
Phylogeny and Virulence Factors of Escherichia coli Isolated from Dogs with Pyometra

Abstract
:1. Introduction
2. Material and Methods
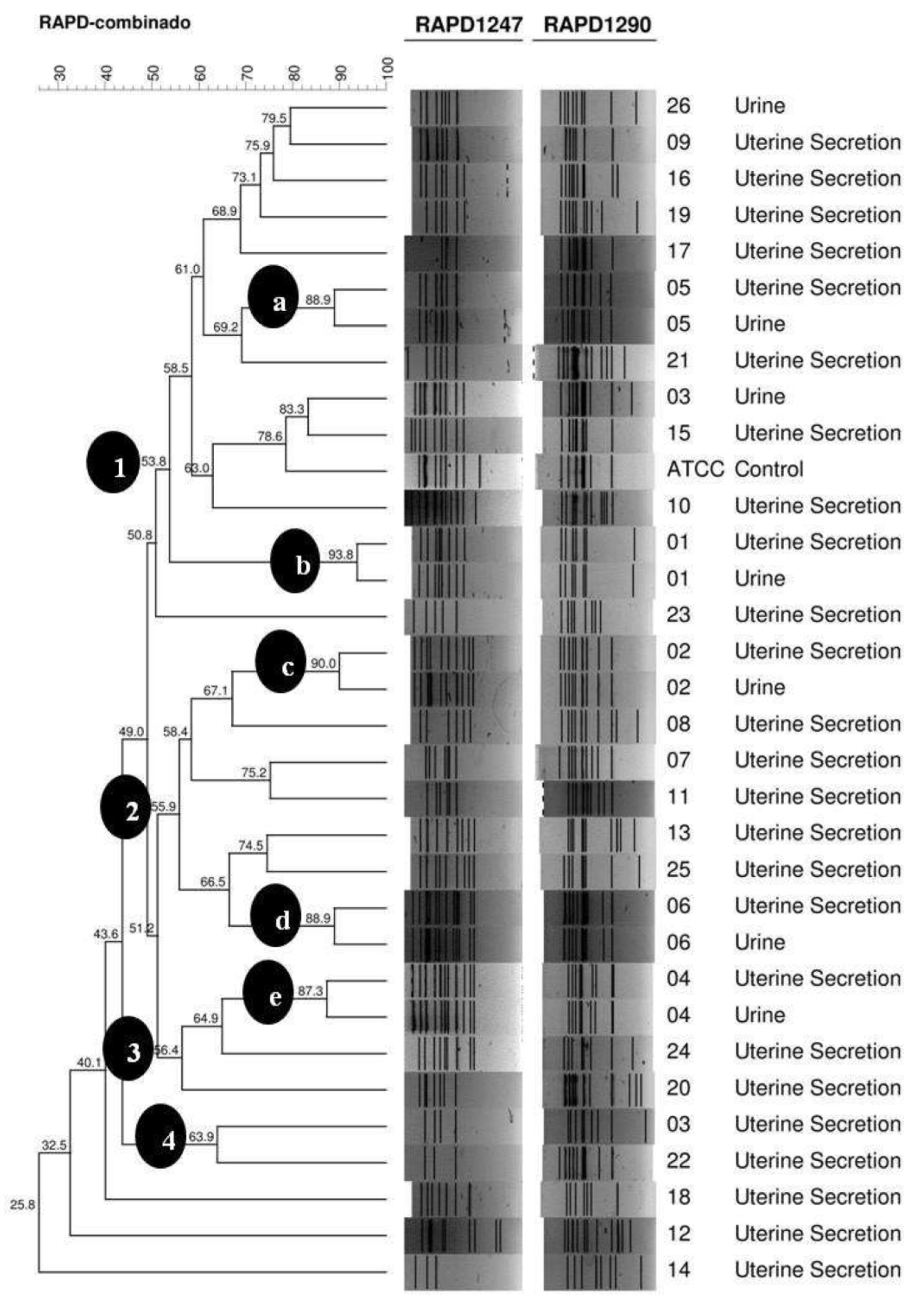
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Dam, T.; Das, P. Plasmids—Potential tool for the investigation of the gene transfer in Mycobacterium tuberculosis. J. Med. Microbiol. 2006, 55, 479–480. [Google Scholar] [PubMed] [Green Version]
- Egenvall, A.; Hagman, R.; Bonnett, B.N.; Hedhammar, A.; Olson, P.; Lagerstedt, A.S. Breed risk of pyometra in insured dogs in Sweden. J. Vet. Intern. Med. 2001, 15, 530–538. [Google Scholar] [PubMed]
- Fransson, B.; Lagerstedt, A.S.; Hellmen, E.; Jonsson, P. Bacteriology findings, blood chemistry profile and plasma endotoxin levels in bitches with pyometra or other uterine diseases. J. Vet. Med. 1997, 44, 417–423. [Google Scholar]
- Hagman, R.; Kühn, I. Escherichia coli strains isolated from the uterus and urinary bladder of bitches suffering from pyometra: Comparison by restriction enzyme digestion and pulsed- field gel electrophoresis. Vet. Microbiol. 2002, 84, 143–153. [Google Scholar]
- Waters, D.J.; Kengeri, S.S.; Maras, A.H.; Suckow, C.L.; Chiang, E.C. Life course analysis of the impact of mammary cancer and pyometra on age-anchored life expectancy in female Rottweilers: Implications for envisioning ovary conservation as a strategy to promote healthy longevity in pet dogs. Vet. J. 2017, 224, 25–37. [Google Scholar]
- Coggan, J.A.; Melville, P.A.; Oliveira, C.M.D.; Faustino, M.; Moreno, A.M.; Benites, N.R. Microbiological and histopathological aspects of canine pyometra. Braz. J. Microbiol. 2008, 39, 477–483. [Google Scholar]
- Da Silva, N.; Junqueira, V.C.A.; Silveira, N.F.A.; Taniwaki, M.H.; Gomes, R.A.R.; Okazaki, M.M. Escherichia coli O157:H7. In Manual De Métodos De Análise Microbiológica De Alimentos e Agua, 5th ed.; Manual of Methods for Microbiological Analysis of Food and Water; Blucher: São Paulo, Brazil, 2017; pp. 253–262. [Google Scholar]
- Hagman, R. Clinical and molecular characteristics of pyometra in female dogs. Reprod. Domest. Anim. 2012, 47, 323–325. [Google Scholar]
- Harada, K.; Okada, E.; Shimizu, T.; Kataoka, Y.; Sawada, T.; Takahashi, T. Antimicrobial resistance, virulence profiles and phylogenetic groups of faecal Escherichia coli isolates: A comparative analysis between dogs and their owners in Japan. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 139–144. [Google Scholar]
- Henriques, S.; Silva, E.; Lemsaddek, A.; Lopes-da-Costa, L.; Mateus, L. Genotypic and phenotypic comparison of Escherichia coli from uterine infections with different outcomes: Clinical metritis in the cow and pyometra in the bitch. Vet. Microbiol. 2014, 170, 109–116. [Google Scholar]
