
Tryptophan Production Maximization in a Fed-Batch Bioreactor with Modified E. coli Cells, by Optimizing Its Operating Policy Based on an Extended Structured Cell Kinetic Model


Abstract
1. Introduction
2. E. coli T5 Strain and the Experimental FBR
2.1. The Used E. coli Strain
2.2. Experimental Bioreactor and the Recorded Kinetic Data







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| The FBR Initial Conditions | ||
|---|---|---|
| Parameter | Nominal (Initial) Value | Obs. |
| Bioreactor type | DASGIP parallel bioreactor system, Eppendorf (completely automated) | [74] |
| Bioreactor mixing | Both mechanical and sparkling gas (O2) | [74] |
| Oxygen supply | Pure oxygen sparging | [74] |
| ) (g DW·L−1) | 0.16 Experimental data of Chen [74] (Figure 8c) | With the courtesy of Chen [74] |
| Batch time (tf ) | 3780 min (63 h) | |
| Cell content dilution rate (μ), (1/min) | 1.25 × 10−5–0.015 | Estimated 0.0017 [52] |
| ) | 0.015 L h−1 | Maintained quasi-constant |
| ) | 0.5 L (initial) | Variable, due to the continuous feeding of the FBR |
| 3330.5 mM | Maintained constant by Chen [74] | |
| at (t = 0) | 194.53 mM Experimental data of Chen [74] (Figure 7) | [74] |
| Temperature/pH | 37 °C/6.8 | [74] |
| )], and facilities | 3 L, automatic control of pH, DO, temperature | [74] |
| ) | 565.5 g DW·L cytosol−1 | [52] |
| Initial concentrations for the glycolytic cell species (in mM) | = 0.6003 = 0.2729 = 2.6729 = 2.6706 = 4.27 [AMDTP]total = 5.82 | Measured by Chassagnole et al. [52] |
| Initial concentrations for the TRP synthesis operon species (in μM) | (t = 0) = 0.01 (t = 0) = 3.32 (nM) (t = 0) = 0.01 (t = 0) = 928 (nM) | Measured by Bhartiya et al. [81] |
| (t = 0) = 0.164 | This paper; data of Chen [74] | |
| Species Mass Balance | Auxiliary Relationships and Estimated Rate Constants |
|---|---|
|
Glucose = control variables to be optimized; j = 1,…, (equal time-arcs) (t = 0) is given in (Table 1) for the nominal FBR of Chen [74] For the optimal FBR with adopted = 5, the feeding policy is (Footnote a): | = constant [33,86,87] (ii) results from solving the thermodynamic equilibrium relationship , i.e., (iii) μ = cell dilution rate (Table 1) (iv) The initial values of cell species concentrations are given in Table 1 (see also footnote (b)) (v) The lump of Figure 3 includes species belonging to the TCA cycle; there are no measurements on this lump, so it was excluded from data fitting (vi) The adopted value for by Maria [35] is = 1/43.63 (at QSS) [88]; was re-estimated from experimental data by Maria [35], resulting in = 0.467 (vii) See Table 3 for the flux expressions |
| Species inside the cell | |
| Liquid volume dynamics in Table 1; j = 1,…, (equal time-arcs) |
(viii) For the adopted = 5, the feeding policy is (see footnote (a)) |
