

The Characterization of Serum-Free Media on Human Mesenchymal Stem Cell Fibrochondrogenesis

 , , and
, , and
Abstract

1. Introduction
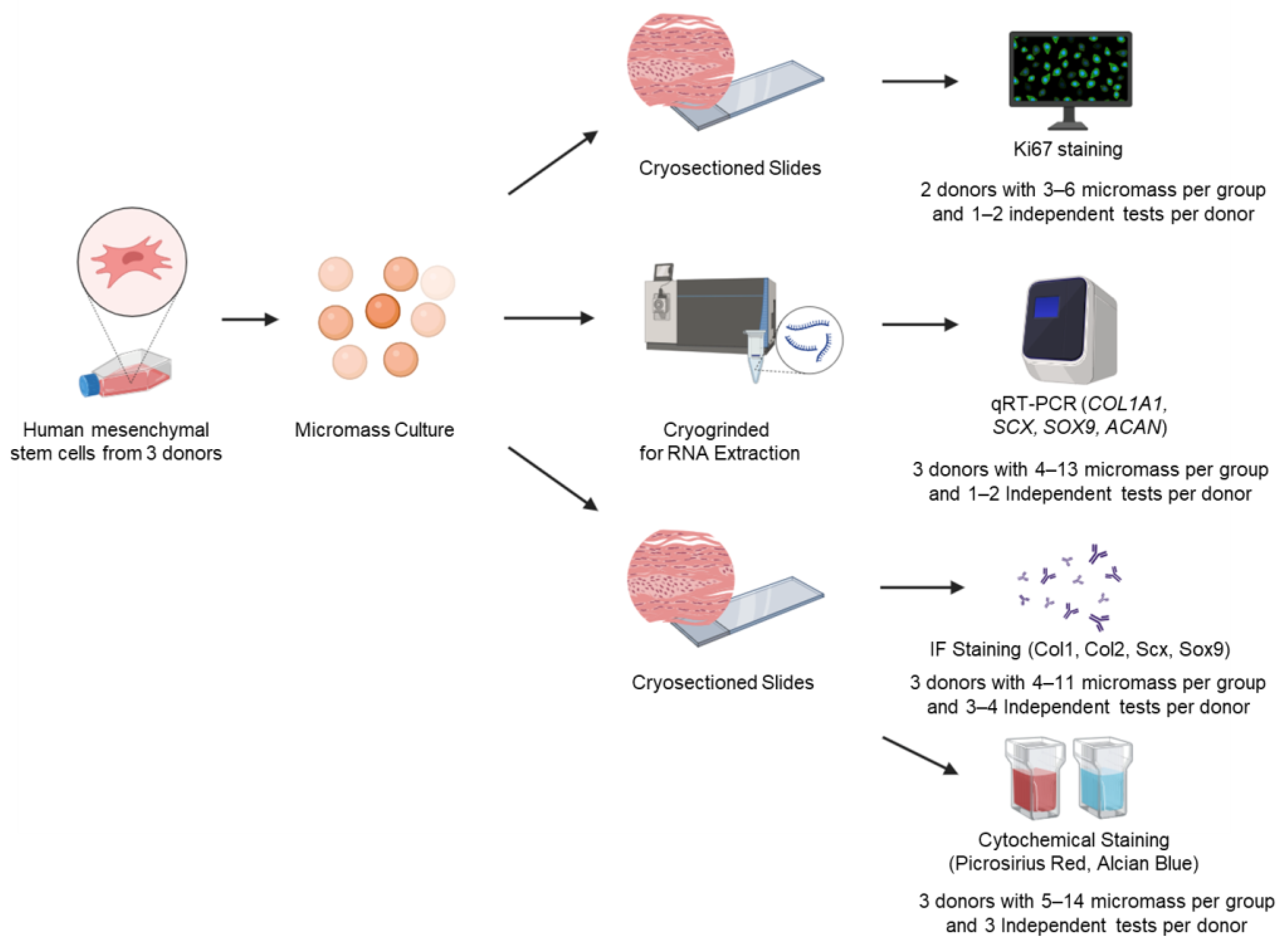
2. Materials and Methods
2.1. Cell Culture
2.2. Fibrochondrogenic Differentiation
2.3. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.4. Cytochemical and Immunofluorescence Staining
2.5. The Semi-Quantification of Cytochemical and Immunofluorescence Images
2.6. Statistical Analysis
3. Results
3.1. The Effect of Fibrochondrogenic Serum-Free Media on hMSC Proliferation Using Immunofluorescence Staining
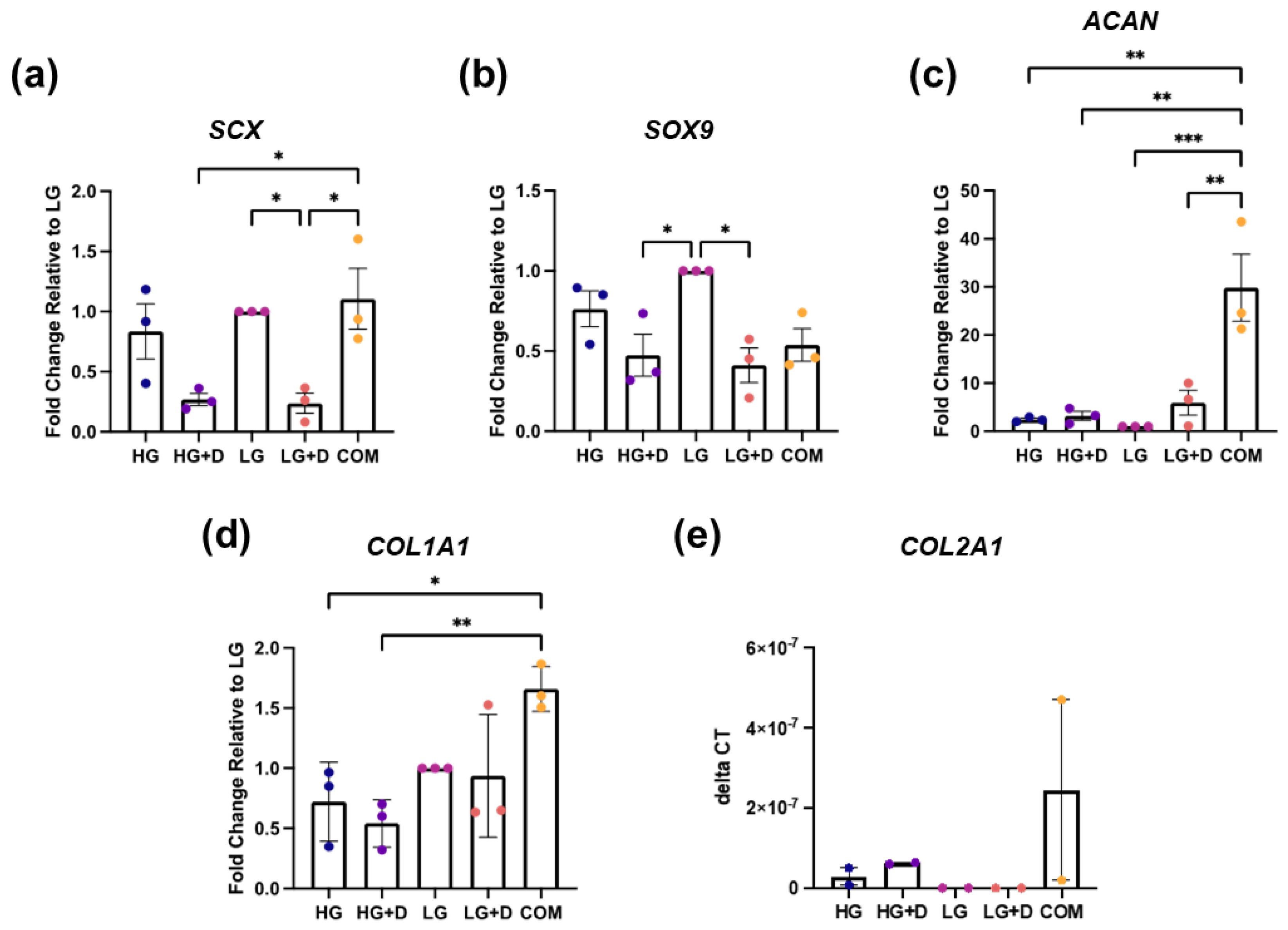
3.2. The Effect of Serum-Free Media on hMSC Fibrochondrogenic Differentiation Using qRT-PCR of Transcription Factor (TF) and ECM Genes
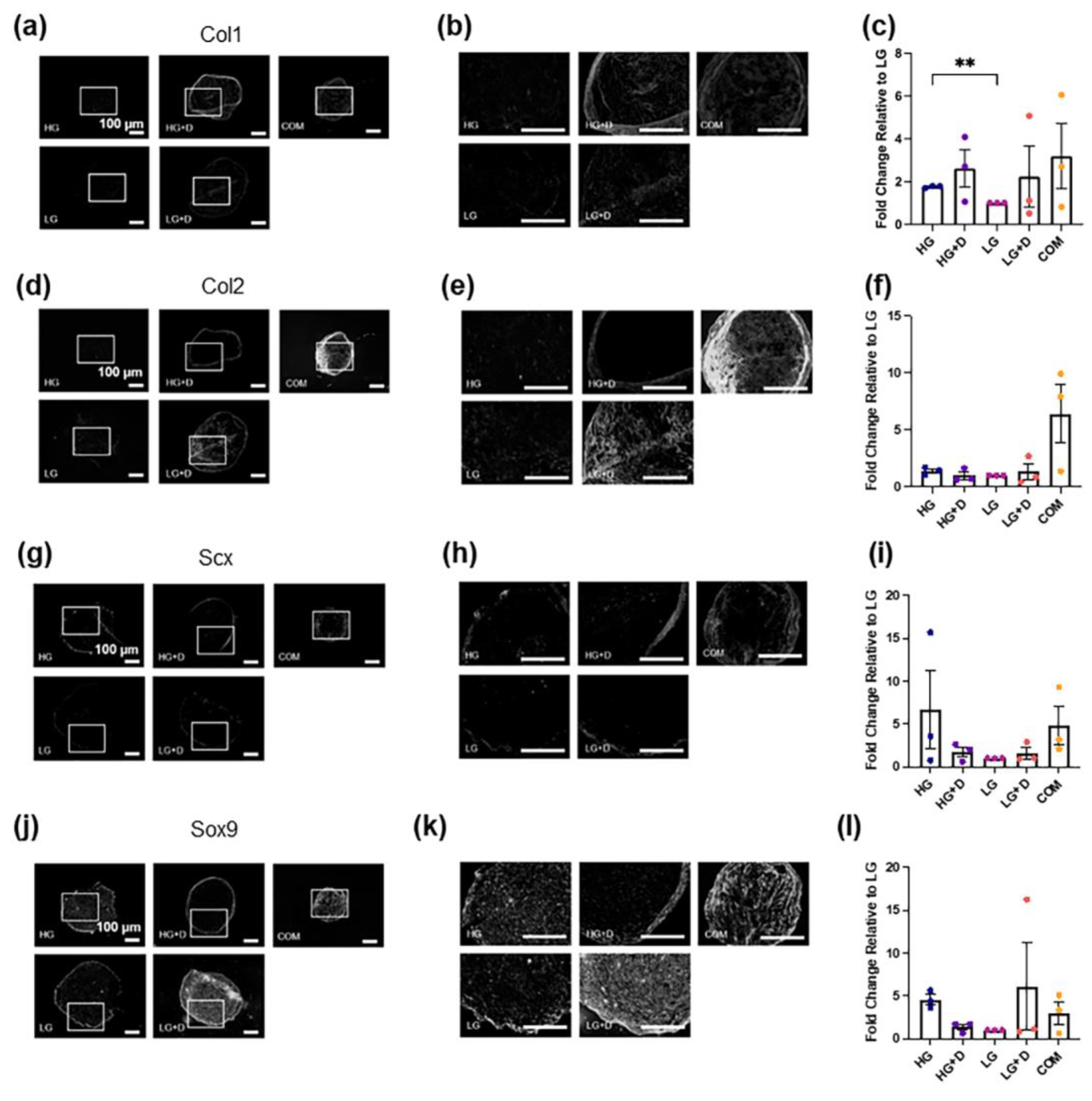
3.3. The Effect of Serum-Free Media on hMSC Fibrochondrogenic Differentiation Using Immunofluorescence Staining of TF and ECM Proteins
3.4. The Effect of Serum-Free Media on hMSC Fibrochondrogenic Differentiation Using the Total Collagen and GAG Content
3.5. The Identification of Serum-Free Media with the Greatest Fibrochondrogenic Potential
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Benjamin, M.; Ralphs, J.R. Fibrocartilage in tendons and ligaments—An adaptation to compressive load. J. Anat. 1998, 193 Pt 4, 481–494. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Fu, P.; Wu, H.; Pei, M. Meniscus, articular cartilage and nucleus pulposus: A comparative review of cartilage-like tissues in anatomy, development and function. Cell Tissue Res. 2017, 370, 53–70. [Google Scholar] [CrossRef]
- Kwan, K.Y.C. Developing a mechanically robust-, growth factor-patterned multiphasic biomaterial for rotator cuff repair. In School of Biomedical Sciences; The Chinese University of Hong Kong: Hong Kong SAR, China, 2021; pp. 1–16. [Google Scholar]
- Eleswarapu, S.V.; Responte, D.J.; Athanasiou, K.A. Tensile properties, collagen content, and crosslinks in connective tissues of the immature knee joint. PLoS ONE 2011, 6, e26178. [Google Scholar] [CrossRef] [PubMed]
- Fox, A.J.; Bedi, A.; Rodeo, S.A. The basic science of human knee menisci: Structure, composition, and function. Sports Health 2012, 4, 340–351. [Google Scholar] [CrossRef]
- Makris, E.A.; Hadidi, P.; Athanasiou, K.A. The knee meniscus: Structure-function, pathophysiology, current repair techniques, and prospects for regeneration. Biomaterials 2011, 32, 7411–7431. [Google Scholar] [CrossRef]
- Reina-Romo, E.; Gómez-Benito, M.J.; García-Aznar, J.M.; Domínguez, J.; Doblaré, M. Modeling distraction osteogenesis: Analysis of the distraction rate. Biomech. Model. Mechanobiol. 2009, 8, 323–335. [Google Scholar] [CrossRef]
- Benjamin, M.; Evans, E.J. Fibrocartilage. J. Anat. 1990, 171, 1–15. [Google Scholar]
