Abstract
The Qinghai-Xizang Plateau, known for its high altitude, geological history of plate collision, crustal uplift, and special ecology factors, provides an ideal environment for studying fungal biodiversity in extreme environmental conditions. Some species within the Conocybe, containing secondary metabolites such as psilocybin, phallotoxins, and amatoxins, have potential medicinal value for treating psychiatric disorders and for use in drug development. This study investigates Conocybe (Bolbitiaceae, Agaricales) on the Plateau, based on specimens collected over the past decade, using morphological and molecular phylogenetic analyses. Seven species were identified, including four new species: C. alticola, C. alticoprophila, C. versicolor, and C. yadongensis. Molecular analyses, utilizing multi-gene sequence data (ITS, nrLSU, and tef-1α), support the taxonomic position of these new species within this genus as new species. Detailed descriptions, illustrations, photographs, line drawings, and comparisons with related species are provided for the new taxa. This study enriches the species diversity of Conocybe on the Qinghai-Tibet Plateau, further enhancing our understanding of fungal biodiversity in this region.
1. Introduction
The Qinghai-Xizang Plateau (abbreviation Q-X Plateau), with an average elevation exceeding 4000 m, and referred to as the “roof” of the world, is the highest and youngest plateau on earth [1]. It is known for its complex geological evolution, including plate collisions, crustal uplift, diverse climatic conditions, and unique ecological features [2,3,4,5,6]. As an “ecological island” isolated from the surrounding lower regions, the plateau has fostered a distinctive high-altitude biogeographic system [7,8]. These exceptional natural conditions have not only promoted the formation of rich and unique biodiversity in the region, but also made it an invaluable resource for global scientific research [9,10,11]. The region’s diverse plant communities and unique environmental conditions provide an ideal natural platform for studying fungal adaptive evolution, making the Q-X Plateau a critical area for exploring high-altitude ecosystems and fungal biodiversity [12,13,14]. Due to challenges posed by remote location, limited accessibility and harsh climatic conditions, research on fungal communities in this region has progressed slowly, leaving numerous valuable fungal resources yet to be explored [15,16].
Conocybe Fayod belongs to Bolbitiaceae (Agaricales) [17], is characterized by its small to medium-sized delicate basidiomata, conical pileus, adnexed to adnate and brown-rusty lamellae, powdery or pilose stipes, lecythiform cheilocystidia, and basidiospores with germ pore [18,19]. Most Conocybe species are saprophytic and are widely distributed in different vegetation types, particularly in fertile soils and herbivore dung, as well as on plant debris, decaying wood, and sawdust [18,20,21,22]. The genus Conocybe is noted for its chemical diversity and biological activities, despite some species containing neurotoxic or cytotoxic substances [23,24,25,26]. It has been believed that secondary metabolites like psilocybin, with potential therapeutic effects, and toxic cyclic peptides such as amatoxins and phallotoxins may cause severe health issues [27,28,29,30].
Conocybe is divided into more than 10 sections based on morphological characteristics, including sect. Conocybe Fayod, sect. Mixtae Singer, sect. Candidae Watling, and sect. Pilosellae Singer [18,31,32,33,34]. Currently, approximately 270 species of Conocybe have been identified worldwide, with 44 recorded in China [18,19,35,36,37]. Recent studies have focused on Conocybe specimens from various regions in China. Song and Bau (2023) described seven new species and one new record from sect. Pilosellae collected in northeastern, northern, and central–southern China [18]. In 2024, Wang et al. described C. himalayana Ke Wang, T.Z. Wei & P. Hong from sect. Pilosellae based on five specimens collected from the Q-X Plateau [16], and Lu et al. reported two new species from sect. Conocybe based on four specimens from the Yunnan-Guizhou Plateau [36]. However, research on Conocybe in the Q-X Plateau remains limited. This study investigates Conocybe specimens collected over the past decade from the plateau, combining macroscopic, microscopic, and molecular methods to explore species diversity and distribution in the region.
2. Materials and Methods
2.1. Samplings and Morphological Study
The specimens for this study were collected from Lhasa Municipality, Qamdo Municipality, Shigatse Municipality, Lingzhi Prefecture, and the Ali Region in the Xizang Autonomous Region in China between 2015 and 2024. Photographs were captured using a Canon EOS 80D camera (Tokyo, Japan), and specimens were carefully packed individually in aluminum foil to prevent mixing or damage. Specimens were dried completely using a food dryer at 50 °C, sealed in plastic bags, and deposited in the Mycological Herbarium of the Institute of Microbiology, Chinese Academy of Sciences (HMAS). Microscopic features such as basidia, basidiospores, pileipellis, and cheilocystidia were observed under an Olympus CX31 light microscope (Tokyo, Japan), with at least 30 measurements obtained for each feature. Microscopic examination followed the methodology of Largent [38]. Tissues were treated with 5% KOH and sterile water for observation. Measurements are presented as (a)b–c(d), where b–c represents at least 90% of the measured values, while a and d indicate the lowest value and the highest value, respectively. The Q value is the length/width ratio of basidiospores, with Qm representing the mean Q value ± standard deviation [39]. The color designations were determined using the Methuen Handbook of Colour [40].
2.2. DNA Extraction, PCR and Sequencing
DNA was extracted from dried specimens using the Broad-spectrum Plant Rapid Genomic DNA Kit (Biomed, Beijing, China), following the manufacturer’s instructions. The primer pairs ITS1F/ITS4 [41], LR7/LR0R [42], and EF983F/EF2218R [43] were used to amplify the nuclear internal transcribed spacer (ITS), the large subunit nuclear ribosomal DNA (nrLSU), and the translation elongation factor subunit 1 alpha (tef-1α) regions, respectively. The polymerase chain reaction (PCR) procedure was performed under the following conditions: an initial denaturation at 94 °C for 5 min, followed by 35 cycles of denaturation at 94 °C for 60 s, annealing at 53 °C (ITS), 50 °C (nrLSU), or 55 °C (tef-1α) for 60 s, and extension at 72 °C for 90 s, with a final extension at 72 °C for 10 min [44,45,46]. The PCR products were detected by electrophoresis and sent to BGI Genomics Co., Ltd. (Beijing, China) for purification and sequencing.
2.3. Phylogenetic Analyses
Based on BLAST searches against GenBank and previous studies, we analyzed the nrITS, nrLSU and tef-1α sequences of 124 taxa, as detailed in Table 1. The sequences were aligned using Muscle version 3.6 separately [47], then manually adjusted in BioEdit version 7.0.4 to remove the ambiguous areas [48]. The aligned sequences were assembled in PhyloSuite version 1.2.3 [49]. Maximum likelihood (ML) analysis was performed on the concatenated sequences using raxmlGUI 1.3 with a GTRGAMMA model and 1000 rapid bootstrap replicates [50]. The best partitioning scheme and evolutionary models for three predefined partitions were selected using PartitionFinder2 v2.1.1 [51], based on the greedy algorithm and AICc criterion: GTR+I+G for ITS, GTR+I+G for nrLSU, and SYM+I+G for tef-1α. Bayesian Inference (BI) analysis was conducted using MrBayes v3.2.7a [52], with six Markov chains run for two million generations and trees sampled every 100th generation. The burn-ins were determined using Tracer version 1.6, with an ESS value higher than 200. The remaining trees were used to calculate Bayesian posterior probabilities (PP). The trees were displayed in Interactive Tree of Life (iTOL) version 6 [53].