| Biomass dynamics ; in (Table 1) | (ix) The biomass growth inhibition corresponds to a modified Contois model [85] The estimated rate constants by Maria [35] are = 1.05·10−4 (1/min·mM), = 10.19, = 1.8036·10−2 (1/min), = 7.334 × 10−2 |
| Reactions | Rate Expressions | Estimated Rate Constants (Units in mM, min) |
|---|---|---|
| GLC import system glc + pep → f6p + pyr pyr + atp → pep + adp + h glc + atp → f6p + adp +h | Modification for the T5 strain | = 1.1191 (1/min) = 3487.5 (mM) = 0 = 0 |
| f6p + atp → fdp + adp + h | = 1.0437 = 2 = 0.062028 (mM/min) = 6.16423 (mM) = 25 μM = 60 μM | |
| fdp + 2 adp (+ 2 nad + 2 p) ↔ 2 pep + 2 atp (+ 2 nadh + 2 h + 2 h2o) | = 4602.3 (1/min) = 31.917 (1/min) = 0.05 = 3 | |
| pep + adp + h → pyr + atp | = 1.331879 m = 4 = 0.1333655 (mM/min) = 1.146443 (mM) = 0.2 (mM) = 9.3 (mM) | |
| pyr → products (accoa, cit, succ, lac, etoh, ac, …) | = 693.3544 (1/min) = 395.525 (mM) = 2.6814 | |
| atp → adp +h | = 552.38 (1/min) | |
| 2 adp ↔ atp + amp | K = 1 | |
| (i) Termonia and Ross [86,87] indicated experimental evidence of a very fast reversible reaction catalyzed by AKase, with the equilibrium being quickly reached (ii) The k6 constant takes values according to the microorganism phenotype (related to the gene encoding the enzyme ATPase that catalyzes this reaction) (iii) = constant [86,87] (iv) results from solving the following thermodynamic equilibrium relationship: , i.e., . | ||
3. Bioprocess and Bioreactor Dynamic Model
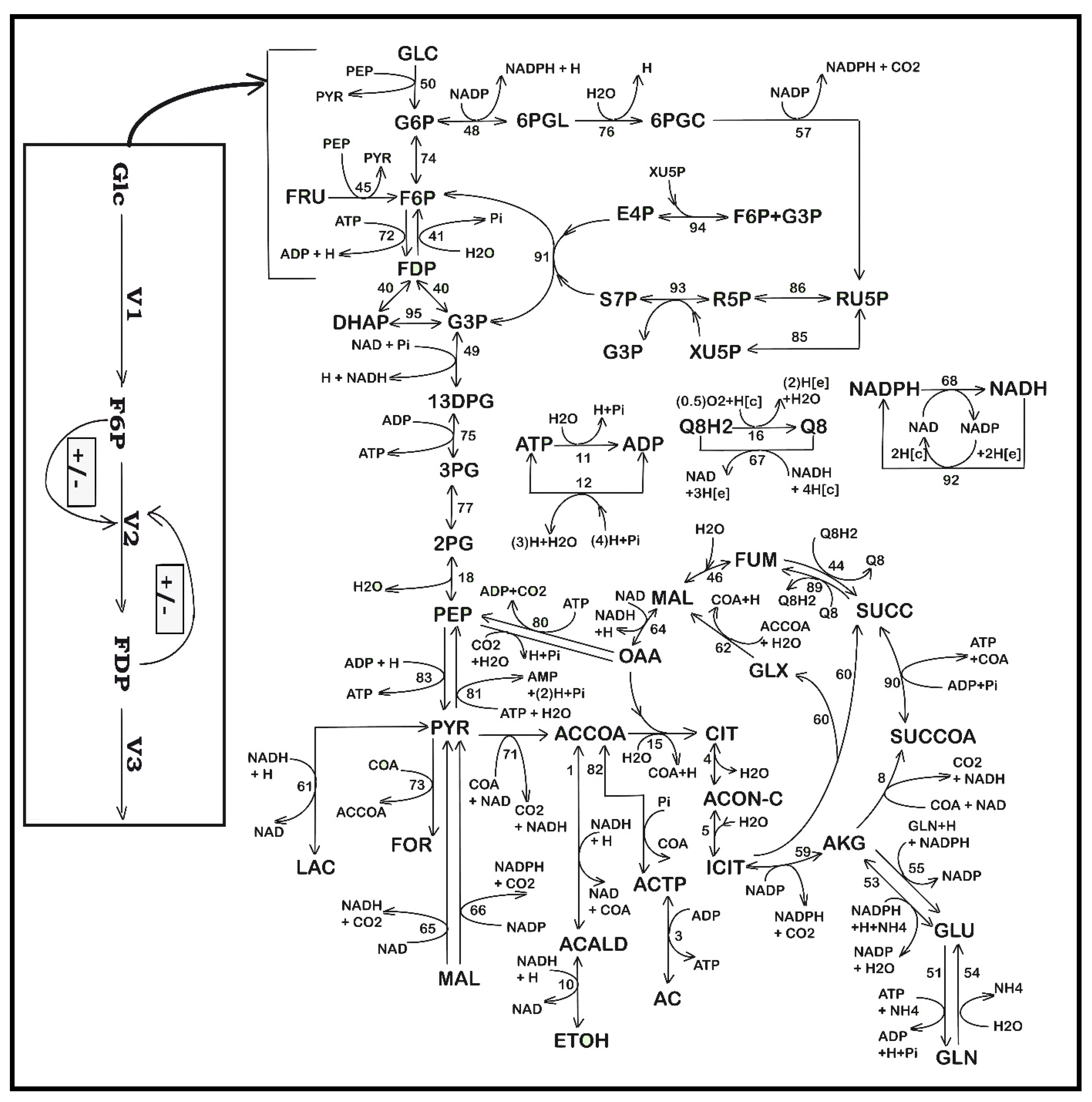
3.1. The Structured Hybrid Kinetic Model of Maria
3.1.1. The Biomass [X] Growth
3.1.2. The FBR Dynamic Model
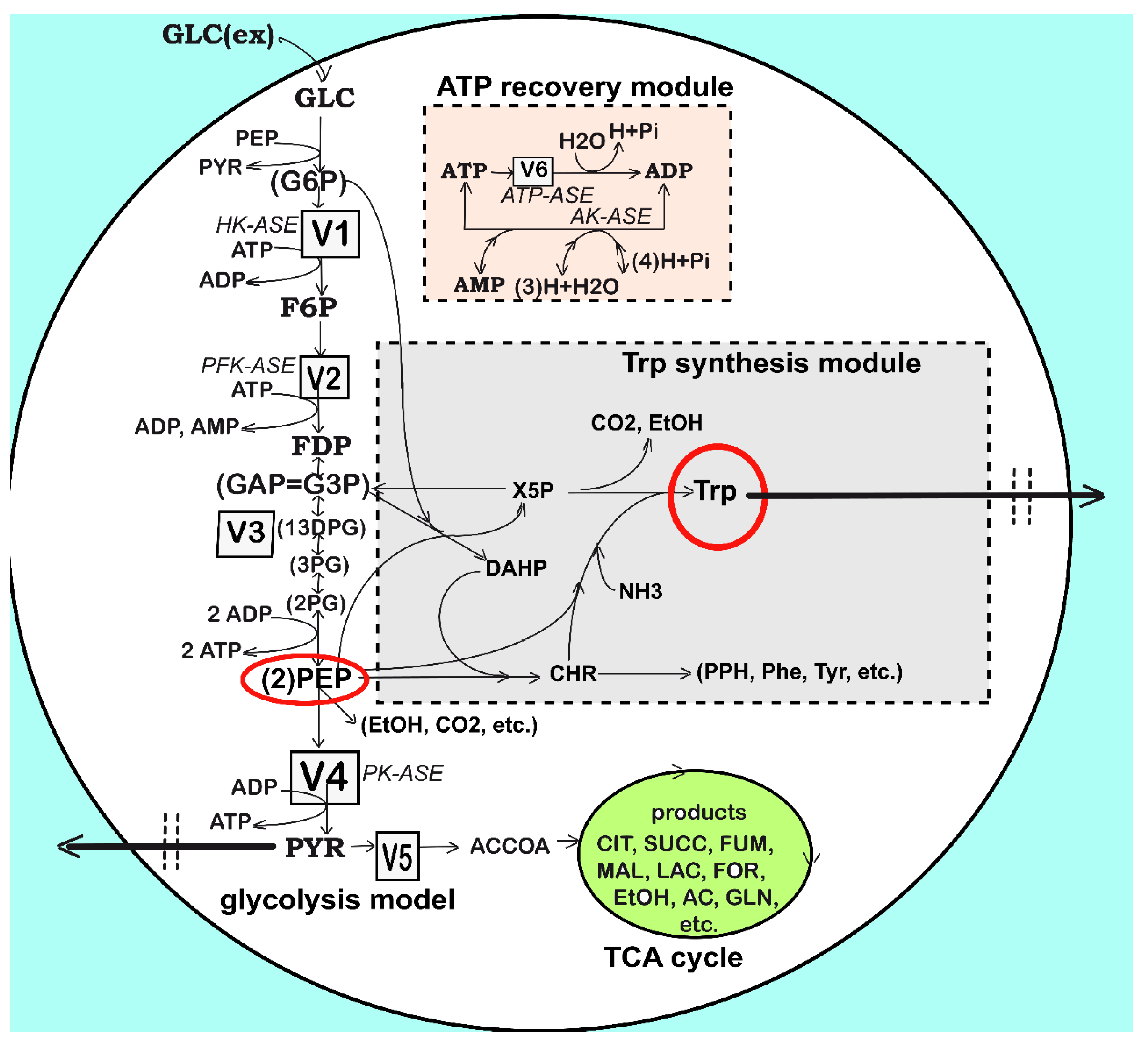
3.1.3. Module [a] Glycolysis and Module [b] ATP Recovery System
3.1.4. Module [c] TRP Synthesis
- i.
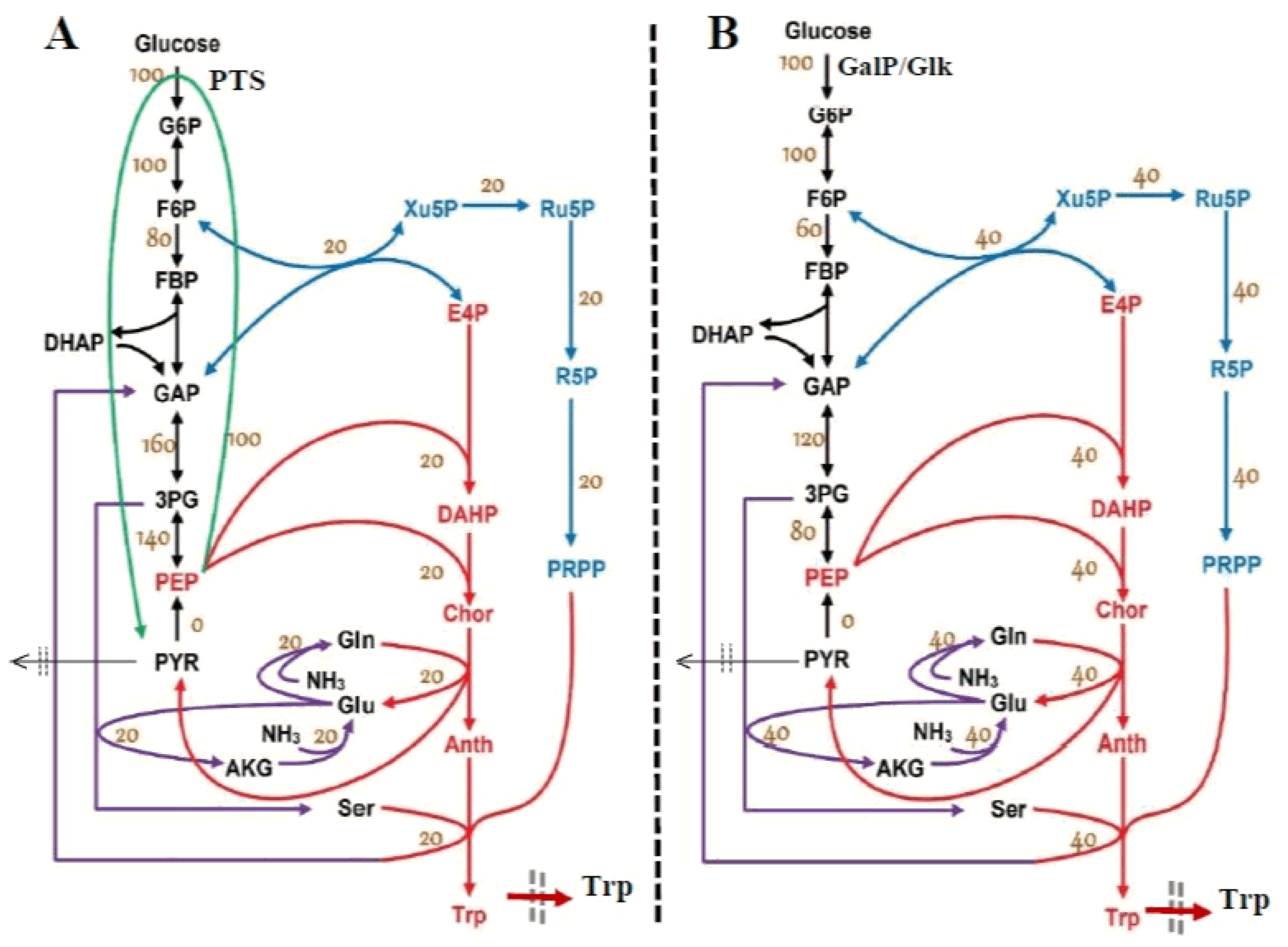
- An explicit connection of the TRP module to the glycolysis module [a] pathway was introduced through the PEP precursor sharing node (in Figure 3). Consequently, PEP is included as a substrate in the TRP mass balance (dcTRP/dt in Table 4), while the PEP consumption term is also considered in the PEP balance of the glycolysis model according to the recommended fluxes ratios of Stephanopoulos and Simpson [88], as a first guess (Table 2). Analysis of this model suggests that intensifying TRP synthesis clearly depends on the glycolysis intensity (average levels of glycolytic species) and its dynamics (QSS or oscillatory). In fact, as remarked by Li et al. [78] and by Chen and Zeng [76], the PEP precursor is the limiting factor for TRP synthesis. This is why intense efforts have been made to increase its production through glycolysis intensification [33,34]. This can be realized by optimizing the FBR operating policy (as in this paper) and/or by using (also in this paper) the modified E. coli T5 strain culture of Chen et al. [73] and Chen [74].
- ii.