- Ker, D.F.E.; Wang, D.; Behn, A.W.; Wang, E.T.H.; Zhang, X.; Zhou, B.Y.; Mercado-Pagán, Á.E.; Kim, S.; Kleimeyer, J.; Gharaibeh, B.; et al. Functionally graded, bone- and tendon-like polyurethane for rotator cuff repair. Adv. Funct. Mater. 2018, 28, 1707107. [Google Scholar] [CrossRef]
- Sihvonen, R.; Paavola, M.; Malmivaara, A.; Itälä, A.; Joukainen, A.; Nurmi, H.; Kalske, J.; Järvinen, T.L. Arthroscopic partial meniscectomy versus sham surgery for a degenerative meniscal tear. N. Engl. J. Med. 2013, 369, 2515–2524. [Google Scholar] [CrossRef]
- Katz, J.N. Lumbar disc disorders and low-back pain: Socioeconomic factors and consequences. J. Bone Joint Surg. Am. 2006, 88 (Suppl. 2), 21–24. [Google Scholar] [CrossRef]
- Chase, L.G.; Yang, S.; Zachar, V.; Yang, Z.; Lakshmipathy, U.; Bradford, J.; Boucher, S.E.; Vemuri, M.C. Development and characterization of a clinically compliant xeno-free culture medium in good manufacturing practice for human multipotent mesenchymal stem cells. Stem Cells Transl. Med. 2012, 1, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Mendicino, M.; Bailey, A.M.; Wonnacott, K.; Puri, R.K.; Bauer, S.R. MSC-based product characterization for clinical trials: An FDA perspective. Cell Stem Cell 2014, 14, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Uccelli, A.; Moretta, L.; Pistoia, V. Mesenchymal stem cells in health and disease. Nat. Rev. Immunol. 2008, 8, 726–736. [Google Scholar] [CrossRef] [PubMed]
- Fabre, H.; Ducret, M.; Degoul, O.; Rodriguez, J.; Perrier-Groult, E.; Aubert-Foucher, E.; Pasdeloup, M.; Auxenfans, C.; McGuckin, C.; Forraz, N.; et al. Characterization of different sources of human MSCs expanded in serum-free conditions with quantification of chondrogenic induction in 3D. Stem Cells Int. 2019, 2019, 2186728. [Google Scholar] [CrossRef]
- Kim, H.J.; Park, J.S. Usage of human mesenchymal stem cells in cell-based therapy: Advantages and disadvantages. Dev. Reprod. 2017, 21, 1–10. [Google Scholar] [CrossRef]
- Indrawattana, N.; Chen, G.; Tadokoro, M.; Shann, L.H.; Ohgushi, H.; Tateishi, T.; Tanaka, J.; Bunyaratvej, A. Growth factor combination for chondrogenic induction from human mesenchymal stem cell. Biochem. Biophys. Res. Commun. 2004, 320, 914–919. [Google Scholar] [CrossRef]
- Li, S.; Sengers, B.G.; Oreffo, R.O.; Tare, R.S. Chondrogenic potential of human articular chondrocytes and skeletal stem cells: A comparative study. J. Biomater. Appl. 2015, 29, 824–836. [Google Scholar] [CrossRef]
- Skog, M.; Muhonen, V.; Nystedt, J.; Narcisi, R.; Kontturi, L.S.; Urtti, A.; Korhonen, M.; van Osch, G.J.; Kiviranta, I. Xeno-free chondrogenesis of bone marrow mesenchymal stromal cells: Towards clinical-grade chondrocyte production. Cytotechnology 2015, 67, 905–919. [Google Scholar] [CrossRef]
