
Phylogenetic and Morphological Analyses Reveal Twelve New Species of the Genus Patellaria (Dothideomycetes, Ascomycota) from Mexico

 , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
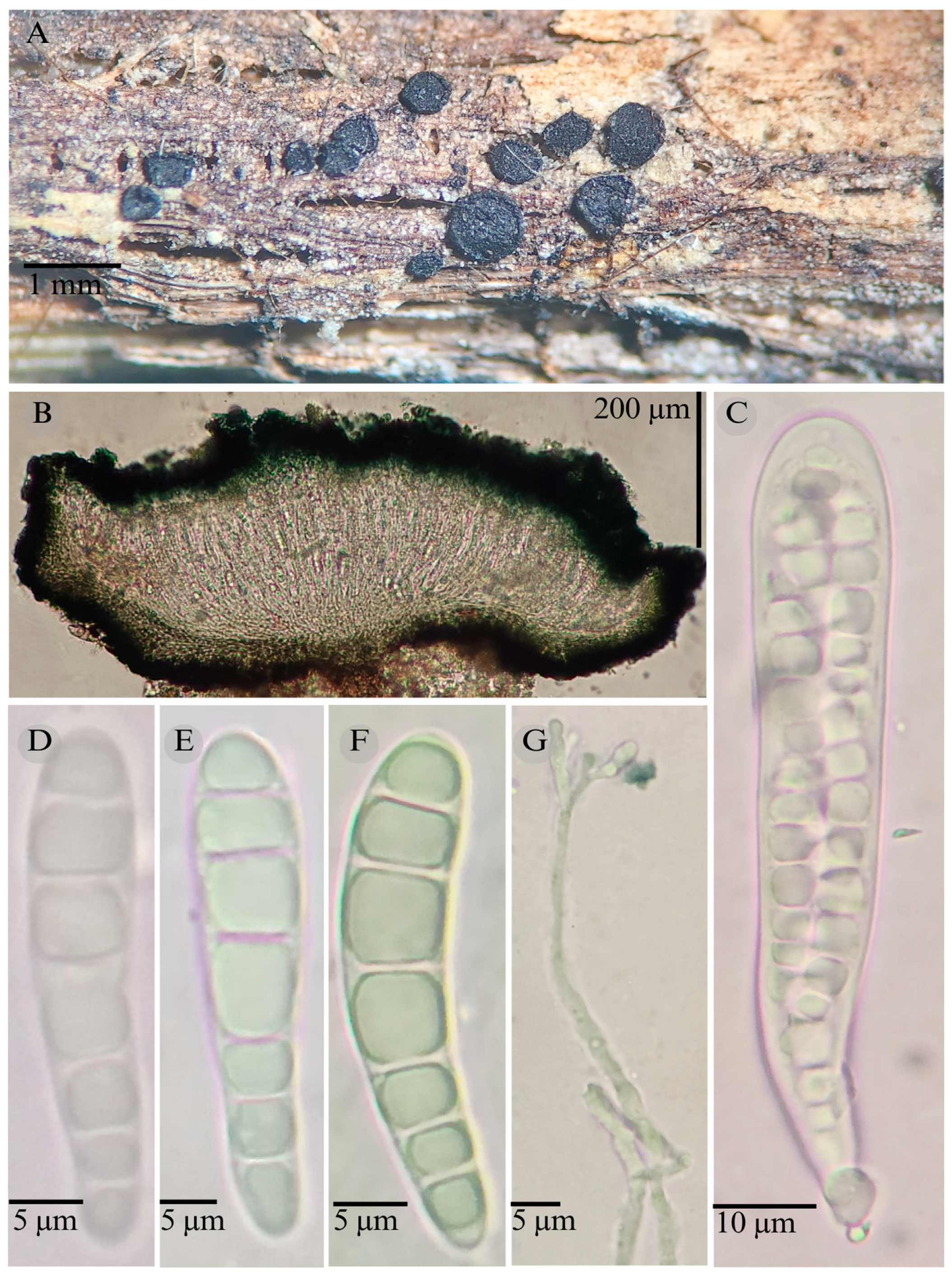
2.1. Morphological Studies
2.2. Amplification and Sequencing
2.3. Phylogenetic Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Loci/Segment | Primer | Sequence 5′-3′ | T (°C) | Reference |
|---|---|---|---|---|
| ITS | ITS5 | GGAAGTAAAAGTCGTAACAAGG | 58 | [33] |
| ITS4 | TCCTCCGCTTATTGATATGC | 58 | [33] | |
