The Inhibition of Prolyl Oligopeptidase as New Target to Counteract Chronic Venous Insufficiency: Findings in a Mouse Model



 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Saphene Vein Ligation (SVL) Model
2.3. Experimental Groups
- Group 1: Sham + vehicle, control group to which the saphenous vein ligation was not performed, orally administered with saline for 7 days (n = 8);
- Group 2: Sham + Simvastatin, group to which the saphenous vein ligation was not performed, orally administered with Simvastatin (20 mg/kg) for 7 days (n = 8);
- Group 3: Sham + KYP-2047 group to which the saphenous vein ligation was not performed; KYP-2047 (10 mg/kg) was intraperitoneal (i.p.) administered for 7 days (n = 8);
- Group 4: Saphene vein ligation (SVL), group subjected to ligation of the saphenous vein, orally administered with saline for 7 days (n = 8);
- Group 5: SVL + Simvastatin, group subjected to ligation of the saphenous vein, orally administered with Simvastatin (20 mg/kg), 30 min after saphene vein ligation, for 7 days (n = 8);
- Group 6: SVL + KYP-2047, subjected to ligation of the saphenous vein, i.p. administered with KYP-2047 (10 mg/kg), 30 min after saphene vein ligation, for 7 days (n = 8)
2.4. Histological Analysis
2.5. Masson Trichrome Staining
2.6. Van Gieson Staining
2.7. Toluidine Blue Staining
2.8. Western Blot Analysis for Interleukin 1β (IL-1β), Tumor Necrosis Factor α (TNF-α), Transforming Growth Factor β (TGFβ1), Vascular Endothelial Growth Factor (VEGF), α-Smooth Muscle Actin (αSMA) and Prolyl Endopeptidase
2.9. ELISA Kit Assay for eNOS and IL-8, Pro-Collagen 1 Alpha, and TGFβ1
2.10. Myeloperoxidase (MPO) Activity
2.11. Primary Culture of Vascular Smooth Muscle Cells (VSMCs) from Murine Saphene Vein: Tissue Block Culture Study
2.11.1. Experimental Groups for In Vivo Study
- Group 1: Sham + vehicle, control group to which the saphenous vein ligation was not performed, orally administered with saline for 7 days (n = 8);
- Group 2: Saphene vein ligation (SVL), group subjected to ligation of the saphenous vein, orally administered with saline for 7 days (n = 8);
- Group 3: SVL + KYP-2047, group subjected to ligation of the saphenous vein, administered with KYP-2047 (i.p., 10 mg/kg), 30 min after saphene vein ligation, for 7 days (n = 8).
2.11.2. Immunofluorescence Analysis
2.11.3. ELISA Kit for Endoglin
2.12. Materials
2.13. Statistical Analysis
3. Results
3.1. The Expression of POP in CVI Mouse Model
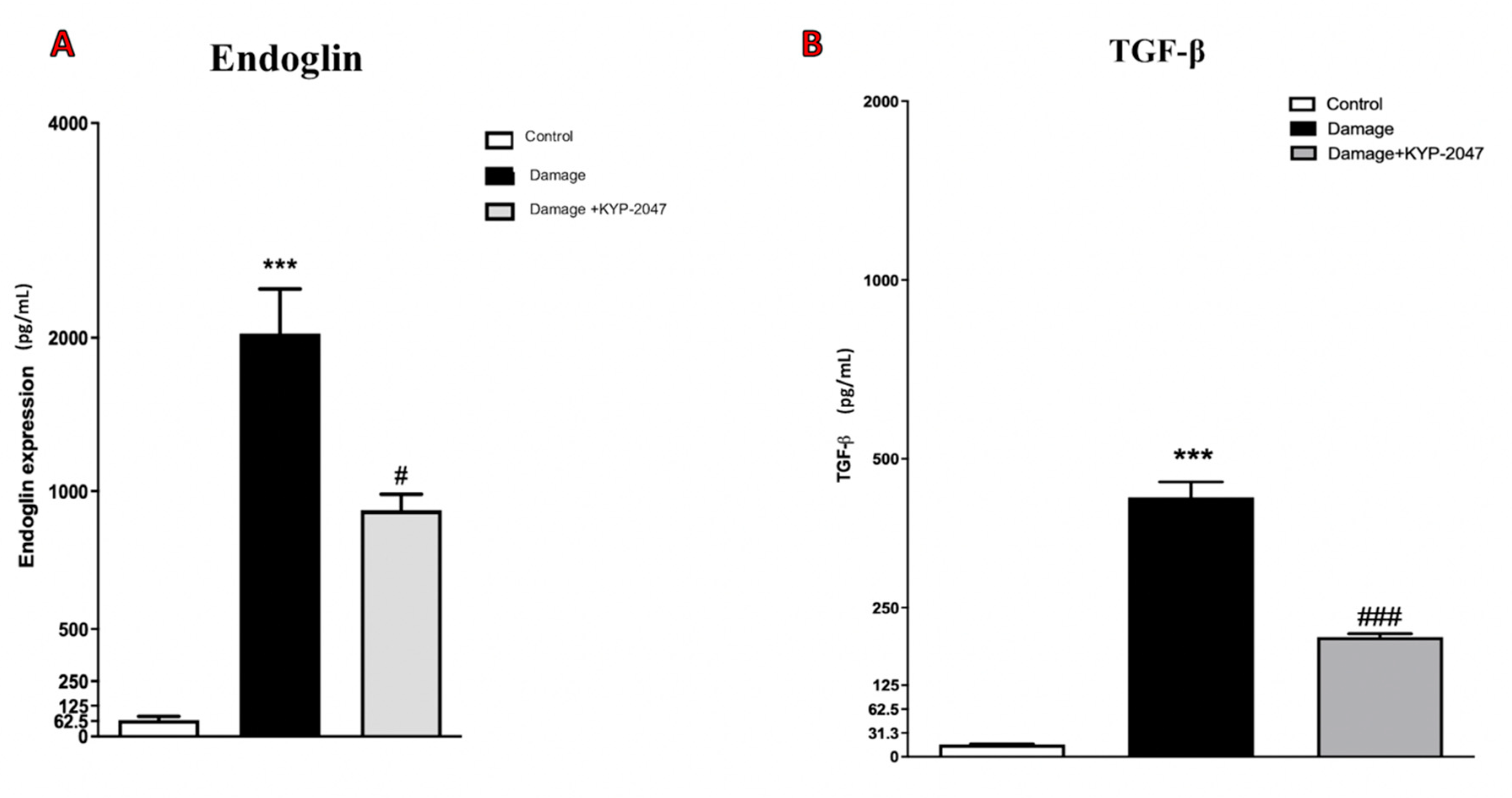
3.2. Role of KYP-2047 Treatment on TGF-β1 and IL-8, as Vascular Markers in CVI
3.3. Role of KYP-2047 Treatment on Angiogenesis Modulation and Vasodilation
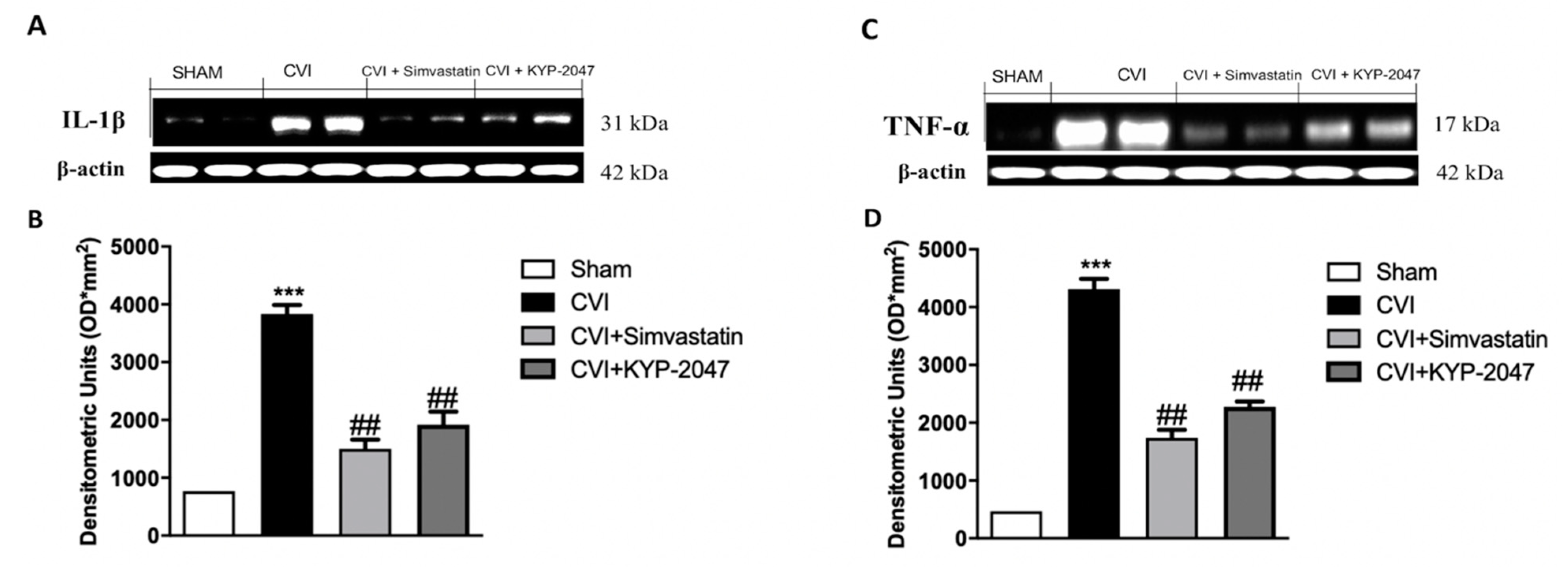