- Mueller, M.B.; Blunk, T.; Appel, B.; Maschke, A.; Goepferich, A.; Zellner, J.; Englert, C.; Prantl, L.; Kujat, R.; Nerlich, M.; et al. Insulin is essential for in vitro chondrogenesis of mesenchymal progenitor cells and influences chondrogenesis in a dose-dependent manner. Int. Orthop. 2013, 37, 153–158. [Google Scholar] [CrossRef]
- Shao, X.X.; Duncan, N.A.; Lin, L.; Fu, X.; Zhang, J.Y.; Yu, C.L. Serum-free media for articular chondrocytes in vitro expansion. Chin. Med. J. 2013, 126, 2523–2529. [Google Scholar] [CrossRef]
- Kaji, D.A.; Montero, A.M.; Patel, R.; Huang, A.H. Transcriptional profiling of mESC-derived tendon and fibrocartilage cell fate switch. Nat. Commun. 2021, 12, 4208. [Google Scholar] [CrossRef] [PubMed]
- Koay, E.J.; Athanasiou, K.A. Development of serum-free, chemically defined conditions for human embryonic stem cell-derived fibrochondrogenesis. Tissue Eng. Part A 2009, 15, 2249–2257. [Google Scholar] [CrossRef] [PubMed]
- Richter, W. Mesenchymal stem cells and cartilage in situ regeneration. J. Intern. Med. 2009, 266, 390–405. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Caplan, A.I.; Welter, J.F.; Baskaran, H. Glucose availability affects extracellular matrix synthesis during chondrogenesis in vitro. Tissue Eng. Part A 2021, 27, 1321–1332. [Google Scholar] [CrossRef]
- Hoben, G.M.; Willard, V.P.; Athanasiou, K.A. Fibrochondrogenesis of hESCs: Growth factor combinations and cocultures. Stem Cells Dev. 2009, 18, 283–292. [Google Scholar] [CrossRef]
- Abbott, R.D.; Purmessur, D.; Monsey, R.D.; Iatridis, J.C. Regenerative potential of TGFβ3 + Dex and notochordal cell conditioned media on degenerated human intervertebral disc cells. J. Orthop. Res. 2012, 30, 482–488. [Google Scholar] [CrossRef]
- Yin, X.; Motorwala, A.; Vesvoranan, O.; Levene, H.B.; Gu, W.; Huang, C.Y. Effects of glucose deprivation on ATP and proteoglycan production of intervertebral disc cells under hypoxia. Sci. Rep. 2020, 10, 8899. [Google Scholar] [CrossRef]
- Heard, B.J.; Barton, K.I.; Agbojo, O.M.; Chung, M.; Sevick, J.L.; Bader, T.J.; Martin, C.R.; Shrive, N.G.; Hart, D.A. Molecular response of rabbit menisci to surgically induced hemarthrosis and a single intra-articular dexamethasone treatment. J. Orthop. Res. 2019, 37, 2043–2052. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Mifune, Y.; Inui, A.; Nishimoto, H.; Yamaura, K.; Mukohara, S.; Shinohara, I.; Kuroda, R. Influence of diabetes-induced glycation and oxidative stress on the human rotator cuff. Antioxidants 2022, 11, 743. [Google Scholar] [CrossRef]
- Marlina, M.; Rahmadian, R.; Armenia, A.; Aviani, J.K.; Sholihah, I.A.; Kusuma, H.S.W.; Azizah, A.M.; Elida, N.; Widowati, W. Conditioned medium of IGF1-induced synovial membrane mesenchymal stem cells increases chondrogenic and chondroprotective markers in chondrocyte inflammation. Biosci. Rep. 2021, 41, BSR20202038. [Google Scholar] [CrossRef]
