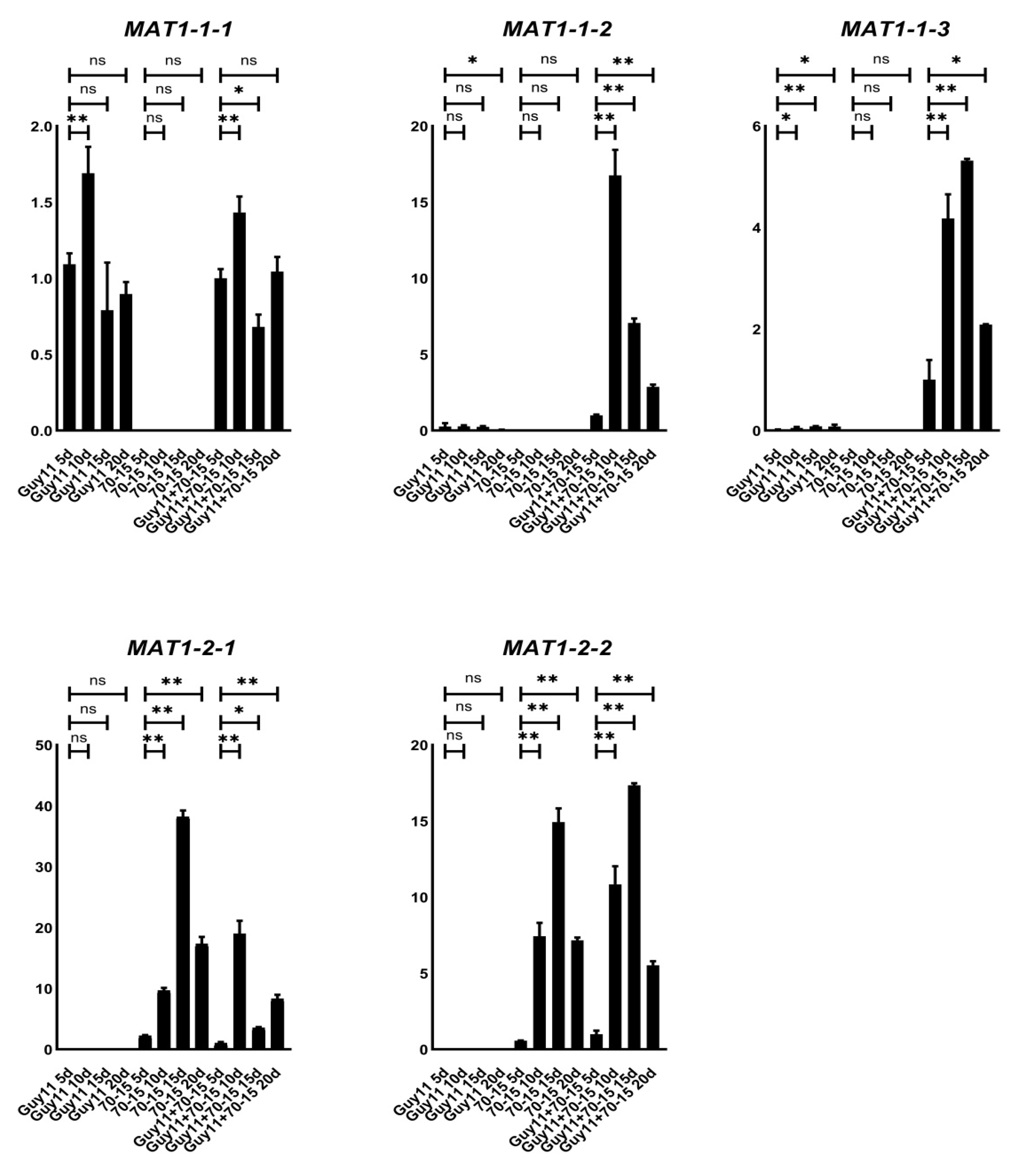
3.2. MAT1-1-2 and MAT1-1-3 Genes Are Up-Regulated during Sexual Reproduction
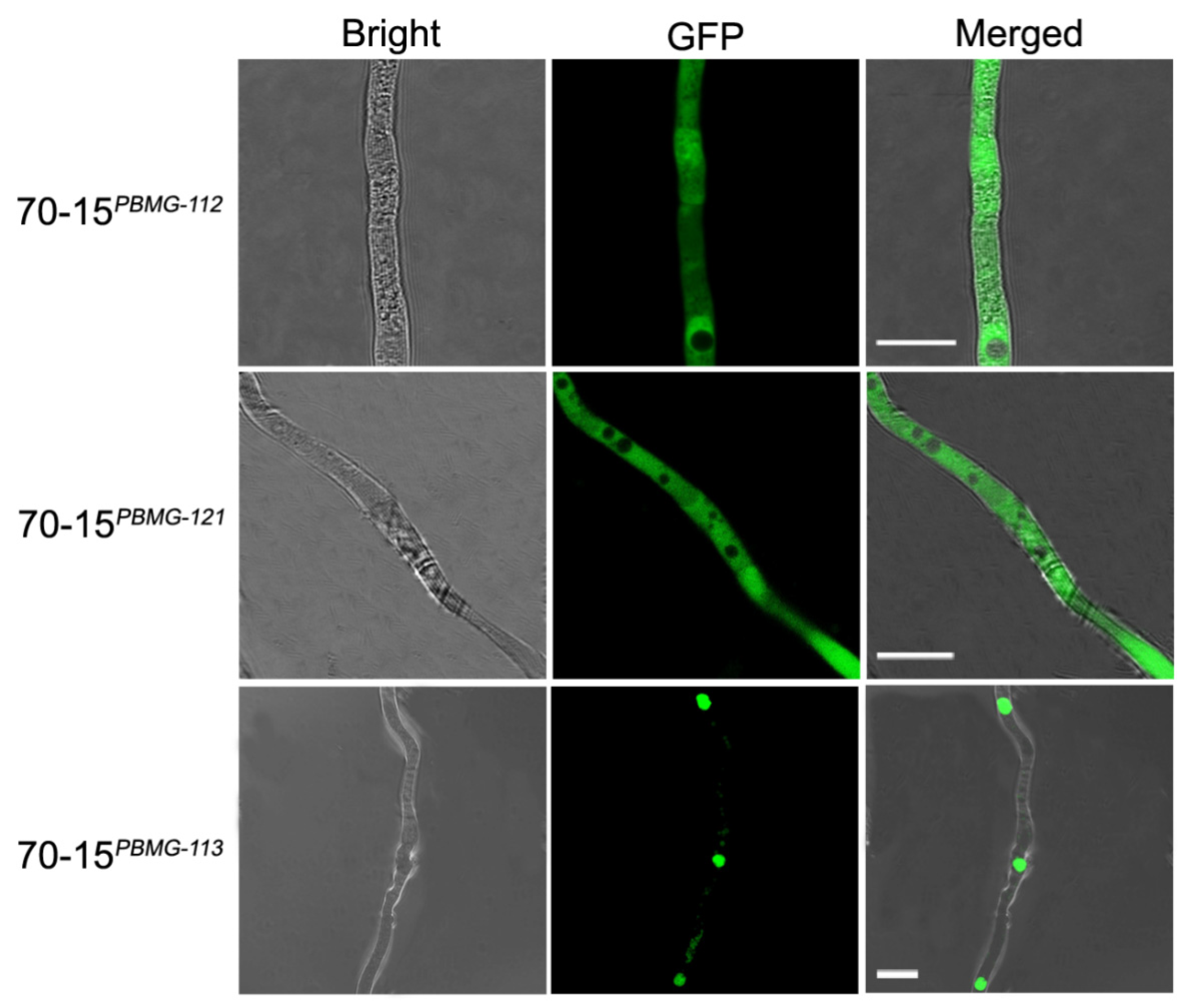
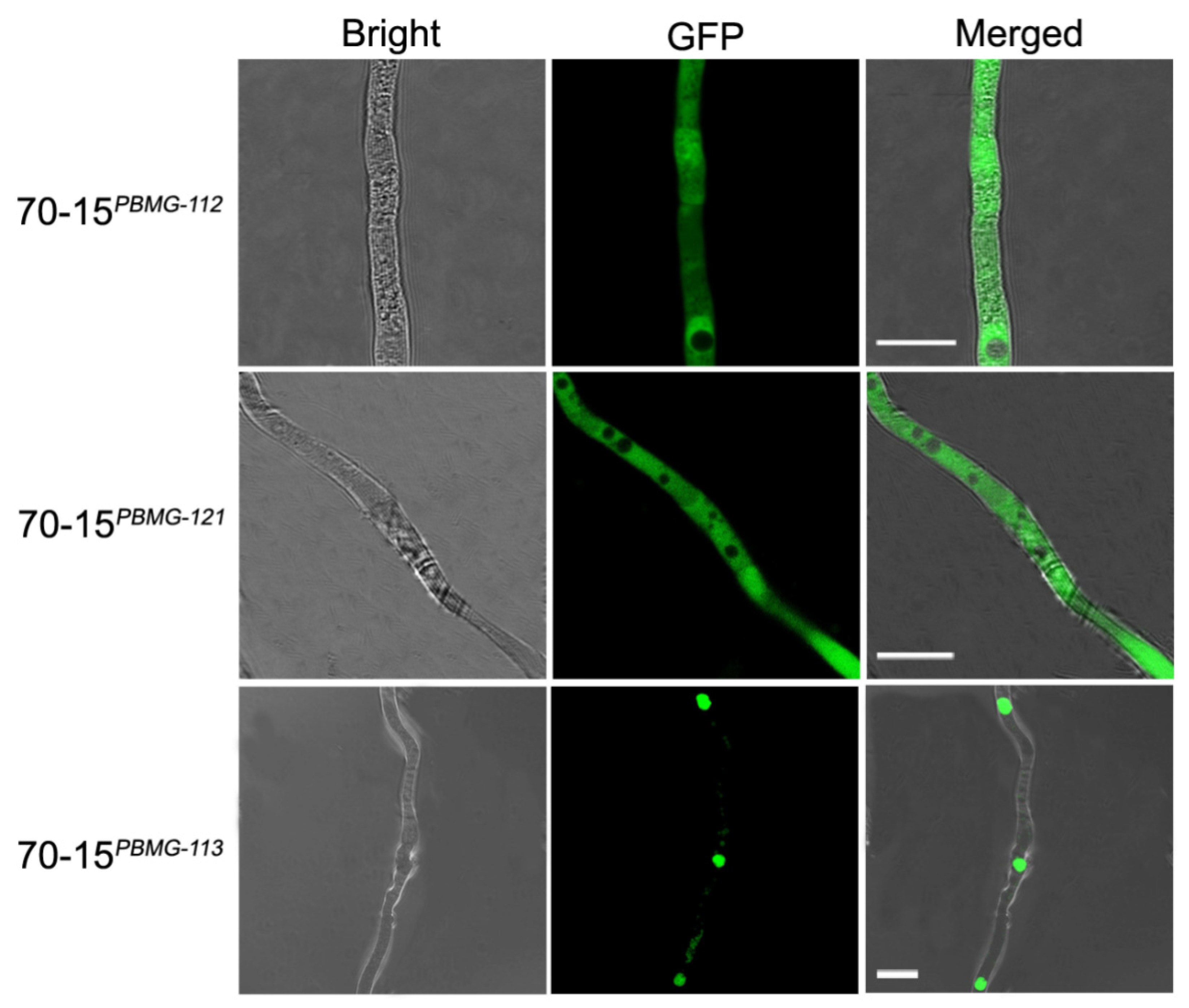
3.3. Subcellular Distribution of the MAT Proteins
3.4. MAT Genes Contribute Differently to the Formation of Sexual Structures
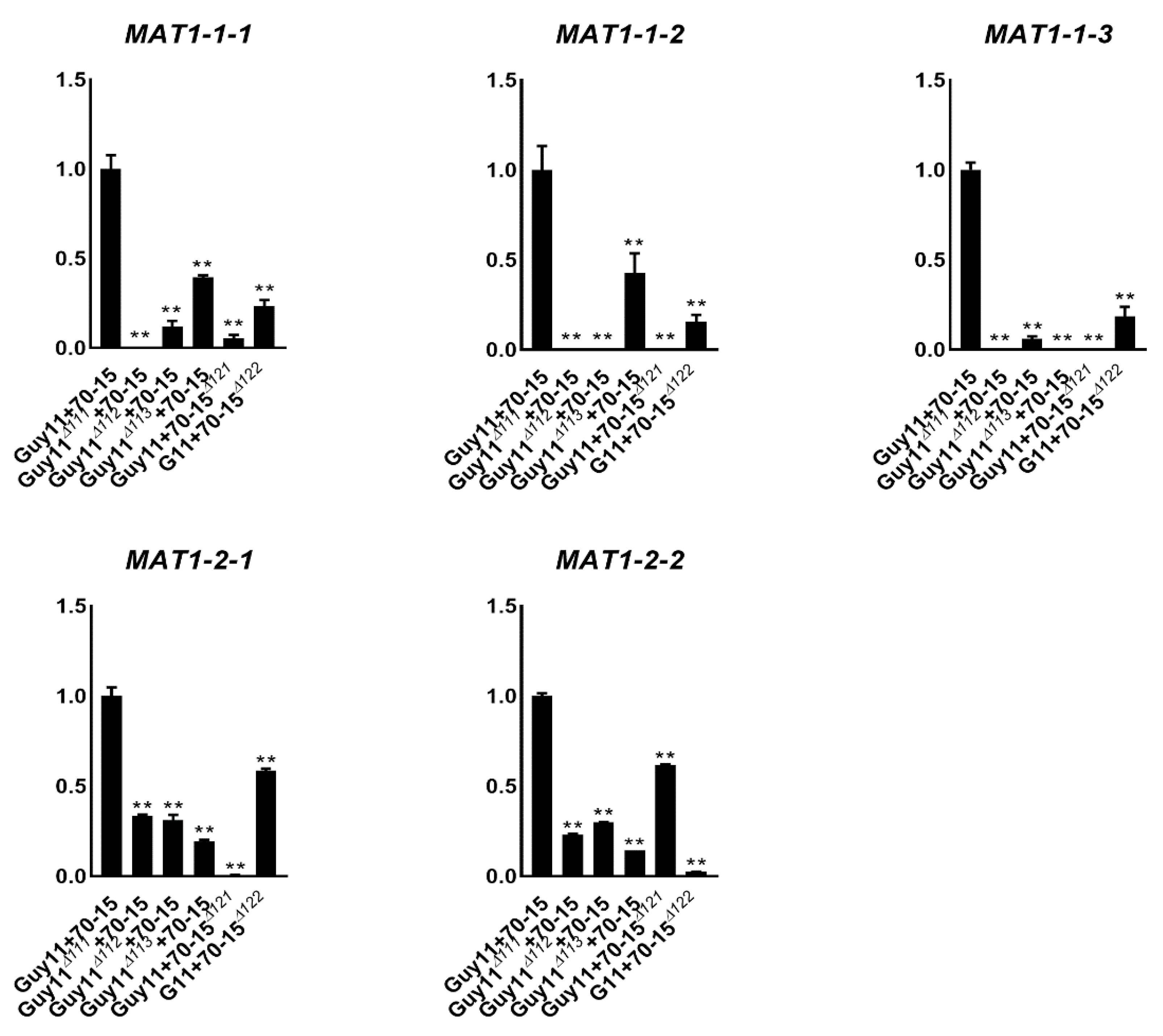
3.5. Deleting Any of the Individual MAT Genes Down-Regulated the Others
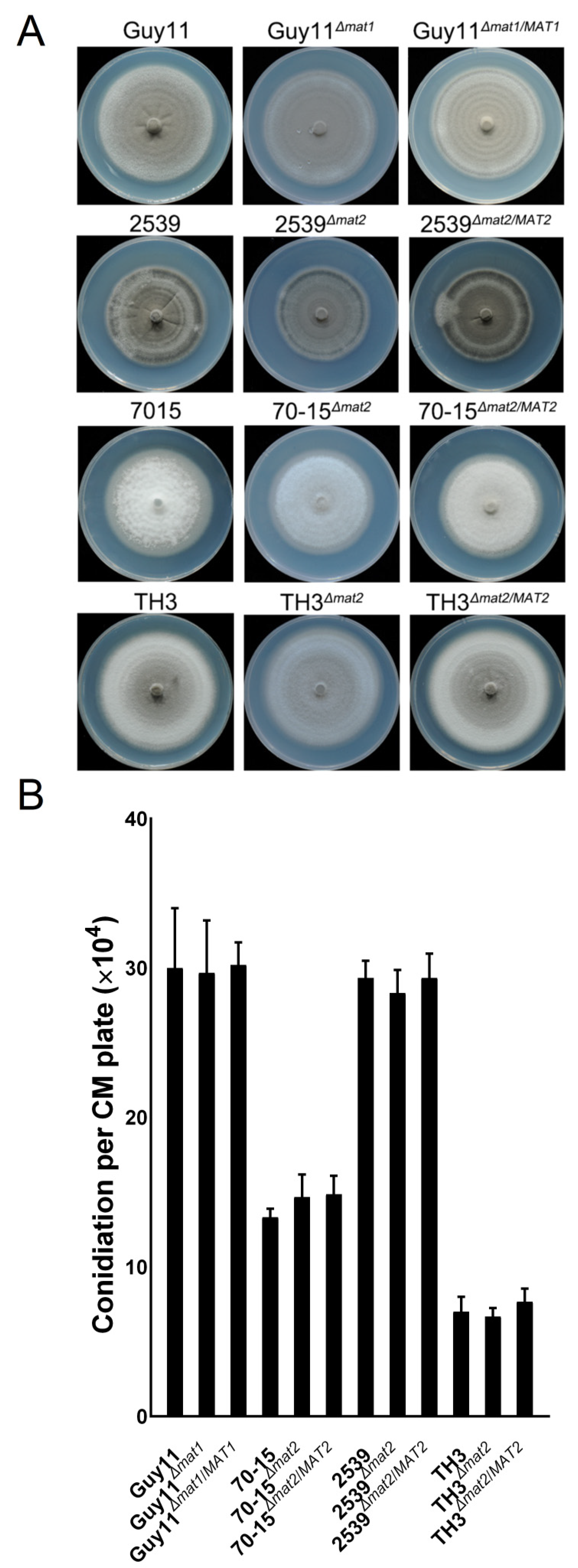
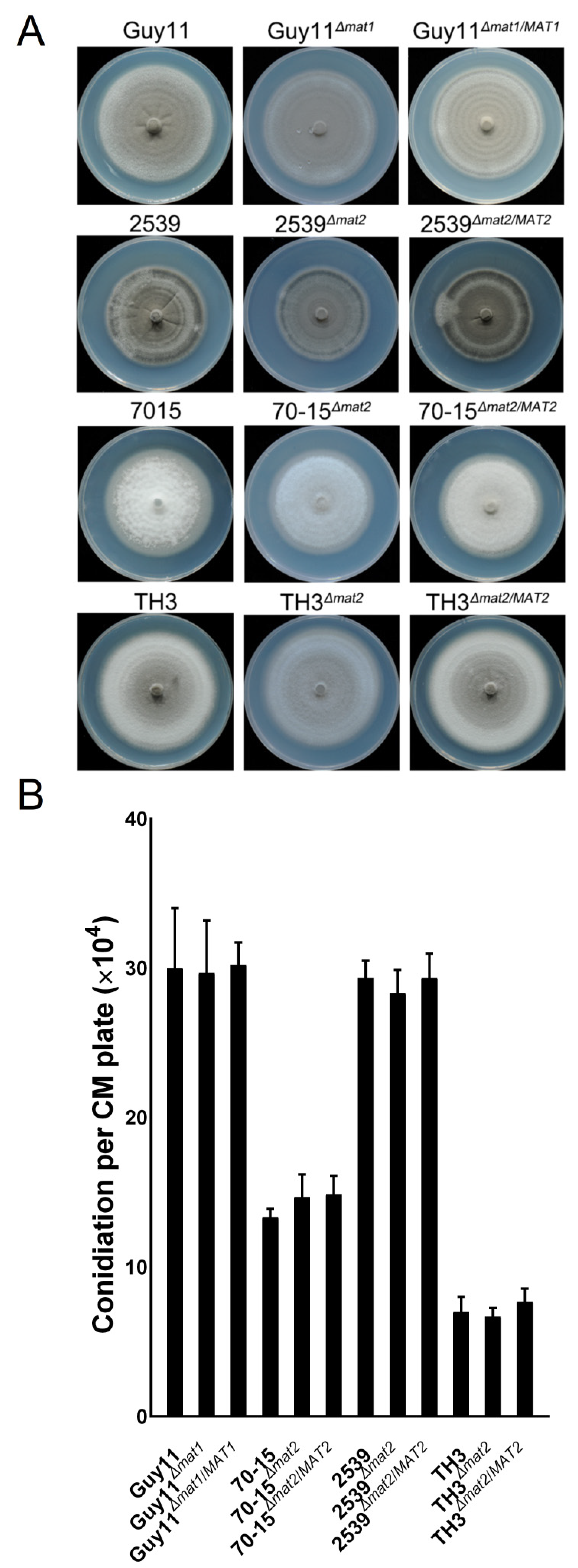
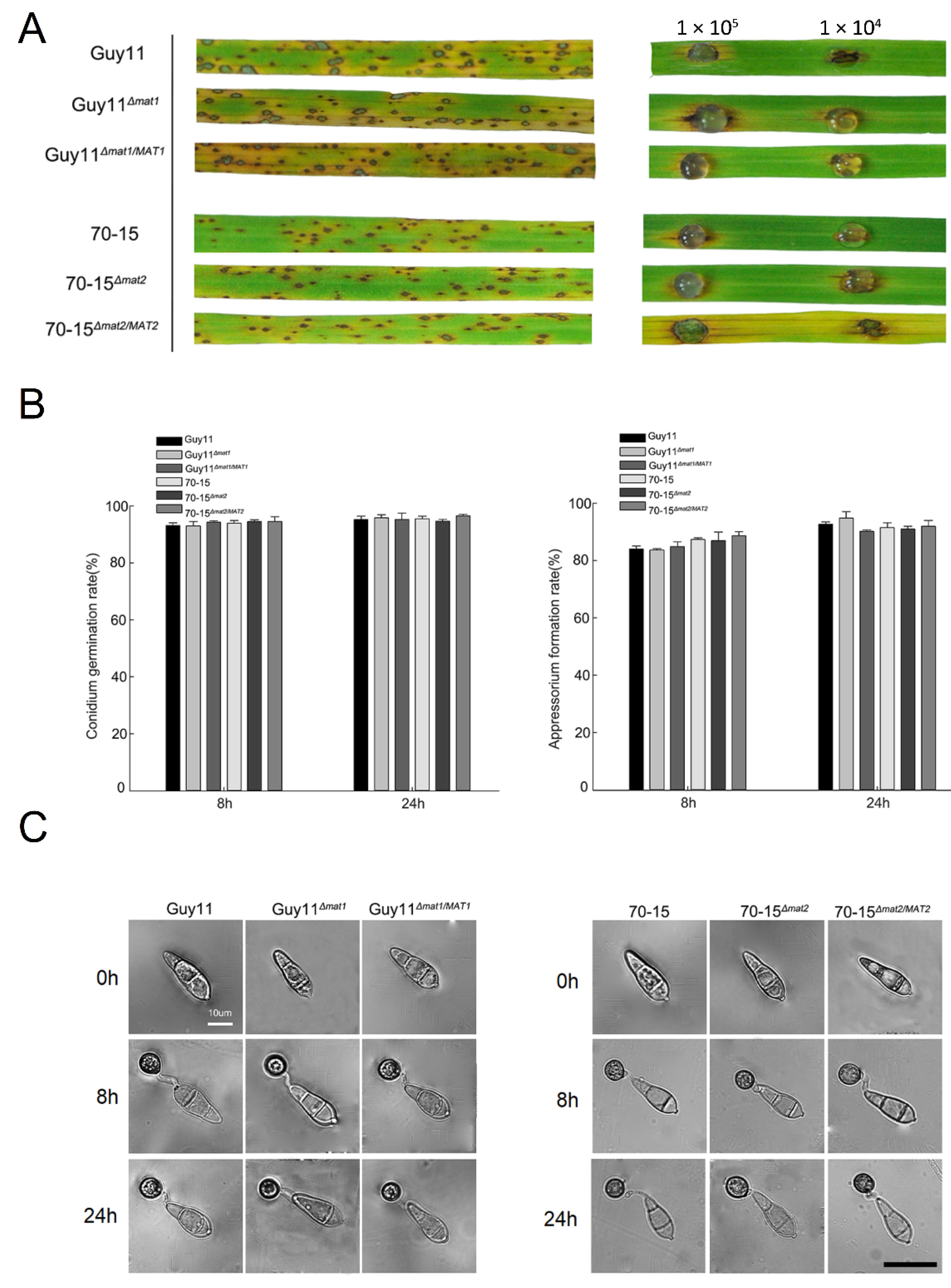
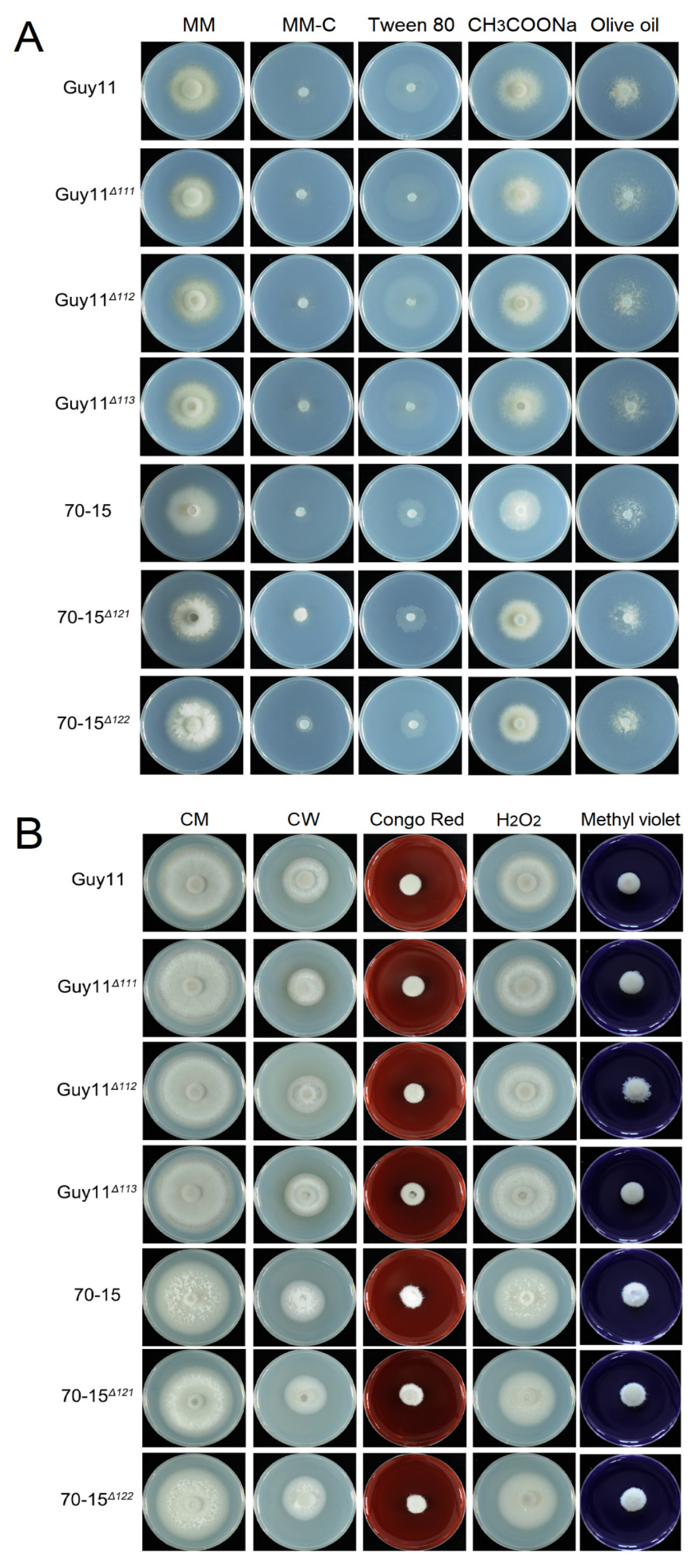
3.6. The MAT Loci Are not Required in Vegetative Growth, Asexual Reproduction or Pathogenicity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Nieuwenhuis, B.P.S.; James, T.Y. The frequency of sex in fungi. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 10, 371. [Google Scholar] [CrossRef] [Green Version]
- Fu, C.; Coelho, M.A.; David-Palma, M.; Priest, S.J.; Heitman, J. Genetic and genomic evolution of sexual reproduction: Echoes from LECA to the fungal kingdom. Curr. Opin. Genet. Dev. 2019, 10, 70–75. [Google Scholar] [CrossRef]
