
Carbon Nanotube-Based Scaffolds for Cardiac Tissue Engineering—Systematic Review and Narrative Synthesis



Abstract
1. Introduction
2. Methodology
2.1. Objectives
2.2. Methods
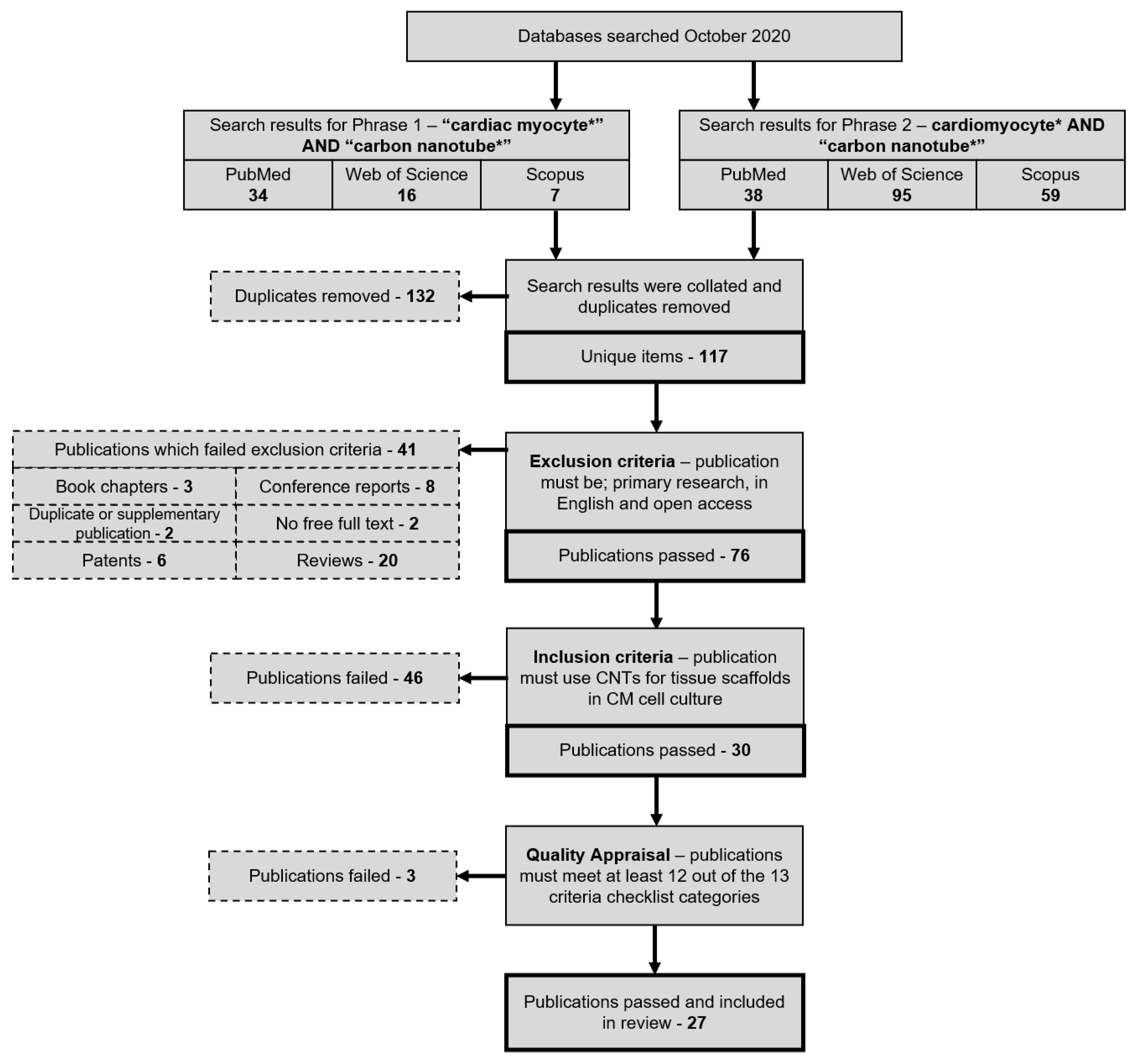
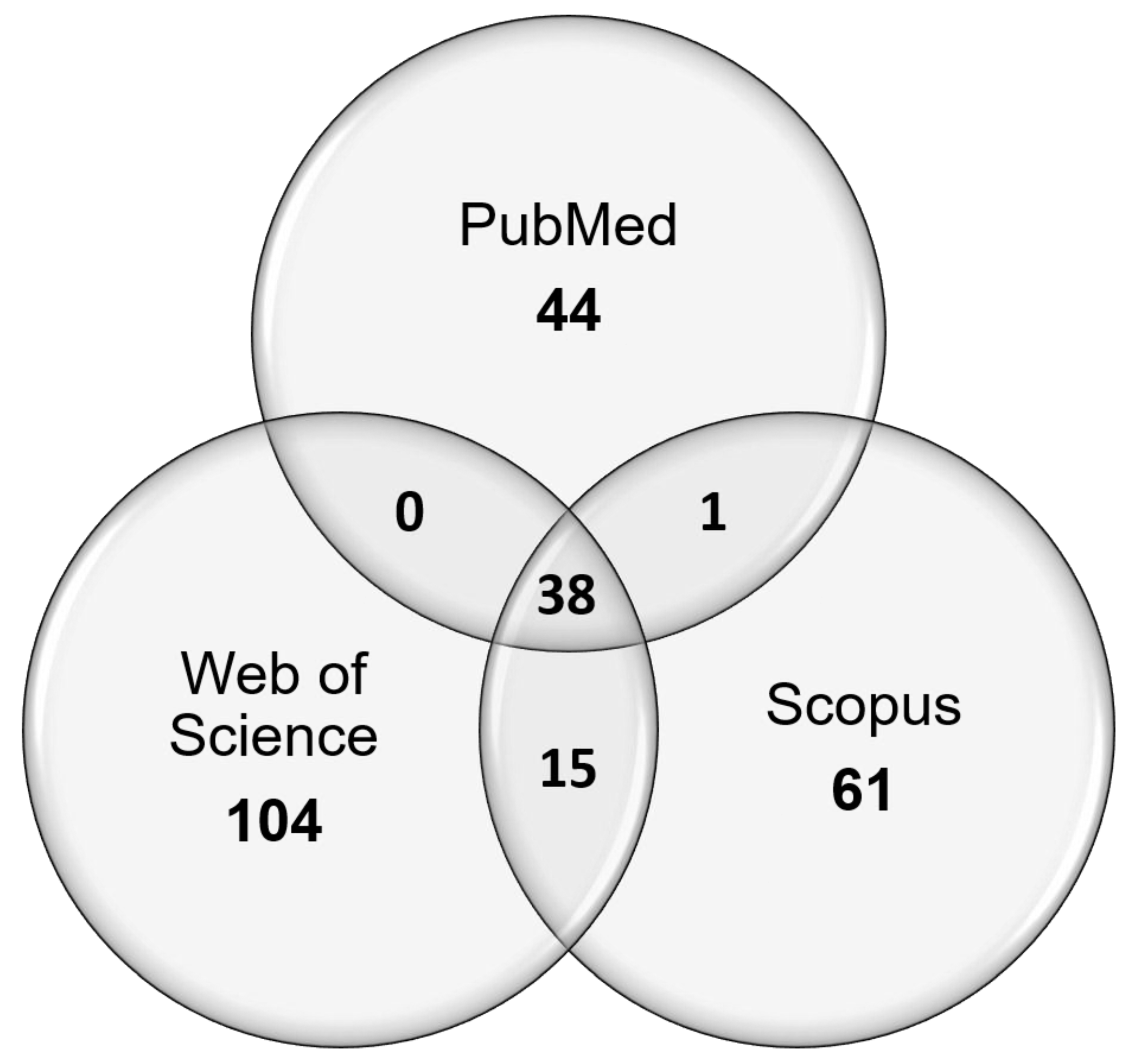
2.3. Search Strategy
2.4. Search Attrition Criteria
2.5. Article Processing and Selection
2.6. Quality Appraisal
- Problem Statement, Conceptual Framework, and Research Question
- Reference to the Literature and Documentation
- Relevance
- Research Design
- Instrumentation, Data Collection and Quality Control
- Population and Sample
- Data Analysis and Statistics
- Reporting of Statistical Analyses
- Presentation of Results
- Discussion and Conclusions: Interpretation
- Title, Authors, and Abstract
- Presentation and Documentation
- Scientific Conduct
2.7. Data Extraction
3. Results
3.1. Search Breakdown
3.2. Quality Appraisal
3.3. Experimental Design
3.4. Experimental Materials
3.5. Experimental Platforms
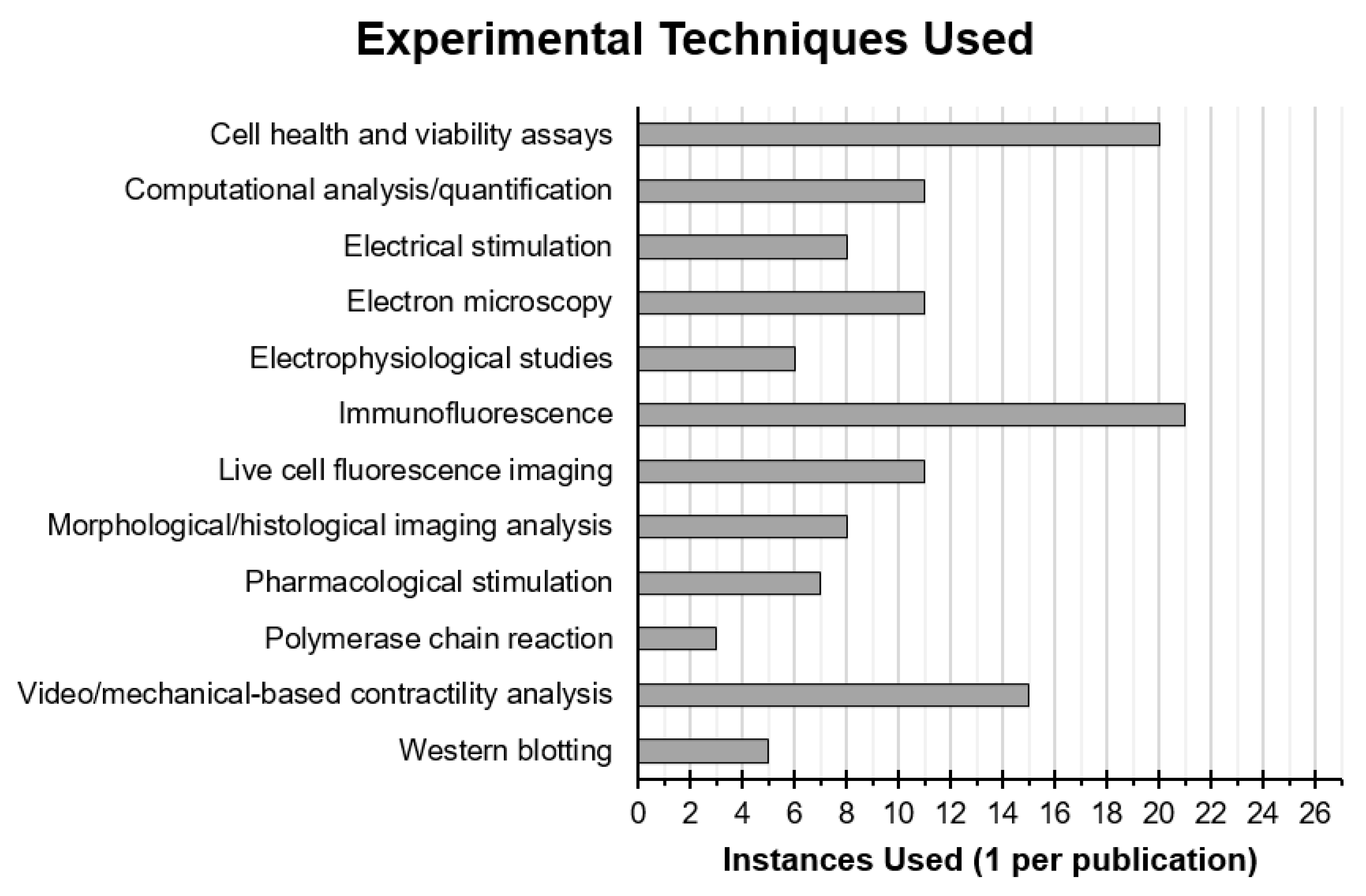
3.6. Experimental Techniques
3.7. Experimental Results
4. Discussion
4.1. The Effects of Carbon Nanotubes on Cell Structure
4.2. The Effects of Carbon Nanotubes on Cell Function
5. Limitations of This Study
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- WHO (World Health Organization). Global Health Estimates 2016: Deaths by Cause, Age, Sex, by Country and by Region, 2000–2016; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- WHO (World Health Organization). The Atlas of Heart Disease and Stroke—The Future. Available online: https://www.who.int/cardiovascular_diseases/resources/atlas/en/ (accessed on 12 March 2021).
- Wilkins, E.; Wilson, L.; Wickramasinghe, K.; Bhatnagar, P.; Leal, J.; Luengo-Fernandez, R.; Burns, R.; Rayner, M.; Townsend, N. European Cardiovascular Disease Statistics, 2017th ed.; European Heart Network: Brussels, Belgium, 2017. [Google Scholar]
- Hwang, T.J.; Lauffenburger, J.C.; Franklin, J.M.; Kesselheim, A.S. Temporal Trends and Factors Associated With Cardiovascular Drug Development, 1990 to 2012. JACC Basic Transl. Sci. 2016, 1, 301–308. [Google Scholar] [CrossRef]
- Pomeroy, J.E.; Helfer, A.; Bursac, N. Biomaterializing the promise of cardiac tissue engineering. Biotechnol. Adv. 2019, 42, 107353. [Google Scholar] [CrossRef]
- Jawad, H.; Ali, N.N.; Lyon, A.R.; Chen, Q.Z.; Harding, S.E.; Boccaccini, A.R. Myocardial tissue engineering: A review. J. Tissue Eng. Regen. Med. 2007, 1, 327–342. [Google Scholar] [CrossRef] [PubMed]
- Curtis, M.W.; Russell, B. Cardiac Tissue Engineering. J. Cardiovasc. Nurs. 2009, 24, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Wong, J.; Wen, J.; Wang, S.; Wang, C.; Spiering, S.; Kan, N.G.; Forcales, S.; Puri, P.L.; Leone, T.C.; et al. Studying arrhythmogenic right ventricular dysplasia with patient-specific iPSCs. Nature 2013, 494, 105–110. [Google Scholar] [CrossRef]
- Lee, E.K.; Tran, D.D.; Keung, W.; Chan, P.; Wong, G.; Chan, C.W.; Costa, K.D.; Li, R.A.; Khine, M. Machine Learning of Human Pluripotent Stem Cell-Derived Engineered Cardiac Tissue Contractility for Automated Drug Classification. Stem. Cell Rep. 2017, 9, 1560–1572. [Google Scholar] [CrossRef]
- Nguyen, A.H.; Marsh, P.; Schmiess-Heine, L.; Burke, P.J.; Lee, A.; Lee, J.; Cao, H. Cardiac tissue engineering: State-of-the-art methods and outlook. J. Biol. Eng. 2019, 13, 57. [Google Scholar] [CrossRef]
- Chaudhuri, R.; Ramachandran, M.; Moharil, P.; Harumalani, M.; Jaiswal, A.K. Biomaterials and cells for cardiac tissue engineering: Current choices. Mater. Sci. Eng. C 2017, 79, 950–957. [Google Scholar] [CrossRef] [PubMed]
- Veerman, C.C.; Kosmidis, G.; Mummery, C.L.; Casini, S.; Verkerk, A.O.; Bellin, M. Immaturity of Human Stem-Cell-Derived Cardiomyocytes in Culture: Fatal Flaw or Soluble Problem? Stem. Cells Dev. 2015, 24, 1035–1052. [Google Scholar] [CrossRef] [PubMed]
- Göktepe, S.; Abilez, O.J.; Parker, K.K.; Kuhl, E. A multiscale model for eccentric and concentric cardiac growth through sarcomerogenesis. J. Theor. Biol. 2010, 265, 433–442. [Google Scholar] [CrossRef]
- Samarel, A.M. Costameres, focal adhesions, and cardiomyocyte mechanotransduction. Am. J. Physiol. Hear. Circ. Physiol. 2005, 289, H2291–H2301. [Google Scholar] [CrossRef] [PubMed]
- Jongsma, H.J.; Wilders, R. Gap junctions in cardiovascular disease. Circ. Res. 2000, 86, 1193–1197. [Google Scholar] [CrossRef]
- Gregorio, C.C.; Antin, P.B. To the heart of myofibril assembly. Trends Cell Biol. 2000, 10, 355–362. [Google Scholar] [CrossRef]
- Louch, W.E.; Sheehan, K.A.; Wolska, B.M. Methods in cardiomyocyte isolation, culture, and gene transfer. J. Mol. Cell. Cardiol. 2011, 51, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Zuppinger, C.; Gibbons, G.; Dutta-Passecker, P.; Segiser, A.; Most, H.; Suter, T.M. Characterization of cytoskeleton features and maturation status of cultured human iPSC-derived cardiomyocytes. Eur. J. Histochem. 2017, 61, 145–153. [Google Scholar] [CrossRef]
- Giacomelli, E.; Meraviglia, V.; Campostrini, G.; Cochrane, A.; Cao, X.; van Helden, R.W.J.; Krotenberg Garcia, A.; Mircea, M.; Kostidis, S.; Davis, R.P.; et al. Human-iPSC-Derived Cardiac Stromal Cells Enhance Maturation in 3D Cardiac Microtissues and Reveal Non-cardiomyocyte Contributions to Heart Disease. Cell Stem Cell 2020, 26, 862–879. [Google Scholar] [CrossRef]
- King, A.A.K.; Matta-Domjan, B.; Large, M.J.; Matta, C.; Ogilvie, S.P.; Bardi, N.; Byrne, H.J.; Zakhidov, A.; Jurewicz, I.; Velliou, E.; et al. Pristine carbon nanotube scaffolds for the growth of chondrocytes. J. Mater. Chem. B 2017, 5, 8178–8182. [Google Scholar] [CrossRef]
- Brunner, E.W.; Jurewicz, I.; Heister, E.; Fahimi, A.; Bo, C.; Sear, R.P.; Donovan, P.J.; Dalton, A.B. Growth and proliferation of human embryonic stem cells on fully synthetic scaffolds based on carbon nanotubes. ACS Appl. Mater. Interfaces 2014, 6, 2598–2603. [Google Scholar] [CrossRef]
- Matta-Domjan, B.; King, A.; Totti, S.; Matta, C.; Dover, G.; Martinez, P.; Zakhidov, A.; La Ragione, R.; Macedo, H.; Jurewicz, I.; et al. Biophysical interactions between pancreatic cancer cells and pristine carbon nanotube substrates: Potential application for pancreatic cancer tissue engineering. J. Biomed. Mater. Res. Part B Appl. Biomater. 2017, 106, 1637–1644. [Google Scholar] [CrossRef]
- Heister, E.; Brunner, E.W.; Dieckmann, G.R.; Jurewicz, I.; Dalton, A.B. Are carbon nanotubes a natural solution? Applications in biology and medicine. ACS Appl. Mater. Interfaces 2013, 5, 1870–1891. [Google Scholar] [CrossRef]
- Gorain, B.; Choudhury, H.; Pandey, M.; Kesharwani, P.; Abeer, M.M.; Tekade, R.K.; Hussain, Z. Carbon nanotube scaffolds as emerging nanoplatform for myocardial tissue regeneration: A review of recent developments and therapeutic implications. Biomed. Pharmacother. 2018, 104, 496–508. [Google Scholar] [CrossRef]
- Singh, E.; Srivastava, R.; Kumar, U.; Katheria, A.D. Carbon Nanotube: A Review on Introduction, Fabrication Techniques and Optical Applications. Nanosci. Nanotechnol. Res. Vol. 2017, 4, 120–126. [Google Scholar] [CrossRef]
- Dresselhaus, M.; Dresselhaus, G.; Avouris, P. Carbon Nanotubes—Synthesis, Structure, Properties, and Applications; Dresselhaus, M., Dresselhaus, G., Avouris, P., Eds.; Springer: New York, NY, USA, 2001; Volume 53. [Google Scholar]
- Iijima, S. Helical microtubules of graphitic carbon. Nature 1991, 354, 56–58. [Google Scholar] [CrossRef]
- Schroeder, V.; Savagatrup, S.; He, M.; Lin, S.; Swager, T.M. Carbon nanotube chemical sensors. Chem. Rev. 2019, 119, 599–663. [Google Scholar] [CrossRef]