- Duarte-Olivenza, C.; Hurle, J.M.; Montero, J.A.; Lorda-Diez, C.I. Modeling the differentiation of embryonic limb chondroprogenitors by cell death and cell senescence in high density micromass cultures and their regulation by FGF signaling. Cells 2022, 12, 175. [Google Scholar] [CrossRef] [PubMed]
- Rajpar, I.; Barrett, J.G. Optimizing growth factor induction of tenogenesis in three-dimensional culture of mesenchymal stem cells. J. Tissue Eng. 2019, 10, 2041731419848776. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Zhang, X.; Wang, D.; Tuan, R.S.; Ker, D.F.E. Synergistic effects of growth factor-based serum-free medium and tendon-like substrate topography on tenogenesis of mesenchymal stem cells. Biomater. Adv. 2023, 146, 213316. [Google Scholar] [CrossRef]
- Wang, D.; Pun, C.C.M.; Huang, S.; Tang, T.C.M.; Ho, K.K.W.; Rothrauff, B.B.; Yung, P.S.H.; Blocki, A.M.; Ker, E.D.F.; Tuan, R.S. Tendon-derived extracellular matrix induces mesenchymal stem cell tenogenesis via an integrin/transforming growth factor-β crosstalk-mediated mechanism. FASEB J. 2020, 34, 8172–8186. [Google Scholar] [CrossRef]
- Timmer, K.B.; Killian, M.L.; Harley, B.A.C. Paracrine signals influence patterns of fibrocartilage differentiation in a lyophilized gelatin hydrogel for applications in rotator cuff repair. Biomater. Sci. 2024, 12, 4806–4822. [Google Scholar] [CrossRef]
- Qin, S.; Wang, W.; Liu, Z.; Hua, X.; Fu, S.; Dong, F.; Li, A.; Liu, Z.; Wang, P.; Dai, L.; et al. Fibrochondrogenic differentiation potential of tendon-derived stem/progenitor cells from human patellar tendon. J. Orthop. Transl. 2020, 22, 101–108. [Google Scholar] [CrossRef]
- Armiento, A.R.; Alini, M.; Stoddart, M.J. Articular fibrocartilage—Why does hyaline cartilage fail to repair? Adv. Drug Deliv. Rev. 2019, 146, 289–305. [Google Scholar] [CrossRef]
- Uchida, A.; Mihata, T.; Hasegawa, A.; Noguchi, Y.; Neo, M. Superior capsule reconstruction for irreparable rotator cuff tears yields good clinical outcomes for patients with and without diabetes mellitus. Arthroscopy 2024, 24, S0749–S8063. [Google Scholar] [CrossRef]
- Lin, Y.C.; Li, Y.J.; Rui, Y.F.; Dai, G.C.; Shi, L.; Xu, H.L.; Ni, M.; Zhao, S.; Chen, H.; Wang, C.; et al. The effects of high glucose on tendon-derived stem cells: Implications of the pathogenesis of diabetic tendon disorders. Oncotarget 2017, 8, 17518–17528. [Google Scholar] [CrossRef]
- Ullah, M.; Hamouda, H.; Stich, S.; Sittinger, M.; Ringe, J. A reliable protocol for the isolation of viable, chondrogenically differentiated human mesenchymal stem cells from high-density pellet cultures. Biores Open Access 2012, 1, 297–305. [Google Scholar] [CrossRef]