- Hopkins, K.L.; Hilton, A.C. Optimisation of random amplification of polymorphic DNA analysis for molecular subtyping of Escherichia coli O157. Lett. Appl. Microbiol. 2001, 32, 126–130. [Google Scholar]
- Johnson, J.R.; Kaster, N.; Kuskowski, M.A.; Ling, G.V. Identification of urovirulence traits in Escherichia coli by comparison of urinary and rectal E. coli isolates from dogs with urinary tract infection. J. Clin. Microbiol. 2003, 41, 337–345. [Google Scholar] [PubMed] [Green Version]
- Kuhnert, P.; Boerlin, P.; Frey, J. Target genes for virulence assessment of Escherichia coli isolates from water, food and the environment. FEMS Microbiol. Rev. 2000, 24, 107–117. [Google Scholar] [PubMed] [Green Version]
- Pereira, A.D.R.V.C. Antibioresistência Em Piometra (In Portuguese) Canina Anti-Bioresistance in Canine Pyometra. Master’s Thesis, Lusofona Veterinary Medicine of University of Humanities and Technologies, Lisboa, Portugal, November 2011. [Google Scholar]
- Slatter, D.H. Manual De Cirurgia De Pequenos Animais Small Animal Surgery Handbook, 3rd ed.; Manole: São Paulo, Brazil, 2007. (In Portuguese) [Google Scholar]
- Chew, D.J.; DiBartola, S.P.; Schenck, P. Canine and Feline Nephrology and Urology, 2nd ed.; Elsevier Saunders: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Sidjabat, H.E.; Chin, J.J.C.; Chapman, T.; Wu, K.; Ulett, G.C.; Ong, C.Y.; Schembri, M.A.; Johnson, J.R.; Trott, D.J. Colonisation dynamics and virulence of two clonal groups.s of multidrug-resistant Escherichia coli isolated from dogs. Microbes Infect. 2009, 11, 100–107. [Google Scholar] [PubMed]
- Borsoi, A.; Santin, E.; Santos, L.R.; Salle, C.T.P.; Moraes, H.L.S.; Nascimento, V.P. Inoculation of newly hatched broiler chicks with two Brazilian isolates of Salmonella Heidelberg strains with different virulence gene profiles, antimicrobial resistence and pulse field gel electrophoresis patterns to intestinal changes evaluation. Poult. Sci. 2009, 88, 750–758. [Google Scholar] [PubMed]
- Yamamoto, S.; Terai, A.; Yuri, K.; Kurazono, H.; Takeda, Y.; Yoshida, O. Detection of urovirulence factors in Escherichia coli by multiple polymerase chain reaction. FEMS Immunol. Med. Microbiol. 1995, 12, 85–90. [Google Scholar]
- Stenske, K.A.; Bemis, D.A.; Gillespie, B.E.; Oliver, S.P.; Draughon, F.A.; Matteson, K.J.; Bartges, J.W. Prevalence of urovirulence genes cnf, hlyD, sfa/foc and papGIII in faecal Escherichia coli from healthy dogs and their owners. Am. J. Vet. Res. 2009, 70, 1401–1406. [Google Scholar]
- Tarchouna, M.; Ferjani, A.; Ben-Selma, W.; Boukadida, J. Distribution of uropathogenic virulence genes in Escherichia coli isolated from patients with urinary tract infection. Int. J. Infect. Dis. 2013, 17, e450–e453. [Google Scholar]
- De Medici, D.; Croci, L.; Delibato, E.; Di Pasquale, S.; Filetici, E.; Toti, L. Evaluation of DNA extraction methods for use in combination with SYBR green I real-time PCR to detect Salmonella enterica serotype enteritidis in poultry. Appl. Environ. Microbiol. 2003, 69, 3456–3461. [Google Scholar]