Table 1.
Sequences used in the phylogenetic analysis in this study. Missing sequences are indicated by “–”.
3. Results
3.1. Phylogeny
In the phylogenetic analysis, 24 specimens from seven Conocybe species were included, with the following four species from the Psathyrellaceae Vilgalys, Moncalvo & Redhead selected as outgroups: Psathyrella piluliformis (Bull.) P.D. Orton, P. amygdalinospora T. Bau & J.Q. Yan, Candolleomyces sichuanicus R.L. Zhao, B. Cao & X.X. Han, and C. singeri (A.H. Sm.) D. Wächt. & A. Melzer. Furthermore, it is noteworthy that Song (2024), through morphological observations and sequence comparisons, proposes that Pholiotina pleurocystidiata Hauskn. & Krisai should be considered a synonym of Psathyrella piluliformis [62,64]. In total, 72 new sequences were generated in this study, which were from 24 specimens from Qinghai-Xizang Plateau, China, all with the nrITS, nrLSU, and tef-1α sequences. The combined dataset with 3310 characters including gaps (864 for nrITS, 1299 for nrLSU, and 1147 for tef-1α) was included in the phylogenetic analyses. The phylogenetic trees of ML and MrBayes were almost identical. The ML tree is shown in Figure 1 with bootstrap values and Bayesian posterior probabilities indicated on the branches.

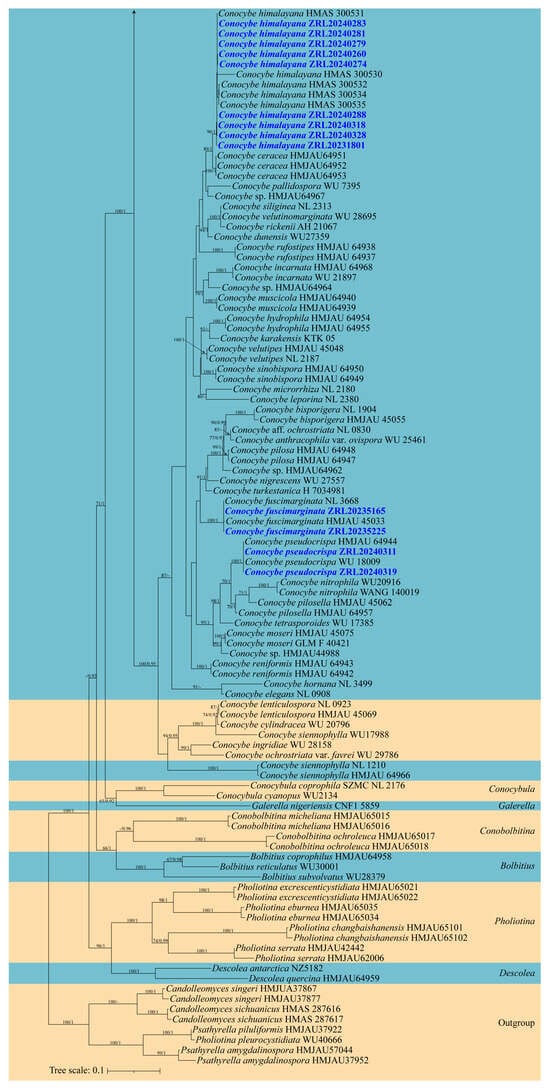
Figure 1.
Molecular phylogenetic analyses of Conocybe species by the maximum likelihood (ML) method based on combined nrITS, nrLSU and tef-1α sequences. Maximum likelihood bootstrap support values (ML) ≥ 60% and Bayesian posterior probabilities (PP) ≥ 0.90 are shown at the nodes as ML/PP. Sequences newly generated in this study are highlighted in colored font.
3.2. Taxonomy
Conocybe alticola R.L. Zhao & X.X. Han, sp. nov., Figure 2

Figure 2.
Basidiomata and microscopic features of Conocybe alticola. (A–D) Basidiomata: (A–C) ZRL20240316 (holotype); (D) ZRL20240317, (E) Basidiospores, (F) Basidia, (G) Cheilocystidia, (H) Pileipellis. Scale bars: 10 mm (A–D); 5 μm (E); 10 μm (F–H).
Fungal Names: FN 572250
Holotype: China. Xizang Autonomous Region, Ali Region, Zhada County, Rural Road 748, Qinipu, N 31°45′43″ E 79°32′49″, 4058 m asl, 5 August 2024, Xi-Xi Han, Lin-Hui Liu ZRL20240316 (holotype HMAS 287991). GenBank: PQ699270 (nrITS), PQ699294 (nrLSU), PQ836632 (tef-1α).
Etymology: alticola (Latin) refers to the habitat of the species, which is adapted to high-altitude environments.
Diagnosis: Conocybe alticola is distinguished by its hemispherical, obtusely conical to convex pileus with a non-striate surface and a pale yellow hue, the stipe slightly enlarged at base, sometimes forming a bulb. The narrowly ellipsoid to oblong basidiospores, with a germ pore. Grows on alpine meadows in summer.
Macroscopic description: Pileus 1.2–2.0 cm diam., hemispherical, obtusely conical to convex, not hygrophanous, faintly pubescent, non-striate, edge decurved, sometimes with slightly undulated edge, carnosus; yellowish white (4A2) to greyish yellow (4B3), blond (4C4) to olive brown (4D6) at the center, edge paler, pruinose surface. Context thin, fleshy, yellowish white (4A2) to blond (4C4), indistinct odor. Lamellae are distant to nearly distant, narrowly adnate to adnexed, unequal in length, orange–white (5A2) to greyish yellow (4B3) with pale edge when young, light brown (6D7) when mature. Stipes 1.3–3.8 × 0.2–0.3 cm, cylindrical, slightly enlarged at base, sometimes forming a bulb, slightly longitudinally striated, with blond (4C4) pruinose, yellowish white (4A2) to greyish yellow (4C5). Odor not distinctive. Taste indistinct.
Microscopic description: Basidiospores (9.4)9.5–10.5(11.1) × (5.5)6.0–6.7(7.1) μm, Q = 1.48–1.70, Qm = 1.59 (±0.11), narrowly ellipsoid to oblong, with germ pore, slightly thick-walled, contains oil droplets, pale yellow (4A3) to greyish yellow (4C6) in water, greyish orange (6B3) to light brown (6D7) in 5% KOH. Basidia (23.5)24.5–27.1(28.5) × (8.1)9.0–10.0(10.4) μm, clavate, sometimes with vacuolar contents, two or four spored. Cheilocystidia (14.5)16.7–21.2(25.2) × (6.9)8.0–9.8(10.7) μm, lecythiform, with 3.9–4.6 μm wide capitula. Pileipellis hymeniform, consist of broadly clavate, obovoid, or spheropedunculate elements, (25.9)29.1–42.7(54.2) × (10.2)12.3–18.8(20.4) μm. Pleurocystidia clavate or lecythiform, and rare in number. Pileocystidia not observed. Clamp connections present.