- Ker, D.F.E.; Chu, B.; Phillippi, J.A.; Gharaibeh, B.; Huard, J.; Weiss, L.E.; Campbell, P.G. Engineering spatial control of multiple differentiation fates within a stem cell population. Biomaterials 2011, 32, 3413–3422. [Google Scholar] [CrossRef] [PubMed]
- Ker, D.F.; Sharma, R.; Wang, E.T.; Yang, Y.P. Development of mRuby2-transfected C3H10T1/2 fibroblasts for musculoskeletal tissue engineering. PLoS ONE 2015, 10, e0139054. [Google Scholar] [CrossRef] [PubMed]
- Ker, D.F.E.; Nain, A.S.; Weiss, L.E.; Wang, J.; Suhan, J.; Amon, C.H.; Campbell, P.G. Bioprinting of growth factors onto aligned sub-micron fibrous scaffolds for simultaneous control of cell differentiation and alignment. Biomaterials 2011, 32, 8097–8107. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Ker, D.F.E.; Ng, K.W.; Li, K.; Gharaibeh, B.; Safran, M.; Cheung, E.; Campbell, P.; Weiss, L.; Yang, Y.P. Combinatorial mechanical gradation and growth factor biopatterning strategy for spatially controlled bone-tendon-like cell differentiation and tissue formation. NPG Asia Mater. 2021, 13, 26. [Google Scholar] [CrossRef]
- Hidalgo Perea, S.; Lyons, L.P.; Nishimuta, J.F.; Weinberg, J.B.; McNulty, A.L. Evaluation of culture conditions for in vitro meniscus repair model systems using bone marrow-derived mesenchymal stem cells. Connect. Tissue Res. 2020, 61, 322–337. [Google Scholar] [CrossRef]
- Qi, X.; Su, T.; Zhang, M.; Tong, X.; Pan, W.; Zeng, Q.; Zhou, Z.; Shen, L.; He, X.; Shen, J. Macroporous hydrogel scaffolds with tunable physicochemical properties for tissue engineering constructed using renewable polysaccharides. ACS Appl. Mater. Interfaces 2020, 12, 13256–13264. [Google Scholar] [CrossRef]
- Su, T.; Zhang, M.; Zeng, Q.; Pan, W.; Huang, Y.; Qian, Y.; Dong, W.; Qi, X.; Shen, J. Mussel-inspired agarose hydrogel scaffolds for skin tissue engineering. Bioact. Mater. 2021, 6, 579–588. [Google Scholar] [CrossRef]
- Blitz, E.; Sharir, A.; Akiyama, H.; Zelzer, E. Tendon-bone attachment unit is formed modularly by a distinct pool of Scx- and Sox9-positive progenitors. Development 2013, 140, 2680–2690. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, D.; Mak, K.K.; Tuan, R.S.; Ker, D.F.E. Engineering musculoskeletal grafts for multi-tissue unit repair: Lessons from developmental biology and wound healing. Front. Physiol. 2021, 12, 691954. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, X.; Huang, S.; Liu, Y.; Fu, B.S.; Mak, K.K.; Blocki, A.M.; Yung, P.S.; Tuan, R.S.; Ker, D.F.E. Engineering multi-tissue units for regenerative medicine: Bone-tendon-muscle units of the rotator cuff. Biomaterials 2021, 272, 120789. [Google Scholar] [CrossRef]
- Madry, H.; Rey-Rico, A.; Venkatesan, J.K.; Johnstone, B.; Cucchiarini, M. Transforming growth factor beta-releasing scaffolds for cartilage tissue engineering. Tissue Eng. Part B Rev. 2014, 20, 106–125. [Google Scholar] [CrossRef] [PubMed]