3.4. Treatment with KYP-2047 on Cytokines Expression in SVL-Damaged Mice
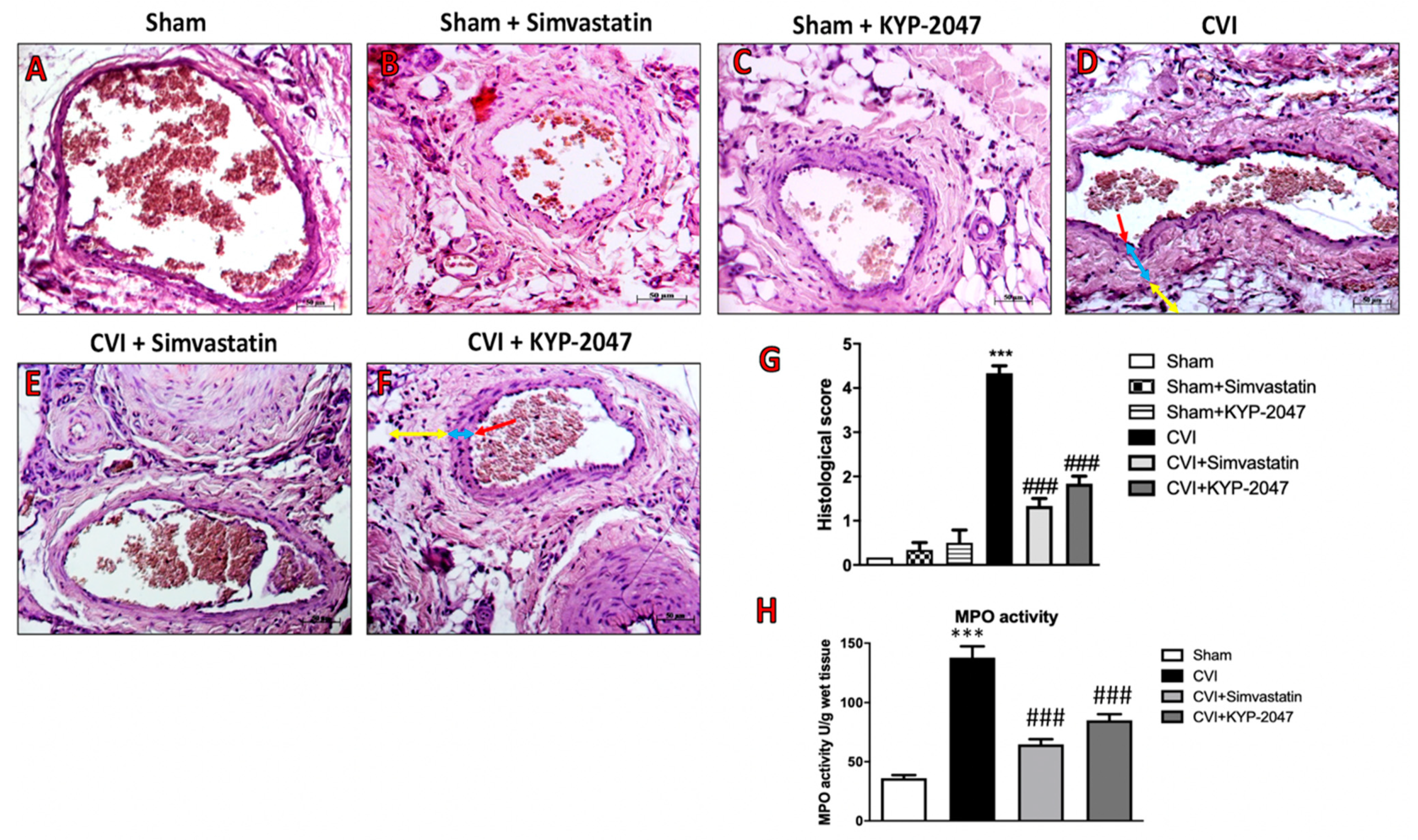
3.5. Effects of KYP-2047 on Histological Damage and Neutrophilic Activation Induced by SVL in Mice
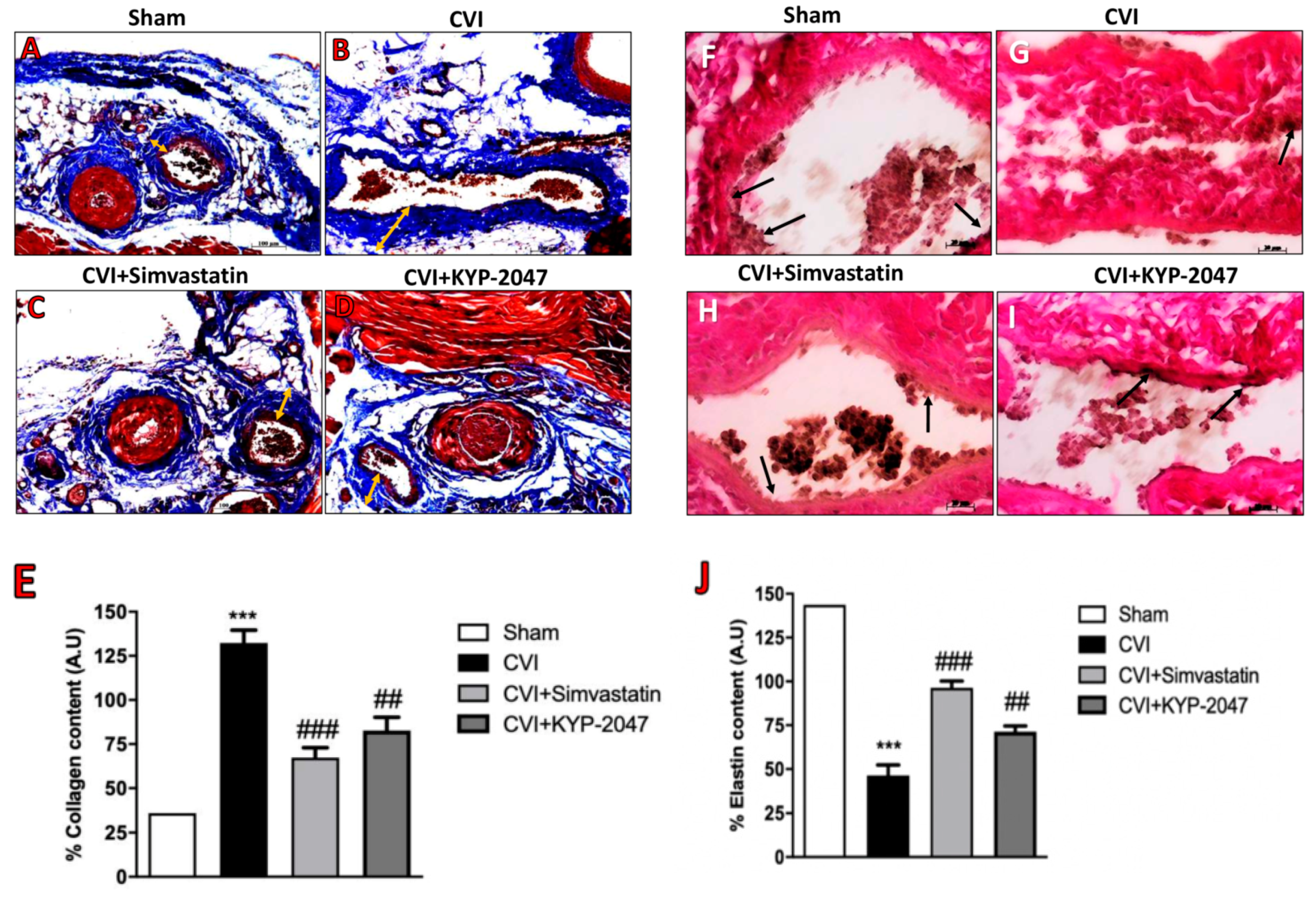
3.6. Role of KYP-2047 Treatment in the Collagen Content Reduction and Elastin Replacement
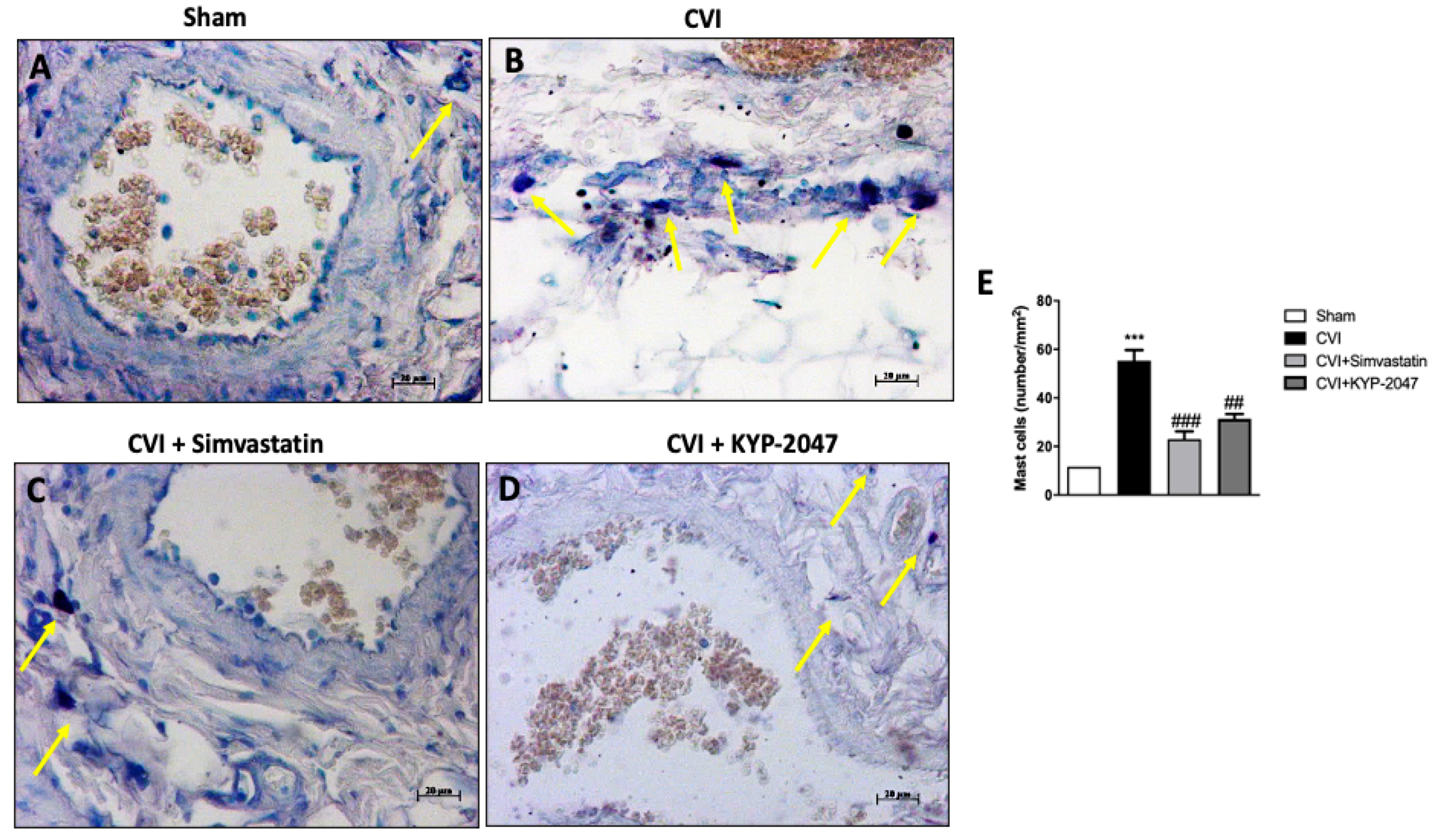
3.7. Role of KYP-2047 Treatment in Preventing Mast Cell Degranulation
3.8. Evaluation of KYP-2047 in a Saphene Vein Block Culture Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Availability of Data and Materials
References
- Onida, S.; Davies, A.H. Predicted burden of venous disease. Phlebology 2016, 31, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Raffetto, J.D.; Mannello, F. Pathophysiology of chronic venous disease. Int. Angiol. 2014, 33, 212–221. [Google Scholar] [PubMed]
- Lichota, A.; Gwozdzinski, L.; Gwozdzinski, K. Therapeutic potential of natural compounds in inflammation and chronic venous insufficiency. Eur. J. Med. Chem. 2019, 176, 68–91. [Google Scholar] [CrossRef] [PubMed]
- Santler, B.; Goerge, T. Chronic venous insufficiency-a review of pathophysiology, diagnosis, and treatment. J. Dtsch. Derm. Ges. 2017, 15, 538–556. [Google Scholar] [CrossRef]
- Youn, Y.J.; Lee, J. Chronic venous insufficiency and varicose veins of the lower extremities. Korean J. Int. Med. 2019, 34, 269–283. [Google Scholar] [CrossRef]
- Hyder, O.N.; Soukas, P.A. Chronic Venous Insufficiency: Novel Management Strategies for an Under-diagnosed Disease Process. R. I. Med. J. 2017, 100, 37–39. [Google Scholar]
- Eberhardt, R.T.; Raffetto, J.D. Chronic venous insufficiency. Circulation 2014, 130, 333–346. [Google Scholar] [CrossRef]
- Darvall, K.A.; Bate, G.R.; Adam, D.J.; Silverman, S.H.; Bradbury, A.W. Duplex ultrasound outcomes following ultrasound-guided foam sclerotherapy of symptomatic recurrent great saphenous varicose veins. Eur. J. Vasc. Endovasc. Surg. 2011, 42, 107–114. [Google Scholar] [CrossRef]
- Verajankorva, E.; Rautio, R.; Giordano, S.; Koskivuo, I.; Savolainen, O. The Efficiency of Sclerotherapy in the Treatment of Vascular Malformations: A Retrospective Study of 63 Patients. Plast. Surg. Int. 2016, 2016, 2809152. [Google Scholar] [CrossRef]
- Vuylsteke, M.E.; Martinelli, T.; Van Dorpe, J.; Roelens, J.; Mordon, S.; Fourneau, I. Endovenous laser ablation: The role of intraluminal blood. Eur. J. Vasc. Endovasc. Surg. 2011, 42, 120–126. [Google Scholar] [CrossRef]
- Feldo, M.; Wojciak-Kosior, M.; Sowa, I.; Kocki, J.; Bogucki, J.; Zubilewicz, T.; Kesik, J.; Bogucka-Kocka, A. Effect of Diosmin Administration in Patients with Chronic Venous Disorders on Selected Factors Affecting Angiogenesis. Molecules 2019, 24, 3316. [Google Scholar] [CrossRef] [PubMed]