| nLSU | LROR | ACCCGCTGAACTTAAGC | 48 | [33] |
| LR3 | GGTCCGTGTTTCAAGAC | 48 | [33] | |
| 0mtSSU | MS1 | CAGCAGTCAAGAATATTAGTCAATG | 63 | [33] |
| MS2 | GCGGATTATCGAATTAAATAAC | 53 | [33] |
3. Results
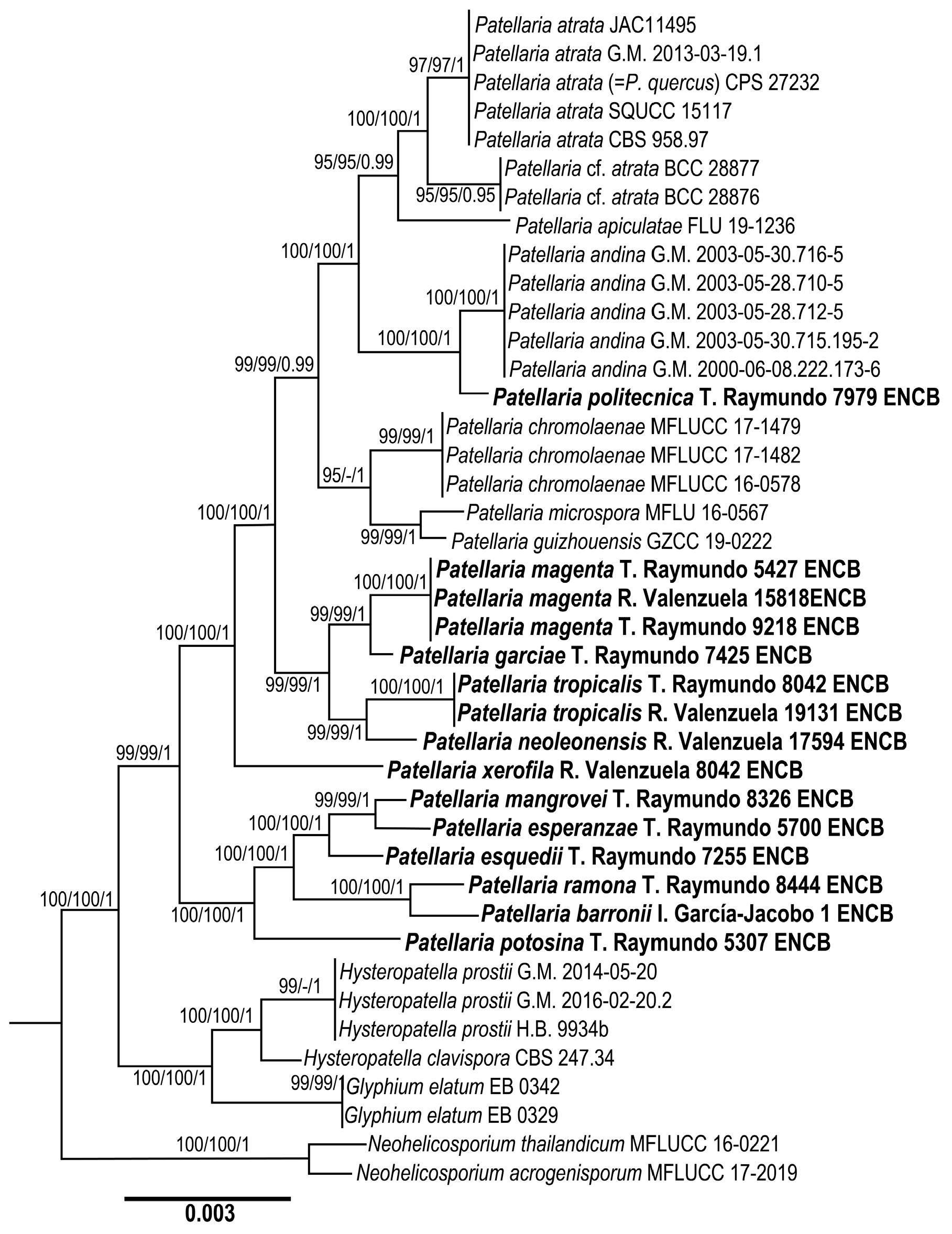
3.1. Phylogenetic Analysis
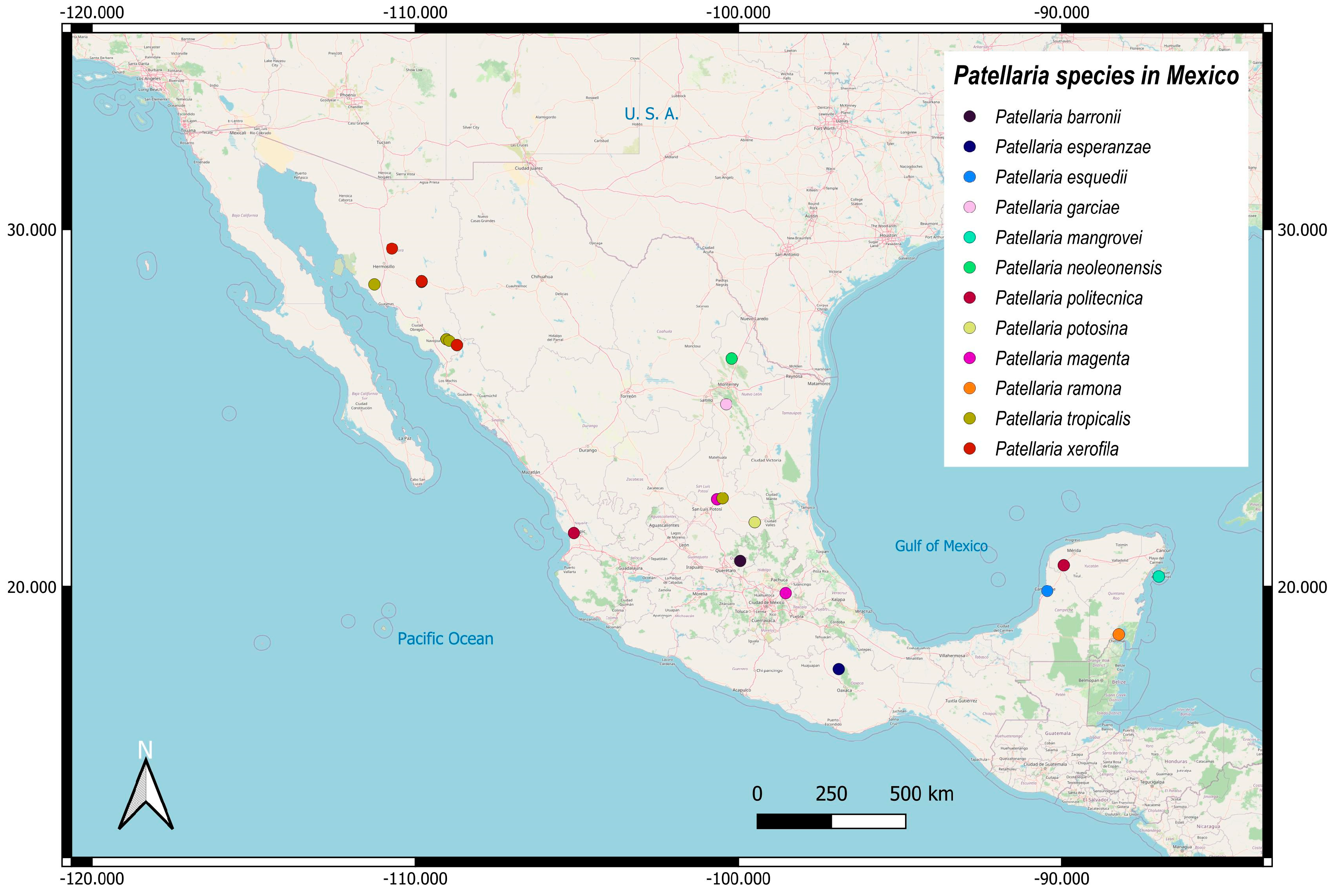
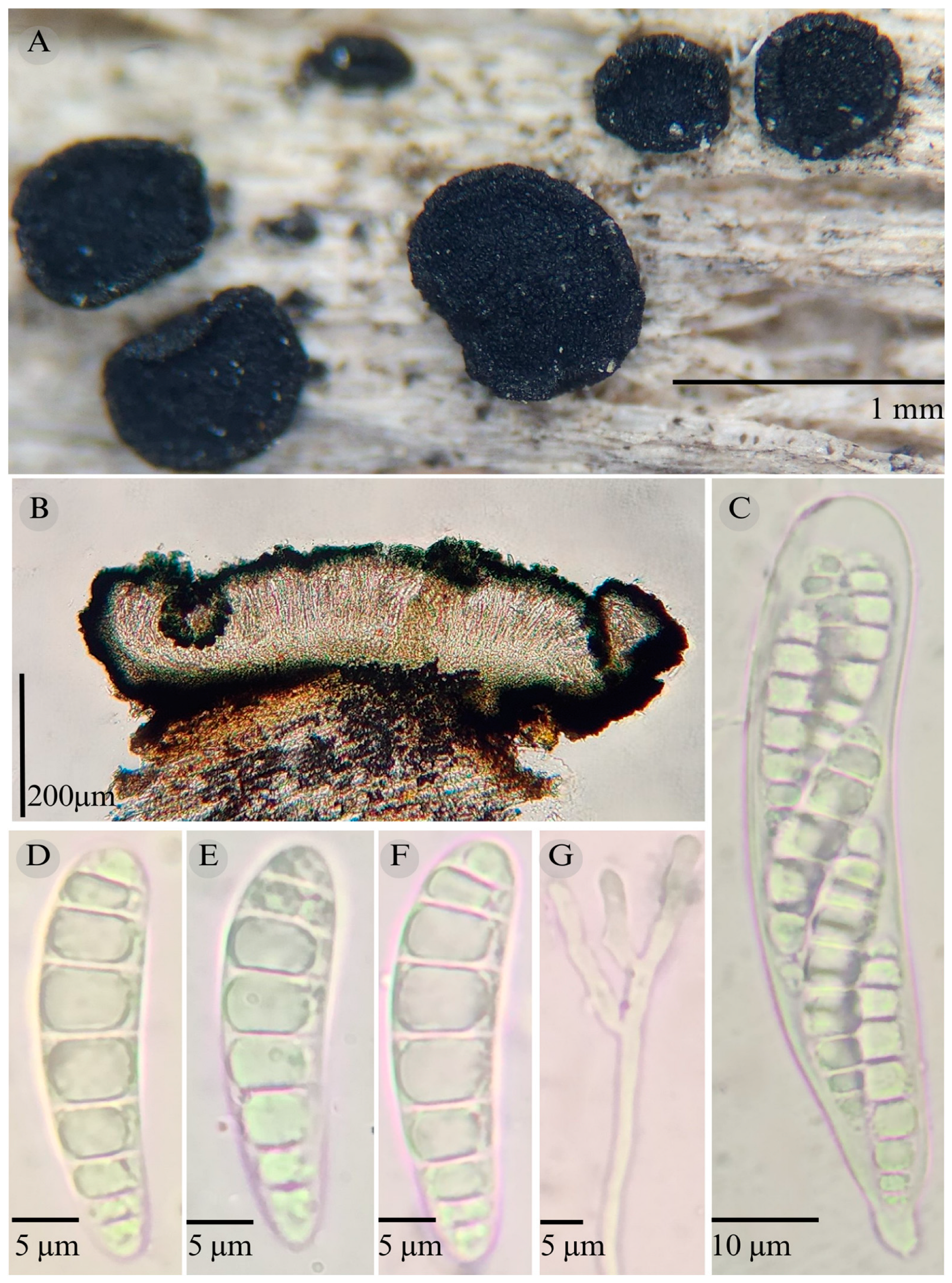
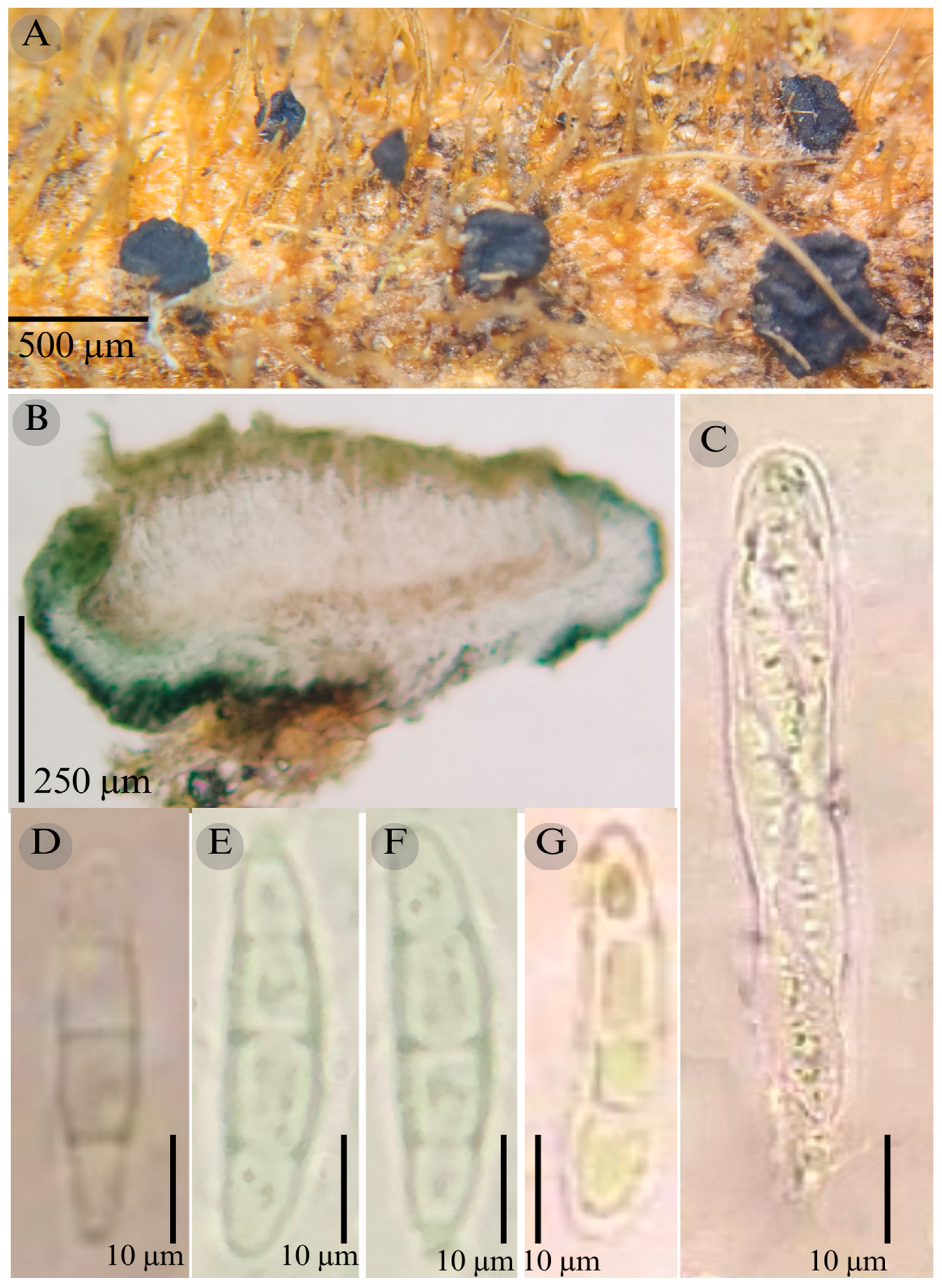
3.2. Taxonomy
- Key to Patellaria species forming ascomata supported by molecular and morphological data
- 1.