- Qu, D.; Zhu, J.P.; Childs, H.R.; Lu, H.H. Nanofiber-based transforming growth factor-β3 release induces fibrochondrogenic differentiation of stem cells. Acta Biomater. 2019, 93, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, H.; Kagawa, K.; Minamizaki, T.; Nakano, M.; Aubin, J.E.; Kozai, K.; Tsuga, K.; Yoshiko, Y. Developmental impairments of craniofacial bone and cartilage in transgenic mice expressing FGF10. Bone Rep. 2023, 18, 101692. [Google Scholar] [CrossRef]
- Ganji, E.; Leek, C.; Duncan, W.; Patra, D.; Ornitz, D.M.; Killian, M.L. Targeted deletion of Fgf9 in tendon disrupts mineralization of the developing enthesis. FASEB J. 2023, 37, e22777. [Google Scholar] [CrossRef]
- Prabhath, A.; Vernekar, V.N.; Esdaille, C.J.; Eisenberg, E.; Lebaschi, A.; Badon, M.; Seyedsalehi, A.; Dzidotor, G.; Tang, X.; Dyment, N.; et al. Pegylated insulin-like growth factor-1 biotherapeutic delivery promotes rotator cuff regeneration in a rat model. J. Biomed. Mater. Res. A 2022, 110, 1356–1371. [Google Scholar] [CrossRef]
- Han, Y.S.; Bang, O.S.; Jin, E.J.; Park, J.H.; Sonn, J.K.; Kang, S.S. High dose of glucose promotes chondrogenesis via PKCalpha and MAPK signaling pathways in chick mesenchymal cells. Cell Tissue Res. 2004, 318, 571–578. [Google Scholar] [CrossRef]
- Mackay, A.M.; Beck, S.C.; Murphy, J.M.; Barry, F.P.; Chichester, C.O.; Pittenger, M.F. Chondrogenic differentiation of cultured human mesenchymal stem cells from marrow. Tissue Eng. 1998, 4, 415–428. [Google Scholar] [CrossRef]
- Chang, T.C.; Hsu, M.F.; Wu, K.K. High glucose induces bone marrow-derived mesenchymal stem cell senescence by upregulating autophagy. PLoS ONE 2015, 10, e0126537. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Nan, L.P.; Wang, F.; Zhou, S.F.; Wang, J.C.; Feng, X.M.; Zhang, L. The effect of high glucose on the biological characteristics of nucleus pulposus-derived mesenchymal stem cells. Cell Biochem. Funct. 2020, 38, 130–140. [Google Scholar] [CrossRef]
- Wu, Y.F.; Wang, H.K.; Chang, H.W.; Sun, J.; Sun, J.S.; Chao, Y.H. High glucose alters tendon homeostasis through downregulation of the AMPK/Egr1 pathway. Sci. Rep. 2017, 7, 44199. [Google Scholar] [CrossRef]
- Lui, P.P.Y. Tendinopathy in diabetes mellitus patients-epidemiology, pathogenesis, and management. Scand. J. Med. Sci. Sports 2017, 27, 776–787. [Google Scholar] [CrossRef] [PubMed]
- Kwan, C.K.; Fu, S.C.; Yung, P.S. A high glucose level stimulate inflammation and weaken pro-resolving response in tendon cells—A possible factor contributing to tendinopathy in diabetic patients. Asia-Pac. J. Sports Med. Arthrosc. Rehabil. Technol. 2020, 19, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zuncheddu, D.; Della Bella, E.; Petta, D.; Bärtschi, C.; Häckel, S.; Deml, M.C.; Stoddart, M.J.; Grad, S.; Basoli, V. Effect of glucose depletion and fructose administration during chondrogenic commitment in human bone marrow-derived stem cells. Stem Cell Res. Ther. 2022, 13, 533. [Google Scholar] [CrossRef]