- Potente, M.; Gerhardt, H.; Carmeliet, P. Basic and therapeutic aspects of angiogenesis. Cell 2011, 146, 873–887. [Google Scholar] [CrossRef]
- Herouy, Y.; Kreis, S.; Mueller, T.; Duerk, T.; Martiny-Baron, G.; Reusch, P.; May, F.; Idzko, M.; Norgauer, Y. Inhibition of angiogenesis in lipodermatosclerosis: Implication for venous ulcer formation. Int. J. Mol. Med. 2009, 24, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Glowinski, J.; Glowinski, S. Generation of reactive oxygen metabolites by the varicose vein wall. Eur. J. Vasc. Endovasc. Surg. 2002, 23, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Saharay, M.; Shields, D.A.; Porter, J.B.; Scurr, J.H.; Coleridge Smith, P.D. Leukocyte activity in the microcirculation of the leg in patients with chronic venous disease. J. Vasc. Surg. 1997, 26, 265–273. [Google Scholar] [CrossRef]
- Myohanen, T.T.; Tenorio-Laranga, J.; Jokinen, B.; Vazquez-Sanchez, R.; Moreno-Baylach, M.J.; Garcia-Horsman, J.A.; Mannisto, P.T. Prolyl oligopeptidase induces angiogenesis both in vitro and in vivo in a novel regulatory manner. Br. J. Pharm. 2011, 163, 1666–1678. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Wang, J.; He, L.N.; Li, B.H.; Ding, Y.N.; Chen, Y.W.; Fan, J.G. Prolyl oligopeptidase attenuates hepatic stellate cell activation through induction of Smad7 and PPAR-gamma. Exp. Med. 2017, 13, 780–786. [Google Scholar] [CrossRef]
- Brandt, I.; Scharpe, S.; Lambeir, A.M. Suggested functions for prolyl oligopeptidase: A puzzling paradox. Clin. Chim. Acta. 2007, 377, 50–61. [Google Scholar] [CrossRef]
- Boersma, D.; van Haelst, S.T.; van Eekeren, R.R.; Vink, A.; Reijnen, M.M.; de Vries, J.P.; de Borst, G.J. Macroscopic and Histologic Analysis of Vessel Wall Reaction After Mechanochemical Endovenous Ablation Using the ClariVein OC Device in an Animal Model. Eur. J. Vasc. Endovasc. Surg. 2017, 53, 290–298. [Google Scholar] [CrossRef][Green Version]
- Svarcbahs, R.; Julku, U.H.; Myohanen, T.T. Inhibition of Prolyl Oligopeptidase Restores Spontaneous Motor Behavior in the alpha-Synuclein Virus Vector-Based Parkinson’s Disease Mouse Model by Decreasing alpha-Synuclein Oligomeric Species in Mouse Brain. J. Neurosci. 2016, 36, 12485–12497. [Google Scholar] [CrossRef]