- Turgeon, B.G.; Yoder, O.C. Proposed nomenclature for mating type genes of filamentous ascomycetes. Fungal Genet. Biol. 2000, 31, 1–5. [Google Scholar] [CrossRef]
- Seibel, C.; Tisch, D.; Kubicek, C.P.; Schmoll, M. ENVOY is a major determinant in regulation of sexual development in Hypocrea jecorina (Trichoderma reesei). Eukaryot. Cell 2012, 11, 885–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.-W.; Yun, S.-H.; Lee, T.; Turgeon, B.G. Altering sexual reproductive mode by interspecific exchange of MAT loci. Fungal Genet. Biol. 2011, 48, 714–724. [Google Scholar] [CrossRef]
- Zheng, Q.; Hou, R.; Ma, J.; Wu, Z.; Wang, G.; Wang, C.; Xu, J.R. The MAT locus genes play different roles in sexual reproduction and pathogenesis in Fusarium graminearum. PLoS ONE 2013, 8, e66980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.-K.; Jo, S.-M.; Kim, G.-Y.; Kim, D.-W.; Kim, Y.-K.; Yun, S.-H. A large-scale functional analysis of putative target genes of mating-type loci provides insight into the regulation of sexual development of the cereal pathogen Fusarium graminearum. PLoS Genet. 2015, 11, e1005486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, R.; Kan, J.A.L.V.