- Von Sydow, G.A.C.M.O.N. Avaliação Da Ocorrência De Fatores De Virulência Em Estirpes De Escherichia coli Em Fezes De Cães Errantes. Master’s Thesis, University of Veterinary Medicine and Animal Science of São Paulo, São Paulo, Brazil, August 2005. (In Portuguese). [Google Scholar]
- Coggan, J.A. Estudo Microbiológico De Conteúdo Intra-Uterino E Histopatológico De Utero De Cadelas Com Piometra E Pesquisa De Fatores De Virulência Em Cepas De E. coli E O Potencial Risco à Saúde Humana. Master’s Thesis, University of Veterinary Medicine and Animal Science of São Paulo, São Paulo, Brazil, September 2005. (In Portuguese). [Google Scholar]
- Hagman, R. Canine pyometra: What is new? Reprod. Domest. Anim. 2017, 52, 288–292. [Google Scholar]
- Berl, C.A.; Franco, C.R.; Baltazar, F.N.; Cortez, M.B.X.; Trevisan, R.; Cirillo, T.; Júnior, W. Perfil de suscetibilidade a antimicrobianos de bactérias isoladas da secreção uterina de cadelas com piometra atendidas em hospital veterinário localizado em São Paulo, SP, Brasil, no período de 2010 a 2015. Antimicrobial susceptibility profile of bacteria isolated from uterine secretion of bitches with pyometra treated at a veterinary hospital located in São Paulo, SP, Brazil, from 2010 to 2015. Rev. MV Z 2018, 16, 36–42. (In Portuguese) [Google Scholar]
- Chen, Y.; Liu, Z.; Zhang, Y.; Zhang, Z.; Lei, L.; Xia, Z. Increasing prevalence of ESBL-producing multidrug-resistant Escherichia coli from diseased pets in Beijing, China from 2012 to 2017. Front. Microbiol. 2019, 10, 2852. [Google Scholar] [PubMed]
- Chen, Y.M.; Wright, P.J.; Lee, C.S.; Browning, G.F. Uropathogenic virulence factors in isolates of Escherichia coli from clinical cases of canine pyometra and faeces of healthy bitches. Vet. Microbiol. 2003, 96, 57–69. [Google Scholar]
- Nguyen, M.C.P.; Woerther, P.L.; Bouvet, M.; Andremont, A.; Leclercq, R.; Canu, A. Escherichia coli as reservoir for macrolide resistance genes. Emerg. Infect. Dis. 2009, 15, 1648–1650. [Google Scholar]
- Osugui, L.; de Castro, A.P.; Iovine, R.; Irino, K.; Carvalho, V.M. Virulence genotypes, antibiotic resistance and the phylogenetic background of extraintestinal pathogenic Escherichia coli isolated from urinary tract infections of dogs and cats in Brazil. Vet. Microbiol. 2014, 171, 242–247. [Google Scholar]
- Pretzer, S.D. Clinical presentation of canine pyometra and mucometra: A review. Theriogenology 2008, 70, 359–363. [Google Scholar]
- Rippere-Lampe, K.E.; O’Brien, A.D.; Conran, R.; Lockman, H. A Mutation of gene encoding cytotoxic necrotizing factor type 1 (cnf1) attenuates the virulence of uropathogenic Escherichia coli. Infect. Immun. 2001, 69, 3954–3964. [Google Scholar]