- Species with allantoid ascospores and 3–6 septa ……………………….…………………………………………………………2
- 1.
- Species with subfusiform, obclavate, or clavate ascospores; 3–13 septa……………………………………………………………………………3
- 2.
- Ascomata < 700 µm in diam; ascospores 27–34 × 5.2–6 µm, allantoid, 3–7 septa; growing in dead stems of Chromolaenae odorata from Thailand …………………………………………………………………P. chromolaenae
- 2.
- Ascomata > 700 µm in diam; ascospores 21–32 × 4–5 µm, allantoid, 3–6 septa; growing in dead wood of Mexico ……………………………………………………………………P. politecnica
- 3.
- Ascospores < 20 µm long, subfusiform or claviform …………………………………………………………………………………4
- 3.
- Ascospores > 20 µm long, obclavate or claviform …………………………………………………………………………………5
- 4.
- Subfusiform ascospores with three septa growing in the stem of the genus Cyathea ……………………………………………………………………P. esperanzae
- 4.
- Ellipsoid ascospores with eight septa growing in dead wood ……………………………………………………………………P. microspora
- 5.
- Obclavate ascospores, 5–6 septa …………………………………………………………………………………6
- 5.
- Clavate ascospores, 4–13 septa …………………………………………………………………………………7
- 6.
- Discoid ascomata, smooth and thick rim, green jungle coloration in KOH (25F8); unilobed pedicellate asci, ascospores 30–32 × 5–7 µm ( = 31 × 6, n = 30); growing in tropical rain forest ………………………………………………………………………P. esquedii
- 6.
- Discoid ascomata, striate and thin rim, coloration nautical blue in KOH (24E7); bilobed pedicel asci, ascospores 30–33 × 6–7 µm ( = 32 × 6, n = 30); growing in Quercus forest “Chaparral” ………………………………………………………………………P. potosina
- 7.
- Clavate ascospores, slightly curved, 4–8 septa, unilobed asci …………………………………………………………………………………8
- 7.
- Clavate ascospores, 8–10 (13) septa, bilobed asci …………………………………………………………………………………15
- 8.
- Ascospores < 30 µm long …………………………………………………………………………………9
- 8.
- Ascospores > 30 µm long …………………………………………………13
- 9.
- Species growing in temperate forests …………………………………………………………………………P. atrata
- 9.
- Species growing in tropical forests or mangroves…………………………………………………………………10
- 10.
- Species growing in mangroves ………………………………………………………………………………11
- 10.
- Species growing in tropical forests; ascospores 26–30 µ ………………………………………………………………………………12
- 11.
- Ascomata with thick rim and turquoise pigmentation in KOH (24F8), ascospores = 26 × 7, 5–6 septa; growing in Rhizophora apiculata ……………………………………………………………………P. apiculate
- 11.
- Ascomata with a thin rim and olive brown pigmentation in KOH (4E8), ascospores = 29 × 7, n = 30, 5–6 septa; growing in Rhizophora mangle ……………………………………………………………………P. mangrovei
- 12.
- Ascomata with thick rim and turquoise pigmentation in KOH (24F8), asci, short pedicellate, ascospores = 28 × 6, n = 30, 4–6 septa; growing in tropical rain forests………………………………………………………………P. ramona
- 12.
- Ascomata with a thin rim with deep green pigmentation in KOH (25D8), asci with long pedicellate, ascospores = 28 × 6, n = 30, 5–7 septa; growing in tropical dry forests ………………………………………………………………P. tropicalis
- 13.
- Ascospores with 5–6 septa ……………………………………………………………P. ghizhuensis
- 13.
- Ascospores with six or more septa……………………………………………………………………14
- 14.
- Ascomata with a thin rim, green jungle pigmentation in KOH (25F8), asci with short pedicellate, ascospores 31–35 × 6–8 µm, = 33 × 7 µm, 6–8 septa; …………………………………………………………P. barronii
- 14.
- Ascomata with thick rim turquoise pigmentation in KOH (24F8), asci long with pedicellate, ascospores 28–30 × 6–7 µm, = 30 × 6 µm, 5–7 septa…………………………………………………………………P. xerofila
- 15.
- Species growing in temperate coniferous forest, ascomata with thin and smooth rims, green jungle pigmentation in KOH(CLA), ascospores with clavate blunt apex ……………………………………………………………………P. garciae
- 15.
- Species in tropical dry forest, ascomata with thick and striate rim, English green pigmentation in KOH, ascospores clavate with conic apex……………………………………………………………………16
- 16.
- Ascospores 32–40 × 6–7 µm, = 36 × 6.2 µm, n = 30, 8–10 (13) septa, asci with long pedicellate, ……………………………………………………………………P. purpurea
- 16.
- Ascospores 38–44 × 6–7 µm, = 40 × 6.6 µm, n = 30, 8–10 septa, asci with short pedicellate………………………………………P. neolonensis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fries, E.M. Systema Mycologicum; Gryphiswaldiae: Sumtibus Emesti, Mauritius, 1822; Volume 2, p. 275. [Google Scholar] [CrossRef]
- Species Fungorum. Available online: http://www.speciesfungorum.org/Index.htm (accessed on 30 September 2024).