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Pietro, A.D.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.E. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Notteghem, J.L.; Silue, D. Distribution of the mating type alleles in Magnaporthe grisea populations pathogenic on rice. Phytopathology 1992, 82, 423–426. [Google Scholar] [CrossRef]
- Wilson, A.M.; Wilken, P.M.; Van Der Nest, M.A.; Wingfield, M.J.; Wingfield, B.D. It’s all in the genes: The regulatory pathways of sexual reproduction in filamentous ascomycetes. Genes 2019, 10, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautier, V.; Tong, L.; Nguyen, T.-S.; Debuchy, R.; Silar, P. PaPro1 and IDC4, Two genes controlling stationary phase, sexual development and cell degeneration in Podospora anserina. Fungi 2018, 4, 85. [Google Scholar] [CrossRef] [Green Version]
- Meng, S.; Brown, D.; Ebbole, D.; Torto, T.; Oh, Y.; Deng, J.; Mitchell, T.; Dean, R. Gene Ontology annotation of the rice blast fungus, Magnaporthe oryzae. BMC Microbiol. 2009, 9, S8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benkhali, A.; Coppin, E.; Brun, S.; Peraza-Reyes, L.; Martin, T.; Dixelius, C.; Lazar, N.; Van Tilbeurgh, H.; Debuchy, R. A network of HMG-Box transcription factors regulates sexual cycle in the fungus Podospora anserina. PLoS Genet. 2013, 9, e1003642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, D.; Xu, P.; Shen, Y.; Li, C.; Adreit, H.; Milazzo, J.; Ravigne, V.; Bazin, E.; Notteghem, J.-L.; Fournier, E.; et al. Sex at the origin: An Asian population of the rice blast fungus Magnaporthe oryzae reproduces sexually. Mol. Ecol. 2012, 21, 1330–1344. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lu, L.; Jia, Y.; Wang, Q.; Fukuta, Y.; Li, C. Characterization of field isolates of Magnaporthe oryzae with mating type, DNA fingerprinting, and pathogenicity assays. Plant Dis. 2016, 100, 298–303. [Google Scholar] [CrossRef]
- Li, L.; Wang, J.; Zhang, Z.; Wang, Y.; Liu, M.; Jiang, H.; Chai, R.; Mao, X.; Qiu, H.; Liu, F.; et al. MoPex19, which is essential for maintenance of peroxisomal structure and woronin bodies, is required for metabolism and development in the rice blast fungus. PLoS ONE 2014, 9, e85252. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Lee, T.; Lee, Y.W.; Yun, S.H.; Turgeon, B.G. Shifting fungal reproductive mode by manipulation of mating type genes: Obligatory heterothallism of Gibberella zeae. Mol. Microbiol. 2003, 50, 145–152. [Google Scholar] [CrossRef]
- Rodenburg, S.Y.A.; Terhem, R.B.; Veloso, J.; Stassen, J.H.M.; van Kan, J.A.L. Functional analysis of mating type genes and transcriptome analysis during fruiting body development of Botrytis cinerea. mBio 2018, 9, e01939-17. [Google Scholar] [CrossRef] [Green Version]
- Klix, V.; Nowrousian, M.; Ringelberg, C.; Loros, J.J.; Dunlap, J.C.; Pöggeler, S. Functional characterization of MAT1-1-specific mating-type genes in the homothallic ascomycete Sordaria macrospora provides new insights into essential and nonessential sexual regulators. Eukaryot. Cell 2010, 9, 894–905. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, A.V.-B.; An, Z.; Metzenberg, R.L.; Glass, N.L. Characterization of matA-2, matA-3 and ΔmatA mating-type mutants of Neurospora crassa. Genetics 1998, 148, 1069–1079. [Google Scholar] [CrossRef]
- Arnaise, S.; Debuchy, R.; Picard, M. What is a Bona Fide Mating-Type Gene? Internuclear complementation of mat mutants in Podospora anserina. Mol. Gen. Genet. 1997, 256, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Doughan, B.; Rollins, A. Characterization of MAT gene functions in the life cycle of Sclerotinia sclerotiorum reveals a lineage-specific MAT gene functioning in apothecium morphogenesis. Fungal Biol. 2016, 120, 1105–1117. [Google Scholar] [CrossRef]
- Gu, Z.; Li, L.; Wang, J.; Chai, R.; Wang, Y.; Zhang, Z.; Mao, X.; Qiu, H.; Sun, G. Observation of sexual structure of Magnaporthe oryzae via calcofluor white and nile red staining. Chin. J. Rice Sci. 2016, 30, 668–672. [Google Scholar]
- Guo, X.; Li, L.; Dong, B.; Wang, J.; Chai, R.; Zhang, Z.; Mao, X.; Qiu, H.; Hao, Z.; Wang, Y.; et al. Lighting the cellular structures of sexual generation in Magnaporthe oryzae with fluorescent proteins and fluorescent dyes. Chin. J. Cell Biol. 2018, 7, 1138–1145. [Google Scholar]
- Wang, J.; Guo, X.; Ling, L.; Qiu, H.; Zhen, Z.; Wang, Y.; Sun, G. Application of the fluorescent dye Bodipy in the study of lipid dynamics of the rice blast fungus Magnaporthe oryzae. Molecules 2018, 23, 1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Guy11 | TH3 | TH16 | |