Habit and habitat: Summer solitary or scattered in the alpine meadows. So far found only in the Xizang Autonomous Region, China.
Other specimens examined: China. Xizang Autonomous Region, Ali Region, Gaer County, Zuozuo Township, N 32°23′55″ E 80°14′59″, 4409 m asl, 31 July 2024, Xi-Xi Han, Lin-Hui Liu ZRL20240256 (HMAS 287982); Xizang Autonomous Region, Ali Region, Gaer County, Zuozuo Township, N 32°23′55″ E 80°14′59″, 4409 m asl, 31 July 2024, Xi-Xi Han, Lin-Hui Liu ZRL20240259 (HMAS 287983); Xizang Autonomous Region, Ali Region, Zhada County, Rural Road 748, Qinipu, N 31°45′43″ E 79°32′49″, 4058 m asl, 5 August 2024, Xi-Xi Han, Lin-Hui Liu ZRL20240317 (HMAS 287992); Xizang Autonomous Region, Ali Region, Zhada County, Paerlong, N 31°15′39″ E 79°54′16″, 4179 m asl, 6 August 2024, Xi-Xi Han, Lin-Hui Liu ZRL20240327 (HMAS 287995).
Notes: Phylogenetically and morphologically, the new species C. alticola is closely related to C. ammophila (Figure 1) [67], but can be distinguished by its hemispherical, obtusely conical to convex, non-striate, carnosus pileus; pale yellow hue stipe; narrower, greyish orange to light brown in 5% KOH basidiospores and two or four-spored basidia, while C. ammophila, originally described from Greenland, has a pale stipe, broader spores, and four-spored basidia [67,68]. Conocybe alticola is widely distributed in the alpine and subalpine regions of the southwestern Q-X Plateau.
Conocybe alticoprophila R.L. Zhao & X.X. Han, sp. nov., Figure 3
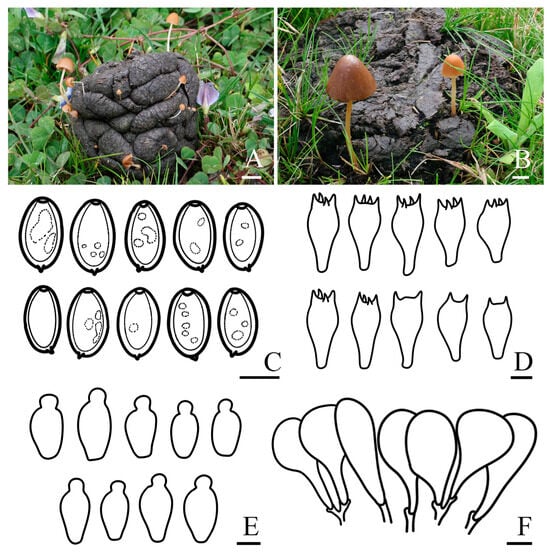
Figure 3.
Basidiomata and microscopic features of Conocybe alticoprophila. (A,B) Basidiomata: (A) ZRL20220041; (B) ZRL20220046 (holotype), (C) Basidiospores, (D) Basidia, (E) Cheilocystidia, (F) Pileipellis. Scale bars: 10 mm (A,B); 10 μm (C–F).
Fungal Names: FN 572251
Holotype: China. Xizang Autonomous Region, Shigatse Municipality, Yadong County, Lower Yadong Township, N 27°13′12″ E 88°34′48″, 2872 m asl, 26 July 2022, Rui-Lin Zhao, Bin Cao ZRL20220046 (holotype HMAS 287975). GenBank: PQ699274 (nrITS), PQ699298 (nrLSU), PQ836636 (tef-1α).
Etymology: “alti-” derived from the Latin word “altus”, meaning “high-altitude”; “-coprophila” derived from the Greek words “copros” and “philos”, meaning “dung” and “fondness”, respectively.
Diagnosis: Conocybe alticoprophila is characterized by its small to medium-sized basidiomata, yellowish-brown, hygrophanous pileus, which is translucently striate to 4/5 from the edge to the center. The basidiospores are slightly amygdaliform, and basidia are two or four spored. Grows on cow dung in grasslands.
Macroscopic description: Pileus 0.2–3.0 cm diam., obtusely conical, hemispherical to conical–convex, hygrophanous, translucently striate to 4/5 from the edge to the center when moist, surface smooth, edge decurved, sometimes with slightly undulated edge; greyish orange (5B4) to yellowish brown (5E5) at the center, turning orange–white (5A2) to yellowish brown (5D8) towards edge. Context thin, and indistinct odor. Lamellae are moderately crowded, narrowly adnate to adnexed, orange–white (5A2) to yellowish brown (5D5). Stipes 0.8–5.2 × 0.1–0.3 cm, cylindrical, fragile, slightly enlarged at the base forming a weak bulb, slightly longitudinally striate, with orange–white (5A2) pruinose, orange–white (5A1) to brownish orange (5C5). Odor not distinctive. Taste indistinct.
Microscopic description: Basidiospores (15.9)16.4–17.7(18.0) × (9.7)10.0–11.0(12.0) μm, Q = 1.55–1.71, Qm = 1.63 (±0.08), narrowly ellipsoid to oblong, slightly amygdaliform, with germ pore, thick-walled, contains oil droplets, pale orange (5A3) to brownish yellow (5C7) in water, light brown (6D6) to dark brown (6F7) in 5% KOH. Basidia (26.3)27.1–31.4(33.5) × (14.2)14.7–15.8(16.6) μm, clavate to broadly clavate, sometimes with vacuolar contents, two or four spored. Cheilocystidia (19.7)21.5–27.0(30.7) × (11.2)12.2–15.5(17.5) μm, lecythiform, with 4.7–7.0 μm wide capitula. Pileipellis hymeniform, consist of clavate or spheropedunculate elements, (26.2)27.6–44.0(47.3) × (10.5)12.9–22.0(24.0) μm. Pleurocystidia lecythiform, and rare in number. Pileocystidia not observed. Clamp connections present.
Habit and habitat: Summer solitary or scattered on cow dung in grasslands. So far found only in the Xizang Autonomous Region, China.
Other specimens examined: China. Xizang Autonomous Region, Shigatse Municipality, Yadong County, Lower Yadong Township, N 27°13′12″ E 88°34′48″, 2872 m asl, 26 July 2022, Rui-Lin Zhao, Bin Cao ZRL20220041 (HMAS 287973); Xizang Autonomous Region, Shigatse Municipality, Yadong County, Lower Yadong Township, N 27°13′12″ E 88°34′48″, 2872 m asl, 26 July 2022, Rui-Lin Zhao, Bin Cao ZRL20220050 (HMAS 287976).