- Blackburn, J.L.; Ferguson, A.J.; Cho, C.; Grunlan, J.C. Carbon-Nanotube-Based Thermoelectric Materials and Devices. Adv. Mater. 2018, 30, 1704386. [Google Scholar] [CrossRef] [PubMed]
- Avouris, P.; Freitag, M.; Perebeinos, V. Carbon-nanotube photonics and optoelectronics. Nat. Photonics 2008, 2, 341–350. [Google Scholar] [CrossRef]
- Jurewicz, I.; Worajittiphon, P.; King, A.A.K.; Sellin, P.J.; Keddie, J.L.; Dalton, A.B. Locking carbon nanotubes in confined lattice geometries—A route to low percolation in conducting composites. J. Phys. Chem. B 2011, 115, 6395–6400. [Google Scholar] [CrossRef]
- Peng, B.; Locascio, M.; Zapol, P.; Li, S.; Mielke, S.L.; Schatz, G.C.; Espinosa, H.D. Measurements of near-ultimate strength for multiwalled carbon nanotubes and irradiation-induced crosslinking improvements. Nat. Nanotechnol. 2008, 3, 626–631. [Google Scholar] [CrossRef]
- Filleter, T.; Bernal, R.; Li, S.; Espinosa, H.D. Ultrahigh strength and stiffness in cross-linked hierarchical carbon nanotube bundles. Adv. Mater. 2011, 23, 2855–2860. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.F.; Files, B.S.; Arepalli, S.; Ruoff, R.S. Tensile loading of ropes of single wall carbon nanotubes and their mechanical properties. Phys. Rev. Lett. 2000, 84, 5552–5555. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.F.; Lourie, O.; Dyer, M.J.; Moloni, K.; Kelly, T.F.; Ruoff, R.S. Strength and breaking mechanism of multiwalled carbon nanotubes under tensile load. Science 2000, 287, 637–640. [Google Scholar] [CrossRef]
- Salvetat, J.P.; Briggs, G.A.D.; Bonard, J.M.; Bacsa, R.R.; Kulik, A.J.; Stöckli, T.; Burnham, N.A.; Forró, L. Elastic and shear moduli of single-walled carbon nanotube ropes. Phys. Rev. Lett. 1999, 82, 944–947. [Google Scholar] [CrossRef]
- Bhatia, R.; Kumari, K.; Rani, R.; Suri, A.; Pahuja, U.; Singh, D. A critical review of experimental results on low temperature charge transport in carbon nanotubes based composites. Rev. Phys. 2018, 3, 15–25. [Google Scholar] [CrossRef]
- Kumanek, B.; Janas, D. Thermal conductivity of carbon nanotube networks: A review. J. Mater. Sci. 2019, 54, 7397–7427. [Google Scholar] [CrossRef]
- Ausman, K.D.; Piner, R.; Lourie, O.; Ruoff, R.S.; Korobov, M. Organic solvent dispersions of single-walled carbon nanotubes: Toward solutions of pristine nanotubes. J. Phys. Chem. B 2000, 104, 8911–8915. [Google Scholar] [CrossRef]
- Furtado, C.A.; Kim, U.J.; Gutierrez, H.R.; Pan, L.; Dickey, E.C.; Eklund, P.C. Debundling and Dissolution of Single-Walled Carbon Nanotubes in Amide Solvents. J. Am. Chem. Soc. 2004, 126, 6095–6105. [Google Scholar] [CrossRef]
- Bergin, S.D.; Sun, Z.; Streich, P.; Hamilton, J.; Coleman, J.N. New solvents for nanotubes: Approaching the dispersibility of surfactants. J. Phys. Chem. C 2010, 114, 231–237. [Google Scholar] [CrossRef]
- Klumpp, C.; Kostarelos, K.; Prato, M.; Bianco, A. Functionalized carbon nanotubes as emerging nanovectors for the delivery of therapeutics. Biochim. Biophys. Acta Biomembr. 2006, 1758, 404–412. [Google Scholar] [CrossRef]
- Giordani, S.; Bergin, S.D.; Nicolosi, V.; Lebedkin, S.; Kappes, M.M.; Blau, W.J.; Coleman, J.N. Debundling of single-walled nanotubes by dilution: Observation of large populations of individual nanotubes in amide solvent dispersions. J. Phys. Chem. B 2006, 110, 15708–15718. [Google Scholar] [CrossRef] [PubMed]
- Vaisman, L.; Wagner, H.D.; Marom, G. The role of surfactants in dispersion of carbon nanotubes. Adv. Colloid Interface Sci. 2006, 128, 37–46. [Google Scholar] [CrossRef]
- Arechabala, B.; Coiffard, C.; Rivalland, P.; Coiffard, L.J.M.; de Roeck-Holtzhauer, Y. Comparison of cytotoxicity of various surfactants tested on normal human fibroblast cultures using the neutral red test, MTT assay and LDH release. J. Appl. Toxicol. 1999, 19, 163–165. [Google Scholar] [CrossRef]
- Joshi, D.R.; Adhikari, N. An Overview on Common Organic Solvents and Their Toxicity. J. Pharm. Res. Int. 2019, 28, 1–18. [Google Scholar] [CrossRef]
- Dai, H. Carbon nanotubes: Synthesis, integration, and properties. Acc. Chem. Res. 2002, 35, 1035–1044. [Google Scholar] [CrossRef]
- Chen, C.; Bai, X.; Ding, Y.; Lee, I.S. Electrical stimulation as a novel tool for regulating cell behavior in tissue engineering. Biomater. Res. 2019, 23, 25. [Google Scholar] [CrossRef]
- Li, Y.; Huang, G.; Zhang, X.; Wang, L.; Du, Y.; Lu, T.J.; Xu, F. Engineering cell alignment in vitro. Biotechnol. Adv. 2014, 32, 347–365. [Google Scholar] [CrossRef]
- Radisic, M.; Park, H.; Shing, H.; Consi, T.; Schoen, F.J.; Langer, R.; Freed, L.E.; Vunjak-Novakovic, G. Functional assembly of engineered myocardium by electrical stimulation of cardiac myocytes cultured on scaffolds. Proc. Natl. Acad. Sci. USA 2004, 101, 18129–18134. [Google Scholar] [CrossRef]
- Sathaye, A.; Bursac, N.; Sheehy, S.; Tung, L. Electrical pacing counteracts intrinsic shortening of action potential duration of neonatal rat ventricular cells in culture. J. Mol. Cell. Cardiol. 2006, 41, 633–641. [Google Scholar] [CrossRef]
- Martherus, R.S.R.M.; Vanherle, S.J.V.; Timmer, E.D.J.; Zeijlemaker, V.A.; Broers, J.L.; Smeets, H.J.; Geraedts, J.P.; Ayoubi, T.A.Y. Electrical signals affect the cardiomyocyte transcriptome independently of contraction. Physiol. Genom. 2010, 42, 283–289. [Google Scholar] [CrossRef][Green Version]
- Kim, M.H.; Kino-Oka, M.; Kawase, M.; Yagi, K.; Taya, M. Response of human epithelial cells to culture surfaces with varied roughnesses prepared by immobilizing dendrimers with/without d-glucose display. J. Biosci. Bioeng. 2007, 103, 192–199. [Google Scholar] [CrossRef]
- Riva, J.J.; Malik, K.M.P.; Burnie, S.J.; Endicott, A.R.; Busse, J.W. What is your research question? An introduction to the PICOT format for clinicians. J. Can. Chiropr. Assoc. 2012, 56, 167–171. [Google Scholar]
- Shamseer, L.; Moher, D.; Clarke, M.; Ghersi, D.; Liberati, A.; Petticrew, M.; Shekelle, P.; Stewart, L.A.; Altman, D.G.; Booth, A.; et al. Preferred reporting items for systematic review and meta-analysis protocols (prisma-p) 2015: Elaboration and explanation. BMJ 2015, 349, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Bordage, G.; Caelleigh, A.; Steinecke, A.; Bland, C.; Crandall, S.; McGaghie, W.; Pangaro, L.; Penn, G.; Regehr, G.; Shea, J. Appendix 1—Checklist of Review Criteria. Acad. Med. 2001, 76, 958–959. [Google Scholar]
- Sun, L.; Chen, Z.; Bian, F.; Zhao, Y. Bioinspired Soft Robotic Caterpillar with Cardiomyocyte Drivers. Adv. Funct. Mater. 2020, 30, 1907820. [Google Scholar] [CrossRef]
- Wang, T.; Migliori, B.; Miccoli, B.; Shin, S.R. Bioinspired soft robot with incorporated microelectrodes. J. Vis. Exp. 2020, 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Zhang, X.; Li, B.; Huang, G.; Xu, F.; Zhang, X. Solvent-Free Fabrication of Carbon Nanotube/Silk Fibroin Electrospun Matrices for Enhancing Cardiomyocyte Functionalities. ACS Biomater. Sci. Eng. 2020, 6, 1630–1640. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Primo, F.; Kumar, S.A.; Manciu, F.S.; Joddar, B. Fabrication of surfactant-dispersed HiPco single-walled carbon nanotube-based alginate hydrogel composites as cellular products. Int. J. Mol. Sci. 2019, 20, 4802. [Google Scholar] [CrossRef]
- Chen, Z.; Fu, F.; Yu, Y.; Wang, H.; Shang, Y.; Zhao, Y. Cardiomyocytes-Actuated Morpho Butterfly Wings. Adv. Mater. 2019, 31, 1805431. [Google Scholar] [CrossRef]
- Lee, J.; Manoharan, V.; Cheung, L.; Lee, S.; Cha, B.H.; Newman, P.; Farzad, R.; Mehrotra, S.; Zhang, K.; Khan, F.; et al. Nanoparticle-Based Hybrid Scaffolds for Deciphering the Role of Multimodal Cues in Cardiac Tissue Engineering. ACS Nano 2019, 13, 12525–12539. [Google Scholar] [CrossRef]