- Siqueira, A.K.; Ribeiro, M.G.; Leite, D.D.S.; Tiba, M.R.; de Moura, C.; Lopes, M.D.; Prestes, N.C.; Salerno, T.; da Silva, A.V. Virulence factors in Escherichia coli strains isolated from urinary tract infection and pyometra cases and from faeces of healthy dogs. Res. Vet. Sci. 2009, 86, 206–210. [Google Scholar]
- Sandholm, M.; Vasenius, H.; Kivistö, A. Pathogenesis of canine pyometra. J. Am. Vet. Med. Assoc. 1975, 167, 1006–1010. [Google Scholar]
- Maluta, R.P.; Borges, C.A.; Beraldo, L.G.; Cardozo, M.V.; Voorwald, F.A.; Santana, A.M.; Rigobelo, E.C.; Toniollo, G.H.; Ávila, F.A. Frequencies of virulence genes and pulse field gel electrophoresis fingerprints in Escherichia coli isolates from canine pyometra. Vet. J. 2014, 202, 393–395. [Google Scholar]
- Trabulsi, L.R.; Alterthum, F. Microbiologia, 4th ed.; Atheneu: New York, NY, USA, 2005. [Google Scholar]
- Tramuta, C.; Nucera, D.; Robino, P.; Salvarani, S.; Nebbia, P. Virulence factors and genetic variability of uropathogenic Escherichia coli isolated from dogs and cats in Italy. J. Vet. Sci. 2011, 12, 49–55. [Google Scholar]
- Hagman, R. Pyometra in small animals. Vet. Clin. N. Am. Small Anim. Pract. 2018, 48, 639–661. [Google Scholar]
- Sary, K.; Fairbrother, J.M.; Arsenault, J.; de Lagarde, M.; Boulianne, M. Antimicrobial resistance and virulence gene profiles among Escherichia coli isolates from retail chicken carcasses in Vietnam. Foodborne Pathog. Dis. 2019, 16, 298–306. [Google Scholar] [PubMed]
- Tiba, M.R. Genotypic Determination of Virulence Factors in Samples of Escherichia coli Isolated from Cystitis. Rev. Inst. Med. Trop. São Paulo 2008, 50, 255–260. [Google Scholar] [PubMed] [Green Version]
- Santos Filho, J.C.B. Research of Bacteria and Their Sensitivity to Antimicrobials in Dogs with Pyometra, with Special Interest in the Characterization of Extra-Intestinal Pathogenic Escherichia coli (ExPEC). Master’s Thesis, Veterinary Medicine of University of São Paulo (UNIP), São Paulo, Brazil, 2008. [Google Scholar]
- Mulvey, M.A. Adhesion and entry of uropathogenic Escherichia coli. Cell Microbiol. 2002, 4, 257–271. [Google Scholar] [PubMed]
- Wadas, B.; Kühn, I.; Lagerstedt, A.S.; Jonsson, P. Biochemical phenotypes of Escherichia coli in dogs: Comparison of isolates from bitches suffering from pyometra and urinary tract infection with isolates from faeces of healthy dogs. Vet. Microbiol. 1996, 52, 293–300. [Google Scholar] [PubMed]

{kind=link}
| Genes | Sequence 5′→3′ | Molecular Weight | Positive Control |
|---|---|---|---|
| pap-1(R) | GCAACAGCAACGCTGGTTGCATCAT | 336 bp [19] a | E. coli J96 [20,21] c |
| pap-2(F) | AGAGAGAGCCACTCTTATACGGACA | ||
| hly-1(R) | AACAAGGATAAGCACTGTTCTGGCT | 1177 bp [19] a | E. coli J96 [20,21] c |
| hly-2(F) | ACCATATAAGCGGTCATTCCCGTCA | ||
| cnf-1(R) | AAGATGGAGTTTCCTATGCAGGAG | 498 bp [19] a | E. coli J96 [20,21] c |