- Eschrich, J.; Meyer, R.; Kuk, H.; Wagner, A.H.; Noppeney, T.; Debus, S.; Hecker, M.; Korff, T. Varicose Remodeling of Veins Is Suppressed by 3-Hydroxy-3-Methylglutaryl Coenzyme A Reductase Inhibitors. J. Am. Heart Assoc. 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Somers, P.; Knaapen, M. The histopathology of varicose vein disease. Angiology 2006, 57, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.J.; Chen, P.R.; Huang, Y.L.; Hsu, H.H. Unique wreath-like smooth muscle proliferation of the pulmonary vasculature in pulmonary veno-occlusive disease versus pulmonary arterial hypertension. J. Med. Assoc. 2020, 119, 300–309. [Google Scholar] [CrossRef]
- Vass, D.G.; Ainsworth, R.; Anderson, J.H.; Murray, D.; Foulis, A.K. The value of an elastic tissue stain in detecting venous invasion in colorectal cancer. J. Clin. Pathol. 2004, 57, 769–772. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, R.; Fusco, R.; Gugliandolo, E.; Crupi, R.; Evangelista, M.; Granese, R.; Cuzzocrea, S. Co-micronized Palmitoylethanolamide/Polydatin Treatment Causes Endometriotic Lesion Regression in a Rodent Model of Surgically Induced Endometriosis. Front Pharm. 2016, 7, 382. [Google Scholar] [CrossRef] [PubMed]
- Casili, G.; Lanza, M.; Filippone, A.; Campolo, M.; Paterniti, I.; Cuzzocrea, S.; Esposito, E. Dimethyl fumarate alleviates the nitroglycerin (NTG)-induced migraine in mice. J. Neuroinflammation 2020, 17, 59. [Google Scholar] [CrossRef] [PubMed]
- Lanza, M.; Campolo, M.; Casili, G.; Filippone, A.; Paterniti, I.; Cuzzocrea, S.; Esposito, E. Sodium Butyrate Exerts Neuroprotective Effects in Spinal Cord Injury. Mol. Neurobiol. 2019, 56, 3937–3947. [Google Scholar] [CrossRef]
- Campolo, M.; Casili, G.; Paterniti, I.; Filippone, A.; Lanza, M.; Ardizzone, A.; Scuderi, S.A.; Cuzzocrea, S.; Esposito, E. Effect of a Product Containing Xyloglucan and Pea Protein on a Murine Model of Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 3596. [Google Scholar] [CrossRef]
- Casili, G.; Campolo, M.; Lanza, M.; Filippone, A.; Scuderi, S.; Messina, S.; Ardizzone, A.; Esposito, E.; Paterniti, I. Role of ABT888, a Novel Poly(ADP-Ribose) Polymerase (PARP) Inhibitor in Countering Autophagy and Apoptotic Processes Associated to Spinal Cord Injury. Mol. Neurobiol. 2020, 57, 4394–4407. [Google Scholar] [CrossRef]
- Porter, J.M.; Moneta, G.L. Reporting standards in venous disease: An update. International Consensus Committee on Chronic Venous Disease. J. Vasc. Surg. 1995, 21, 635–645. [Google Scholar] [CrossRef]
- Castro-Ferreira, R.; Cardoso, R.; Leite-Moreira, A.; Mansilha, A. The Role of Endothelial Dysfunction and Inflammation in Chronic Venous Disease. Ann. Vasc. Surg. 2018, 46, 380–393. [Google Scholar] [CrossRef] [PubMed]
- Guardiola, S.; Prades, R.; Mendieta, L.; Brouwer, A.J.; Streefkerk, J.; Nevola, L.; Tarrago, T.; Liskamp, R.M.J.; Giralt, E. Targeted Covalent Inhibition of Prolyl Oligopeptidase (POP): Discovery of Sulfonylfluoride Peptidomimetics. Cell Chem. Biol. 2018, 25, 1031–1037 e1034. [Google Scholar] [CrossRef] [PubMed]