- Roshanbinfar, K.; Mohammadi, Z.; Sheikh-Mahdi Mesgar, A.; Dehghan, M.M.; Oommen, O.P.; Hilborn, J.; Engel, F.B. Carbon nanotube doped pericardial matrix derived electroconductive biohybrid hydrogel for cardiac tissue engineering. Biomater. Sci. 2019, 7, 3906–3917. [Google Scholar] [CrossRef]
- Vaithilingam, J.; Sanjuan-Alberte, P.; Campora, S.; Rance, G.A.; Jiang, L.; Thorpe, J.; Burroughs, L.; Tuck, C.J.; Denning, C.; Wildman, R.D.; et al. Multifunctional Bioinstructive 3D Architectures to Modulate Cellular Behavior. Adv. Funct. Mater. 2019, 29, 1902016. [Google Scholar] [CrossRef]
- Hou, J.; Xie, Y.; Ji, A.; Cao, A.; Fang, Y.; Shi, E. Carbon-Nanotube-Wrapped Spider Silks for Directed Cardiomyocyte Growth and Electrophysiological Detection. ACS Appl. Mater. Interfaces 2018, 10, 6793–6798. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, V.; Bosi, S.; Penã, B.; Baj, G.; Long, C.S.; Sbaizero, O.; Giacca, M.; Prato, M.; Mestroni, L. 3D Carbon-Nanotube-Based Composites for Cardiac Tissue Engineering. ACS Appl. Bio Mater. 2018, 1, 1530–1537. [Google Scholar] [CrossRef]
- Wang, L.; Dou, W.; Malhi, M.; Zhu, M.; Liu, H.; Plakhotnik, J.; Xu, Z.; Zhao, Q.; Chen, J.; Chen, S.; et al. Microdevice Platform for Continuous Measurement of Contractility, Beating Rate, and Beating Rhythm of Human-Induced Pluripotent Stem Cell-Cardiomyocytes inside a Controlled Incubator Environment. ACS Appl. Mater. Interfaces 2018, 10, 21173–21183. [Google Scholar] [CrossRef] [PubMed]
- Ahadian, S.; Davenport Huyer, L.; Estili, M.; Yee, B.; Smith, N.; Xu, Z.; Sun, Y.; Radisic, M. Moldable elastomeric polyester-carbon nanotube scaffolds for cardiac tissue engineering. Acta Biomater. 2017, 52, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Peña, B.; Bosi, S.; Aguado, B.A.; Borin, D.; Farnsworth, N.L.; Dobrinskikh, E.; Rowland, T.J.; Martinelli, V.; Jeong, M.; Taylor, M.R.G.; et al. Injectable Carbon Nanotube-Functionalized Reverse Thermal Gel Promotes Cardiomyocytes Survival and Maturation. ACS Appl. Mater. Interfaces 2017, 9, 31645–31656. [Google Scholar] [CrossRef]
- Ren, J.; Xu, Q.; Chen, X.; Li, W.; Guo, K.; Zhao, Y.; Wang, Q.; Zhang, Z.; Peng, H.; Li, Y.G. Superaligned Carbon Nanotubes Guide Oriented Cell Growth and Promote Electrophysiological Homogeneity for Synthetic Cardiac Tissues. Adv. Mater. 2017, 29, 1702713. [Google Scholar] [CrossRef]
- Roshanbinfar, K.; Hilborn, J.; Varghese, O.P.; Oommen, O.P. Injectable and thermoresponsive pericardial matrix derived conductive scaffold for cardiac tissue engineering. RSC Adv. 2017, 7, 31980–31988. [Google Scholar] [CrossRef]
- Sherrell, P.C.; Cieślar-Pobuda, A.; Ejneby, M.S.; Sammalisto, L.; Gelmi, A.; de Muinck, E.; Brask, J.; Łos, M.J.; Rafat, M. Rational Design of a Conductive Collagen Heart Patch. Macromol. Biosci. 2017, 17, 1600446. [Google Scholar] [CrossRef]
- Sun, H.; Tang, J.; Mou, Y.; Zhou, J.; Qu, L.; Duval, K.; Huang, Z.; Lin, N.; Dai, R.; Liang, C.; et al. Carbon nanotube-composite hydrogels promote intercalated disc assembly in engineered cardiac tissues through β1-integrin mediated FAK and RhoA pathway. Acta Biomater. 2017, 48, 88–99. [Google Scholar] [CrossRef]
- Sun, H.; Zhou, J.; Huang, Z.; Qu, L.; Lin, N.; Liang, C.; Dai, R.; Tang, L.; Tian, F. Carbon nanotube-incorporated collagen hydrogels improve cell alignment and the performance of cardiac constructs. Int. J. Nanomed. 2017, 12, 3109–3120. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, L.; Guo, B.; Ma, P.X. Interwoven Aligned Conductive Nanofiber Yarn/Hydrogel Composite Scaffolds for Engineered 3D Cardiac Anisotropy. ACS Nano 2017, 11, 5646–5659. [Google Scholar] [CrossRef]
- Yu, H.; Zhao, H.; Huang, C.; Du, Y. Mechanically and Electrically Enhanced CNT-Collagen Hydrogels as Potential Scaffolds for Engineered Cardiac Constructs. ACS Biomater. Sci. Eng. 2017, 3, 3017–3021. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, J.; Xu, G.; Wei, J.; Zhang, Z.; Li, X. Tuning the conductivity and inner structure of electrospun fibers to promote cardiomyocyte elongation and synchronous beating. Mater. Sci. Eng. C 2016, 69, 865–874. [Google Scholar] [CrossRef]
- Shin, S.R.; Shin, C.; Memic, A.; Shadmehr, S.; Miscuglio, M.; Jung, H.Y.; Jung, S.M.; Bae, H.; Khademhosseini, A.; Tang, X.; et al. Aligned Carbon Nanotube-Based Flexible Gel Substrates for Engineering Biohybrid Tissue Actuators. Adv. Funct. Mater. 2015, 25, 4486–4495. [Google Scholar] [CrossRef]
- Sun, H.; Lü, S.; Jiang, X.X.; Li, X.; Li, H.; Lin, Q.; Mou, Y.; Zhao, Y.; Han, Y.; Zhou, J.; et al. Carbon nanotubes enhance intercalated disc assembly in cardiac myocytes via the β1-integrin-mediated signaling pathway. Biomaterials 2015, 55, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Kharaziha, M.; Shin, S.R.; Nikkhah, M.; Topkaya, S.N.; Masoumi, N.; Annabi, N.; Dokmeci, M.R.; Khademhosseini, A. Tough and flexible CNT-polymeric hybrid scaffolds for engineering cardiac constructs. Biomaterials 2014, 35, 7346–7354. [Google Scholar] [CrossRef]
- Pok, S.; Vitale, F.; Eichmann, S.L.; Benavides, O.M.; Pasquali, M.; Jacot, J.G. Biocompatible carbon nanotube-chitosan scaffold matching the electrical conductivity of the heart. ACS Nano 2014, 8, 9822–9832. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.; Wang, P.; Wang, L.; Xiao, L.; Zhang, F.; Na, J. Facile oxidation of superaligned carbon nanotube films for primary cell culture and genetic engineering. J. Mater. Chem. B 2014, 2, 471–476. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, J.; Sun, H.; Qiu, X.; Mou, Y.; Liu, Z.; Zhao, Y.; Li, X.; Han, Y.; Duan, C.; et al. Engineering the heart: Evaluation of conductive nanomaterials for improving implant integration and cardiac function. Sci. Rep. 2014, 4, 3733. [Google Scholar] [CrossRef]
- Martinelli, V.; Cellot, G.; Toma, F.M.; Long, C.S.; Caldwell, J.H.; Zentilin, L.; Giacca, M.; Turco, A.; Prato, M.; Ballerini, L.; et al. Carbon nanotubes instruct physiological growth and functionally mature syncytia: Nongenetic engineering of cardiac myocytes. ACS Nano 2013, 7, 5746–5756. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.R.; Jung, S.M.; Zalabany, M.; Kim, K.; Zorlutuna, P.; Kim, S.B.; Nikkhah, M.; Khabiry, M.; Azize, M.; Kong, J.; et al. Carbon-nanotube-embedded hydrogel sheets for engineering cardiac constructs and bioactuators. ACS Nano 2013, 7, 2369–2380. [Google Scholar] [CrossRef]
- Martinelli, V.; Cellot, G.; Toma, F.M.; Long, C.S.; Caldwell, J.H.; Zentilin, L.; Giacca, M.; Turco, A.; Prato, M.; Ballerini, L.; et al. Carbon nanotubes promote growth and spontaneous electrical activity in cultured cardiac myocytes. Nano Lett. 2012, 12, 1831–1838. [Google Scholar] [CrossRef] [PubMed]
- Karousis, N.; Tagmatarchis, N.; Tasis, D. Current progress on the chemical modification of carbon nanotubes. Chem. Rev. 2010, 110, 5366–5397. [Google Scholar] [CrossRef]
- Shoulders, M.D.; Raines, R.T. Collagen Structure and Stability Ann Rev Biochemistry. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef]
- Chang, C.W.; Dalgliesh, A.J.; López, J.E.; Griffiths, L.G. Cardiac extracellular matrix proteomics: Challenges, techniques, and clinical implications. Proteom. Clin. Appl. 2016, 10, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Byron, A.; Humphries, J.D.; Humphries, M.J. Defining the extracellular matrix using proteomics. Int. J. Exp. Pathol. 2013, 94, 75–92. [Google Scholar] [CrossRef] [PubMed]