- Kutorga, E.; Hawksworth, D.L. A re-assessment of the genera referred to the family Patellariaceae (Ascomcota). Syst. Ascomycetum 1997, 15, 1–110. [Google Scholar]
- Dayarathne, M.C.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Devadatha, B.; Sarma, V.V.; Khongphinitbunjong, K.; Chonunti, P.; Hyde, K.D. Morpho-molecular characterization of microfungi associated with marine based habitats. Mycosphere 2020, 11, 1–188. [Google Scholar] [CrossRef]
- Yacharoen, S.; Tian, Q.; Chommunti, P.; Boonmee, S.; Chukeatirote, E.; Bhat, J.D.; Hyde, K.D. Patellariaceae revisited. Mycosphere 2015, 6, 290–326. [Google Scholar] [CrossRef]
- Hernandez-Restrepo, M.; Schumacher, R.K.; Wingfield, M.J.; Ishtiaq, A.; Cai, L.; Duong, T.A.; Edwards, J.; Gene, J.; Groenewald, J.Z.; Sana, J.; et al. Fungal systematics and evolution: FUSE 2. Sydowia 2016, 68, 193–230. [Google Scholar]
- Mapook, A.; Hyde, K.D.; McKenzie, E.H.C.; Jones, E.B.G.; Bhat, D.J.; Jeewon, R.; Stadler, M.; Samarakoon, M.C.; Malaithong, M.; Tanunchai, B.; et al. Taxonomic and phylogenetic contributions to fungi associated with the invasive weed Chromolaena odorata (Siam weed). Fungal Divers. 2020, 101, 1–175. [Google Scholar] [CrossRef]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Lücking, R.; Boonmee, S.; Bhat, J.D.; Liu, N.G.; et al. Refined families of Dothideomycetes: Orders and families incertae sedis in Dothideomycetes. Fungal Divers. 2020, 105, 17–318. [Google Scholar] [CrossRef]
- Zhang, J.F.; Liu, J.K.; Hyde, K.D.; Chen, Y.; Ran, H.; Liu, Z. Ascomycetes from karst landscapes of Guizhou Province, China. Fungal Divers. 2023, 122, 1–160. [Google Scholar] [CrossRef]
- Chacón, S.; Tapia, F. Algunas especies saprobias de dothideomycetes y lecanoromycetes (Pezizomycotina: Ascomycota) en México. Rev. Mex. Biodivers. 2016, 87, 1169–1176. [Google Scholar] [CrossRef]
- Méndez-Mayboca, F.; Checa, J.; Esqueda, M.; Chacón, S. New records of Loculoascomycetes from natural protected areas in Sonora, Mexico. Mycotaxon 2010, 111, 19–30. [Google Scholar] [CrossRef]
- Esqueda, M.; Coronado, M.L.; Gutiérrez, A.; Lizárraga, M.; Raymundo, T.; Valenzuela, R. Hongos de la Reserva de la Biósfera El Pinacate y Gran desierto de Altar, 1st ed.; Centro de Investigación en Alimentación y Desarrollo (CIAD): Hermosillo, Mexico, 2013; p. 23. [Google Scholar]
- García-Martínez, Y.; Heredia Abarca, G.; Guzmán-Guillermo, J.; Valenzuela, R.; Raymundo, T. Hongos asociados al mangle rojo Rhizophora mangle (Rhizophoraceae) en la Reserva de la Biosfera Isla Cozumel, Quintana Roo, México. Acta Bot. Mex. 2021, 128. [Google Scholar] [CrossRef]