- Ligi, D.; Croce, L.; Mosti, G.; Raffetto, J.D.; Mannello, F. Chronic Venous Insufficiency: Transforming Growth Factor-beta Isoforms and Soluble Endoglin Concentration in Different States of Wound Healing. Int. J. Mol. Sci. 2017, 18, 2206. [Google Scholar] [CrossRef] [PubMed]
- Sugden, W.W.; Meissner, R.; Aegerter-Wilmsen, T.; Tsaryk, R.; Leonard, E.V.; Bussmann, J.; Hamm, M.J.; Herzog, W.; Jin, Y.; Jakobsson, L.; et al. Endoglin controls blood vessel diameter through endothelial cell shape changes in response to haemodynamic cues. Nat. Cell Biol. 2017, 19, 653–665. [Google Scholar] [CrossRef]
- Ruiz-Ortega, M.; Rodriguez-Vita, J.; Sanchez-Lopez, E.; Carvajal, G.; Egido, J. TGF-beta signaling in vascular fibrosis. Cardiovasc. Res. 2007, 74, 196–206. [Google Scholar] [CrossRef]
- Gordon, K.J.; Blobe, G.C. Role of transforming growth factor-beta superfamily signaling pathways in human disease. Biochim. Biophys. Acta 2008, 1782, 197–228. [Google Scholar] [CrossRef]
- Sola Ldel, R.; Aceves, M.; Duenas, A.I.; Gonzalez-Fajardo, J.A.; Vaquero, C.; Crespo, M.S.; Garcia-Rodriguez, C. Varicose veins show enhanced chemokine expression. Eur. J. Vasc. Endovasc. Surg. 2009, 38, 635–641. [Google Scholar] [CrossRef]
- Heidemann, J.; Ogawa, H.; Dwinell, M.B.; Rafiee, P.; Maaser, C.; Gockel, H.R.; Otterson, M.F.; Ota, D.M.; Lugering, N.; Domschke, W.; et al. Angiogenic effects of interleukin 8 (CXCL8) in human intestinal microvascular endothelial cells are mediated by CXCR2. J. Biol. Chem. 2003, 278, 8508–8515. [Google Scholar] [CrossRef]
- Surendran, S.; Ramegowda, K.S.; Suresh, A.; Binil Raj, S.S.; Lakkappa, R.K.B.; Kamalapurkar, G.; Radhakrishnan, N.; Chandrasekharan, C.K. Arterialization and anomalous vein wall remodeling in varicose veins is associated with upregulated FoxC2-Dll4 pathway. Lab. Invest. 2016, 96, 399–408. [Google Scholar] [CrossRef]
- Papapetropoulos, A.; Garcia-Cardena, G.; Madri, J.A.; Sessa, W.C. Nitric oxide production contributes to the angiogenic properties of vascular endothelial growth factor in human endothelial cells. J. Clin. Invest. 1997, 100, 3131–3139. [Google Scholar] [CrossRef]
- Marumo, T.; Schini-Kerth, V.B.; Busse, R. Vascular endothelial growth factor activates nuclear factor-kappaB and induces monocyte chemoattractant protein-1 in bovine retinal endothelial cells. Diabetes 1999, 48, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Grudzinska, E.; Czuba, Z.P. Immunological aspects of chronic venous disease pathogenesis. Cent. Eur. J. Immunol. 2014, 39, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.D. Neutrophil activation and mediators of inflammation in chronic venous insufficiency. J. Vasc. Res. 1999, 36, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Wali, M.A.; Eid, R.A. Changes of elastic and collagen fibers in varicose veins. Int. Angiol. 2002, 21, 337–343. [Google Scholar]