Notes: C. alticoprophila is a coprophilous species characterized by a small basidioma, conical and yellowish-brown pileus, and a cylindrical stipe with finely pruinose surface, it is similar to C. pubescens (Gillet) Kühner [69]. However, C. pubescens has a smaller basidiospores and pileus lack translucently striate when moist. The pileus of C. alticoprophila is obtusely conical, hemispherical to conical–convex with a cylindrical stipe that is slightly enlarged at the base forming a weak bulb. Morphologically, it resembles C. watlingii Hauskn, and is closely related phylogenetically. In contrast, the basidiospores and basidia of C. watlingii are narrower, its pileus lacks translucently striate edges when moist, and the cheilocystidia are slightly narrower with smaller capitula [70]. C. alticoprophila is phylogenetically and morphologically closely related to C. cylindrospora T. Bau & J. Liu, but the latter has smaller basidiospores that are cylindrical to ellipsoid-oblong, four-spored basidia which are also smaller, and smaller cheilocystidia [19]. The Conocybe sp. from Uruguay (PP949255 and PP949256) in the NCBI GenBank shows a high sequence similarity with the ITS of C. alticoprophila, due to partial sequence gaps and the lack of morphological information for the specimens, we cannot confirm whether they represent the same species.
Conocybe versicolor R.L. Zhao & X.X. Han, sp. nov., Figure 4

Figure 4.
Basidiomata and microscopic features of Conocybe versicolor. (A–C) Basidiomata: (A,B) ZRL20220299 (holotype); (C) ZRL20152517, (D) Basidiospores, (E) Basidia, (F) Cheilocystidia, (G) Pileipellis. Scale bars: 10 mm (A–C); 5 μm (D); 10 μm (E–G).
Fungal Names: FN 572252
Holotype: China. Xizang Autonomous Region, Shigatse Municipality, Yadong County, Lower Yadong Township, N 27°25′20″ E 88°55′6″, 3254 m asl, 27 July 2022, Rui-Lin Zhao, Bin Cao ZRL20220299 (holotype HMAS 287978). GenBank: PQ699290 (nrITS), PQ699314 (nrLSU), PQ836639 (tef-1α).
Etymology: versi- = variable, -color = color, referring to the basidiomata of the species having variable colors.
Diagnosis: Pileus surface of Conocybe versicolor is smooth with a slightly undulated edge. The stipe is cylindrical, hallow, slightly enlarged at the base, covered with scattered to dense pruinose, and no radicating structures were observed. Basidiospores are ellipsoid to oblong. Grow on the ground in mixed forests.
Macroscopic description: Pileus 2.9–4.6 cm diam., obtusely conical, conical–convex to campanulate with or without distinct broad umbo, hardly expanding, sometimes wrinkled, edge slightly undulated, slightly hygrophanous, not striate, surface smooth; brownish orange (5C5) to yellowish brown (5D8) at the center, edge orange–white (5A2) to greyish orange (5B4). Context thin, yellowish white (3A2) to greyish yellow (3C4), and odor indistinct. Lamellae are slightly crowded, narrowly adnate to adnexed, unequal in length, with smooth edges. Stipes 9.7–15.1 × 0.2–0.4 cm, cylindrical, hallow, slightly enlarged at the base, with scattered to dense orange–white (5A2) pruinose, and no radicating structures were observed. Odor not distinctive. Taste indistinct.
Microscopic description: Basidiospores (8.2)8.4–9.1(9.7) × (4.9)5.4–6.1(6.6) μm, Q = 1.43–1.63, Qm = 1.53 (±0.10), ellipsoid to oblong, slightly angular, with germ pore, thick-walled, contains oil droplets, yellowish white (4A2) to greyish yellow (4B4) in water, brownish orange (6C6) to brown (6E8) in 5% KOH. Basidia (18.2)19.7–22.3(23.6) × (7.4)7.8–8.6(9.3) μm, clavate, with vacuolar contents, two or four-spored. Cheilocystidia (15.1)16.9–19.6(20.9) × (6.9)7.5–9.0(10.0) μm, lecythiform, with 3.4–4.5 μm wide capitula. Pileipellis hymeniform, consist of broadly clavate, obovoid, or spheropedunculate elements, (26.9)33.3–44.9(52.4) × (14.0)17.1–23.2(26.5) μm. Pleurocystidia clavate or lecythiform, and rare in number. Pileocystidia not observed. Clamp connections present.
Habit and habitat: Summer solitary on the ground in mixed forests. So far found only in the Xizang Autonomous Region, China.
Other specimens examined: China. Xizang Autonomous Region, Lingzhi Prefecture, Lulang Town, N 29°46′18″ E 94°44′12″, 3326 m asl, 22 September 2015, Sheng-Yu Su ZRL20152517 (HMAS 287972).
Notes: Conocybe versicolor has a conical–convex to campanulate pileus, pale orange to greyish orange, resembling C. solitaria K.A. Thomas, Hauskn. & Manim. It shares similar basidiospore size and clavate basidia with C. solitaria, but the latter can be distinguished by its decurved pileus margin with faint radial striations, and larger basidia [71]. Additionally, C. versicolor has a similar stipe, basidiospores size and two or four-spored basidia to C. humicola (Thiers) Hauskn., Krisai & Voglmary, but C. humicola differs in having smaller basidiomata, shorter basidia, shorter cheilocystidia and a pileipellis consistsing of roundish-stipitate elements [67]. Phylogenetically, C. versicolor is closely related to C. antipus (Lasch) Kühner, which was originally described from North America. Both species share similar basidiospore sizes and occasionally wrinkled pileus, but C. antipus has slightly larger four-spored basidia, and long radicating stipes up to 40 mm [68,70].
Conocybe yadongensis R.L. Zhao & X.X. Han, sp. nov., Figure 5

Figure 5.
Basidiomata and microscopic features of Conocybe yadongensis. (A–C) Basidiomata ZRL20220042 (holotype), (D) Basidiospores, (E) Basidia, (F) Cheilocystidia, (G) Pileipellis. Scale bars: 10 mm (A–C); 5 μm (D); 10 μm (E–G).
Fungal Names: FN 572253
Holotype: China. Xizang Autonomous Region, Shigatse Municipality, Yadong County, Lower Yadong Township, N 27°13′12″ E 88°34′48″, 2872 m asl, 26 July 2022, Rui-Lin Zhao, Bin Cao ZRL20220042 (holotype HMAS 287974). GenBank: PQ699291 (nrITS), PQ699315 (nrLSU), PQ836640 (tef-1α).
Etymology: yadongensis (Latin) refers to Yadong County, Xizang, the locality of the type specimen.
Diagnosis: Conocybe yadongensis is characterized by small- to medium-sized basidiomata with a yellowish-brown pileus that is translucently striate and has a surface that is barely pruinose. The stipe is faintly longitudinally striated, and covered with strong pruinosity. The basidiospores are narrowly ellipsoid to oblong. Grows in the grassland.
Macroscopic description: Pileus 0.7–2.2 cm diam., hemispherical when young, obtusely conical, convex to plano-convex when mature, hygrophanous, translucently striate to 4/5 from the edge to the center when moist, faintly pubescent, surface hardly pruinose, edge slightly undulated; brownish orange (5C5) to yellowish brown (5E8) at the center, edge orange–white (5A2) to brownish orange (5C4). Context thin, slightly fragile, orange–white (5A2) to brownish orange (5C5), odor indistinct. Lamellae are slightly crowded, narrowly adnate to adnexed, unequal in length, surface spotted, pale orange (5A3) to light brown (5D6). Stipes 1.6–5.4 × 0.1–0.3 cm, cylindrical, hallow, slightly enlarged at the base, strongly pruinose and slightly longitudinally striate, greyish orange (5B3) to light brown (5D6).