- Lundgren, E.; Terracio, L.; Mårdh, S.; Borg, T.K. Extracellular matrix components influence the survival of adult cardiac myocytes in vitro. Exp. Cell Res. 1985, 158, 371–381. [Google Scholar] [CrossRef]
- Tonelli, F.M.P.; Santos, A.K.; Gomes, K.N.; Lorençon, E.; Guatimosim, S.; Ladeira, L.O.; Resende, R.R. Carbon nanotube interaction with extracellular matrix proteins producing scaffolds for tissue engineering. Int. J. Nanomed. 2012, 7, 4511–4529. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Noorman, M.; van der Heyden, M.A.G.; van Veen, T.A.B.; Cox, M.G.P.J.; Hauer, R.N.W.; de Bakker, J.M.T.; van Rijen, H.V.M. Cardiac cell-cell junctions in health and disease: Electrical versus mechanical coupling. J. Mol. Cell. Cardiol. 2009, 47, 23–31. [Google Scholar] [CrossRef]
- Jiang, Y.; Park, P.; Hong, S.M.; Ban, K. Maturation of cardiomyocytes derived from human pluripotent stem cells: Current strategies and limitations. Mol. Cells 2018, 41, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Scuderi, G.J.; Butcher, J. Naturally engineered maturation of cardiomyocytes. Front. Cell Dev. Biol. 2017, 5, 50. [Google Scholar] [CrossRef]
- Sjöblom, B.; Salmazo, A.; Djinović-Carugo, K. α-Actinin structure and regulation. Cell. Mol. Life Sci. 2008, 65, 2688–2701. [Google Scholar] [CrossRef]
- Sharma, S.; Jackson, P.G.; Makan, J. Cardiac troponins. J. Clin. Pathol. 2004, 57, 1025–1026. [Google Scholar] [CrossRef]
- Wu, X.; Sun, Z.; Foskett, A.; Trzeciakowski, J.P.; Meininger, G.A.; Muthuchamy, M. Cardiomyocyte contractile status is associated with differences in fibronectin and integrin interactions. Am. J. Physiol. Heart Circ. Physiol. 2010, 298, 2071–2081. [Google Scholar] [CrossRef] [PubMed]
- Hersch, N.; Wolters, B.; Dreissen, G.; Springer, R.; Kirchgessner, N.; Merkel, R.; Hoffmann, B. The constant beat: Cardiomyocytes adapt their forces by equal contraction upon environmental stiffening. Biol. Open 2013, 2, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Franke, W.W.; Borrmann, C.M.; Grund, C.; Pieperhoff, S. The area composita of adhering junctions connecting heart muscle cells of vertebrates. I. Molecular definition in intercalated disks of cardiomyocytes by immunoelectron microscopy of desmosomal proteins. Eur. J. Cell Biol. 2006, 85, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.K.; Hanson, D.A.; Schlaepfer, D.D. Focal adhesion kinase: In command and control of cell motility. Nat. Rev. Mol. Cell Biol. 2005, 6, 56–68. [Google Scholar] [CrossRef]
- Ribeiro-Rodrigues, T.M.; Martins-Marques, T.; Morel, S.; Kwak, B.R.; Girao, H. Role of connexin 43 in different forms of intercellular communication—Gap junctions, extracellular vesicles and tunnelling nanotubes. J. Cell Sci. 2017, 3619–3630. [Google Scholar] [CrossRef]
- Zuppinger, C.; Eppenberger-Eberhardt, M.; Eppenberger, H.M. N-Cadherin: Structure, function and importance in the formation of new intercalated disc-like cell contacts in cardiomyocytes. Heart Fail. Rev. 2000, 5, 251–257. [Google Scholar] [CrossRef]
- Vermij, S.H.; Abriel, H.; van Veen, T.A.B. Refining the molecular organization of the cardiac intercalated disc. Cardiovasc. Res. 2017, 113, 259–275. [Google Scholar] [CrossRef]
- Israeli-Rosenberg, S.; Manso, A.M.; Okada, H.; Ross, R.S. Integrins and integrin-associated proteins in the cardiac myocyte. Circ. Res. 2014, 114, 572–586. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.E.; Li, L.; Xia, X.; Fu, W.; Liao, Q.; Lan, C.; Yang, D.; Chen, H.; Yue, R.; Zeng, C.; et al. Dedifferentiation, proliferation, and redifferentiation of adult mammalian cardiomyocytes after ischemic injury. Circulation 2017, 136, 834–848. [Google Scholar] [CrossRef] [PubMed]
- Richardson, W.J.; Clarke, S.A.; Alexander Quinn, T.; Holmes, J.W. Physiological implications of myocardial scar structure. Compr. Physiol. 2015, 5, 1877–1909. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, P.; Tang, X.; Saif, T.A.; Bashir, R. Stiffness of the substrate influences the phenotype of embryonic chicken cardiac myocytes. J. Biomed. Mater. Res. Part A 2010, 95, 1261–1269. [Google Scholar] [CrossRef]
- Poland, C.A.; Duffin, R.; Kinloch, I.; Maynard, A.; Wallace, W.A.H.; Seaton, A.; Stone, V.; Brown, S.; MacNee, W.; Donaldson, K. Carbon nanotubes introduced into the abdominal cavity of mice show asbestos-like pathogenicity in a pilot study. Nat. Nanotechnol. 2008, 3, 423–428. [Google Scholar] [CrossRef]
- Chen, M.; von Mikecz, A. Formation of nucleoplasmic protein aggregates impairs nuclear function in response to SiO2 nanoparticles. Exp. Cell Res. 2005, 305, 51–62. [Google Scholar] [CrossRef]
- Alarifi, S.; Ali, D. Mechanisms of multi-walled carbon nanotubes-induced oxidative stress and genotoxicity in mouse fibroblast cells. Int. J. Toxicol. 2015, 34, 258–265. [Google Scholar] [CrossRef]
- Zhou, L.; Forman, H.J.; Ge, Y.; Lunec, J. Multi-walled carbon nanotubes: A cytotoxicity study in relation to functionalization, dose and dispersion. Toxicol. Vitr. 2017, 42, 292–298. [Google Scholar] [CrossRef]
- Valli, H.; Ahmad, S.; Fraser, J.A.; Jeevaratnam, K.; Huang, C.L.H. Pro-arrhythmic atrial phenotypes in incrementally paced murine Pgc1β−/−hearts: Effects of age. Exp. Physiol. 2017, 102, 1619–1634. [Google Scholar] [CrossRef]
- Mlinarić, A.; Horvat, M.; Smolčić, V.Š. Dealing with the positive publication bias: Why you should really publish your negative results. Biochem. Med. 2017, 27, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Harrison, B.S.; Atala, A. Carbon nanotube applications for tissue engineering. Biomaterials 2007, 28, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Home Office. Annual Statistics of Scientific Procedures on Living Animals Great Britain 2018; UK Government: London, UK, 2019; ISBN 9781528613361. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Publication | Categories of Checklist of Review Criteria [56] | Total Criteria Met | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | ||
| Sun 2020 [57] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Wang 2020 [58] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Zhao 2020 [59] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Alvarez-Primo 2020 [60] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Chen 2019 [61] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 11 | ||
| Lee 2019 [62] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Roshanbinfar 2019 [63] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Vaithilingam 2019 [64] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Hou 2018 [65] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Martinelli 2018 [66] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Wang 2018 [67] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Ahadian 2017 [68] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 13 |
| Peña 2017 [69] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Ren 2017 [70] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 13 |
| Roshanbinfar 2017 [71] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Sherrell 2017 [72] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Sun 2017 (1) [73] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Sun 2017 (2) [74] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Wu 2017 [75] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 11 | ||
| Yu 2017 [76] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Liu 2016 [77] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Shin 2015 [78] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Sun 2015 [79] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Kharaziha 2014 [80] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Pok 2014 [81] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 13 |