- Wu, T.J.; Fong, Y.C.; Lin, C.Y.; Huang, Y.L.; Tang, C.H. Glucose enhances aggrecan expression in chondrocytes via the PKCα/p38-miR141-3p signaling pathway. J. Cell Physiol. 2018, 233, 6878–6887. [Google Scholar] [CrossRef]
- Xiao, Y.; Peperzak, V.; van Rijn, L.; Borst, J.; de Bruijn, J.D. Dexamethasone treatment during the expansion phase maintains stemness of bone marrow mesenchymal stem cells. J. Tissue Eng. Regen. Med. 2010, 4, 374–386. [Google Scholar] [CrossRef]
- Stewart, A.A.; Byron, C.R.; Pondenis, H.C.; Stewart, M.C. Effect of dexamethasone supplementation on chondrogenesis of equine mesenchymal stem cells. Am. J. Vet. Res. 2008, 69, 1013–1021. [Google Scholar] [CrossRef]
- Derfoul, A.; Perkins, G.L.; Hall, D.J.; Tuan, R.S. Glucocorticoids promote chondrogenic differentiation of adult human mesenchymal stem cells by enhancing expression of cartilage extracellular matrix genes. Stem Cells 2006, 24, 1487–1495. [Google Scholar] [CrossRef]
- Park, J.S.; Na, K.; Woo, D.G.; Yang, H.N.; Park, K.H. Determination of dual delivery for stem cell differentiation using dexamethasone and TGF-beta3 in/on polymeric microspheres. Biomaterials 2009, 30, 4796–4805. [Google Scholar] [CrossRef]
- Na, K.; Kim, S.; Woo, D.G.; Sun, B.K.; Yang, H.N.; Chung, H.M.; Park, K.H. Combination material delivery of dexamethasone and growth factor in hydrogel blended with hyaluronic acid constructs for neocartilage formation. J. Biomed. Mater. Res. A 2007, 83, 779–786. [Google Scholar] [CrossRef]
- Lu, H.; Yang, H.; Shen, H.; Ye, G.; Lin, X.J. The clinical effect of tendon repair for tendon spontaneous rupture after corticosteroid injection in hands: A retrospective observational study. Medicine 2016, 95, e5145. [Google Scholar] [CrossRef]
- Zhang, J.; Keenan, C.; Wang, J.H. The effects of dexamethasone on human patellar tendon stem cells: Implications for dexamethasone treatment of tendon injury. J. Orthop. Res. 2013, 31, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Puzzitiello, R.N.; Patel, B.H.; Forlenza, E.M.; Nwachukwu, B.U.; Allen, A.A.; Forsythe, B.; Salzler, M.J. Adverse impact of corticosteroids on rotator cuff tendon health and repair: A systematic review of basic science studies. Arthrosc. Sports Med. Rehabil. 2020, 2, e161–e169. [Google Scholar] [CrossRef] [PubMed]
- Alm, J.J.; Heino, T.J.; Hentunen, T.A.; Vaananen, H.K.; Aro, H.T. Transient 100 nM dexamethasone treatment reduces inter- and intraindividual variations in osteoblastic differentiation of bone marrow-derived human mesenchymal stem cells. Tissue Eng. Part C Methods 2012, 18, 658–666. [Google Scholar] [CrossRef]