Microscopic description: Basidiospores (8.2)8.4–9.1(9.5) × (4.8)5.0–5.5(5.7) μm, Q = 1.59–1.74, Qm = 1.66 (±0.07), narrowly ellipsoid to oblong, some lentiform, with germ pore, slightly thin walled, contains oil droplets, orange–white (5A2) to greyish orange (5B4) in water, brownish orange (5C5) to yellowish brown (5E7) in 5% KOH. Basidia (14.6)16.1–19.5(21.8) × (6.6)7.6–10.0(11.1) μm, clavate, sometimes with vacuolar contents, two or four spored. Cheilocystidia (18.3)20.4–24.4(26.5) × (9.2)9.4–11.8(13.5) μm, lecythiform, with 5.3–7.2 μm wide capitula. Pileipellis hymeniform, consist of broadly clavate, obovoid, or spheropedunculate elements, (26.9)34.3–50.9(65.9) × (13.3)17.0–27.0(34.7) μm. Pleurocystidia utriform, broadly clavate or lecythiform, and rare in number. Pileocystidia not observed. Clamp connections present.
Habit and habitat: Summer scattered in the grassland. So far found only in the Xizang Autonomous Region, China.
Notes: In the phylogenetic analyses, C. yadongensis is closely related to C. coniferarum E.F. Malysheva, C. echinata (Velen.) Singer, and C. qujingensis W.H. Lu, Karunarathna & Tibpromma (Figure 1). However, C. coniferarum has longer stipe with brownish hues, broader basidiospores and four-spored basidia [22], while C. echinata has a dark brown to dull brown pileus and larger cheilocystidia [35,72]. Additionally, C. qujingensis has a longer stipe, larger cheilocystidia, and pileipellis consists of clavate and spheropedunculate elements [36].
4. Discussion
This study revisits the taxonomy of Conocybe specimens from the Q-X Plateau based on the phylogenetic frameworks established by Tóth et al. [21], Lu et al. [36], and Song and Bau [62], alongside the morphological classification by Hausknecht [34]. We incorporated macrofungal specimens collected from the Q-X Plateau between 2015 and 2024, and employed a concatenated dataset (including ITS, nrLSU, and tef1-α sequences) to reassess the classification of Conocybe in this region. Based on the results, we describe four new species: C. alticola, C. alticoprophila, C. versicolor, and C. yadongensis, from various locations, including the Ali Region, Shigatse Municipality, and Lingzhi Prefecture. These newly identified species expand the known diversity of Conocybe on the Q-X Plateau and offer preliminary insights into the species adaptation to high-altitude, cold-climate habitats. Additionally, there are three known species, C. himalayana, C. pseudocrispa (Hauskn.) Arnolds, and C. fuscimarginata, from Lhasa Municipality, Qamdo Municipality and the Ali Region. Notably, the latter two species are reported here for the first time from the Q-X Plateau. This study broadens the understanding of Conocybe diversity in this unique high-altitude region and provides foundational data on its distribution patterns in cold environments.
The formation of basidioma is closely influenced by environmental factors, such as temperature, light, and nutrients [73]. On the Q-X Plateau, most Conocybe taxa exhibit temperate characteristics, but C. alticola, which we have newly described and has tough basidioma and relatively short stipes, is an exception. This aligns with the toughness-protection hypothesis proposed by Krah et al., which suggests that tough-fleshed basidioma reduce water loss, helping macrofungi cope with extreme microclimatic fluctuations [74]. Despite extensive surveys across various regions of Hengduan Mountains and Xizang Autonomous Region, C. alticola has only been found in the Ali region of Xizang Autonomous Region, which is known for long daylight hours, low temperatures, intense sunlight, and high altitudes (with an average elevation exceeding 4500 m) [75,76].
Phylogenetically, C. alticola is clustered with C. qujingensis, C. echinate, and C. coniferarum, forming a distinct lineage, and is sister to C. ammophila (Figure 1), which is described as being found only in cold climates with extreme temperature fluctuations and low rainfall, such as Greenland, Mongolia, and Russia. It is one of the stoutest members of the genus [68], which aligns with the characteristics of C. alticola. In contrast, C. qujingensis, C. echinata, and C. coniferarum exhibit slender stipes and relatively fragile pileus [22,35,36]. The variation in macroscopic morphology partially reveals that species within the genus may undergo adaptive evolution changes when facing extreme environmental stress. To further substantiate this conclusion, additional sampling outside the current study area is required to explore the broader distribution of these species.
Currently, a total of 15 Conocybe species have been recorded from the Q-X Plateau, including C. apala (Fr.) Arnolds, C. fragilis (Peck) Singer, C. himalayana, C. macrocephala Kühner & Watling, C. macrospora (G.F. Atk.) Hauskn., C. ochracea (Kühner) J. Favre, C. siliginea (Fr.) Kühner, C. subovalis Kühner & Watling, C. tenera (Schaeff.) Kühner, and six species identified in this study [77,78,79,80,81,82,83,84,85]. In addition, Conocybe is primarily distributed in Jilin Province and Inner Mongolia, where 26 and 9 species have been reported, respectively [36]. Limited records have also been made in regions such as Guangxi Province, Hunan Province, Hubei Province, Yunnan Province, Taiwan, Tianjin Municipality, and Xinjiang Uygur Autonomous Region [19,57,77,78,85]. However, in comparison to the total number of known species, the diversity of Conocybe in China remains relatively underrepresented.
The classification of Conocybe remains challenging, primarily due to the discordance between the clades within genus and morphological sections, as well as the presence of morphologically similar species, which complicates accurate species identification [21,62]. Traditional identification relies on morphological features such as pileus, lamellae, stipe, which are influenced by environmental conditions and growth stages, leading to variation. Additionally, some species lack sufficient and reliable molecular data, making it difficult to conduct a phylogenetic analysis for accurate classification [36]. Therefore, a combined approach using multi-gene phylogenetic analysis and morphological characteristics, along with the inclusion of multidimensional regional samples, is essential for further clarifying the distribution and classification of this genus in China.
This study describes four new species and reports the first records of two known species from the Qinghai-Xizang Plateau. By combining morphological and molecular phylogenetic analyses, it provides preliminary insights into the adaptive evolution of macrofungi in extreme high-altitude cold climates, contributing new perspectives on fungal diversity and species distribution on the Plateau. It also offers valuable data for understanding species distribution patterns and environmental adaptations in high-altitude ecosystems and provides empirical support for the toughness-protection hypothesis.