|---|---|---|---|
| Guy1 | 1.3 ± 0.3 | ||
| Guy11Δmat | 0 | ||
| Guy11Δmat1/MAT | 1.1 ± 0.1 | ||
| TH3 | 1.3 ± 0.3 | ||
| TH3Δmat2 | 0 | ||
| TH3Δmat2/MAT2 | 1.4 ± 0.2 | ||
| 70-15 | 1.0 ± 0.2 | ||
| 70-15Δmat2 | 0 | ||
| 70-15Δmat2/MAT2 | 0.9 ± 0.1 | ||
| 2539 | 1.5 ± 0.3 | ||
| 2539Δmat2 | 0 | ||
| 2539Δmat2/MAT2 | 1.5 ± 0.3 |
| TH3 | TH16 | Guy11 | |
|---|---|---|---|
| Guy11 | 1.2 ± 0.3 | ||
| Guy11Δ111 | 0 | ||
| Guy11Δ112 | 1.2 ± 0.3 | ||
| Guy11Δ113 | 0 | ||
| Guy11Δ111/111 | 1.1 ± 0.2 | ||
| Guy11Δ112/112 | 1.2 ± 0.3 | ||
| Guy11Δ113/113 | 1.2 ± 0.2 | ||
| 70-15 | 1.0 ± 0.3 | ||
| 70-15Δ121 | 0 | ||
| 70-15Δ122 | 1.0 ± 0.2 | ||
| 70-15Δ121/121 | 0.9 ± 0.2 | ||
| 2539 | 1.3 ± 0.1 | ||
| 2539Δ122 | 1.3 ± 0.3 | ||
| TH3 | 1.6 ± 0.3 | ||
| TH3Δ122 | 1.6 ± 0.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.-y.; Wang, S.-z.; Zhang, Z.; Hao, Z.-n.; Shi, X.-x.; Li, L.; Zhu, X.-m.; Qiu, H.-p.; Chai, R.-y.; Wang, Y.-l.; et al. MAT Loci Play Crucial Roles in Sexual Development but Are Dispensable for Asexual Reproduction and Pathogenicity in Rice Blast Fungus Magnaporthe oryzae. J. Fungi 2021, 7, 858. https://doi.org/10.3390/jof7100858