- Raymundo, T.; Martínez-Pineda, M.; Reyes, P.H.; Cobos-Villagrán, A.; Martínez, Y.A.G.; Tun, A.A.; Valenzuela, R. Ascomicetos de la Reserva de la Biosfera Isla Cozumel, México. Act. Bot. Mex. 2021, 128. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Methuen: London, UK, 1978; pp. 1–254. [Google Scholar]
- Martínez-González, C.R.; Ramírez-Mendoza, R.; Jiménez-Ramírez, J.; Gallegos-Vázquez, C.; Luna-Vega, I. Improved method for genomic DNA extraction for Opuntia Mill. (Cactaceae). Plant Methods 2017, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Wanasinghe, D.N.; Cheewangkoon, R.; Al-Sadi, A.M. Uncovering the hidden taxonomic diversity of fungi in Oman. Fungal Divers. 2021, 106, 229–268. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Muller, K.; Quandt, D.; Muller, J.; Neinhuis, C. PhyDER-Phylogenetic Data Editor. Program Distributed by the Authors, Ver. 10.0. 2005. Available online: https://www.phyde.de (accessed on 3 September 2024).
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.31. 2017. Available online: http://mesquiteproject.org (accessed on 3 September 2024).
- Swofford, D.L. Phylogenetic Analysis Using Parsimony (PAUP*) 4.0; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MrBayes: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Frandsen, P.B.; Calcott, B.; Mayer, C.; Lanfear, R. Automatic selection of partitioning schemes for phylogenetic analyses using iterative k-means clustering of site rates. BMC Evol. Biol. 2015, 15, 13. [Google Scholar] [CrossRef]
- Lanfear, R.; Calcott, B.; Kainer, D.; Mayer, C.; Stamatakis, A. Selecting optimal partitioning schemes for phylogenomic datasets. BMC Evol. Biol. 2014, 14, 82. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2016, 34, 772–773. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree v.1.4.2. A Graphical Viewer of Phylogenetic Trees. 2014. Available online: https://tree.bio.ed.ac.uk/software/figtree (accessed on 3 September 2024).
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 135–322. [Google Scholar] [CrossRef]
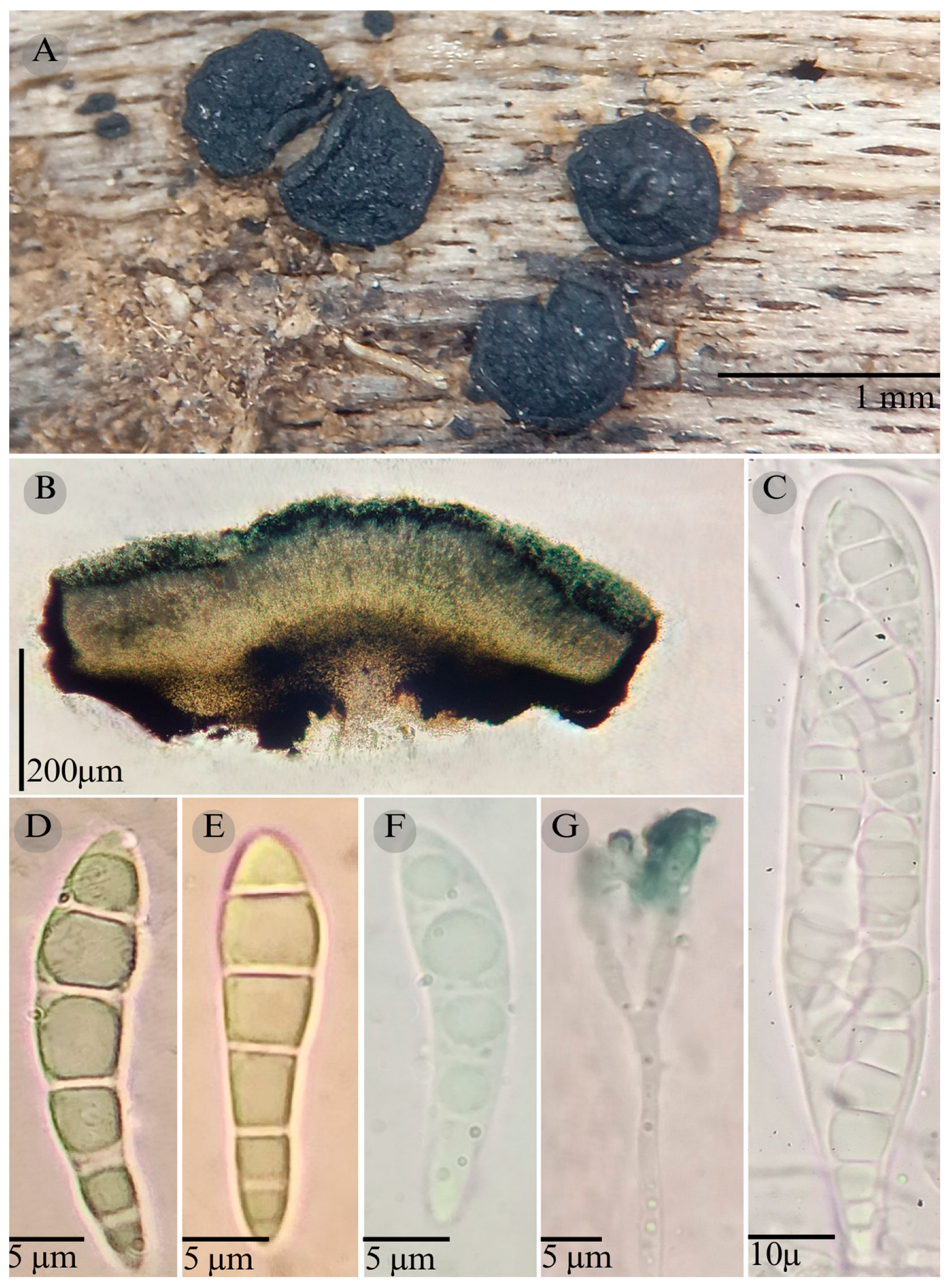
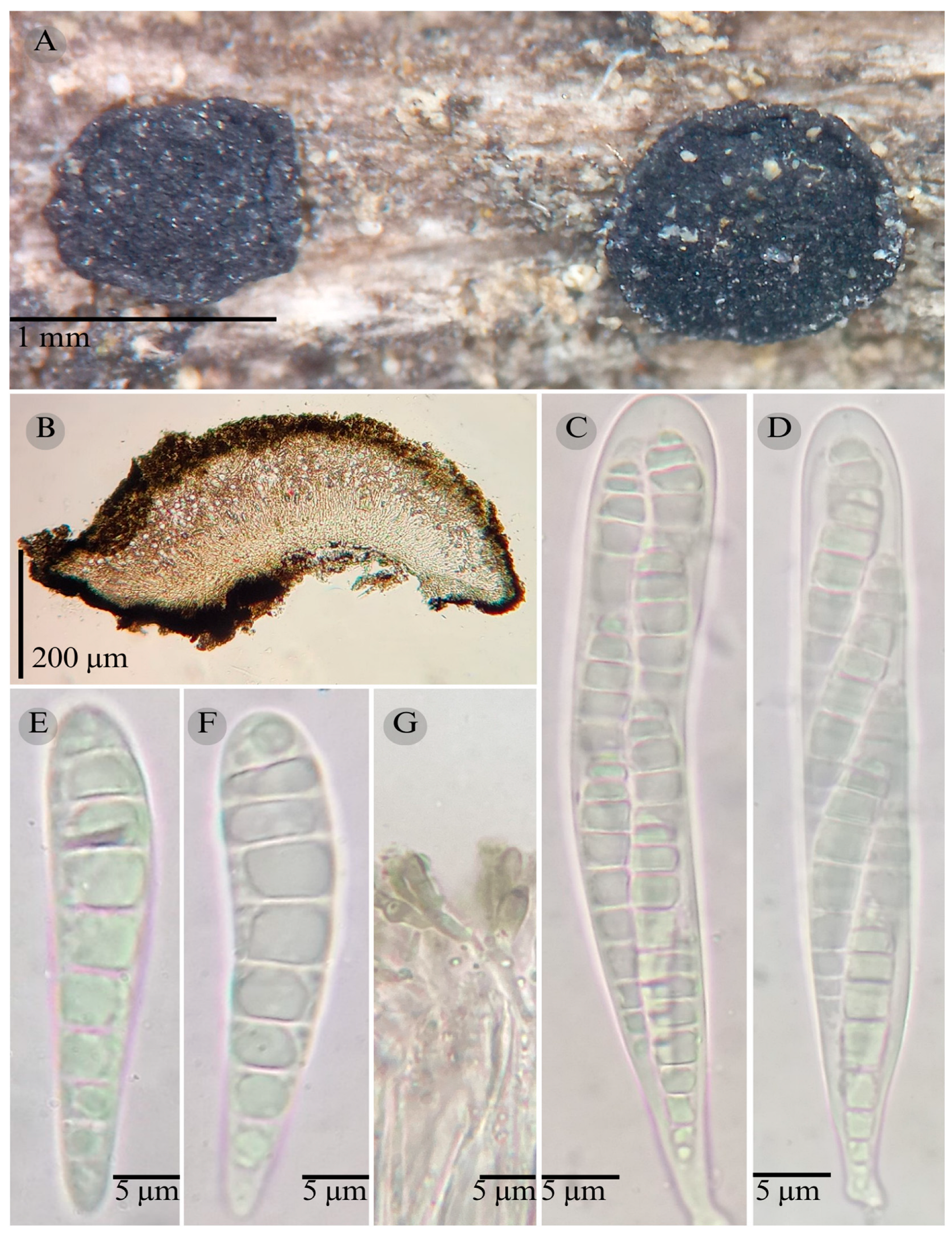
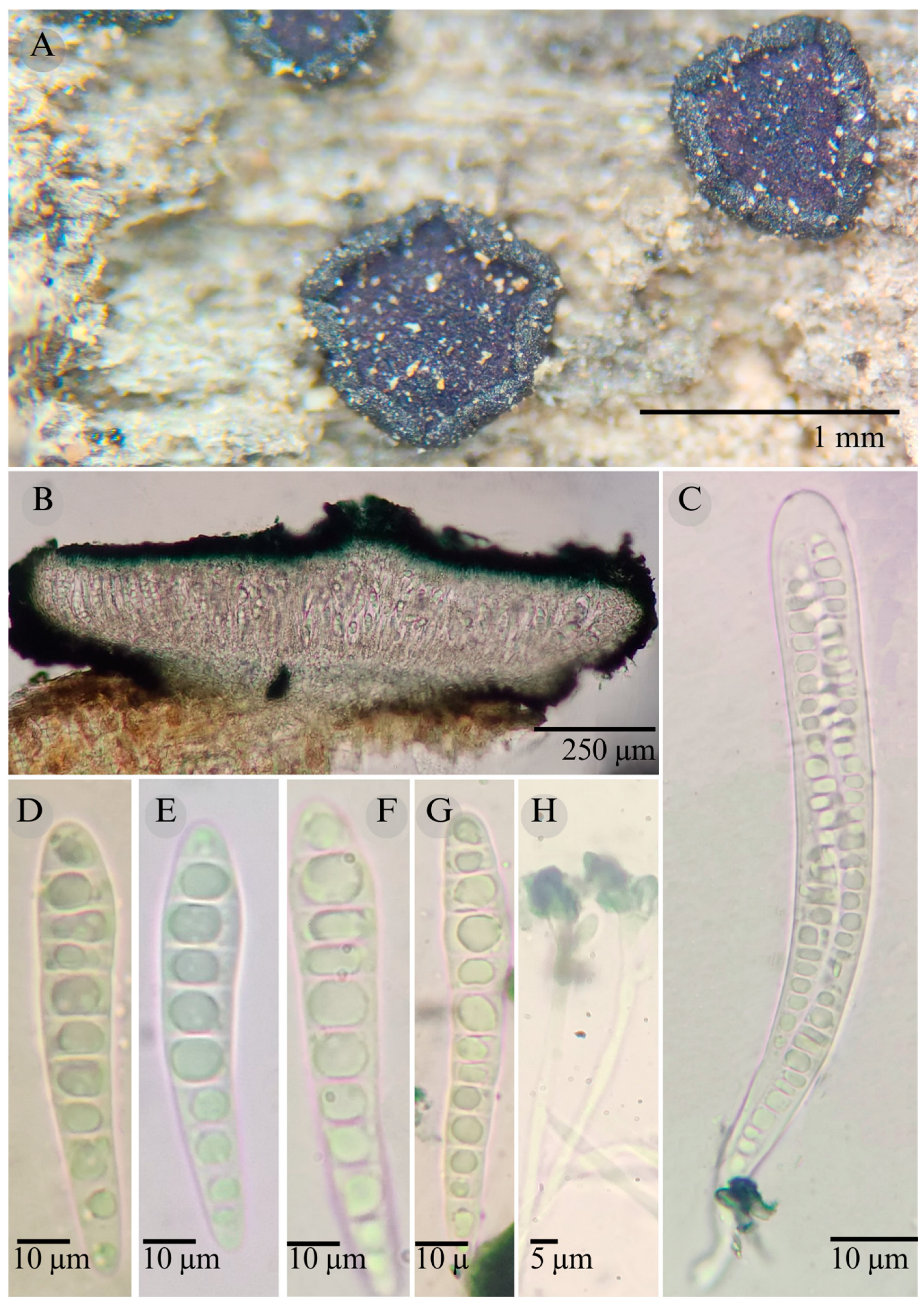
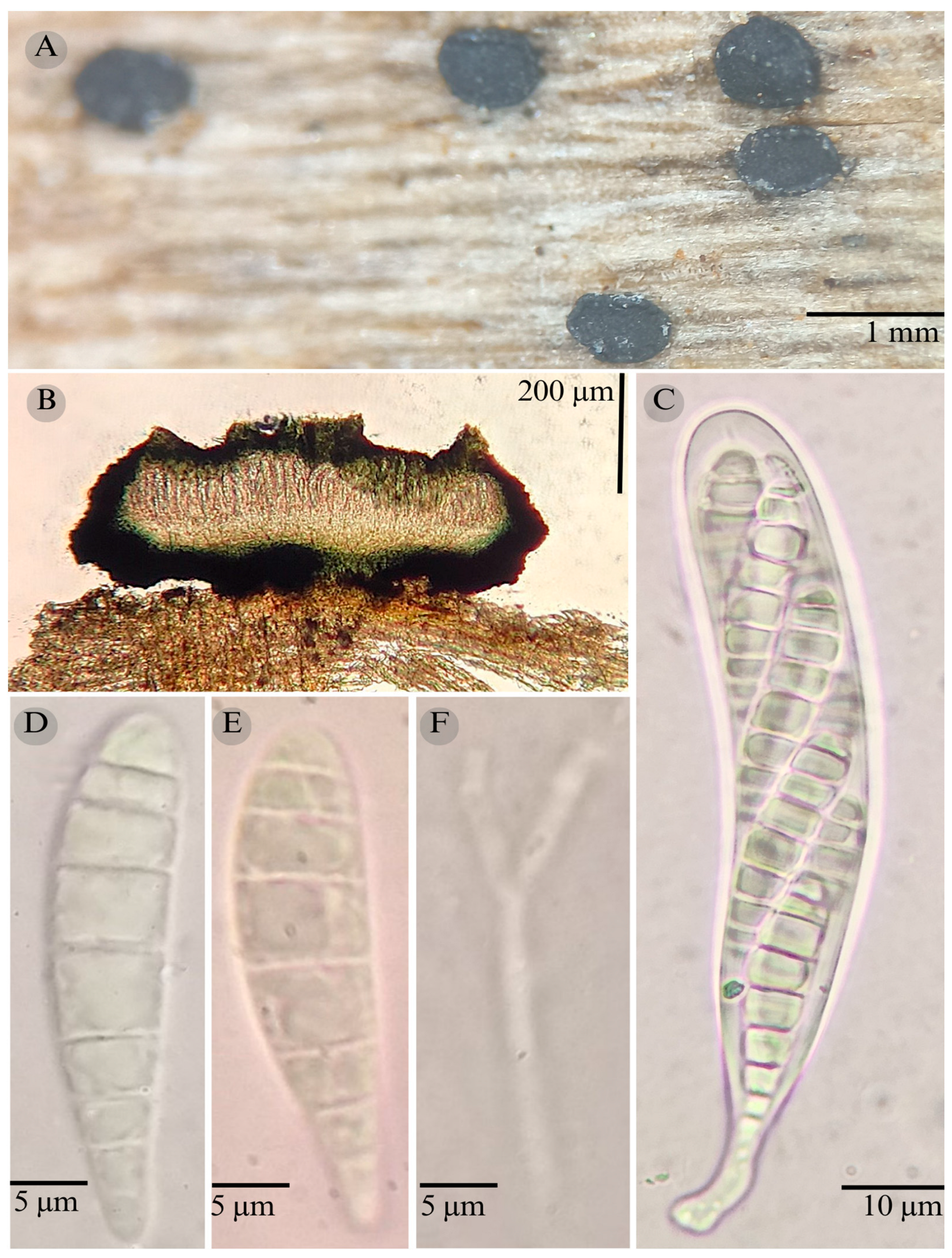

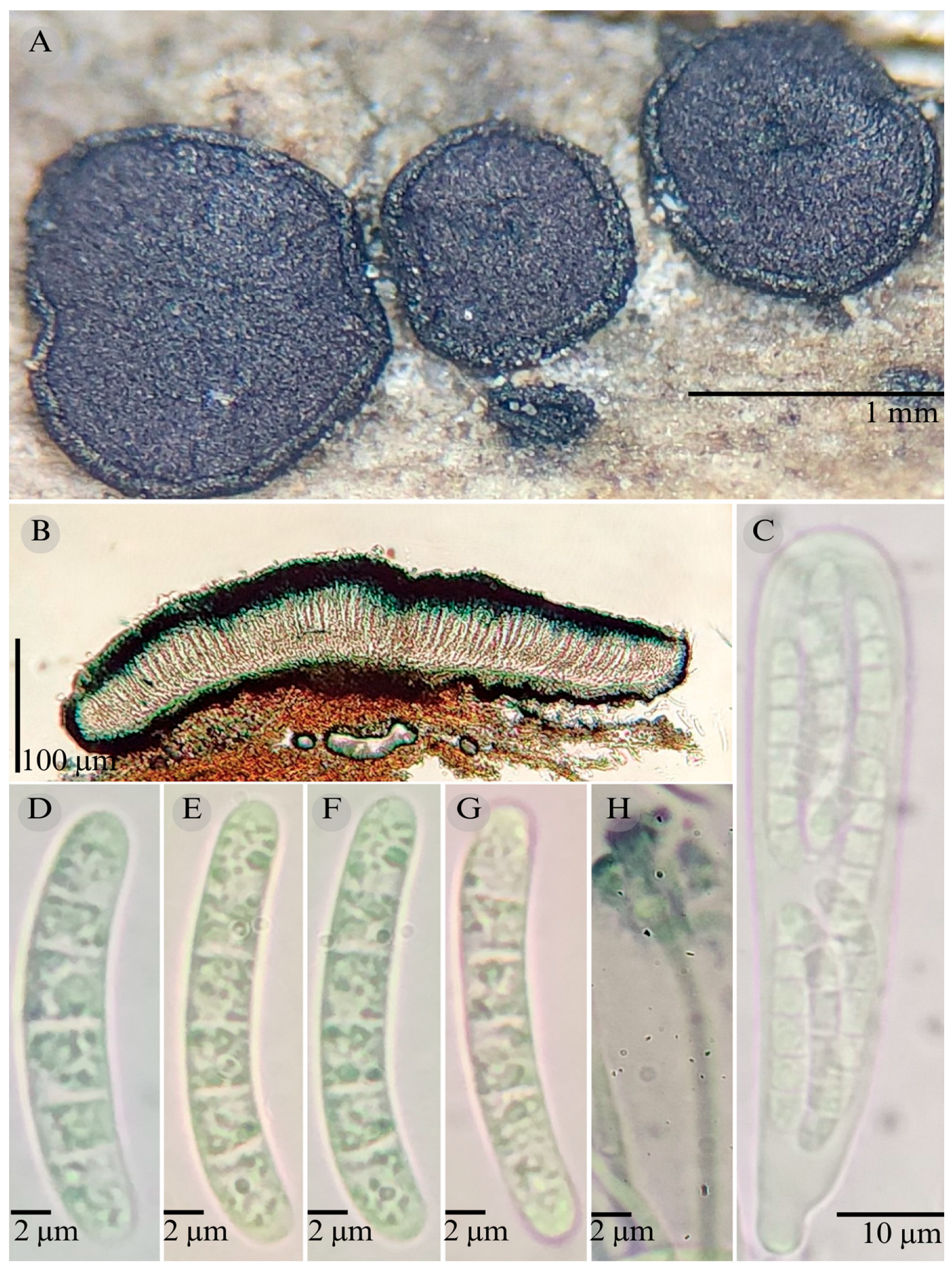
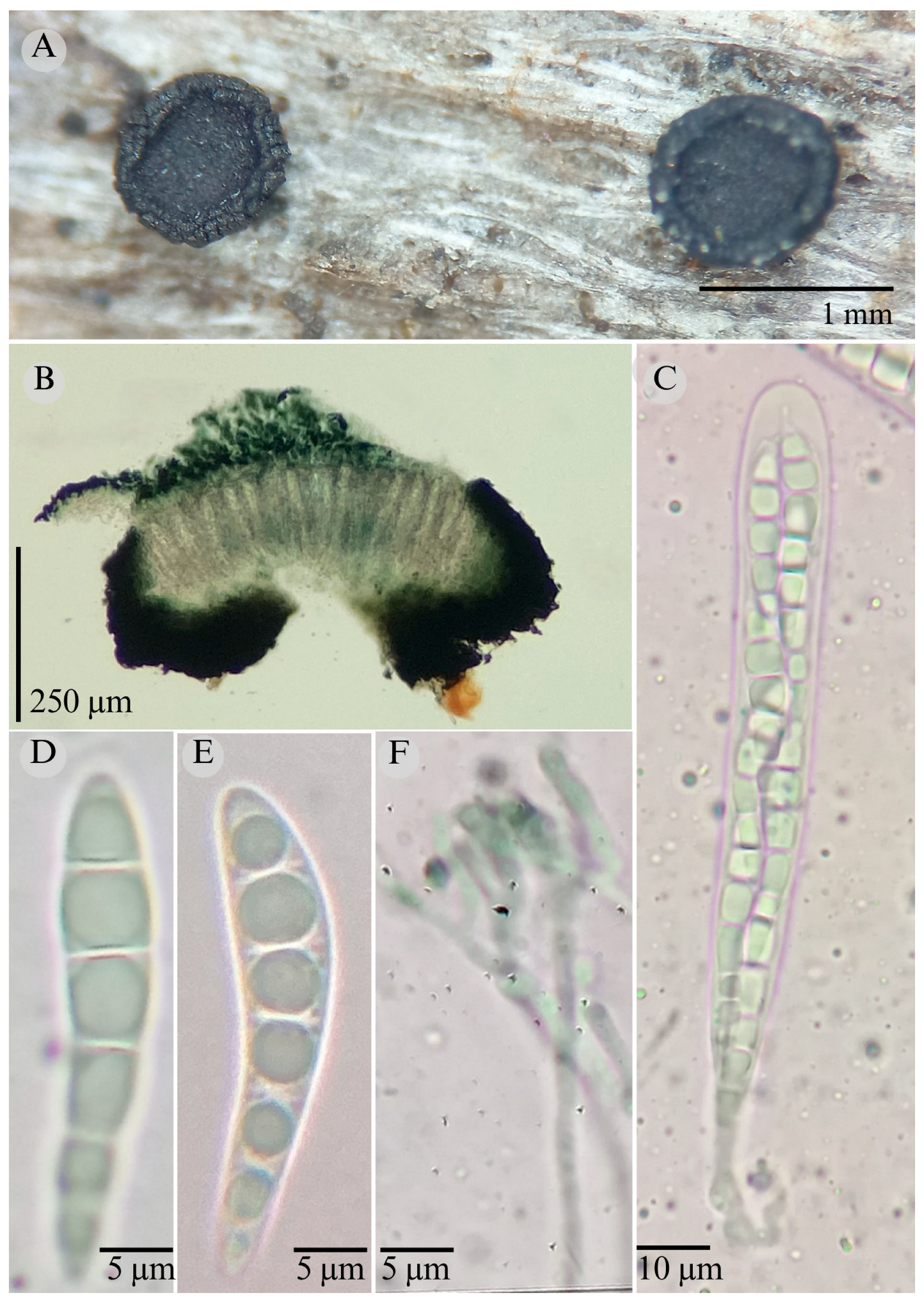
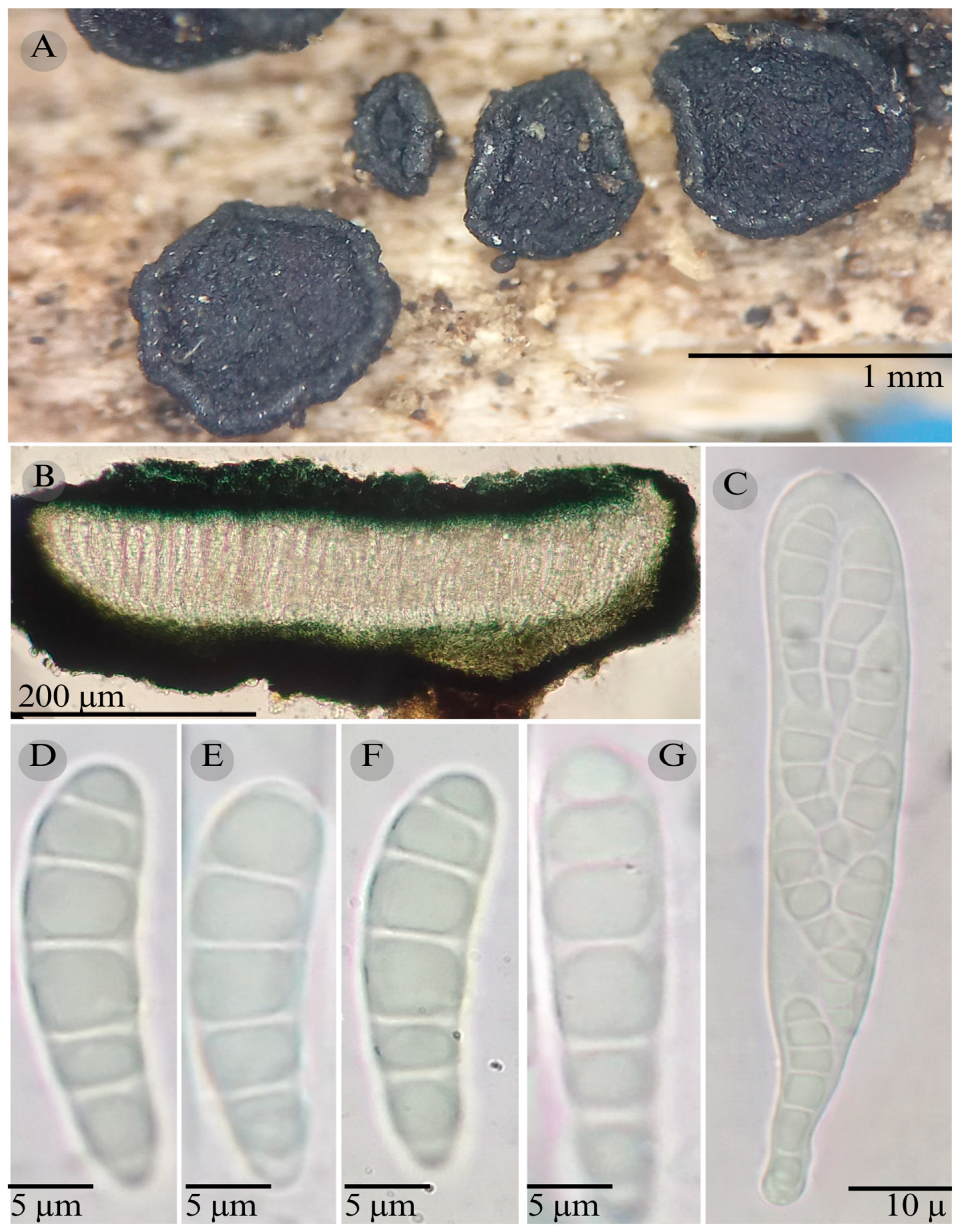
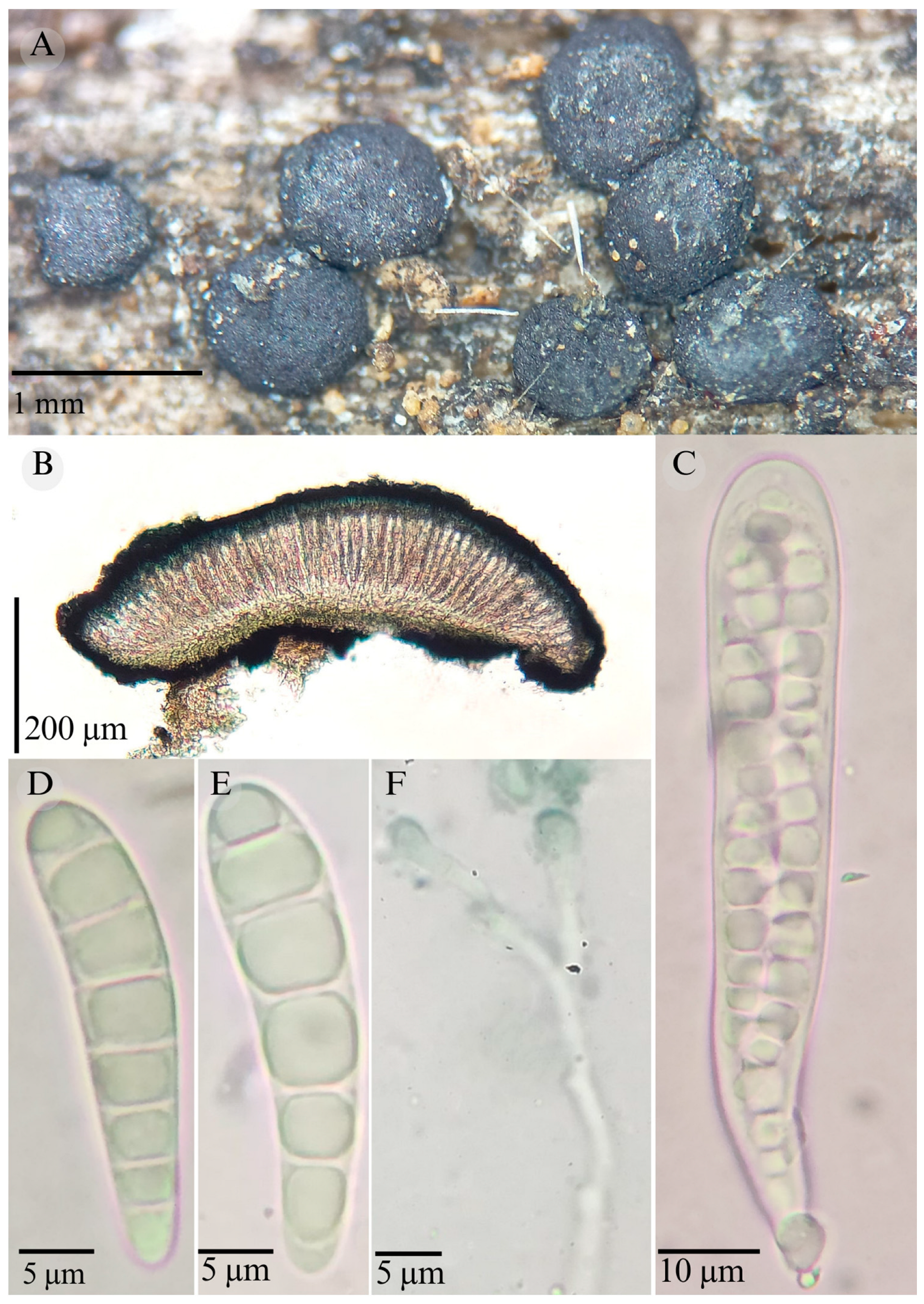
| Species | Strain No. | GenBank Accession No. | ||