Author Contributions
Conceptualization, X.-X.H. and D.P.; field sampling, X.-X.H., D.P., B.C., J.-X.L., X.-Y.Z., L.-H.L. and R.-L.Z.; molecular experiments and data analysis, X.-X.H., L.-H.L. and D.P.; morphological observation, X.-X.H.; writing—original draft preparation, X.-X.H.; writing—review and editing, N.T., K.D.H., R.-L.Z. and B.C.; project administration, B.C.; funding acquisition, D.P. and R.-L.Z. All authors have read and agreed to the published version of the manuscript.
Funding
The Project of Science and Technology Programs of Tibet (XZ202202YD0031C), the Survey of Wildlife Resources in Key Areas of Tibet (ZL202203601), the Biodiversity Survey and Assessment Project of the Ministry of Ecology and Environment, China (2019HJ2096001006), and the National Natural Science Foundation of China (31961143010).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All sequence data are available in NCBI GenBank following the accession numbers in the manuscript.
Acknowledgments
We thank the Mycological Herbarium, Institute of Microbiology, Chinese Academy of Sciences (HMAS), and all the individuals in our laboratory. We thank Shaun Pennycook (Manaaki Whenua Landcare Research, New Zealand) for his valuable suggestions on the fungal nomenclature.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhu, Q.; Chen, H.; Peng, C.; Liu, J.; Piao, S.; He, J.-S.; Wang, S.; Zhao, X.; Zhang, J.; Fang, X. An early warning signal for grassland degradation on the Qinghai-Tibetan Plateau. Nat. Commun. 2023, 14, 6406. [Google Scholar] [CrossRef]
- Mao, K.S.; Wang, Y.; Liu, J.Q. Evolutionary origin of species diversity on the Qinghai–Tibet Plateau. J. Syst. Evol. 2021, 59, 1142–1158. [Google Scholar] [CrossRef]
- Renner, S.S. Available data point to a 4-km-high Tibetan Plateau by 40 Ma, but 100 molecular-clock papers have linked supposed recent uplift to young node ages. J. Biogeogr. 2016, 43, 1479–1487. [Google Scholar] [CrossRef]
- Yang, M.X.; Werth, S.; Wang, L.S.; Scheidegger, C. Phylogeographic analyses of an epiphytic foliose lichen show multiple dispersal events westward from the Hengduan Mountains of Yunnan into the Himalayas. Ecol. Evol. 2022, 12, e9308. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Deane, D.C.; Sui, X.; Fang, S.; Chu, C.; Liu, Y.; He, F. Testing multiple hypotheses for the high endemic plant diversity of the Tibetan Plateau. Glob. Ecol. Biogeogr. 2019, 28, 131–144. [Google Scholar] [CrossRef]
- Ding, L.; Kapp, P.; Cai, F.; Garzione, C.N.; Xiong, Z.; Wang, H.; Wang, C. Timing and mechanisms of Tibetan Plateau uplift. Nat. Rev. Earth Environ. 2022, 3, 652–667. [Google Scholar] [CrossRef]
- Deng, T.; Wu, F.; Zhou, Z.; Su, T. Tibetan Plateau: An evolutionary junction for the history of modern biodiversity. Sci. China Earth Sci. 2020, 63, 172–187. [Google Scholar] [CrossRef]
- Liu, H.; Wang, W.; Song, G.; Qu, Y.; Li, S.-H.; Fjeldså, J.; Lei, F. Interpreting the process behind endemism in China by integrating the phylogeography and ecological niche models of the Stachyridopsis ruficeps. PLoS ONE 2012, 7, e46761. [Google Scholar] [CrossRef]
- Yang, T.; Adams, J.M.; Shi, Y.; He, J.S.; Jing, X.; Chen, L.; Tedersoo, L.; Chu, H. Soil fungal diversity in natural grasslands of the Tibetan Plateau: Associations with plant diversity and productivity. New Phytol. 2017, 215, 756–765. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wang, N.; Yu, L. Soil fungal community composition differs significantly among the Antarctic, Arctic, and Tibetan Plateau. Extremophiles 2020, 24, 821–829. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, Y.; Liu, F.; Liu, X.; Wang, Q. Diversity patterns of wetland angiosperms in the Qinghai-Tibet Plateau, China. Diversity 2022, 14, 777. [Google Scholar] [CrossRef]
- Li, Q.; He, G.; Wen, T.; Zhang, D.; Liu, X. Distribution pattern of soil fungi community diversity in alpine meadow in Qilian Mountains of eastern Qinghai-Tibetan Plateau. Ecol. Indic. 2022, 141, 109054. [Google Scholar] [CrossRef]
- Phurbu, D.; Huang, J.-E.; Song, S.; Ni, Z.; Zhou, X.; Li, S.; Cai, L.; Liu, F. Diversity of culturable fungi in six Tibetan Plateau lakes, with descriptions of eight new taxa. Mycology 2024, 1–20. [Google Scholar] [CrossRef]
- Yang, N.; Li, X.; Liu, D.; Zhang, Y.; Chen, Y.; Wang, B.; Hua, J.; Zhang, J.; Peng, S.; Ge, Z. Diversity patterns and drivers of soil bacterial and fungal communities along elevational gradients in the Southern Himalayas, China. Appl. Soil Ecol. 2022, 178, 104563. [Google Scholar] [CrossRef]
- Han, X.; Liu, D.; Zhang, M.; He, M.; Li, J.; Zhu, X.; Wang, M.; Thongklang, N.; Zhao, R.; Cao, B. Macrofungal diversity and distribution patterns in the primary forests of the shaluli mountains. J. Fungi 2023, 9, 491. [Google Scholar] [CrossRef]
- Wang, K.; Liu, S.-L.; Liu, X.-Z.; Hong, P.; Wei, H.-W.; Wang, Y.; Phurbu, D.; Zhou, L.-W.; Wei, T.-Z. Catalogue of fungi in China 3. New taxa of macrofungi from southern Xizang, China. Mycology 2024, 1–33. [Google Scholar] [CrossRef]
- He, M.-Q.; Cao, B.; Liu, F.; Boekhout, T.; Denchev, T.T.; Schoutteten, N.; Denchev, C.M.; Kemler, M.; Gorjón, S.P.; Begerow, D. Phylogenomics, divergence times and notes of orders in Basidiomycota. Fungal Divers. 2024, 126, 127–406. [Google Scholar] [CrossRef]
- Song, H.-B.; Bau, T. Conocybe section Pilosellae in China: Reconciliation of taxonomy and phylogeny reveals seven new species and a new record. J. Fungi 2023, 9, 924. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Bau, T. New species and new records in the genus Conocybe (Bolbitaceae) from China. Phytotaxa 2018, 357, 261–274. [Google Scholar] [CrossRef]