| Tao 2014 [82] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 13 |
| Zhou 2014 [83] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 11 | ||
| Martinelli 2013 [84] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Shin 2013 [85] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 12 | |
| Martinelli 2012 [86] | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 13 |
| Total Passed | 27 | |||||||||||||
| Publication | Materials Used | Techniques Used to Assess CM Viability & Function | Results & Discussion | Limitations Suggested by the Authors | Further Work Suggested by the Authors | |||
|---|---|---|---|---|---|---|---|---|
| Biological | Synthetic | |||||||
| Cells | Source | CNTs | Scaffold Material and Design | |||||
| Sun 2020 [57] | Neonatal cardiomyocytes | 1- to 2-day old rat pups | No information given | Caterpillar-inspired soft robot consisting of a magnetised GelMA layer with asymmetric claws, an aligned CNT-coated GelMA layer and an iridescent colour layer. | Confocal imaging of F-actin plus DAPI, MTT assay, SEM, visual analysis of CM contractions and soft robot actuation, pStim by isoproterenol, disease modelling by induced hyperkalemia. | Aligned CNTs improved electrical and mechanical properties of the scaffold, as well as improving biocompatibility by increasing cytoskeletal alignment and cell elongation. CM beating frequency was adversely affected by increased scaffold thickness and increased stiffness, influenced by GelMA percentage. pStim increased CM beating and speed of soft robot movement along PDMS “racetrack”. Increasing potassium concentration to induce hyperkalemia caused CM contractions to slow down and eventually stop, however pStim caused continuation of beating. | Only hyperkalemia tested, while there are many other “varieties of heart disease in actual clinical practice”. The microfluidics system was expected to perform more complex functions. | Further research needs to improve the platform and explore more applications for soft robots in clinical treatments. |
| Wang 2020 [58] | Neonatal cardiomyocytes | 2-day old Sprague Dawley rat pups | Purchased MWNTs—carboxylic functionalisation | Manta ray-inspired soft robot consisting of a gold electrode embedded between micropatterned PEGDA and GelMA-MWNT hydrogel layers. | Confocal IF of α-actinin and Cx43, video recording of CM contractions and soft robot actuation, field and direct eStim. | Precise spacing of the CNT-GelMa micropattern improved CM orientation and alignment—75µm spacing induced alignment and CMs were interconnected for synchronous beating. CMs on CNT-GelMA showed good alignment and elongation as well as uniaxial sarcomere alignment with interconnecting structures between cells. This was improved where cells were directly above the gold electrodes. eStim allowed a moderate amount of control over soft robot actuations without harming CMs. CNT cytotoxicity was acknowledged but said to be limited by only using CNT concentrations up to 5 mg/mL in the scaffold. | Parts of the production process are technically very challenging. | Optimise the fabrication process to improve ease of production. |
| Zhao 2020 [59] | Neonatal ventricular cardiomyocytes | Neonatal rat pups | Purchased MWNTs—98% purity, 10–30 µm length, 10–20 nm diameter, carboxylic functionalisation. | Random and aligned electrospun fibres consisting of Bombyx mori silk fibroin, PEO, gum arabic and varying concentrations of CNTs. | Live/Dead assay, confocal IF of α-actinin, cTnI, Cx43 plus F-actin and DAPI, custom software analysis of CM alignment, ImageJ analysis of nuclear aspect ratio and IF staining. | Cell viability maintained at >90% at day 7 showing insignificant effect from CNTs, solvent-free method and the nanofibrous morphology of the scaffolds. Aligned scaffolds guided cell growth parallel to the fibre direction. Scaffolds containing CNTs increased cell elongation. Aligned scaffolds induced more rod-shaped CM morphology and anisotropic organization. Scaffold alignment was more dominant in this regard than CNT content. CNTs increased sarcomeric organization and expression of α-actinin and cTnI. Cx43 expression was increased in CNT scaffolds and with scaffold alignment. | Challenge remains of producing entirely interconnected networks of gap junctions in engineered cardiac tissues which will function normally when transplanted and not induce arrhythmia. | No further work suggested. |
| Alvarez-Primo 2019 [60] | AC16 human cardiomyocytes | Derived from the fusion of primary ventricular cardiomyocytes and fibroblasts | Purchased SWNTs—pristine, produced by HiPco process | SWNTs dissolved in three different surfactants—SDS, CTAB and PF108—were mixed with sodium alginate to produce hydrogel scaffolds. | Live/Dead assay | 80% cell viability was observed in scaffolds which used PF108, and no cells, alive or dead, were detected on the other scaffolds. | No limitations given. | Assess whether PEO, poly(pyrrole), poly(thiopene) or poly(aniline) could be used as dispersion agents for SWNTs in tissue engineering applications. Development of biodegradable SWNT-alginate hydrogels. |
| Lee 2019 [62] | Neonatal cardiomyocytes | 2-day old Sprague Dawley rat pups | Purchased MWNTs—95% purity, 30 ± 15 nm length, 5 ± 20 nm diameter, carboxylic functionalisation | CNTs, along with GO and rGO, were dispersed in GelMA to produce thin hydrogel films. | PicoGreen assay, confocal IF of vinculin, α-actinin, cTnI, Cx43 plus F-actin, CellProfiler, ImageJ, and MATLAB analysis of IF staining, video recording and MATLAB analysis of CM contractions, excitation threshold testing by eStim, whole cell patch clamp, pStim by blebbistatin, qRT-PCR of Cx43, α-actinin, cTnI, integrin subunits α1, α2, α3, α5, α7, and β1, melusin, dystrophin, β-actin, α-tubulin, RhoA, Rac1, PAK, CDC42, TAZ, vinculin, YAP and SRC. | CMs on CNT and rGO films displayed more classic CM morphology, with increased elongation and cell spreading. Surface chemistry of nanomaterials had a greater effect on cell morphology than their geometry. CNT scaffold maintained highest cell viability after 5 days in culture. CNT-GelMA suggested as optimal nanomaterial scaffold based on cell maturation, retention, and viability. CNT and rGO scaffolds showed increased expression of all IF stained proteins compared to GO. CMs on CNTs displayed markedly increased upstroke velocity and action potential duration. Based on action potential features, cells on CNTs more closely mimic ventricular CMs and GO cells more closely mimic atrial CMs, while rGO cells displayed intermediate characteristics. CMs on CNTs and rGO had lower excitation thresholds and could be externally paced after 1 day. CNTs showed increased expression of Cx43 and α-actinin compared to native myocardial tissue, however no difference in terms of cTnI expression. CMs on CNT scaffolds showed increased expression of integrin subunits α1, α2, α3 and β1 after 3 and 5 days of culture, while rGO CMs showed increased expression of α5, α7 and β1 after the same time periods. CMs are commonly known to use integrin heterodimers α1β1 and α2β1 to bind to cardiac ECM proteins. From qRT-PCR of cytoskeletal, Rho family, mechanotransduction and growth proteins, CMs on CNTs showed increased expression of proteins related to growth and mechanotransduction as well as melusin, RhoA and Rac1. | Fibroblast contamination in isolated cells may have contributed to DNA content in PicoGreen assay, therefore skewing results. | No further work suggested. |