|---|---|---|---|---|
| nLSU | ITS | SSU | ||
| Glyphium elatum | EB 0329 | KM220937 | KM220943 | KM220951 |
| Glyphium elatum | EB 0342 | KM220938 | KM220944 | KM220952 |
| Glyphium elatum | EB 0388 | KM220940 | KM220946 | ----- |
| Hysteropatella clavispora | CBS 247.34 | AY541493 | ----- | ----- |
| Hysteropatella prostii | H.B. 9934b | KT876980 | KT876980 | ----- |
| Hysteropatella prostii | G.M. 2014-05-20 | KM220949 | KM220949 | ----- |
| Hysteropatella prostii | G.M. 2016-02-20.2 | MT341324 | MT341324 | MT341324 |
| Neohelicosporium acrogenisporum | MFLUCC 17-2019 | MH558871 | NR160376 | ----- |
| Neohelicosporium thailandicum | MFLUCC 16-0221 | MF467941 | MF467928 | MF467928 |
| Patellaria andina | G.M. 2000-06-08.222.173-6 | MT273251 | ----- | ----- |
| Patellaria andina | G.M. 2003-05-30.716-5 | MT293616 | MT293616 | ----- |
| Patellaria andina | G.M. 2003-05-30.715.195-2 | MT273250 | MT273250 | ----- |
| Patellaria andina | G.M. 2003-05-28.712-5 | MT293619 | MT293619 | ----- |
| Patellaria andina | G.M. 2003-05-28.710-5 | MT293624 | MT293624 | ----- |
| Patellaria apiculatae | MFLU 19-1236 | MN017860 | MN047094 | MN017925 |
| Patellaria atrata | JAC11495 | MK431438 | MK432702 | MK432702 |
| Patellaria atrata | CBS 958.97 | GU301855 | ----- | GU296181 |
| Patellaria atrata | G.M. 2013-03-19.1 | MN565888 | MN565888 | ----- |
| Patellaria atrata | SQUCC 15117 | MW077152 | MW077143 | ----- |
| Patellaria barronii | S. García 1 ENCB | PQ766174 | PQ817147 | PQ773270 |
| Patellaria cf. atrata | BCC 28877 | GU371829 | ----- | GU371837 |
| Patellaria cf. atrata | BCC 28876 | GU371828 | ----- | GU371836 |
| Patellaria chromolaenae | MFLUCC 16-0578 | MW142387 | MW136695 | MW127179 |
| Patellaria chromolaenae | MFLUCC 17-1482 | MT214475 | MT214381 | MT214426 |
| Patellaria chromolaenae | MFLUCC 17-1479 | MT214474 | MT214380 | ----- |
| Patellaria esperanzae | T. Raymundo 5700 ENCB | PQ766175 | PQ817148 | PQ773271 |
| Patellaria esquedii | T. Raymundo 7255 ENCB | PQ766176 | PQ817149 | PQ773272 |
| Patellaria garciae | T. Raymundo 7425 ENCB | PQ766177 | PQ817150 | PQ773273 |
| Patellaria guizhouensis | GZCC 19-0222 | OR209665 | ----- | OR134437 |
| Patellaria mangrovei | T. Raymundo 8326 ENCB | PQ766178 | PQ817151 | PQ773274 |
| Patellaria microspora | MFLU 16-0567 | MW142388 | NR175661 | |
| Patellaria neoleonensis | R. Valenzuela 17594 ENCB | PQ766179 | PQ817152 | PQ773275 |
| Patellaria politecnica | T. Raymundo 7979 ENCB | PQ766180 | PQ817153 | PQ773276 |