- Amandeep, K.; Atri, N.; Munruchi, K. Diversity of species of the genus Conocybe (Bolbitiaceae, Agaricales) collected on dung from Punjab, India. Mycosphere 2015, 6, 19–42. [Google Scholar] [CrossRef]
- Tóth, A.; Hausknecht, A.; Krisai-Greilhuber, I.; Papp, T.; Vágvölgyi, C.; Nagy, L.G. Iteratively refined guide trees help improving alignment and phylogenetic inference in the mushroom family Bolbitiaceae. PLoS ONE 2013, 8, e56143. [Google Scholar] [CrossRef]
- Malysheva, E.F. Five new species of Conocybe (Agaricomycetes, Bolbitiaceae) from Russia. Mycol. Prog. 2017, 16, 625–636. [Google Scholar] [CrossRef]
- He, M.Q.; Wang, M.Q.; Chen, Z.H.; Deng, W.Q.; Li, T.H.; Vizzini, A.; Jeewon, R.; Hyde, K.D.; Zhao, R.L. Potential benefits and harms: A review of poisonous mushrooms in the world. Fungal Biol. Rev. 2022, 42, 56–68. [Google Scholar] [CrossRef]
- Pu, C.-J.; Peng, Y.-L.; Li, Z.-H.; He, J.; Huang, R.; Feng, T.; Liu, J.-K. Two highly oxygenated ergosterols from cultures of the basidiomycete Conocybe siliginea. Nat. Prod. Res. 2019, 33, 3037–3043. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Hallen-Adams, H.E.; Walton, J.D. Processing of the phalloidin proprotein by prolyl oligopeptidase from the mushroom Conocybe albipes. J. Biol. Chem. 2009, 284, 18070–18077. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Pu, C.-J.; Wang, M.; Li, Z.-H.; Feng, T.; Zhao, D.-K.; Liu, J.-K. Conosiligins A–D, ring-rearranged tremulane sesquiterpenoids from Conocybe siliginea. J. Nat. Prod. 2020, 83, 2743–2748. [Google Scholar] [CrossRef]
- Erritzoe, D.; Barba, T.; Greenway, K.T.; Murphy, R.; Martell, J.; Giribaldi, B.; Timmermann, C.; Murphy-Beiner, A.; Jones, M.B.; Nutt, D. Effect of psilocybin versus escitalopram on depression symptom severity in patients with moderate-to-severe major depressive disorder: Observational 6-month follow-up of a phase 2, double-blind, randomised, controlled trial. eClinicalMedicine 2024, 76, 102799. [Google Scholar] [CrossRef]
- Griffiths, R.R.; Johnson, M.W.; Carducci, M.A.; Umbricht, A.; Richards, W.A.; Richards, B.D.; Cosimano, M.P.; Klinedinst, M.A. Psilocybin produces substantial and sustained decreases in depression and anxiety in patients with life-threatening cancer: A randomized double-blind trial. J. Psychopharmacol. 2016, 30, 1181–1197. [Google Scholar] [CrossRef]
- Wu, F.; Zhou, L.W.; Yang, Z.L.; Bau, T.; Li, T.H.; Dai, Y.C. Resource diversity of Chinese macrofungi: Edible, medicinal and poisonous species. Fungal Divers. 2019, 98, 1–76. [Google Scholar] [CrossRef]
- Hallen, H.E.; Watling, R.; Adams, G.C. Taxonomy and toxicity of Conocybe lactea and related species. Mycol. Res. 2003, 107, 969–979. [Google Scholar] [CrossRef]
- Matheny, P.B.; Curtis, J.M.; Hofstetter, V.; Aime, M.C.; Moncalvo, J.-M.; Ge, Z.-W.; Yang, Z.-L.; Slot, J.C.; Ammirati, J.F.; Baroni, T.J. Major clades of Agaricales: A multilocus phylogenetic overview. Mycologia 2006, 98, 982–995. [Google Scholar] [CrossRef]
- Hausknecht, A.; Contu, M. Interesting species of Conocybe (Agaricales, Bolbitiaceae) from Gallura (NE Sardinia, Italy). Österr. Z. Pilzk 2007, 16, 157–166. [Google Scholar]
- Watling, R. The genus Conocybe subgenus Pholiotina II. Some European exannulate species and North American annulate species. Persoonia-Mol. Phylogeny Evol. Fungi. 1971, 6, 313–339. [Google Scholar]
- Hausknecht, A. A monograph of the genera Conoybe Fayod, Pholiotina Fayod in Europe. Fungi Eur. 2009, 11, 969. [Google Scholar]
- Prydiuk, M.P. Some rare and interesting Conocybe found in Vyzhnytsia National Nature Park (Ukrainian Carpathians). Mycobiota 2014, 4, 1–24. [Google Scholar] [CrossRef]
- Lu, W.; Suwannarach, N.; Lumyong, S.; Elgorban, A.M.; Dai, D.-Q.; Dutta, A.K.; Han, L.-H.; Tibpromma, S.; Karunarathna, S.C. Molecular phylogeny and morphology reveal two new species of Conocybe (Bolbitiaceae, Agaricales) from southwest China. N. Z. J. Bot. 2024, 62, 501–518. [Google Scholar] [CrossRef]
- Ullah, T.; Ullah, K.; Saba, M.; Shah, F.H. Conocybe karakensis sp. nov.(Bolbitiaceae, Agaricales) from Pakistan. Phytotaxa 2023, 584, 135–148. [Google Scholar] [CrossRef]
- Largent, D. How to Identify Mushrooms to Genus Vol. I. Macroscopic Features; Mad River Press: Eureka, CA, USA, 1986; Volume 166. [Google Scholar]
- Bau, T.; Yan, J.Q. Two new rare species of Candolleomyces with pale spores from China. MycoKeys 2021, 80, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour; Methuen & Co. Ltd.: London, UK, 1967. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Hopple Jr, J.S.; Vilgalys, R. Phylogenetic relationships in the mushroom genus Coprinus and dark-spored allies based on sequence data from the nuclear gene coding for the large ribosomal subunit RNA: Divergent domains, outgroups, and monophyly. Mol. Phylogenetics Evol. 1999, 13, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Örstadius, L.; Ryberg, M.; Larsson, E. Molecular phylogenetics and taxonomy in Psathyrellaceae (Agaricales) with focus on psathyrelloid species: Introduction of three new genera and 18 new species. Mycol. Prog. 2015, 14, 25. [Google Scholar] [CrossRef]
- Li, J.X.; Cao, B.; Phurbu, D.; He, M.Q.; Zhu, X.Y.; Parra, L.A.; Zhao, R.L. The revision of the taxonomic system of Lycoperdaceae. Mycosphere 2024, 15, 4919–5016. [Google Scholar] [CrossRef]
- Mou, G.-F.; Bau, T. Asproinocybaceae fam. nov. (Agaricales, Agaricomycetes) for Accommodating the Genera Asproinocybe and Tricholosporum, and Description of Asproinocybe sinensis and Tricholosporum guangxiense sp. nov. J. Fungi 2021, 7, 1086. [Google Scholar] [CrossRef] [PubMed]
- Han, X.X.; Phurbu, D.; Ma, G.F.; Li, Y.Z.; Mei, Y.J.; Liu, D.M.; Lin, F.C.; Zhao, R.L.; Thongklang, N.; Cao, B. A Taxonomic Study of Candolleomyces Specimens from China Revealed Seven New Species. J. Fungi 2024, 10, 499. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v6: Recent updates to the phylogenetic tree display and annotation tool. Nucleic Acids Res. 2024, 52, gkae268. [Google Scholar] [CrossRef] [PubMed]