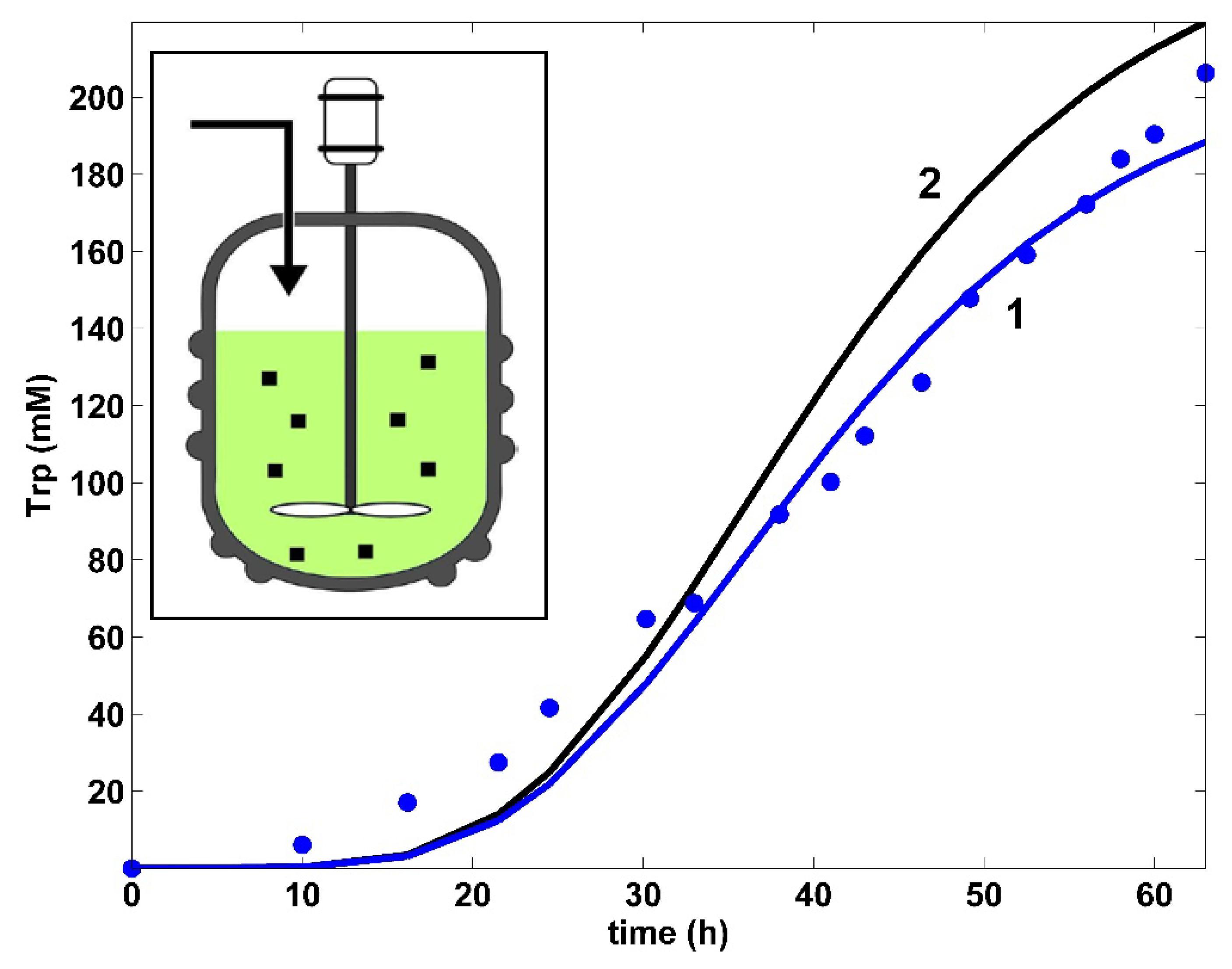
- The TRP synthesis model of Bhartiya et al. [81] (Table 4) includes two terms for the product inhibition, i.e., the C3 term (of allosteric-type) plus a Michaelis–Menten term. Our tests proved that these terms do not adequately fit the TRP experimental kinetic data of Figure 4. Accordingly, the product inhibition term in the TRP balance of Table 4 was replaced by the more adequate Contois-type model, considering a power-law inhibition of the first-order growing TRP at the denominator. Eventually, the rate constants of the TRP [c] kinetic module, the PEP consumption stoichiometry, and the rate constants of the other modules ([a], [b], and [X]) were re-estimated (refined) simultaneously using the whole (complete) hybrid FBR model, as well as all available experimental kinetic trajectories of the key-species offered by Chen [74] (Table 1, and Figure 4, Figure 5, Figure 6, Figure 7 and Figure 8). The initial guesses of the rate constants of the TRP module [c] were adopted from the literature.
- iii.
- The required PEP and GLC dynamic trajectories during estimation were transferred among the modules [a], [b], [c], [X] of the FBR kinetic model, all being available at this point.
- iv.
- In contrast to the literature, in the TRP balance of Table 4, an activation inhibition term was considered by bringing together the substrate (PEP) and the first key enzyme (anthranilate synthase, E) that trigger TRP synthesis [35]. Such an approach was proven to better fit the experimental data of Figure 4, i.e., , u = 1, …, n (where n = 17 denotes the number of experimental points) and to confer more flexibility to the derived model. The estimated negative g constant, of a small negative value, reflects the slight inhibition of TRP synthesis with the substrate PEP, as also suggested in the literature [35].
3.2. Rate Constant Estimation by Maria (2021)
3.3. Ways to Intensify the TRP Production in the FBR
- i.
- The GLC import system efficiency (V1 in Figure 3) is regulated and triggered by the external concentration of glucose and by the subsequent PEP and PYR synthesis (Table 2 and Table 3). The regular GLC uptake system, i.e., the PTS translocation system in the wild strain (of a complex reaction rate expression discussed by [35,48,52]) was replaced in the present studied E. coli T5 strain, as mentioned in Section 2, with a more efficient one (Figure 2B) able to accelerate the GLC uptake flux into the cell at least twofold [74]. Such a modified GLC import was modeled by simple Michaelis–Menten kinetics in the model of Table 3 by accounting for the well-known GLC substrate inhibition.
- ii.
- The quick import of GLC and its conversion to the precursor PEP require important amounts of regenerable ATP and a rapid enough ATP-to-ADP conversion rate, as well as its quick regeneration. The re-estimated rate constants of the kinetic module [b] (pink rectangle in Figure 3, and Section 3.1.3), concomitantly with those of the kinetic module [a] from the experimental data coming from the FBR operated with modified E. coli cells implicitly ensure the requirement that the A(MDT)P energy system is able to support the cell glycolysis (see V2, V4, and V6 expressions in Table 3 and the ATP mass balance in Table 2). On the other hand, limited A(MDT)P energy resources which exist in the cell slow down the GLC import if the ATP use/regeneration is not working fast enough [97]. Such an A(MDT)P resource is linked to the microorganism phenotype. Here, the total A(MDT)P was adopted (Table 1 and Table 3) at the average level recommended by Chassagnole et al. [52].
- iii.
- Additionally, due to the enzyme ATPase and AKase characteristics related to the bacteria genome and cell phenotype (Figure 3), a limited ATP conversion rate can sustain the glycolytic reactions, while the ATP recovery rate is limited by the enzymes participating in the A(MDT)P interconversion reactions (i.e., the K and k6 rate constants in the kinetic model of Table 3). This is why the k6 rate constant was re-estimated here to fit the experimental data, as suggested by Maria et al. [36,49].
- iv.
- At the same time, as glycolysis is a systemic process, with a complex regulatory structure, its dynamics (oscillatory, transient, or QSS) is also related to the rate constants of all involved reactions. Consequently, all these rate constants were considered in the final estimation step of the whole FBR hybrid kinetic model. Similarly, Silva and Yunes [98] found that glycolysis (QSS or oscillatory) is only possible if the external concentration of GLC and the maximum reaction rates controlled by the enzymes PFKase and GKase (which control the V1 and V2 reactions of Figure 3) are within specific intervals. Due to the same reason, the rate constants related to the GLC uptake system in the modified E. coli cell (modified V1 flux in Table 3) were re-estimated to match the experimental kinetic data.
- v.
- As a corollary of the issue (iv), Maria [33,34,36,37] determined the operating conditions leading to glycolytic oscillations or QSS by varying the external factor [GLC]ext and some internal factors such as the total [AMDTP] level and the k6 rate constant of Table 3. Such an investigation was not necessary here, because no oscillatory process was identified in the present operating case.
- vi.
- Simulations by Maria [33,35] revealed that the TRP synthesis efficiency is also strongly influenced by external factors, related to the FBR operating regime, namely, (a) the cell dilution (taken into account as “μ” in the approached hybrid kinetic model of Table 2, (b) the GLC concentration in the external (bulk) phase ( in Table 2), and (c) the optimal operating policy for the control variables. In this paper, such an operating policy will correspond to the time stepwise variation of the feed flow-rate ( in Equation (5)) and of the GLC feeding concentration ( in Equation (6)).
4. Fed-Batch Bioreactor Optimization Problem
4.1. Preliminary Considerations
4.2. Formulation of the Optimization Problem
4.2.1. Selection of the FBR Control Variables
- (a)
- The substrate (j = 1, …, ) whose concentration plays a major role in the cell glycolysis and TRP production;
- (b)
- The liquid feed flow rate (j = 1, …, ), with a GLC solution directly linked to the GLC feeding, responsible for the reactor content dilution.