| Roshanbinfar 2019 [63] | Human induced pluripotent stem cell (hiPSC)-derived cardiomyocytes | hiPSCs differentiated by CHIR-99021-IWR-1-endo protocol | Purchased MWNTs—carboxylic functionalisation modified to carbodihydrazide | Thermoresponsive pericardial tissue-derived hydrogel with CNTs dispersed at 0.5 wt% concentration. | Live/Dead assay, Kymograph and MUSCLEMOTION analysis of CM contractions, IF of cTnI, α-actinin and Cx43 with ImageJ analysis plus DAPI, calcium imaging with Fluo-4, pStim by isoproterenol and epinephrine. | CNTs showed similar viability to Matrigel. Kymography showed synchronous beating on all scaffolds but only CNTs didn’t show arrhythmia. MUSCLEMOTION analysis showed increased contraction amplitude, speed, and BPM, which was enhance but not pathologically by pStim. IF showed no change in cTnI, increase in Cx43 and increased sarcomere length from α-actinin staining. ImageJ analysis of cTnI IF showed improved unidirectional alignment on CNTs. Calcium imaging showed increased BPM, peak amplitude and faster excitation and relaxation. Long term repeat experiments showed no change in IF and pStim results. | No limitations given. | No further work suggested. |
| Vaithilingam 2019 [64] | hiPSC-derived cardiomyocytes | Adult male human skin biopsy | Purchased MWNTs—carboxylic functionalization | PETrA was combined a curing agent and 0.1 wt% CNTs and was 3D printed by 2PP to create aligned, ridged scaffolds. | Live/Dead assay, IF of α-actinin plus F-actin and Hoechst 33258, eStim, ImageJ analysis of IF staining. | IF showed increased alignment and length of sarcomeres. eStim was shown to negatively affect viability in a voltage-dependent manner but low voltage stimulation and 3D scaffold architecture improved myofibril organization. Alignment and aspect ratio were shown to be improved on 3D scaffolds. | Creation of nanoscale scaffold topography wasn’t possible with the ink containing CNTs. | No further work suggested. |
| Hou 2018 [65] | Embryonic atrial & ventricular cardiomyocytes | E11 stage White Leghorn chicken embryos | Synthesized SWNTs—produced by CVD into thin films | Nephila clavata silk fibres wrapped in aligned CNT sheets. | SEM, ePhys via scaffold, pStim by isoproterenol. | SEM showed tight contacts between hybrid fibres and CMs. ePhys via scaffolds revealed increased long-term health and maturity of cells based on action potential amplitude and frequency. | No limitations given. | No further work suggested. |
| Martinelli 2018 [66] | Neonatal ventricular cardiomyocytes | 0- to 1-day old Wistar rat pups | Purchased MWNTs—organic functional groups added using sarcosine and heptanal. | 3D porous PDMS scaffold, surface coated with CNTs. | Alamar Blue assay, EdU assay, confocal IF of α-actinin, cTnI and Cx43 plus Hoechst 33342, calcium imaging by Fluo-4. | Addition of CNTs to 3D-PDMS increased cell retention and viability. Based on IF, CMs in CNT scaffolds were “phenotypically different” from the control scaffold. IF of cTnI showed improved sarcomeric organisation. CNTs increased Cx43 expression and localization to gap junctions. CMs in CNT scaffolds showed increased beating rates with more rhythmic oscillations in intracellular calcium. 8% increase in CM proliferation on CNT scaffolds from controls with no change in proliferation of fibroblasts. | Possible cell ageing, or cytotoxicity, induced by MWNT-coated scaffolds. | No further work suggested. |
| Wang 2018 [67] | hiPSC-derived cardiomyocytes | Purchased from Cellular Dynamics | Purchased CNTs—20–30 nm diameter, 10–30 µm length (MWNTs based on diameter) | Onto a 3-well PDMS culture chamber, 1:5 CNT:PDMS strips were adhered across the culture membrane. On top of which a PDMS-fluorescent bead layer was spin coated. | Confocal IF of α-actinin and MYH7, calcium imaging by Fluo-5, pStim by isoproterenol, verapamil, omecamtiv mecarbil, ivabradine and E-4031, impedence spectroscopy and Poincaré plot analysis of CM contractions. | IF showed increased alignment which improved over time but there was no change in sarcomere length. Calcium imaging showed similar peak intensity, frequency, and duration to controls. Scaffold resistance was shown to increase over time as cells began to adhere and become electrically active. CMs were also shown to contract more regularly as variability between beats decreased. pStim showed all drugs affected CMs as expected, demonstrating the scaffolds’ effectiveness as a drug testing platform. | No limitations given. | No further work suggested. |
| Ahadian 2017 [68] | Neonatal cardiomyocytes | 1- to 2-day old Sprague Dawley rat pups | Purchased MWNTs—40–90 nm diameter, 10–20 µm length, organic functional groups added using nitric and sulphuric acid | PEGDM-124 polymer mesh scaffolds with CNT concentrations of 0.1, 0.5 and 1.5%, coated with 2% gelatine solution. | Live/Dead assay, confocal IF of α-actinin plus F-actin, eStim. | 0.1 and 0.5% CNT scaffolds began beating 2 days earlier than controls, and more rhythmically and synchronously. No difference seen from Live/Dead viability assay. 0.5% CNT scaffolds showed lower excitation threshold than 0% and 0.1%, which were comparable. Characteristic striation of CM cytoskeletal and contractile proteins was observed in all scaffolds. | CNTs cause increased opacity of polymer solutions which can interfere with polymerisation and mechanical properties of UV-cured materials. | Use of published eStim protocols could further improve CM maturity on CNT-containing scaffolds. |
| Peña 2017 [69] | Neonatal ventricular cardiomyocytes | 1- to 3-day old rat pups | Synthesized MWNTs—carboxylic functionalized | EDC-NHS-lysine cross-linked immobilised CNTs in a PHSU-PNIPAAm-lysine reverse thermal gel. | CM contractions by AFM, IF of α-actinin, vimentin, CD31 and Cx43 plus DAPI, EdU assay, calcium imaging by Fluo-4. | IF showed no CD31-positive cells on any scaffolds but reduced vimentin-positive fibroblasts and increased expression and improved localization of α-actinin and Cx43 in CMs. EdU showed CMs possess more proliferative phenotype after day 3. Calcium imaging and AFM showed increased and synchronised contractions from CMs on CNTs. | No limitations given. | Investigate the biocompatibility and in vivo applications of RTG-CNT scaffolds. |
| Ren 2017 [70] | Neonatal cardiomyocytes | 1- to 3-day old Sprague Dawley rat pups | Synthesized MWNTs—6–10 walls, approx. 10 nm diameter, ~200 µm length/Purchased MWNTs—10–30 nm dimeter, 10–30 µm length | Super aligned CNT sheets, produced by CVD, were deposited onto glass or PDMS. Randomly aligned CNT controls were spray deposited onto glass or PDMS. | TUNEL assay, confocal IF of α-actinin and Cx43 plus DAPI, eStim by pacemaker, ImagePro Plus analysis of CM contractility, whole cell patch clamp. | Decreased cell death on SA-CNTs. CMs on SA-CNTs showed increased alignment and elongation. eStim resulted in no changes to cell morphology or viability. Increased expression of Cx43 and improved localization to gap junctions was shown on SA-CNTs. Increased spontaneous and synchronised beating was seen on SA-CNTs, with a decrease in resting membrane potential and action potential duration and an in increase in AP amplitude. Variability in AP duration was also reduced between cells and between consecutive beats on SA-CNTs. | Short CNTs can penetrate the cell membrane and disrupt cell activity. | Investigate biodegradable alternatives to PDMS and the application of scaffolds in cardiac resynchronization therapy. |