| Patellaria potosina | T. Raymundo 5307 ENCB | PQ766181 | PQ817154 | PQ773277 |
| Patellaria magenta | T. Raymundo 5427 ENCB | PQ766182 | PQ817155 | PQ773278 |
| Patellaria magenta | R. Valenzuela 15818 ENCB | PQ766183 | PQ817156 | PQ773279 |
| Patellaria magenta | T. Raymundo 9218 ENCB | PQ766184 | PQ817157 | PQ773280 |
| Patellaria quercus | CPC 27232 | NG059696 | NR152540 | |
| Patellaria ramona | T. Raymundo 8444 ENCB | PQ766185 | PQ817158 | PQ773281 |
| Patellaria tropicalis | T. Raymundo 8042 ENCB | PQ766186 | PQ817159 | PQ773282 |
| Patellaria tropicalis | R. Valenzuela 19131 ENCB | PQ766187 | PQ817160 | PQ773283 |
| Patellaria xerofila | R. Valenzuela 15286 ENCB | PQ766188 | PQ817161 | PQ773284 |
| Species | Ascomata | Rim | Exciple | Sub Hymenium | Asci | Pedicel | Epithecium Color & Key | Colour KOH 10% | Ascospores | Septa | Vegetation |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Patellaria barronii | Discoidal 500–730 µm | Thin striate | 33–40 µm = 37 | 35–40 µm = 37 | 101.3 × 14.8–15.3 µm ( = 101 × 15 µm) | Short unilobed | = 20 µm Color: 25E7 Green Jungle |  | 28–36 × 5–8 = 35 × 7 µm Clavate | 6–8 | XS |
| Patellaria esperanzae | Discoidal 400–600 µm | Thick smooth | 45–52 µm = 47.5 µm | 25–40 µm = 30 µm | 70–95 × 10 µm ( = 83 × 10) | - | = 25 µm Color: 27E5 Olive green |  | 16–20 × 4–5 = 18 × 4 µm Subfusiform | 3 | TMCF |
| Patellaria esquedii | Discoidal to irregular 600–1000 µm | Thick smooth | 35–40 µm = 37.3 µm | 35–40 µm = 37 µm | 85–98 × 13–17 µm ( = 91 × 13.83 µm) | Short unilobed | = 22.3 µm Color: 25E7 Green jungle |  | 27–32 × 6 = 30 × 6 µm Obclavate | 5–6 | TRF |
| Patellaria garciae | Discoidal 700–800 µm | Thin smooth | 50 µm | 30 µm | 100–115 × 15 µm ( = 108 × 15 µm) | Short bilobuled | = 30 µm Color: 25F3 Spruce green |  | 35–42 × 68 = 40 × 7 µm Clavate | 8–10 | CF |
| Patellaria magenta | Discoidal 600–1000 µm | Thick inovulte striate | 40–56 µm = 49.6 µm | 25–37 µm = 30.5 µm | 95–145 × 12–20 µm ( = 120.6 × 13.1 µm) | Long bilobed | = 25 µm Color: 25E7 Green jungle |  | 30–54 × 6–8 µm = 36 × 6 Clavate | 7–13 | TDF, XS |
| Patellaria mangrovei | Discoidal to ovoid 500–700 µm | Thin smooth | 40–43 µm = 42 | 60 µm | 75–90 × 14–16 µm ( = 83 × 15 µm) | Short unilobed | = 20 µm Color: 4E8 Olive brown |  | 25–35 × 6–9 = 30 × 7 µm Clavate | 5–8 | M |
| Patellaria neoleonensis | Discoidal 830–1000 µm | Thick involute striate | 45–62 µm = 55 µm | 20–35 µm = 28 | 93–154 × 12–20 ( = 109.7 × 14.7 µm) | Long bilobed | = 21 µm Color: 24F8 Dark turquoise |  | 32–50 × 6–8 = 40 × 6 µm Clavate | 8–10 | XS |