- Venturi, M.; Bonavina, L.; Annoni, F.; Colombo, L.; Butera, C.; Peracchia, A.; Mussini, E. Biochemical assay of collagen and elastin in the normal and varicose vein wall. J. Surg. Res. 1996, 60, 245–248. [Google Scholar] [CrossRef]
- Kakkos, S.K.; Zolota, V.G.; Peristeropoulou, P.; Apostolopoulou, A.; Geroukalos, G.; Tsolakis, I.A. Increased mast cell infiltration in familial varicose veins: Pathogenetic implications? Int. Angiol. 2003, 22, 43–49. [Google Scholar]
- Yamada, T.; Tomita, S.; Mori, M.; Sasatomi, E.; Suenaga, E.; Itoh, T. Increased mast cell infiltration in varicose veins of the lower limbs: A possible role in the development of varices. Surgery 1996, 119, 494–497. [Google Scholar] [CrossRef]
- Shoab, S.S.; Scurr, J.H.; Coleridge-Smith, P.D. Increased plasma vascular endothelial growth factor among patients with chronic venous disease. J. Vasc. Surg. 1998, 28, 535–540. [Google Scholar] [CrossRef]
- Tian, H.; Huang, J.J.; Golzio, C.; Gao, X.; Hector-Greene, M.; Katsanis, N.; Blobe, G.C. Endoglin interacts with VEGFR2 to promote angiogenesis. Faseb J. 2018, 32, 2934–2949. [Google Scholar] [CrossRef]
- Beebe-Dimmer, J.L.; Pfeifer, J.R.; Engle, J.S.; Schottenfeld, D. The epidemiology of chronic venous insufficiency and varicose veins. Ann. Epidemiol. 2005, 15, 175–184. [Google Scholar] [CrossRef]
- Davies, A.H. The Seriousness of Chronic Venous Disease: A Review of Real-World Evidence. Adv. Ther. 2019, 36, 5–12. [Google Scholar] [CrossRef] [PubMed]
- McLafferty, R.B.; Passman, M.A.; Caprini, J.A.; Rooke, T.W.; Markwell, S.A.; Lohr, J.M.; Meissner, M.H.; Eklof, B.G.; Wakefield, T.W.; Dalsing, M.C. Increasing awareness about venous disease: The American Venous Forum expands the National Venous Screening Program. J. Vasc. Surg. 2008, 48, 394–399. [Google Scholar] [CrossRef] [PubMed]
- McGuinness, C.L.; Humphries, J.; Waltham, M.; Burnand, K.G.; Collins, M.; Smith, A. Recruitment of labelled monocytes by experimental venous thrombi. Thromb. Haemost. 2001, 85, 1018–1024. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.A.; Pearson, C.I. Angiogenesis in the pathogenesis of inflammatory joint and lung diseases. Arthritis Res. 2001, 3, 147–153. [Google Scholar] [CrossRef]
- Gaggar, A.; Jackson, P.L.; Noerager, B.D.; O’Reilly, P.J.; McQuaid, D.B.; Rowe, S.M.; Clancy, J.P.; Blalock, J.E. A novel proteolytic cascade generates an extracellular matrix-derived chemoattractant in chronic neutrophilic inflammation. J. Immunol. 2008, 180, 5662–5669. [Google Scholar] [CrossRef]