| Roshanbinfar 2017 [71] | HL-1 cardiomyocytes | Murine cell line derived from AT-1 atrial cardiomyocyte tumour lineage | Purchased MWNTs—carboxylic functionalisation modified to carbodihydrazide | Thermoresponsive pericardial tissue-derived hydrogel with CNT concentration of 0.5 wt%. | Alamar Blue assay, Live/Dead assay, confocal IF of α-actinin and Cx43, calcium imaging by Fluo-4. | CMs on CNTs displayed a more proliferative phenotype and increased viability. IF showed increased Cx43 expression but no change in α-actinin. CMs on CNTs showed more synchronous, ‘directional’ beating with BPMs twice as high as controls. | No limitations given. | No further work suggested. |
| Sherrell 2017 [72] | HL-1 cardiomyocytes | Murine cell line derived from AT-1 atrial cardiomyocyte tumour lineage | Purchased SWNTs—1.5 nm average diameter/Purchased MWNTs—8 nm average diameter | EDC-NHS cross-linked chitosan-collagen hydrogel scaffolds with CNTs at 0.5 or 2 g/L. | Whole cell patch clamp. | SWNT scaffolds had improved mechanical properties compared to MWNTs as a result of increased, even dispersion throughout the hydrogel. Cells on SWNT scaffolds showed similar electrophysiological phenotype to fibronectin/gelatine-coated controls where cells displayed mostly atrial and few pacemaker-type action potentials. Increased in chitosan surfactant in SWNT scaffolds reduced spontaneous beating. Cells on MWNT scaffolds displayed decreased spontaneous beating and increase pacemaker-type action potentials. Ideal scaffold components identified as 2 g/L of SWNTs and 1% chitosan surfactant. | MWNT scaffolds where thought to decrease spontaneous beating due to blockage of potassium channels. | No further work suggested. |
| Sun 2017 (1) [73] | Neonatal ventricular cardiomyocytes | 1-day old Sprague Dawley rat pups | Purchased SWNTs—0.7–1.3 nm diameter, 5–20 µm length, carboxylic functionalisation | GelMA-CNT solutions at 0, 0.5, 1, and 2 mg/mL were dispensed into 24-well plates and cured using UV. | Live/Dead assay, confocal IF of α-actinin, cTnI, Cx43, N-cadherin, PKP2 and PG plus Hoechst 33258, ImageJ analysis of IF staining, video recording of CM beating, TEM, calcium imaging by Fluo-4, WB of β1-integrin, β-catenin, N-cadherin, p-FAK, FAK and RhoA. | High CM viability up to 100 ppm CNT concentration, any higher displayed cytotoxicity. IF showed increased elongation and alignment based on α-actinin with ID proteins all increasing over time. ImageJ analysis also showed increase sarcomere length but also increase in Z-line width. TEM showed increase in ID structures and same observations as ImageJ. Calcium imaging showed stronger synchronous and rhythmic contractions. WB showed β1-integrin increased while β-catenin was unchanged. Blocking β1-integrin caused a reduction in Cx43. Blocking FAK and RhoA also caused this, but further investigation revealed FAK modulates electrical junctions and RhoA regulates mechanical junctions between cells. | No limitations given | Investigate the underlying mechanisms of ID formation on CNTs and the downstream effects on adherens junctions and desmosomes. Also use human cells in scaffolds to optimise modulus for ID formation. |
| Sun 2017 (2) [74] | Neonatal ventricular cardiomyocytes | 1-day old Sprague Dawley rat pups | Purchased SWNTs—0.8–1.6 nm diameter, 5–30 µm length, 95% purity | Type I collagen-CNT solutions, at 0, 0.5, 1, and 2 mg/mL, were combined with DMEMα and FBS and then gelated at 37 °C. | Live/Dead assay, Alamar Blue assay, confocal IF of α-actinin, cTnI and Cx43 plus F-actin and DAPI, TEM, video recording of CM contractions, calcium imaging by Fluo-4, H&E staining. | Experiments revealed dose-dependent toxicity of CNTs based on viability, with 1 wt% being optimum. F-actin staining showed increased cell adhesion and elongation with thicker and longer actin filaments. IF showed an increase in α-actinin, cTnI and Cx43. TEM and H&E also showed increased elongation, alignment, and visibility of sarcomeric structures. Beating analysis showed CMs started to beat earlier and faster on CNTs. Calcium imaging confirmed stronger synchronous and rhythmic contractions on CNTs. | No limitations given. | No further work suggested. |
| Yu 2017 [76] | Neonatal cardiomyocytes | 1-day old Sprague Dawley rat pups | Purchased MWNTs—30 ± 15 nm diameter, 5–20 µm length, 95% purity, carboxylic functionalisation | Type I collagen solutions, in DMEM, with CNT concentrations at 0, 0.5, 1, 2, 5 and 10 wt% were gelated at 37 °C in moulds or well plates. | Video recording of CM contractions. | The number and scaffold coverage of rhythmically contractile CMs was increased on CNT scaffolds. | No limitations given. | No further work suggested. |
| Liu 2016 [77] | Neonatal cardiomyocytes | 1- to 2-day old Sprague Dawley rat pups | Purchased MWNTs—10–20 nm diameter, 10–20 µm length | Aligned blend and coaxial PELA:CNT electrospun fibres at 0, 1, 2, 3, 4, 5, and 6% CNT concentrations. | LDH release assay, SEM, IF of α-actinin and cTnI, ImageJ analysis of CM elongation and aspect ratio, WB for α-actinin and cTnI, custom software analysis of CM contractions. | 5% CNT scaffolds showed best cell viability, growth, adhesion, elongation and aspect ratio, highest expression of α-actinin and cTnI by IF and WB, and highest BPM. Coaxial fibres outperformed blend fibres in all parameters. While 5% was optimum, 6% CNT scaffolds began to show cytotoxic effects. | No limitations given. | No further work suggested. |
| Shin 2015 [78] | Neonatal ventricular cardiomyocytes | 2-day old Sprague Dawley rat pups | Synthesized MWNTs—~5 nm inner diameter, ~2 nm wall thickness | CNT forest electrodes were sandwiched between layers of PEG and CNT-GelMA hydrogels. | Confocal IF of α-actinin and Cx43, eStim, custom MATLAB analysis of CM contractions. | Increase in sarcomeric α-actinin organization and alignment but a homogenous distribution of Cx43 which showed variability with changes in CNT concentration and stiffness. CMs showed increased BPM on CNT scaffolds and eStim enabled control over synchronous beating. | No limitations given. | No further work suggested. |
| Sun 2015 [79] | Neonatal ventricular cardiomyocytes | 1-day old Sprague Dawley rat pups | Purchased SWNTs—0.7–1.2 nm diameter, 100–1000 µm length, 95% purity | Type 1 Collagen-CNT solutions, prepared at 0, 0.05, 0.1, 0.15 and 0.2 mg/mL, were deposited onto glass substrates and vacuum dried at 60 °C. | Live/Dead assay, Alamar Blue assay, cell retention by DAPI, qDNA assay, H&E staining, confocal IF of cTnI, α-actinin, Cx43 and N-cadherin plus F-actin and DAPI, TEM, calcium imaging by Fluo-4, WB of Cx43, N-cadherin, plakophilin2, plakoglobin, β-integrin, p-ERK, ERK, p-AKT, AKT, p-Src, p-FAK and ILK, RT-PCR of AP-1, c-fos, MEF-2c, NKX2.5, and GATA4. | Low CNT concentrations showed similar viability to controls, but higher concentrations displayed cytotoxicity. CNT scaffolds showed increase cell retention compared to controls. F-actin staining showed thicker and more spread out actin filaments with better cell adhesion. Increased DNA content across all substrates but no difference between CNTs and controls. H&E and IF staining both showed increased alignment and visibility of sarcomeres with IF showing an increase in TnI, Cx43, and NC. TEM displayed more compact and visible I, Z and H bands and an increase in ID structures. Calcium imaging showed stronger synchronous, rhythmic beating on CNTs. WB showed an increase in all ID proteins except β1-integrin. With an increased in p-FAK but no change in p-Src or ILK. Increase in p-FAK also lead to increases in p-ERK but no change in p-AKT. Blocking ERK lead to decrease in Cx43 while blocking AKT showed an increase as seen in normal culture. In addition, blocking β1-integrin also caused a decrease in Cx43 and N-cadherin but only on CNTs. RT-PCR showed increases in GATA4 and MEF-2c but no change in AP-1, c-fos or NKX2.5. | No limitations given. | No further work suggested. |