Wang J-y, Wang S-z, Zhang Z, Hao Z-n, Shi X-x, Li L, Zhu X-m, Qiu H-p, Chai R-y, Wang Y-l, et al. MAT Loci Play Crucial Roles in Sexual Development but Are Dispensable for Asexual Reproduction and Pathogenicity in Rice Blast Fungus Magnaporthe oryzae. Journal of Fungi. 2021; 7(10):858. https://doi.org/10.3390/jof7100858
Chicago/Turabian StyleWang, Jiao-yu, Shi-zhen Wang, Zhen Zhang, Zhong-na Hao, Xiao-xiao Shi, Ling Li, Xue-ming Zhu, Hai-ping Qiu, Rong-yao Chai, Yan-li Wang, and et al. 2021. "MAT Loci Play Crucial Roles in Sexual Development but Are Dispensable for Asexual Reproduction and Pathogenicity in Rice Blast Fungus Magnaporthe oryzae" Journal of Fungi 7, no. 10: 858. https://doi.org/10.3390/jof7100858
APA StyleWang, J.-y., Wang, S.-z., Zhang, Z., Hao, Z.-n., Shi, X.-x., Li, L., Zhu, X.-m., Qiu, H.-p., Chai, R.-y., Wang, Y.-l., Li, L., Liu, X.-h., Feng, X.-x., Sun, G.-c., & Lin, F.-c. (2021). MAT Loci Play Crucial Roles in Sexual Development but Are Dispensable for Asexual Reproduction and Pathogenicity in Rice Blast Fungus Magnaporthe oryzae. Journal of Fungi, 7(10), 858. https://doi.org/10.3390/jof7100858