4.2.2. Objective Function (Ω) Choice
4.2.3. Optimization Problem Constraints
- (a)
- (b)
- (c)
- To limit the excessive consumption of substrate and to prevent the hydrodynamic stress due to the limited reactor volume, feasible searching ranges were imposed on the control/decision variables, i.e.,[GLC]inlet,min = 1000 (mM) ≤ [GLC]inlet,j ≤ [GLC]inlet,max = 4500 (mM),
FL,min = 0.01 (L/h) ≤ FL,j; FL,0 ≤ FL,max = 0.04(L/h); - (d)
- Physical meaning of searching variables:
- (e)
- Physical meaning of state variables:
- (f)
- Limit the maximum cell resources in AMDTP:[ATP] (t) < Total [AMDTP], with [ATP] (t) obtained from solving the FBR model in Equations (1)–(6).
4.2.4. and Operating Alternatives Choice
4.2.5. The Used Numerical Solvers
4.2.6. The Problem Solution Particularities
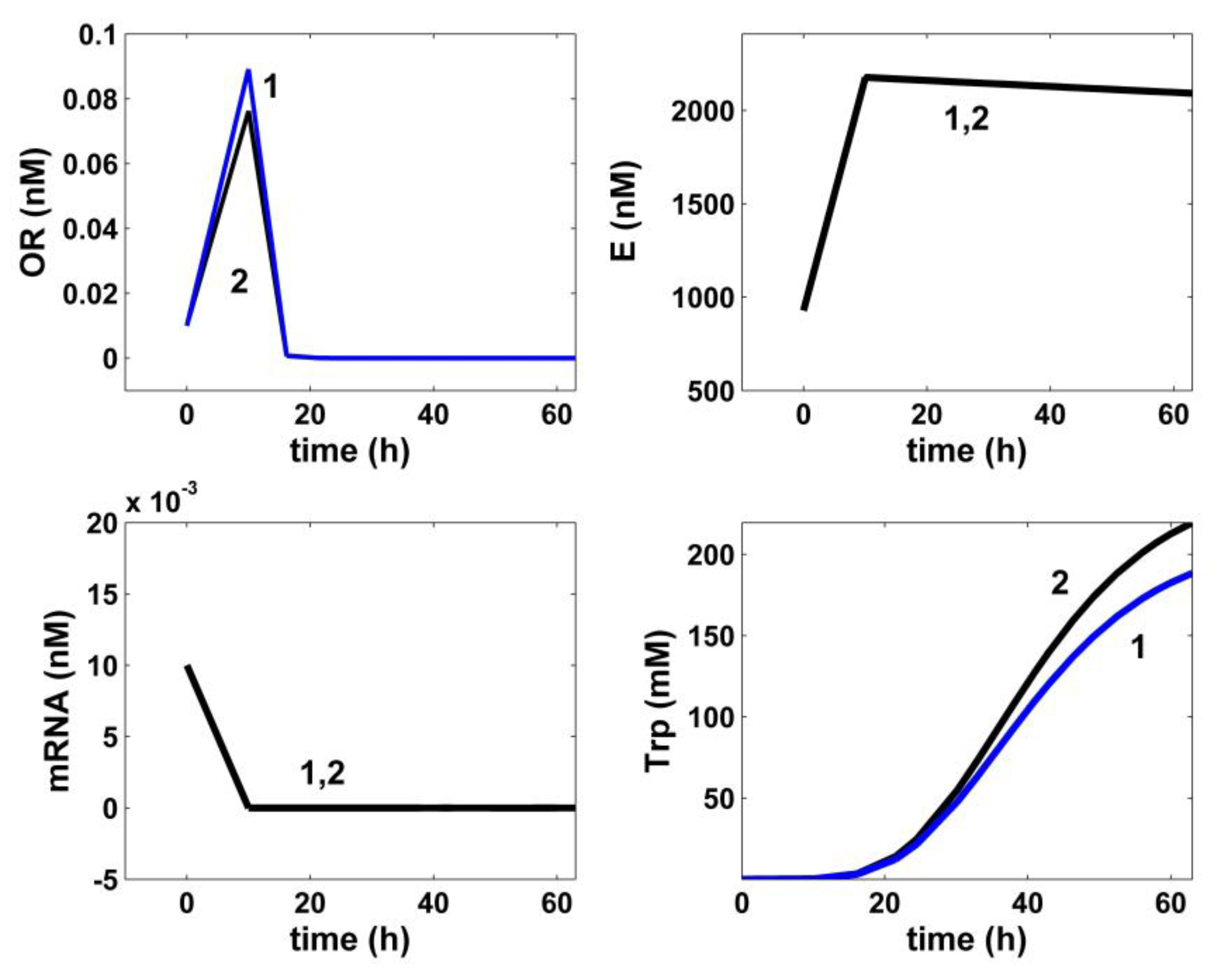
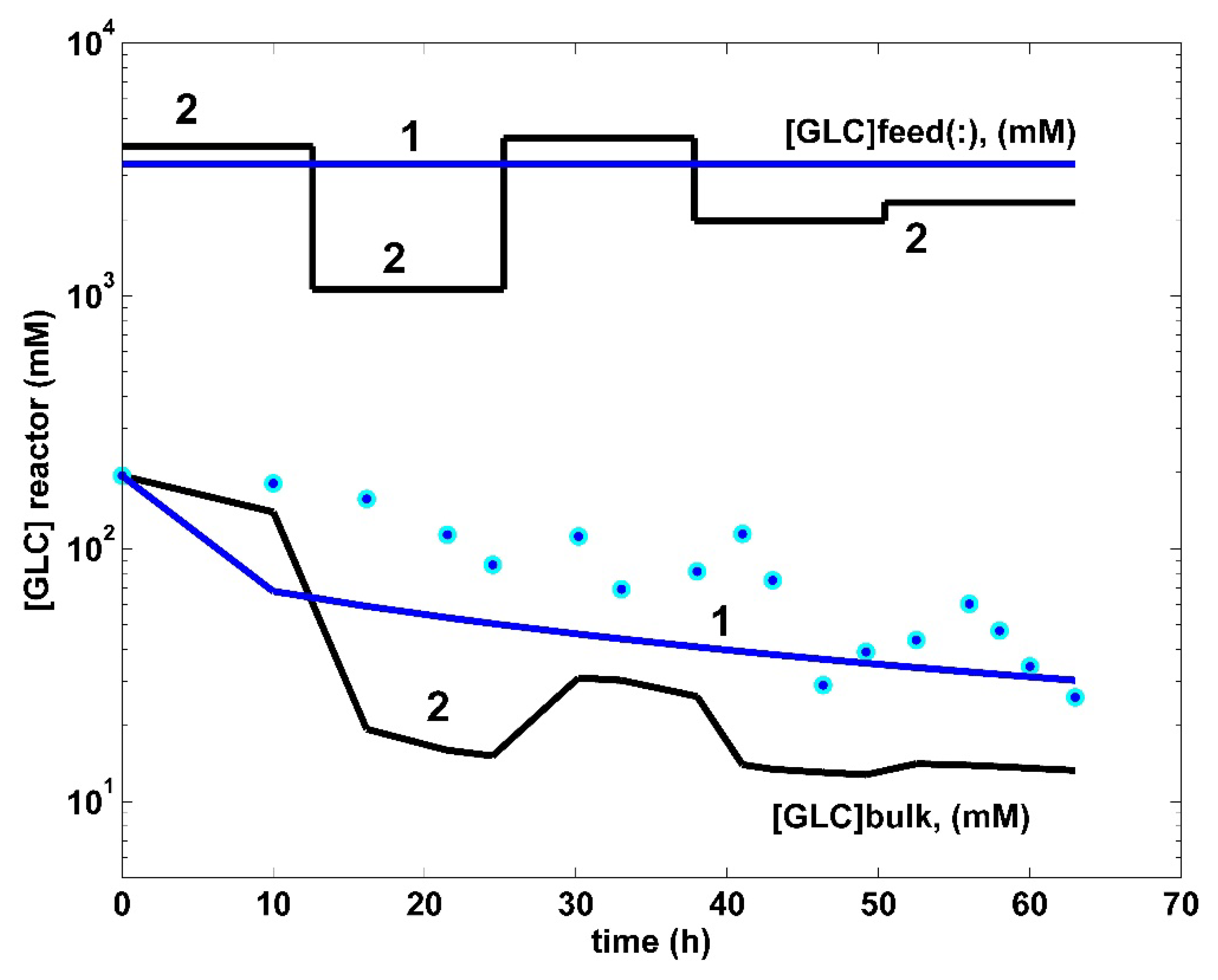
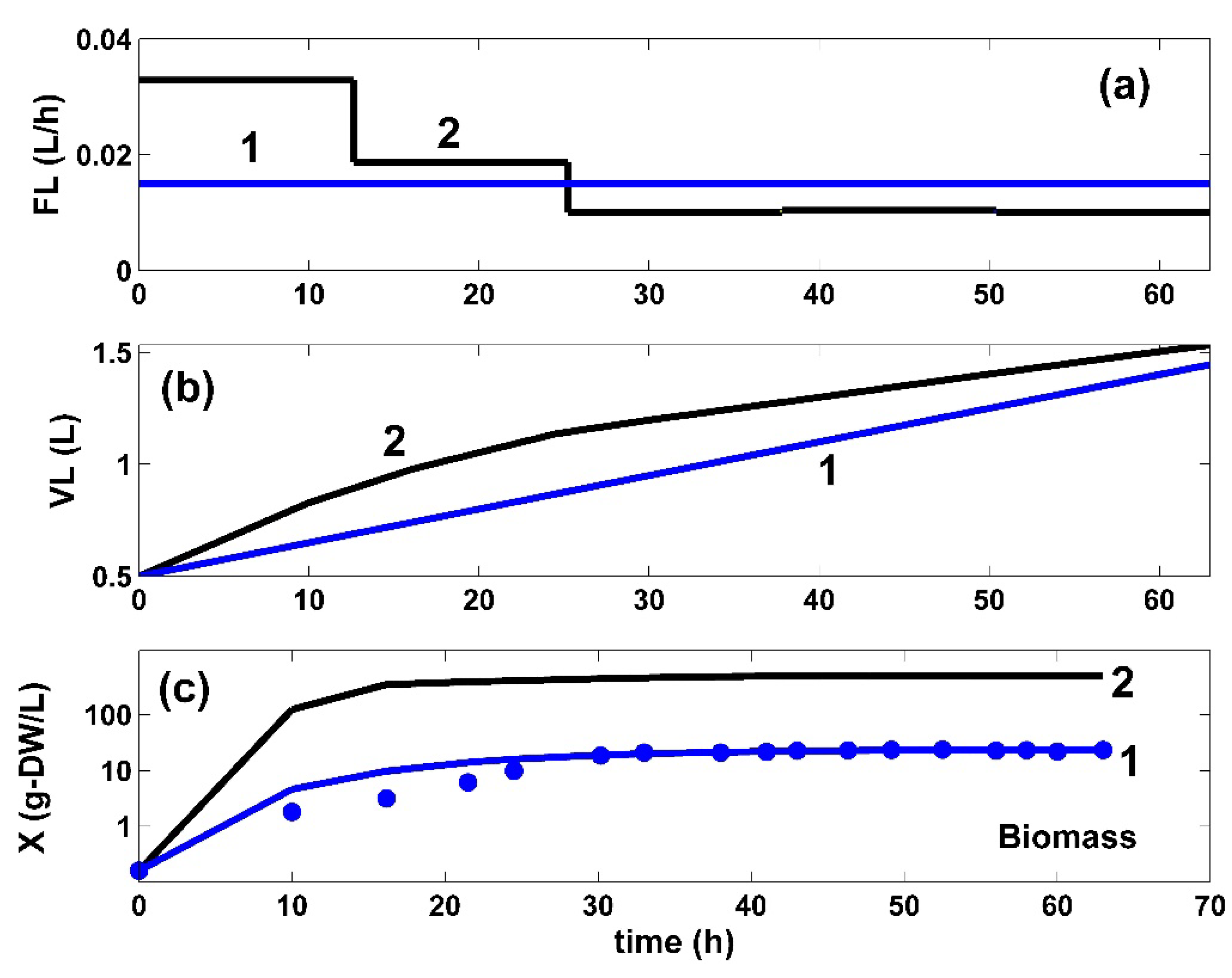
5. Optimization Results and Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Nomenclature
| Species (i) concentration | |
| Biomass concentration | |
| Glucose feeding solution concentration | |
| Glucose feeding solution concentration over the time-arc “J” | |
| Initial glucose concentration in the bioreactor | |
| Glucose concentration in the bulk phase | |
| Liquid feed flow rate in the bioreactor | |
| k, , , K, n, , , , , , , , , , , g,, , , , , , , etc.- | Reaction rates and/or equilibrium constants of the kinetic model |
| Species (i) reaction rate | |
| , | Time, batch time |
| Metabolic fluxes in the glycolysis (Table 2 and Table 3, Figure 3) | |
| Liquid volume in the bioreactor | |
| Stoichiometric coefficients | |
| Greeks | |
| α, β, γ, δ | Reaction rate constants |
| μ | Cell content dilution rate, that is , where denotes the cell cycle |
| Ω | FBR optimization objective function, Equation (10) |
| Biomass density | |
| Subscripts | |
| 0,o | Initial |
| cell | Referring to the cell (inside) |
| ext | External to cell (i.e., in the bulk phase) |
| f | Final |
| inlet | In the feed |
| x | Biomass |
| Abbreviations | |
| 13dpg, pgp | 1,3-Diphosphoglycerate |
| 3pg | 3-Phosphoglycerate |
| 2pg | 2-Phosphoglycerate |
| AA | Amino acid |
| Accoa, acetyl-CoA | Acetyl-coenzyme A |
| AC | acetate |
| ADP, adp | Adenosine diphosphate |
| AK-ase | Adenylate kinase |
| ALE | Adaptive laboratory evolution |
| AMP, amp | Adenosine monophosphate |
| ATP, atp | Adenosine triphosphate |
| ATP-ase | ATP monophosphatase |
| CCM | Central carbon metabolism |
| CIT | Citrate |
| CSTR | Continuously stirred tank reactor |
| DO | Dissolved oxygen |
| DW | Dry mass |
| E | Enzyme anthranilate synthase in TRP synthesis model |
| ETOH | Ethanol |
| ext | External to the cell (i.e., in the bulk phase) |
| FBR | Fed-batch bioreactor |
| FDP, fdp | Fructose-1,6-biphosphate |
| F6P, f6p | Fructose-6-phosphate |
| GalP/Glk | Galactose permease/glucokinase |
| G3P, g3p, GAP, gap, 3PG, 3pg | Glyceraldehyde-3-phosphate |
| 2PG, 2pg | 2-Phosphoglycerate |
| G6P, g6p | Glucose-6-phosphate |
| GLC, glc | Glucose |
| Glc(ex), [GLC]ext | Glucose in the environment (bulk phase) |
| GMO | Genetically modified microorganisms |
| GRC | Genetic regulatory circuits |
| HK-ase | Hexokinase |
| JWS | Silicon Cell project of Olivier and Snoep [55] |
| LAC, lac | Lactate |
| Max (x) | Maxim of (x) |
| MMA | The adaptive random optimization algorithm of Maria [93]; Mihail and Maria [99] |
| mRNA | Tryptophan messenger ribonucleic acid during its encoding gene dynamic transcription and translation |
| NAD(P)H | Nicotinamide adenine dinucleotide (phosphate) reduced |
| NLP | Nonlinear programming |
| ODE | Ordinary differential equations set |
| OR | The complex between O and R (aporepressor of the TRP gene) |
| OT | The total TRP operon |
| P, Pi | Phosphoric acid |
| PEP, pep | Phosphoenolpyruvate |
| 13DPG=PGP | 1,3-Diphosphoglycerate |
| PFK-ase | Phosphofructokinase |
| PK-ase | Pyruvate kinase |
| PTS | Phosphotransferase or the phosphoenolpyruvate–glucose phosphotransferase system |
| PYR, pyr | Pyruvate |
| QSS | Quasi-steady state |
| R5P | Ribose 5-phosphate |
| mRNA | Messenger ribonucleic acid |
| SUCC, suc | Succinate |
| TCA, tca | Tricarboxylic acid cycle |
| TF | Gene expression transcription factors |
| TRP, Trp, trp | Tryptophan |
| X | Biomass |
| Wt. | Weight |
| [x] | Concentration of species x |
References