| Kharaziha 2014 [80] | Neonatal ventricular cardiomyocytes | 2-day old Sprague Dawley rat pups | Purchased MWNTs—30 ± 15 nm diameter, 5–20 µm length, 95% purity, carboxylic functionalisation | Aligned EDC-NHS cross-linked electrospun PG:CNT fibres with 0.05, 0.5 and 1.5% CNT concentrations. | Live/Dead assay, Alamar Blue assay, cell retention by DAPI, IF of α-actinin, Cx43 and cTnI plus DAPI, ImageJ analysis of IF staining, FFT analysis of cell alignment, custom MATLAB analysis of CM contractions, eStim. | 1.5% PG:CNT scaffolds showed the greatest improvement in cell viability, retention, alignment, and contractile function. eStim further improved CM contractile and electrical activity. | No limitations given. | Develop layered and vascularised structures from CNT scaffolds to improve nutrient delivery. |
| Pok 2014 [81] | Neonatal ventricular cardiomyocytes | 1- to 3-day old Sprague Dawley rat pups | Purchased SWNTs—0.98 ± 0.21 nm diameter, 0.1–4 µm length, >90% purity | Chitosan-gelatine hydrogel with 0, 33, 69, 175 ppm CNT concentrations. | Live/Dead assay, SEM, visual analysis of CM contractions, IF of α-actinin and Cx43 plus DAPI, action potential imaging by Di-8-ANEPPS. | Scaffolds containing less than 69 ppm CNTs had 80% viability, but higher concentration showed large drop in viability—indicating CNT toxicity is dose dependent. Analysis of CM BPM showed low CNT concentrations were similar to controls, but medium concentrations displayed BPM close to physiological BPM of rats. IF showed increase in α-actinin and slight increase in Cx43, which was confirmed by WB. Cx43 also showed improved localization to gap junctions. CMs on CNTs showed increased conduction velocity and action potential duration. | Mechanisms of CNT cytotoxicity is unknown and widely debated. | In vivo application of scaffolds in a rat disease model. |
| Tao 2014 [82] | Neonatal cardiomyocytes | 1-day old rat pups | Synthesized MWNTs—3–10 walls, 6–15 nm diameter, pristine modified to oxidized by acid vapour | SA-CNT yarns were pulled from aligned CNT arrays to produce thin films of adjacent yarns. 30 films were stacked, rotating alignment 90° each time, to produce scaffolds. CNTs were then oxidised using nitric acid vapour at 120 °C for 6 h. | Cell retention by Hoechst, visual analysis of cell adhesion and growth, SEM, calcium imaging by Fluo-4. | Higher cell count was observed on oxidised CNTs with increased adherence and growth. Conclusions from SEM were unclear. Calcium imaging showed no difference between CNT scaffolds and controls. | Oxidised CNT films had lower cell density than traditional culture dishes. | Oxidised CNTs provide a starting point for functionalisation to create scaffolds specialized for cell adherence. |
| Martinelli 2013 [84] | Neonatal ventricular cardiomyocytes | 1- to 3-day old rat pups | Purchased MWNTs—20–30 nm (parameter not given) | Isotropic CNT layer on glass substrates from 0.01 mg/mL dispersion in DMF, annealed at 350 °C in N₂ for 20 min. | SEM, IF of α-actinin and Cx43, WB of Cx43, SERCA2a and MYH7, qPCR for αMHC, βMHC, ANP, SERCA2a and skeletal actin, pStim by phenylephrine, whole cell patch clamp, calcium imaging by FURA2. | Increased expression of αMHC and SERCA2a, decreased expression of ANP and no change in βMHC or skA. Changes in SERCA2a and myosin heavy chains were also seen. Increased expression of Cx43 and improved localisation to gap junctions. Stimulation by phenylephrine showed CMs on CNTs did not exert the same hypertrophic response, qPCR revealed increased in αMHC and SERCA2a with a decrease in βMHC and skA but no change in ANP. Patch clamp showed decrease in resting membrane potential. Calcium imaging showed increase in more mature, sporadically contractile CMs. | No limitations given. | No further work suggested. |
| Shin 2013 [85] | Neonatal ventricular cardiomyocytes | 2-day old Sprague Dawley rat pups | Purchased MWNTs—30 ± 15 nm diameter, 5–20 µm length, 95% purity, carboxylic functionalisation | GelMA UV-cured hydrogel thin films made with 0, 1, 3, and 5 mg/mL CNT concentrations. | Live/Dead assay, MTS assay, qDNA assay, confocal IF for α-actinin, cTnI and Cx43 plus F-actin and DAPI, WB for α-actinin, cTnI and Cx43, SEM, custom software analysis of CM contractions, eStim, pStim by 1-heptanol and doxorubin. | CNT scaffolds showed increased cell retention and viability with limited proliferation, indicating CMs are growing but fibroblasts are not dividing. Improved alignment of whole cells and cytoskeletal organization. Increased expression of cTnI and increased visibility and organization of sarcomeric structures. An increase in α-actinin and TnI but only a small increase in Cx43 from WB, but IF indicates Cx43 is instead better localized to gap junctions. CMs also showed increased BPM and synchronization. CMs on CNTs showed a reduced threshold potential when undergoing eStim. CNT scaffolds exhibited a protective effect against pStim. SEM revealed CNTs encourage cell elongation and spreading of filopodia. | No limitations given. | Investigation into inhibitor resistance on high concentration CNT scaffolds. |
| Martinelli 2012 [86] | Neonatal ventricular cardiomyocytes | 1- to 3-day old rat pups | Purchased MWNTs—20–30 nm (parameter not given) | Isotropic CNT layer on glass substrates from 0.01 mg/mL dispersion in DMF, annealed at 350 °C in N₂ for 20 min. | TEM, Alamar Blue assay, IF of α-actinin plus DAPI, BrdU assay, flow cytometry and IF of pHH3, whole cell patch clamp. | Frequent and tight contacts existed between cells and CNTs with some CNTs appearing to penetrate membrane. Increased metabolic activity on CNTs after 36 hrs. IF of α-actinin revealed fewer α-actinin-negative fibroblasts on CNTs. BrdU, pHH3 and flow cytometry all confirmed CMs on CNTs retained a more proliferative phenotype. Patch clamp showed lower resting potential and shorter action potential duration on CNTs. | The precise effects of MWNTs are not understood. | Investigation into underlying effects with regards to genotype and phenotype, and into applications treatment of arrhythmia and conduction diseases. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scott, L.; Jurewicz, I.; Jeevaratnam, K.; Lewis, R. Carbon Nanotube-Based Scaffolds for Cardiac Tissue Engineering—Systematic Review and Narrative Synthesis. Bioengineering 2021, 8, 80. https://doi.org/10.3390/bioengineering8060080
Scott L, Jurewicz I, Jeevaratnam K, Lewis R. Carbon Nanotube-Based Scaffolds for Cardiac Tissue Engineering—Systematic Review and Narrative Synthesis. Bioengineering. 2021; 8(6):80. https://doi.org/10.3390/bioengineering8060080
Chicago/Turabian StyleScott, Louie, Izabela Jurewicz, Kamalan Jeevaratnam, and Rebecca Lewis. 2021. "Carbon Nanotube-Based Scaffolds for Cardiac Tissue Engineering—Systematic Review and Narrative Synthesis" Bioengineering 8, no. 6: 80. https://doi.org/10.3390/bioengineering8060080
APA StyleScott, L., Jurewicz, I., Jeevaratnam, K., & Lewis, R. (2021). Carbon Nanotube-Based Scaffolds for Cardiac Tissue Engineering—Systematic Review and Narrative Synthesis. Bioengineering, 8(6), 80. https://doi.org/10.3390/bioengineering8060080