| Patellaria politecnica | Discoidal 700–860 µm | Thin smooth | 25–30 µm = 28.3 | 10 µm | 55–76 × 10–15 µm ( = 65.18 × 12.60 µm) | Short unilobed | = 18 µm Color: 24E7 Nautic bue |  | 21–32 × 4–5, = 25 × 4 µm Allantoid | 3–6 | TRF |
| Patellaria potosina | Discoidal 550–750 µm | Thin striate | 35–45 µm = 40 µm | 40–55 µm = 45 µm | 75–116 × 11–15 µm ( = 91.42 × 12.56 µm) | Short bilobed | = 22.5 µm Color: 24E7 Nautic bue |  | 31–38 × 5–7 = 31 × 6 µm Obclavate | 5–8 | XS |
| Patellaria ramona | Irregular 450–700 µm | Thin smooth | 25–30 µm | 35–40 µm = 37.5 µm | 75–96 × 15–17 µm ( = 88.5 × 15.8 µm) | Long bilobed | = 15 µm Color: 24F8 Dark turquoise |  | 22–29 × 6 = 2 × 6 µm Clavate | 4–6 | TRF |
| Patellaria tropicalis | Discoidal irregular 530–800 µm | Thin smooth | 25–40 µm = 39.6 | 17–30 µm = 20.28 | 72–110 × 12–15 µm ( = 85.12 × 14.15 µm) | Short unilobed | = 20 µm Color: 25D8 Deep green |  | Obclavate 24–37 × 6–7 = 28 × 6 µm Clavate | 5–7 | TDF |
| Patellaria xerófila | Discoidal irregular 510–660 µm | Thin striate | 14–45 µm = 36 | 14–45 µm = 25 | 82–110 × 11–15 ( = 92.9 × 13.03 µm) | Short unilobed | = 17 µm Color: 24F8 Dark turquoise |  | 26–39 × 5–7 = 30 × 6 µm Clavate | 5–7 | XS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Jacobo, I.; Raymundo, T.; Martínez-González, C.R.; Martínez-Pineda, M.; Valenzuela, R. Phylogenetic and Morphological Analyses Reveal Twelve New Species of the Genus Patellaria (Dothideomycetes, Ascomycota) from Mexico. J. Fungi 2025, 11, 44. https://doi.org/10.3390/jof11010044
García-Jacobo I, Raymundo T, Martínez-González CR, Martínez-Pineda M, Valenzuela R. Phylogenetic and Morphological Analyses Reveal Twelve New Species of the Genus Patellaria (Dothideomycetes, Ascomycota) from Mexico. Journal of Fungi. 2025; 11(1):44. https://doi.org/10.3390/jof11010044
Chicago/Turabian StyleGarcía-Jacobo, Ilian, Tania Raymundo, Cesar R. Martínez-González, Michelle Martínez-Pineda, and Ricardo Valenzuela. 2025. "Phylogenetic and Morphological Analyses Reveal Twelve New Species of the Genus Patellaria (Dothideomycetes, Ascomycota) from Mexico" Journal of Fungi 11, no. 1: 44. https://doi.org/10.3390/jof11010044
APA StyleGarcía-Jacobo, I., Raymundo, T., Martínez-González, C. R., Martínez-Pineda, M., & Valenzuela, R. (2025). Phylogenetic and Morphological Analyses Reveal Twelve New Species of the Genus Patellaria (Dothideomycetes, Ascomycota) from Mexico. Journal of Fungi, 11(1), 44. https://doi.org/10.3390/jof11010044