- Tangtrongsup, S.; Kisiday, J.D. Effects of dexamethasone concentration and timing of exposure on chondrogenesis of equine bone marrow-derived mesenchymal stem cells. Cartilage 2016, 7, 92–103. [Google Scholar] [CrossRef]
- Shintani, N.; Hunziker, E.B. Differential effects of dexamethasone on the chondrogenesis of mesenchymal stromal cells: Influence of microenvironment, tissue origin and growth factor. Eur. Cell Mater. 2011, 22, 302–319, discussion 319–320. [Google Scholar] [CrossRef]
- Jansen, B.J.; Gilissen, C.; Roelofs, H.; Schaap-Oziemlak, A.; Veltman, J.A.; Raymakers, R.A.; Jansen, J.H.; Kögler, G.; Figdor, C.G.; Torensma, R.; et al. Functional differences between mesenchymal stem cell populations are reflected by their transcriptome. Stem Cells Dev. 2010, 19, 481–490. [Google Scholar] [CrossRef]
- Francis, G.L. Albumin and mammalian cell culture: Implications for biotechnology applications. Cytotechnology 2010, 62, 1–16. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HG | HG + D | LG | LG + D | COM | |
|---|---|---|---|---|---|
| Cell Proliferation | |||||
| Ki67 | 3 | 3 | 3 | 5 | 4 |
| qRT-PCR | |||||
| COL1A1 | 3 | 3 | 4 | 4 | 5 |
| COL2A1 | 3 | 4 | / | / | 5 |
| SCX | 5 | 4 | 5 | 4 | 5 |
| SOX9 | 4 | 3 | 5 | 3 | 3 |
| ACAN | 4 | 4 | 3 | 4 | 5 |
| IF Staining | |||||
| Col1a1 | 4 | 4 | 3 | 4 | 5 |
| Col2a1 | 4 | 4 | 4 | 4 | 5 |
| Col10a1 | 4 | 5 | 3 | 4 | 2 |
| Scx | 5 | 3 | 3 | 3 | 4 |
| Sox9 | 4 | 2 | 2 | 5 | 3 |
| Cytochemical Staining | |||||
| Picrosirius Red | 3 | 4 | 2 | 5 | 5 |
| Alcian Blue | 4 | 5 | 4 | 4 | 4 |
| Total Score | 50 | 48 | 41 | 49 | 55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwan, K.Y.C.; Li, K.; Wang, Y.Y.; Tse, W.Y.; Tong, C.Y.; Zhang, X.; Wang, D.M.; Ker, D.F.E. The Characterization of Serum-Free Media on Human Mesenchymal Stem Cell Fibrochondrogenesis. Bioengineering 2025, 12, 546. https://doi.org/10.3390/bioengineering12050546
Kwan KYC, Li K, Wang YY, Tse WY, Tong CY, Zhang X, Wang DM, Ker DFE. The Characterization of Serum-Free Media on Human Mesenchymal Stem Cell Fibrochondrogenesis. Bioengineering. 2025; 12(5):546. https://doi.org/10.3390/bioengineering12050546
Chicago/Turabian StyleKwan, Ka Yu Carissa, Ke Li, Yu Yang Wang, Wai Yi Tse, Chung Yan Tong, Xu Zhang, Dan Michelle Wang, and Dai Fei Elmer Ker. 2025. "The Characterization of Serum-Free Media on Human Mesenchymal Stem Cell Fibrochondrogenesis" Bioengineering 12, no. 5: 546. https://doi.org/10.3390/bioengineering12050546
APA StyleKwan, K. Y. C., Li, K., Wang, Y. Y., Tse, W. Y., Tong, C. Y., Zhang, X., Wang, D. M., & Ker, D. F. E. (2025). The Characterization of Serum-Free Media on Human Mesenchymal Stem Cell Fibrochondrogenesis. Bioengineering, 12(5), 546. https://doi.org/10.3390/bioengineering12050546