- Tenorio-Laranga, J.; Coret-Ferrer, F.; Casanova-Estruch, B.; Burgal, M.; Garcia-Horsman, J.A. Prolyl oligopeptidase is inhibited in relapsing-remitting multiple sclerosis. J. Neuroinflammation 2010, 7, 23. [Google Scholar] [CrossRef]
- Venalainen, J.I.; Juvonen, R.O.; Garcia-Horsman, J.A.; Wallen, E.A.; Christiaans, J.A.; Jarho, E.M.; Gynther, J.; Mannisto, P.T. Slow-binding inhibitors of prolyl oligopeptidase with different functional groups at the P1 site. Biochem. J. 2004, 382, 1003–1008. [Google Scholar] [CrossRef]
- Han, G.; Li, F.; Singh, T.P.; Wolf, P.; Wang, X.J. The pro-inflammatory role of TGFbeta1: A paradox? Int. J. Biol. Sci. 2012, 8, 228–235. [Google Scholar] [CrossRef]
- Serralheiro, P.; Soares, A.; Costa Almeida, C.M.; Verde, I. TGF-beta1 in Vascular Wall Pathology: Unraveling Chronic Venous Insufficiency Pathophysiology. Int. J. Mol. Sci. 2017, 18, 2534. [Google Scholar] [CrossRef]
- Kofler, S.; Nickel, T.; Weis, M. Role of cytokines in cardiovascular diseases: A focus on endothelial responses to inflammation. Clin. Sci. (London) 2005, 108, 205–213. [Google Scholar] [CrossRef]
- Li, A.; Dubey, S.; Varney, M.L.; Dave, B.J.; Singh, R.K. IL-8 directly enhanced endothelial cell survival, proliferation, and matrix metalloproteinases production and regulated angiogenesis. J. Immunol. 2003, 170, 3369–3376. [Google Scholar] [CrossRef] [PubMed]
- Sansilvestri-Morel, P.; Rupin, A.; Badier-Commander, C.; Fabiani, J.N.; Verbeuren, T.J. Chronic venous insufficiency: Dysregulation of collagen synthesis. Angiology 2003, 54, S13–S18. [Google Scholar] [CrossRef] [PubMed]
- Barasch, E.; Gottdiener, J.S.; Aurigemma, G.; Kitzman, D.W.; Han, J.; Kop, W.J.; Tracy, R.P. The relationship between serum markers of collagen turnover and cardiovascular outcome in the elderly: The Cardiovascular Health Study. Circ. Heart Fail 2011, 4, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Sprague, A.H.; Khalil, R.A. Inflammatory cytokines in vascular dysfunction and vascular disease. Biochem. Pharm. 2009, 78, 539–552. [Google Scholar] [CrossRef]
- Takase, S.; Bergan, J.J.; Schmid-Schonbein, G. Expression of adhesion molecules and cytokines on saphenous veins in chronic venous insufficiency. Ann. Vasc. Surg. 2000, 14, 427–435. [Google Scholar] [CrossRef]
- Boada, J.N.; Nazco, G.J. Therapeutic effect of venotonics in chronic venous insufficiency—A meta-analysis. Clin. Drug Invest. 1999, 18, 413–432. [Google Scholar] [CrossRef]
- Novotny, K.; Campr, V. Histopathological changes to the vascular wall after treatment of great saphenous veins using n-butyl-2-cyanoacrylate. Vasa 2019, 48, 399–404. [Google Scholar] [CrossRef]