- Ghose, T.K.; Fiechter, A.; Blakebrough, N. Advances in Biochemical Engineering; Springer: Berlin, Germany, 1977; Volume 7–10. [Google Scholar]
- Moser, A. Bioprocess Technology—Kinetics and Reactors; Springer: Berlin, Germany, 1988. [Google Scholar]
- Maria, G. Model-based optimization of a fed-batch bioreactor for mAb production using a hybridoma cell culture. Molecules 2020, 25, 5648. [Google Scholar] [CrossRef] [PubMed]
- Maria, G.; Peptănaru, I.M. Model-based optimization of mannitol production by using a sequence of batch reactors for a coupled bi-enzymatic process—A dynamic approach. Dynamics 2021, 1, 8. [Google Scholar] [CrossRef]
- Liese, A.; Seelbach, K.; Wandrey, C. Industrial Biotransformations; Wiley-VCH: Weinheim, Germany, 2006. [Google Scholar]
- Maria, G. Enzymatic reactor selection and derivation of the optimal operation policy, by using a model-based modular simulation platform. Comput. Chem. Eng. 2012, 36, 325–341. [Google Scholar] [CrossRef]
- Buchholz, K.; Hempel, D.C. From gene to product (editorial). Eng. Life Sci. 2006, 6, 437. [Google Scholar] [CrossRef]
- Hempel, D.C. Development of biotechnological processes by integrating genetic and engineering methods. Eng. Life Sci. 2006, 6, 443–447. [Google Scholar] [CrossRef]
- Chaudhuri, J.; Al-Rubeai, M. (Eds.) Bioreactors for Tissue Engineering Principles, Design and Operation; Springer: Berlin, Germany, 2005. [Google Scholar]
- Nedovic, V.; Willaert, R. Applications of Cell Immobilisation Technology; Springer: Amsterdam, The Netherlands, 2005. [Google Scholar]
- DiBiasio, D. Introduction to the control of biological reactors. In Chemical Engineering Problems in Biotechnology; Shuler, M.L., Ed.; AIChE: New York, NY, USA, 1989; Volume 1. [Google Scholar]
- Abel, O.; Marquardt, W. Scenario-integrated on-line optimisation of batch reactors. J. Process Control. 2003, 13, 703–715. [Google Scholar] [CrossRef]
- Lee, J.; Lee, K.S.; Lee, J.H.; Park, S. An on-line batch span minimization and quality control strategy for batch and semi-batch processes. Control Eng. Pract. 2001, 9, 901–909. [Google Scholar] [CrossRef]
- Ruppen, D.; Bonvin, D.; Rippin, D.W.T. Implementation of adaptive optimal operation for a semi-batch reaction system. Comput. Chem. Eng. 1998, 22, 185–199. [Google Scholar] [CrossRef]
- Bonvin, D. Optimal operation of batch reactors—A personal view. J. Process Control. 1998, 8, 355–368. [Google Scholar] [CrossRef]
- Rao, M.; Qiu, H. Process Control Engineering: A Textbook for Chemical, Mechanical and Electrical Engineers; Gordon and Breach Science Publ.: Amsterdam, The Netherlands, 1993. [Google Scholar]
- Reuss, M. Computer control of bioreactors present limits and challenges for the future. In Proceedings of the 3rd International Conference on Chemistry Process Control—CPCIII; Morari, M., McAvoy, T.J., Eds.; Elsevier: Amsterdam, The Netherlands, 1986; pp. 12–17. [Google Scholar]
- Agrawal, P.; Koshy, G.; Ramseier, M. An algorithm for operating a fed-batch fermentator at optimum specific-growth rate. Biotechnol. Bioeng. 1989, 33, 115–125. [Google Scholar] [CrossRef]
- Banga, J.R.; Alonso, A.A.; Singh, P.R. Stochastic optimal control of fed-batch bioreactors. In Proceedings of the AIChE Annual Meeting, San Francisco, CA, USA, 13–18 November 1994. [Google Scholar]
- Doran, P.M. Bioprocess Engineering Principles; Elsevier: Amsterdam, The Netherlands, 1995. [Google Scholar]
- Smets, I.Y.; Claes, J.E.; November, E.J.; Bastin, G.P.; Van Impe, J.F. Optimal adaptive control of (bio)chemical reactors: Past, present and future. J. Process Control 2004, 14, 795–805. [Google Scholar] [CrossRef]
- Sarkar, D.; Modak, J.M. Pareto-optimal solutions for multi-objective optimization of fed-batch bioreactors using nondominated sorting genetic algorithm. Chem. Eng. Sci. 2005, 60, 481–492. [Google Scholar] [CrossRef]
- Henson, M.A. Model-based control of biochemical reactors. In The Control Handbook, 2nd ed.; Levine, W., Ed.; Taylor and Francis: New York, NY, USA, 2010. [Google Scholar]
- Henson, M.A.; Muller, D.; Reuss, M. Combined metabolic and cell population modelling for yeast bioreactor control. In Proceedings of the IFAC Symposium on Advanced Control of Chemical Processes; Allgöwer, F., Gao, F., Eds.; Hong Kong University of Science and Technology: Hong Kong, China, 2009; pp. 11–14. [Google Scholar]