- Izhar, A.; Bashir, H.; Khalid, A.N. A new species of Conocybe (Bolbitaceae) from Punjab, Pakistan. Phytotaxa 2019, 402, 251–258. [Google Scholar] [CrossRef]
- Malysheva, E. Conocybe (Bolbitiaceae, Agaricomycetes) in the Russian Far East: New species and new section. Микoлoгия И Фитoпатoлoгия 2012, 46, 232–242. [Google Scholar]
- Malysheva, E.F. Conocybe hausknechtii, a new species of sect. Pilosellae from the Western Caucasus, Russia. Mycotaxon 2013, 121, 159–163. [Google Scholar] [CrossRef]
- Liu, J. Taxonomy and Molecular Phylogeny of Bolbitiaceae in Northeast China; Jilin Agricultural University: Changchun, China, 2018. [Google Scholar]
- Wang, Y.-W.; Tzean, S.-S. Dung-associated, potentially hallucinogenic mushrooms from Taiwan. Taiwania 2015, 60, 160–168. [Google Scholar]
- Siquier, J.; Salom, J. Contributo alla conoscenza dei Generi Conocybe (II) e Pholiotina (II) delle Isole Baleari (Spagna). Conocybe parapilosella sp. nov. Riv. Micol. 2021, 31, 131–155. [Google Scholar]
- Siquier, J.; Salom, J. Contributo alla conescenza del genere Conocybe nelle Isole Baleari (Spagna). I. Riv. Di Micol. 2018, 61, 35–77. [Google Scholar]
- Hausknecht, A.; Broussal, M. Conocybe volvicystidiata, a new species of the section Singerella. Osterr. Z. Für Pilzkd. 2016, 25, 191–199. [Google Scholar]
- Song, H.B.; Bau, T. Resolving the polyphyletic origins of Pholiotina s.l. (Bolbitiaceae, Agaricales) based on Chinese materials and reliable foreign sequences. Mycosphere 2024, 15, 1595–1674. [Google Scholar] [CrossRef]
- Peintner, U.; Bougher, N.L.; Castellano, M.A.; Moncalvo, J.M.; Moser, M.M.; Trappe, J.M.; Vilgalys, R. Multiple origins of sequestrate fungi related to Cortinarius (Cortinariaceae). Am. J. Bot. 2001, 88, 2168–2179. [Google Scholar] [CrossRef]
- Hausknecht, A.; Krisai-Greilhuber, I. Pholiotina pleurocystidiata (Bolbitiaceae), eine neue Art mit Pleuro-zystiden Pholiotina pleurocystidiata (Bolbitiaceae), a new species with pleuro-cystidia. Osterr. Z. Für Pilzkd. 2020, 28, 13–18. [Google Scholar]
- Yan, J.Q.; Bau, T. The Northeast Chinese species of Psathyrella (Agaricales, Psathyrellaceae). MycoKeys 2018, 33, 85–102. [Google Scholar] [CrossRef] [PubMed]
- Bau, T.; Yan, J.-Q. A new genus and four new species in the/Psathyrella sl clade from China. MycoKeys 2021, 80, 115. [Google Scholar] [CrossRef] [PubMed]
- Hausknecht, A.; Krisai-Greilhuber, I.; Voglmayr, H. Type studies in north American species of Bolbitiaceae belonging to the genera Conocybe and Pholiotina. Osterr. Z. Für Pilzkd. 2004, 13, 153–235. [Google Scholar]
- Hausknecht, A.; Kalamees, K.; Knudsen, H.; Mukhin, V. The genera Conocybe and Pholiotina (agaricomycotina, Bolbitiaceae) in temperate Asia. Folia Cryptogam. Est. 2009, 45, 23–47. [Google Scholar]
- Karun, N.; Sridhar, K. Elephant dung-inhabiting macrofungi in the Western Ghats. Curr. Res. Environ. Appl. Mycol. 2015, 5, 60–69. [Google Scholar] [CrossRef]
- Hausknecht, A. Beitrage zur kenntnis der Bolbitiaceae 3. Europaische Conocybe-arten mit wurzelndem oder tief im substrat eingesenktem stiel. Osterr Z Pilzk. 1996, 5, 161–202. [Google Scholar]
- Thomas, K.A.; Hausknecht, A.; Manimohan, P. Bolbitiaceae of Kerala State, India: New species and new and noteworthy records. Osterr. Z. Für Pilzkd. 2001, 10, 87–114. [Google Scholar]
- Prydiuk, M.P. New records of Conocybe species from Ukraine. II. The section Conocybe. Czech Mycol. 2007, 59, 39. [Google Scholar] [CrossRef]
- Sakamoto, Y. Influences of environmental factors on fruiting body induction, development and maturation in mushroom-forming fungi. Fungal Biol. Rev. 2018, 32, 236–248. [Google Scholar] [CrossRef]
- Krah, F.S.; Hagge, J.; Schreiber, J.; Brandl, R.; Müller, J.; Bässler, C. Fungal fruit body assemblages are tougher in harsh microclimates. Sci. Rep. 2022, 12, 1633. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.Y.; Miao, L.Z.; Feng, X.K.; Zhan, X.X. Snow Disaster Risk Assessment Based on Long-Term Remote Sensing Data: A Case Study of the Qinghai-Tibet Plateau Region in Xizang. Remote Sens. 2024, 16, 1661. [Google Scholar] [CrossRef]
- Kong, H.J.; Wang, J.G.; Cai, L.; Cao, J.X.; Zhou, M.; Fan, Y.D. Surface Solar Radiation Resource Evaluation of Xizang Region Based on Station Observation and High-Resolution Satellite Dataset. Remote Sens. 2024, 16, 1405. [Google Scholar] [CrossRef]
- Mao, X.L. Economic Fungi of China; Science and Technology Press: Beijing, China, 1998; pp. 1–762. [Google Scholar]
- Bau, T.; Li, Y. Study on fungal flora diversity in Daqinggou Nature Reserve. Biodivers. Sci. 2000, 8, 73–80. [Google Scholar] [CrossRef]
- Mao, X.L. Alpine macrofungi of East Himalaya and their adaptive characteristics. Mt Res. 1985, 3, 299–307. [Google Scholar]
- Mao, X.L. The resources of macrofungi from the Mt. Namjagbarwa region in Xizang (Tibet), China. Acta Mycol. Sin. 1985, 4, 197–207. [Google Scholar]
- Mao, X.L. The Macrofungi in China; Henan Science and Technology Press: Zhengzhou, China, 2000; Volume 118. [Google Scholar]
- Yuan, M.S.; Sun, P.Q. Mushrooms of Sichuan; Sichuan Science and Technology Press: Chengdu, China, 1995; pp. 1–737. [Google Scholar]
- Bau, T.; Li, Y. Notes on Fungi in Tibet of China. Bull. Bot. Res. 2001, 2, 191–194. [Google Scholar]
- Yuan, M.; Sun, P. The Pictorial Book of Mushrooms of China; Sichuan Science and Technology Press: Chengdu, China, 2007; pp. 1–552. [Google Scholar]
- Zhuang, W.Y. Fungi of Northwestern China; Mycotaxon: Ithaca, NY, USA, 2005; pp. 1–430. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).