| cnf-2(F) | CATTCAGAGTCCTGCCCTCATTATT | ||
| sfa-1(R) | CTCCGGAGAACTGGGTGCATCTTAC | 410 bp [19] a | E. coli J96 [20,21] c |
| sfa-2(F) | CGGAGGAGTAATTACAAACCTGGCA | ||
| afa-1(R) | GCTGGGCAGCAAACTGATAACCTC | 750 bp [20] b | E. coli A30 [21] c |
| afa-2(F) | CATCAAGCTGTTTGTTCGTCCGCCG |
| Animal | Breed | Age (Years) | Uterine Secretion | Urine |
|---|---|---|---|---|
| 01 | Mongrel | 8 | + | + |
| 02 | Mongrel | 12 | + | + |
| 03 | Mongrel | 14 | + | + |
| 04 | Pinscher | 8 | + | + |
| 05 | Rottweiler | 10 | + | + |
| 06 | BassetHound | 10 | + | + |
| 07 | Mongrel | 1 | + | − |
| 08 | Mongrel | 1 | + | − |
| 09 | Mongrel | 4 | + | − |
| 10 | Mongrel | 11 | + | − |
| 11 | Mongrel | 11 | + | − |
| 12 | Mongrel | 13 | + | − |
| 13 | Poodle | 9 | + | − |
| 14 | Poodle | 10 | + | − |
| 15 | Poodle | 10 | + | − |
| 16 | Poodle | 11 | + | − |
| 17 | Poodle | 11 | + | − |
| 18 | Pitt Bull | 8 | + | − |
| 19 | Pitt Bull | 9 | + | − |
| 20 | Pitt Bull | 10 | + | − |
| 21 | Rottweiler | 9 | + | − |
| 22 | Rottweiler | 10 | + | − |
| 23 | BassetHound | 6 | + | − |
| 24 | Sharpei | 4 | + | − |
| 25 | Akita | 8 | + | − |
| 26 | Mongrel | 7 | − | + |
| Prevalence of Virulence Genes | Presence—N (%) |
|---|---|
| Pap | 15 (46.9) |
| Hly | 17 (53.1) |
| Afa | 19 (59.4) |
| Cnf | 22 (68.8) |
| Sfa | 31 (96.9) |
| Virulence Profiles | Presence—N (%) |
| V1: sfa | 8 (25.0) |
| V2: pap, cnf | 1 (3.1) |
| V3: afa, sfa | 1 (3.1) |
| V4: cnf, sfa | 1 (3.1) |
| V5: pap, afa, sfa | 1 (3.1) |
| V6: cnf, hly, sfa | 1 (3.1) |
| V7: cnf, afa, sfa | 3 (9.4) |
| V8: pap, cnf, hly, sfa | 2 (6.2) |
| V9: cnf, afa, hly, sfa | 3 (9.4) |
| V10: all genes | 11 (34.4) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melo, R.T.; Oliveira, R.P.; Silva, B.F.; Monteiro, G.P.; Saut, J.P.E.; Costa, L.R.M.; Dias, S.D.C.; Rossi, D.A. Phylogeny and Virulence Factors of Escherichia coli Isolated from Dogs with Pyometra. Vet. Sci. 2022, 9, 158. https://doi.org/10.3390/vetsci9040158
Melo RT, Oliveira RP, Silva BF, Monteiro GP, Saut JPE, Costa LRM, Dias SDC, Rossi DA. Phylogeny and Virulence Factors of Escherichia coli Isolated from Dogs with Pyometra. Veterinary Sciences. 2022; 9(4):158. https://doi.org/10.3390/vetsci9040158
Chicago/Turabian StyleMelo, Roberta T., Raquel P. Oliveira, Beatryz F. Silva, Guilherme P. Monteiro, João Paulo E. Saut, Letícia R. M. Costa, Sthéfany Da C. Dias, and Daise A. Rossi. 2022. "Phylogeny and Virulence Factors of Escherichia coli Isolated from Dogs with Pyometra" Veterinary Sciences 9, no. 4: 158. https://doi.org/10.3390/vetsci9040158
APA StyleMelo, R. T., Oliveira, R. P., Silva, B. F., Monteiro, G. P., Saut, J. P. E., Costa, L. R. M., Dias, S. D. C., & Rossi, D. A. (2022). Phylogeny and Virulence Factors of Escherichia coli Isolated from Dogs with Pyometra. Veterinary Sciences, 9(4), 158. https://doi.org/10.3390/vetsci9040158