- Bodizs, L.; Titica, M.; Faria, N.; Srinivasan, B.; Dochain, D.; Bonvin, D. Oxygen control for an industrial pilot-scale fed-batch filamentous fungal fermentation. J. Process Control 2007, 17, 595–606. [Google Scholar] [CrossRef][Green Version]
- Ashoori, A.; Moshiri, B.; Khaki-Sedigh, A.; Bakhtiari, M.R. Optimal control of a nonlinear fed-batch fermentation process using model predictive approach. J. Process Control 2009, 19, 1162–1173. [Google Scholar] [CrossRef]
- Roeva, O.; Pencheva, T.; Tzonkov, S.; Arndt, M.; Hitzmann, B.; Kleist, S.; Miksch, G.; Friehs, K.; Flaschel, E. Multiple model approach to modelling of Escherichia coli fed-batch cultivation extracellular production of bacterial phytase. J. Biotechnol. 2007, 10, 592–603. [Google Scholar] [CrossRef][Green Version]
- Maria, G.; Luta, I. Structured cell simulator coupled with a fluidized bed bioreactor model to predict the adaptive mercury uptake by E. coli cells. Comput. Chem. Eng. 2013, 58, 98–115. [Google Scholar] [CrossRef]
- Roubos, J.A. Bioprocess Modeling and Optimization—Fed-Batch Clavulanic acid Production by Streptomyces clavuligerus. Ph.D. Thesis, TU Delft, Delft, The Netherland, 2002. [Google Scholar]
- Zak, D.E.; Vadigepalli, R.; Gonye, G.E.; Doyle, F.J., III; Schwaber, J.S.; Ogunnaike, B.A. Unconventional systems analysis problems in molecular biology: A case study in gene regulatory network modelling. Comput. Chem. Eng. 2005, 29, 547–563. [Google Scholar] [CrossRef]
- Maria, G. Deterministic Modelling Approach of Metabolic Processes in Living Cells—A Still Powerful Tool for Representing the Metabolic Process Dynamics; Juniper Publisher: Irvine, CA, USA, 2017; ISBN 978-1-946628-07-7. Available online: https://juniperpublishers.com/ebooks/Deterministic%20Modelling%20Approach%20of%20Metabolic%20Processes%20in%20Living%20Cells%20-%20A%20Still%20Powerful%20Tool%20for%20Representing%20the%20Metabolic%20Processes%20Dynamics.pdf (accessed on 22 October 2021).
- Maria, G. In-Silico Design of Genetic Modified Micro-Organisms (GMO) of Industrial Use, by Using Systems Biology and (Bio)Chemical Engineering Tools; Juniper Publisher: Irvine, CA, USA, 2018; ISBN 978-1-946628-12-1. Available online: https://juniperpublishers.com/ebooks/In-Silico%20Design%20of%20Genetic%20Modified%20Micro-Organisms%20(GMO)%20of%20Industrial%20Use,%20by%20Using%20Systems%20Biology%20and.pdf (accessed on 22 October 2021).
- Maria, G. In-silico determination of some conditions leading to glycolytic oscillations and their interference with some other processes in E. coli cells. Front. Chem. 2020, 8, 526679–526693. [Google Scholar] [CrossRef]
- Maria, G.; Mihalachi, M.; Gijiu, C.L. In silico optimization of a bioreactor with an E. coli culture for tryptophan production by using a structured model coupling the oscillating glycolysis and tryptophan synthesis. Chem. Eng. Res. Des. 2018, 135, 207–221. [Google Scholar] [CrossRef]
- Maria, G. A CCM-based modular and hybrid kinetic model to simulate the tryptophan synthesis in a fed-batch bioreactor using modified E. coli cells. Comput. Chem. Eng. 2021, 133, 106628–106635. [Google Scholar] [CrossRef]
- Maria, G.; Mihalachi, M.; Gijiu, C.L. Model-based identification of some conditions leading to glycolytic oscillations in E. coli cells. Chem. Biochem. Eng. Q. 2018, 32, 523–533. [Google Scholar] [CrossRef]
- Maria, G.; Gijiu, C.L.; Maria, C.; Tociu, C. Interference of the oscillating glycolysis with the oscillating tryptophan synthesis in the E. coli cells. Comput. Chem. Eng. 2018, 108, 395–407. [Google Scholar] [CrossRef]
- Maria, G.; Xu, Z.; Sun, J. Multi-objective MINLP optimization used to identify theoretical gene knockout strategies for E. coli cell. Chem. Biochem. Eng. Q. 2011, 25, 403–424. Available online: http://silverstripe.fkit.hr/cabeq/past-issues/issue/12 (accessed on 22 October 2021).
- Nielsen, J.; Villadsen, J. Bioreaction Engineering Principles; Springer: Boston, MA, USA, 1994. [Google Scholar]
- Maria, G. An Adaptive Strategy for Solving Kinetic Model Concomitant Estimation-Reduction Problems. Can. J. Chem. Eng. 1989, 67, 825–832. [Google Scholar] [CrossRef]
- Maria, G. A review of algorithms and trends in kinetic model identification for chemical and biochemical systems. Chem. Biochem. Eng. Q. 2004, 18, 195–222. Available online: http://silverstripe.fkit.hr/cabeq/past-issues/issue/41 (accessed on 22 October 2021).