- Wali, M.A.; Dewan, M.; Eid, R.A. Histopathological changes in the wall of varicose veins. Int. Angiol. 2003, 22, 188–193. [Google Scholar]
- Hernandez-Morera, P.; Castano-Gonzalez, I.; Travieso-Gonzalez, C.M.; Mompeo-Corredera, B.; Ortega-Santana, F. Quantification and Statistical Analysis Methods for Vessel Wall Components from Stained Images with Masson’s Trichrome. PLoS ONE 2016, 11, e0146954. [Google Scholar] [CrossRef]
- Satoh, T.; Sugama, K.; Matsuo, A.; Kato, S.; Ito, S.; Hatanaka, M.; Sasaguri, Y. Histamine as an activator of cell growth and extracellular matrix reconstruction for human vascular smooth muscle cells. Atherosclerosis 1994, 110, 53–61. [Google Scholar] [CrossRef]
- Lu, X.; Gong, J.; Dennery, P.A.; Yao, H. Endothelial-to-mesenchymal transition: Pathogenesis and therapeutic targets for chronic pulmonary and vascular diseases. Biochem. Pharm. 2019, 168, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Kovacic, J.C.; Dimmeler, S.; Harvey, R.P.; Finkel, T.; Aikawa, E.; Krenning, G.; Baker, A.H. Endothelial to Mesenchymal Transition in Cardiovascular Disease: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2019, 73, 190–209. [Google Scholar] [CrossRef] [PubMed]
- ten Dijke, P.; Goumans, M.J.; Pardali, E. Endoglin in angiogenesis and vascular diseases. Angiogenesis 2008, 11, 79–89. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casili, G.; Lanza, M.; Scuderi, S.A.; Messina, S.; Paterniti, I.; Campolo, M.; Esposito, E. The Inhibition of Prolyl Oligopeptidase as New Target to Counteract Chronic Venous Insufficiency: Findings in a Mouse Model. Biomedicines 2020, 8, 604. https://doi.org/10.3390/biomedicines8120604
Casili G, Lanza M, Scuderi SA, Messina S, Paterniti I, Campolo M, Esposito E. The Inhibition of Prolyl Oligopeptidase as New Target to Counteract Chronic Venous Insufficiency: Findings in a Mouse Model. Biomedicines. 2020; 8(12):604. https://doi.org/10.3390/biomedicines8120604
Chicago/Turabian StyleCasili, Giovanna, Marika Lanza, Sarah Adriana Scuderi, Salvatore Messina, Irene Paterniti, Michela Campolo, and Emanuela Esposito. 2020. "The Inhibition of Prolyl Oligopeptidase as New Target to Counteract Chronic Venous Insufficiency: Findings in a Mouse Model" Biomedicines 8, no. 12: 604. https://doi.org/10.3390/biomedicines8120604
APA StyleCasili, G., Lanza, M., Scuderi, S. A., Messina, S., Paterniti, I., Campolo, M., & Esposito, E. (2020). The Inhibition of Prolyl Oligopeptidase as New Target to Counteract Chronic Venous Insufficiency: Findings in a Mouse Model. Biomedicines, 8(12), 604. https://doi.org/10.3390/biomedicines8120604