- Edwards, K.; Edgar, T.F.; Manousiouthakis, V.I. Kinetic model reduction using genetic algorithms. Comput. Chem. Eng. 1998, 22, 239–246. [Google Scholar] [CrossRef]
- Gerdtzen, Z.P.; Daoutidis, P.; Hu, W.S. Non-linear reduction for kinetic models of metabolic reaction networks. Metab. Eng. 2004, 6, 140–154. [Google Scholar] [CrossRef]
- Martinez, E.C.; Beltramini, L.J. Lumping upon time-scales: Modeling upon topological factors. Chem. Eng. Sci. 1990, 45, 2103–2108. [Google Scholar] [CrossRef]
- Maria, G. Relations between apparent and intrinsic kinetics of programmable drug release in human plasma. Chem. Eng. Sci. 2005, 60, 1709–1723. [Google Scholar] [CrossRef]
- Eshtewy, N.A.; Scholz, L. Model Reduction for Kinetic Models of Biological Systems. Symmetry 2020, 12, 863. [Google Scholar] [CrossRef]
- Dorka, P. Modelling Batch and Fed-Batch Mammalian Cell Cultures for Optimizing MAb Productivity. Master’s Thesis, University of Waterloo, Waterloo, ON, Canada, 2007. [Google Scholar]
- Maria, G. In silico derivation of a reduced kinetic model for stationary or oscillating glycolysis in E. coli bacterium. Chem. Biochem. Eng. Q. 2014, 28, 509–529. [Google Scholar] [CrossRef]
- Maria, G.; Mihalachi, M.; Gijiu, C.L. Chemical engineering tools applied to simulate some conditions producing glycolytic oscillations in E. coli cells. U.P.B. Sci. Bull. Ser. B Chem. 2018, 80, 27–38. Available online: https://www.scientificbulletin.upb.ro/rev_docs_arhiva/full6c1_852902.pdf (accessed on 22 October 2021).
- Maria, G. A Review of Some Novel Concepts Applied to Modular Modelling of Genetic Regulatory Circuits; Juniper Publisher: Irvine, CA, USA, 2017; ISBN 978-1-946628-03-9. Available online: https://juniperpublishers.com/ebooks/A%20Review%20of%20Some%20Novel%20Concepts%20Applied%20to%20Modular%20Modelling%20of%20Genetic%20Regulatory%20Circuits.pdf (accessed on 22 October 2021).
- Edwards, J.S.; Palsson, B.O. The Escherichia coli MG1655 in silico metabolic genotype: Its definition, characteristics, and capabilities. Proc. Natl. Acad. Sci. USA 2000, 97, 5528–5533. [Google Scholar] [CrossRef]
- Chassagnole, C.; Noisommit-Rizzi, N.; Schmid, J.W.; Mauch, K.; Reuss, M. Dynamic modeling of the central carbon metabolism of Escherichia Coli. Biotechnol. Bioeng. 2002, 79, 53–73. [Google Scholar] [CrossRef]
- Usuda, Y.; Nishio, Y.; Iwatani, S.; Van Dien, S.J.; Imaizumi, A.; Shimbo, K.; Kageyama, N.; Iwahata, D.; Miyano, H.; Matsui, K. Dynamic modeling of Escherichia coli metabolic and regulatory systems for amino-acid production. J. Biotechnol. 2010, 147, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Kadir, T.A.A.; Mannan, A.A.; Kierzek, A.M.; McFadden, J.; Shimizu, K. Modeling and simulation of the main metabolism in Escherichia coli and its several single-gene knockout mutants with experimental verification. Microb. Cell Factories 2010, 9, 88. [Google Scholar] [CrossRef]
- Ceric, S.; Kurtanjek, Z. Model identification, parameter estimation, and dynamic flux analysis of E. coli central metabolism. Chem. Biochem. Eng. Q. 2006, 20, 243–253. [Google Scholar]
- Tusek, A.J.; Kurtanjek, Z. Model and global sensitivity analysis of E. coli central metabolism. In Proceedings MathMod; Troch, I., Breitenecker, F., Eds.; ARGESIM: Vienna, Austria, 2009; p. 253. [Google Scholar]
- Teusink, B.; Passarge, J.; Reijenga, C.A.; Esgalhado, E.; van der Weijden, C.C.; Schepper, M.; Walsh, M.C.; Bakker, B.M.; van Dam, K.; Westerhoff, H.V.; et al. Can yeast glycolysis be understood in terms of in vitro kinetics of the constituent enzymes? Testing biochemistry. Eur. J. Biochem. 2000, 267, 5313–5329. [Google Scholar] [CrossRef]
- Olivier, B.G.; Snoep, J.L. Web-based kinetic modelling using JWS Online. Bioinformatics 2004, 20, 2143–2144. [Google Scholar] [CrossRef]
- Seressiotis, A.; Bailey, J.E. MPS: An algorithm and data base for metabolic pathways synthesis. Biotechnol. Lett. 1986, 8, 837–842. [Google Scholar] [CrossRef]
- Tomita, M.; Hashimoto, K.; Takahashi, K.; Shimizu, T.; Matsuzaki, Y.; Miyoshi, F.; Saito, K.; Tanida, S.; Yugi, K.; Venter, J.C. E-Cell: Software environment for whole cell simulation. Bioinformatics 1999, 15, 72–84. [Google Scholar] [CrossRef]
- Tomita, M. Whole-cell simulation: A grand challenge of the 21st century. Trends Biotechnol. 2001, 19, 205–210. [Google Scholar] [CrossRef]
- Slepchenko, B.M.; Schaff, J.C.; Macara, I.; Loew, L.M. Quantitative cell biology with the Virtual Cell. Trends Cell Biol. 2003, 13, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Schmid, J.W.; Mauch, K.; Reuss, M.; Gilles, E.D.; Kremling, A. Metabolic design based on a coupled gene expression—Metabolic network model of tryptophan production in Escherichia Coli. Metab. Eng. 2004, 6, 364–377. [Google Scholar] [CrossRef]
- Costa, R.S.; Machado, D.; Rocha, I.; Ferreira, E.C. Large scale dynamic model reconstruction for the central carbon metabolism of Escherichia coli. In Distributed Computing, Artificial Intelligence, Bioinformatics, Soft Computing, and Ambient Assisted Living, Proceedings of the IWANN Conference, Salamanca, Spain, 10–12 June 2009; Omatu, S., Ed.; Part II, LNCS 5518; Springer: Berlin, Germany, 2009; pp. 1079–1083. [Google Scholar]
- Costa, R.S.; Machado, D.; Rocha, I.; Ferreira, E.C. Hybrid dynamic modeling of Escherichia coli central metabolic network combining Michaelis–Menten and approximate kinetic equations. BioSystems 2010, 100, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Machado, D.; Zhuang, K.H.; Sonnenschein, N.; Herrgård, M.J. Current challenges in modeling cellular metabolism. Front. Bioeng. Biotechnol. 2015, 2–3, 4–96. [Google Scholar] [CrossRef] [PubMed]
- Styczynski, M.P.; Stephanopoulos, G. Overview of computational methods for the inference of gene regulatory networks. Comput. Chem. Eng. 2005, 29, 519–534. [Google Scholar] [CrossRef]
- Visser, D.; Schmid, J.W.; Mauch, K.; Reuss, M.; Heijnen, J.J. Optimal re-design of primarymetabolism in Escherichia coli using linlog kinetics. Metab. Eng. 2004, 6, 378–390. [Google Scholar] [CrossRef]
- Xiong, J. Essential Bioinformatics; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Rocha, I.; Maia, P.; Evangelista, P.; Vilaça, P.; Soares, S.; Pinto, J.P.; Nielsen, J.; Patil, K.R.; Ferreira, E.C.; Rocha, M. OptFlux: An open-source software platform for in silico metabolic engineering. BMC Syst Biol. 2010, 4, 45. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.H.; Wang, F.S.; Chang, M.S. Multi-objective optimization of enzyme manipulations in metabolic networks considering resilience effects. BMC Syst. Biol. 2011, 5, 145. [Google Scholar] [CrossRef]
- Slominski, A.; Semak, I.; Pisarchik, A.; Sweatman, T.; Szczesniewski, A.; Wortsman, J. Conversion of l-tryptophan to serotonin and melatonin in human melanoma cells. FEBS Lett. 2002, 511, 102–106. [Google Scholar] [CrossRef]
- Chen, M.; Ma, C.; Chen, L.; Zeng, A.P. Integrated laboratory evolution and rational engineering of GalP/Glk-dependent Escherichia coli for higher yield and productivity of l-tryptophan biosynthesis. Metab. Eng. Commun. 2021, 12, e00167. [Google Scholar] [CrossRef]
- Chen, M. Novel Approaches for In Vivo Evolution, Screening and Characterization of Enzymes for Metabolic Engineering of Escherichia coli as Hyper l-tryptophan Producer. Ph.D. Thesis, TU Hamburg, Hamburg, Germany, 2020. [Google Scholar]
- Chen, L. Rational Metabolic Engineering and Systematic Analysis of Escherichia coli for l-tryptophan Bioproduction. Ph.D. Thesis, TU Hamburg, Hamburg, Germany, 2016. [Google Scholar]
- Chen, L.; Zeng, A.P. Rational design and metabolic analysis of Escherichia coli for effective production of l-tryptophan at high concentration. Appl Microbiol Biotechnol. 2017, 101, 559–568. [Google Scholar] [CrossRef]
- Chen, M.; Chen, L.; Zeng, A.P. CRISPR/Cas9-facilitated engineering with growth-coupled and sensor-guided in vivo screening of enzyme variants for a more efficient chorismate pathway in E Coli. Metab. Eng. Commun. 2019, 9, e00094. [Google Scholar] [CrossRef]
- Li, Z.; Wang, H.; Ding, D.; Liu, Y.; Fang, H.; Chang, Z.; Chen, T.; Zhang, D. Metabolic engineering of E. coli for production of chemicals derived from the shikimate pathway. J. Ind. Microbiol. Biotechnol. 2020, 47, 525–535. [Google Scholar] [CrossRef]
- Niu, H.; Li, R.; Liang, Q.; Qi, Q.; Li, Q.; Gu, P. Metabolic engineering for improving l-tryptophan production in E. coli. J. Ind. Microbiol. Biotechnol. 2019, 46, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Carmona, S.B.; Flores, N.; Martínez-Romero, E.; Gosset, G.; Bolívar, F.; Escalante, A. Evolution of an Escherichia coli PTS− strain: A study of reproducibility and dynamics of an adaptive evolutive process. Appl. Microbiol. Biotechnol. 2020, 104, 9309–9325. [Google Scholar] [CrossRef]
- Bhartiya, S.; Chaudhary, N.; Venkatesh, K.V.; Doyle III, F.J. Multiple feedback loop design in the tryptophan regulatory network of E. coli suggests a paradigm for robust regulation of processes in series. J. R. Soc. Interface 2006, 3, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Xiu, Z.L.; Zeng, A.P.; Deckwer, W.D. Model analysis concerning the effects of growth rate and intracellular tryptophan level on the stability and dynamics of tryptophan biosynthesis in bacteria. J. Biotechnol. 1997, 58, 125–140. [Google Scholar] [CrossRef]
- Xiu, Z.L.; Chang, Z.Y.; Zeng, A.P. Nonlinear dynamics of regulation of bacterial trp operon: Model analysis of integrated effects of repression, feedback inhibition, and attenuation. Biotechnol. Prog. 2002, 18, 686–693. [Google Scholar] [CrossRef]
- Chen, L.; Chen, M.; Ma, C.; Zeng, A.P. Discovery of feed-forward regulation in l-tryptophan biosynthesis and its use in metabolic engineering of E. coli for efficient tryptophan bioproduction. Metab. Eng. 2018, 47, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, B.; Zambrano, J. Analysis of simple bioreactor models—A comparison between Monod and Contois kinetics. In Proceedings of the IWA Special International Conference: “Activated Sludge—100 Years and Counting”, Essen, Germany, 12–14 June 2014. [Google Scholar] [CrossRef]
- Termonia, Y.; Ross, J. Oscillations and control features in glycolysis: Numerical analysis of a comprehensive model. Proc. Nat. Acad. Sci. USA 1981, 78, 2952–2956. [Google Scholar] [CrossRef]
- Termonia, Y.; Ross, J. Oscillations and control features in glycolysis: Analysis of resonance effects. Proc. Nat. Acad. Sci. USA 1981, 78, 3563–3566. [Google Scholar] [CrossRef] [PubMed]
- Stephanopoulos, G.; Simpson, T.W. Flux amplification in complex metabolic networks. Chem. Eng. Sci. 1997, 52, 2607–2627. [Google Scholar] [CrossRef]
- Scoban, A.G.; Maria, G. Model-based optimization of the feeding policy of a fluidized bed bioreactor for mercury uptake by immobilized Pseudomonas putida cells. Asia-Pac. J. Chem. Eng. 2016, 11, 721–734. [Google Scholar] [CrossRef]
- Maria, G.; Crisan, M. Operation of a mechanically agitated semi-continuous multi-enzymatic reactor by using the Pareto-optimal multiple front method. J. Process Control. 2017, 53, 95–105. [Google Scholar] [CrossRef]
- Mathews, C.K.; van Holde, K.E.; Ahem, K.G. Biochemistry; Prentice Hall: Hoboken, NJ, USA, 1999. [Google Scholar]
- Calhoun, K.A.; Swartz, J.R. Total amino acid stabilization during cell-free protein synthesis reactions. J. Biotechnol. 2006, 123, 193–203. [Google Scholar] [CrossRef]
- Noor, E.; Eden, E.; Milo, R.; Alon, U. Central Carbon Metabolism as a minimal biochemical walk between precursors for biomass and energy. Mol. Cell 2010, 39, 809–820. [Google Scholar] [CrossRef]
- Kreth, J.; Lengeler, J.W.; Jahreis, K. Characterization of Pyruvate Uptake in Escherichia coli K-12. PLoS ONE 2013, 8, E67125. [Google Scholar] [CrossRef]
- Ruby, E.G.; Nealson, K.H. Pyruvate production and excretion by the luminous marine bacteria. Appl. Environ. Microbiol. 1977, 34, 164–169. Available online: https://aem.asm.org (accessed on 4 May 2021). [CrossRef] [PubMed]
- Maria, G. ARS combination with an evolutionary algorithm for solving MINLP optimization problems. In Modelling, Identification and Control; Hamza, M.H., Ed.; IASTED/ACTA Press: Anaheim, CA, USA, 2003; pp. 112–117. Available online: https://www.actapress.com/Content_of_Proceeding.aspx?proceedingID=213 (accessed on 22 October 2021).
- Bier, M.; Teusink, B.; Kholodenko, B.N.; Westerhoff, H.V. Control analysis of glycolytic oscillations. Biophys. Chem. 1996, 62, 15–24. [Google Scholar] [CrossRef]
- Silva, A.S.; Yunes, J.A. Conservation of glycolytic oscillations in Saccharomyces cerevisiae. Genet. Mol. Res. 2006, 3–5, 525–535. [Google Scholar]
- Bharat, A. Process Analytical Technology (PAT). 2013. Available online: https://www.slideshare.net/anjalibharat19/process-analytical-tchnology%20 (accessed on 7 December 2021).
- Loeblein, C.; Perkins, J.; Srinivasan, B.; Bonvin, D. Performance analysis of on-line batch optimization systems. Comput. Chem. Eng. 1997, 21, S867–S872. [Google Scholar] [CrossRef]
- Maria, G.; Dan, A. Derivation of optimal operating policies under safety and technological constraints for the acetoacetylation of pyrrole in a semi-batch catalytic reactor. Comput. Chem. Eng. 2011, 35, 177–189. [Google Scholar] [CrossRef]
- Maria, G.; Crisan, M. Evaluation of optimal operation alternatives of reactors used for D-glucose oxidation in a bi-enzymatic system with a complex deactivation kinetics. Asia-Pac. J. Chem Eng. 2014, 10, 22–44. [Google Scholar] [CrossRef]
- Mihail, R.; Maria, G. A Modified Matyas Algorithm (MMA) for Random Process Optimization. Comput. Chem. Eng. 1986, 10, 539–544. [Google Scholar] [CrossRef]
- Avili, M.G.; Fazaelipoor, M.H.; Jafari, S.A.; Ataei, S.A. Comparison between batch and fed-batch production of rhamnolipid by Pseudomonas aeruginosa. Iran. J. Biotechnol. 2012, 10, 263–269. Available online: https://www.researchgate.net/publication/288966207 (accessed on 21 October 2021).


| Rate Expression | Kinetic Model Parameters (Units in mM, μM, min) |
|---|---|
; | = 59.062, 1/min·mM = 0.5443, 1/min = 17.796, 1/min = 14.094, 1/min = 1.157, 1/min = 3.53, μM = 1.92 = 0.04, μM (see footnote (d)) |
(see footnotes (a)–(d)) | g = −0.32 = 0.36365, 1/min = 3.9923 = 0.017153, 1/min = 0.071515 |
| E. coli Strain | V1 Flux (in the Initial FBR Conditions) (mM/min) | Total GLC Consumption over the Batch Time (g) | TRP-Production of FBR (mM/min) |
|---|---|---|---|
| Maria et al. [34] (wild strain) | 1.2485 × 102 | 360 | 0.001–0.04 (nonoptimized FBR) |
| Maria [35] (T5 strain) (Table 1) | 1.2526 × 104 | 567 | 0.048 (nominal, nonoptimized FBR) |
| This paper (T5 strain) | 1.2526 × 104 | 532 | 0.06 and higher (*) (optimized FBR) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maria, G.; Renea, L. Tryptophan Production Maximization in a Fed-Batch Bioreactor with Modified E. coli Cells, by Optimizing Its Operating Policy Based on an Extended Structured Cell Kinetic Model. Bioengineering 2021, 8, 210. https://doi.org/10.3390/bioengineering8120210
Maria G, Renea L. Tryptophan Production Maximization in a Fed-Batch Bioreactor with Modified E. coli Cells, by Optimizing Its Operating Policy Based on an Extended Structured Cell Kinetic Model. Bioengineering. 2021; 8(12):210. https://doi.org/10.3390/bioengineering8120210
Chicago/Turabian StyleMaria, Gheorghe, and Laura Renea. 2021. "Tryptophan Production Maximization in a Fed-Batch Bioreactor with Modified E. coli Cells, by Optimizing Its Operating Policy Based on an Extended Structured Cell Kinetic Model" Bioengineering 8, no. 12: 210. https://doi.org/10.3390/bioengineering8120210
APA StyleMaria, G., & Renea, L. (2021). Tryptophan Production Maximization in a Fed-Batch Bioreactor with Modified E. coli Cells, by Optimizing Its Operating Policy Based on an Extended Structured Cell Kinetic Model. Bioengineering, 8(12), 210. https://doi.org/10.3390/bioengineering8120210