CMT-308, a Nonantimicrobial Chemically-Modified Tetracycline, Exhibits Anti-Melanogenic Activity by Suppression of Melanosome Export


Abstract
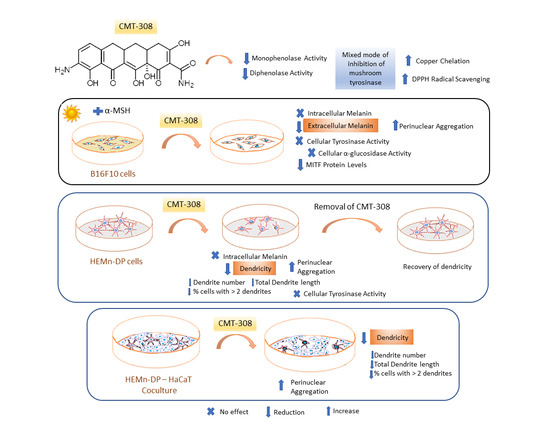
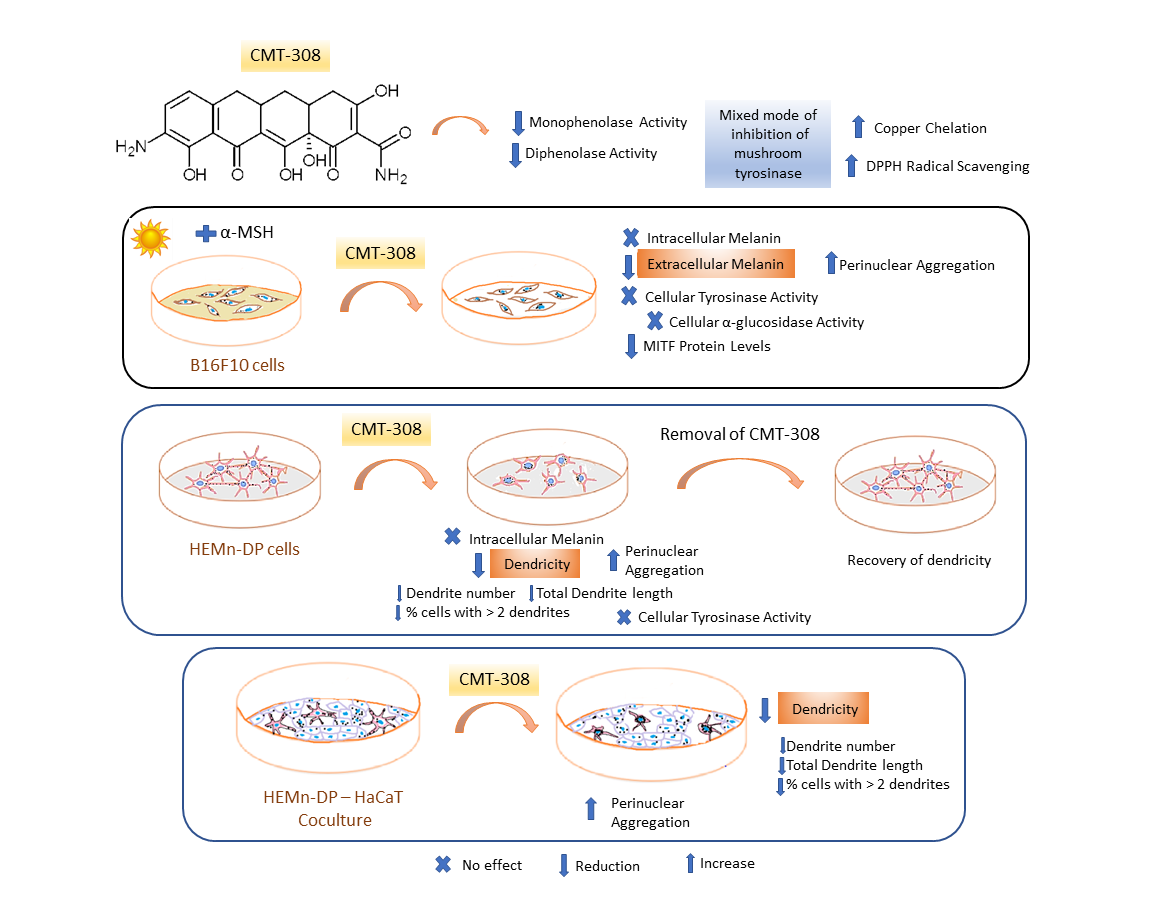{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Mushroom Tyrosinase Activity Using l-TYR and l-DOPA Substrates
2.3. Kinetic Analysis of Enzyme Inhibition
2.4. Copper Chelation Assay
2.5. α-Glucosidase Activity Assay
2.6. DPPH Radical Scavenging Assay
2.7. Cell Culture
2.8. Cytotoxicity Assay
2.9. Melanin Content Estimation
2.10. Cellular Tyrosinase Activity
2.11. Intracellular α-Glucosidase Activity in B16F10 Cells
2.12. Intracellular ROS Measurement in B16F10 Cells
2.13. Estimation of MITF Protein Levels in B16F10 Cells
2.14. Quantification of Dendricity in HEMn-DP Cells
2.15. Quantification of Recovery of Dendricity in HEMn-DP Cells
2.16. HEMn-DP Cocultures with HaCaT Cells
2.16.1. Fontana-Masson (FM) Staining
2.16.2. Quantification of Dendricity in Cocultures
2.17. Statistical Analysis
3. Results
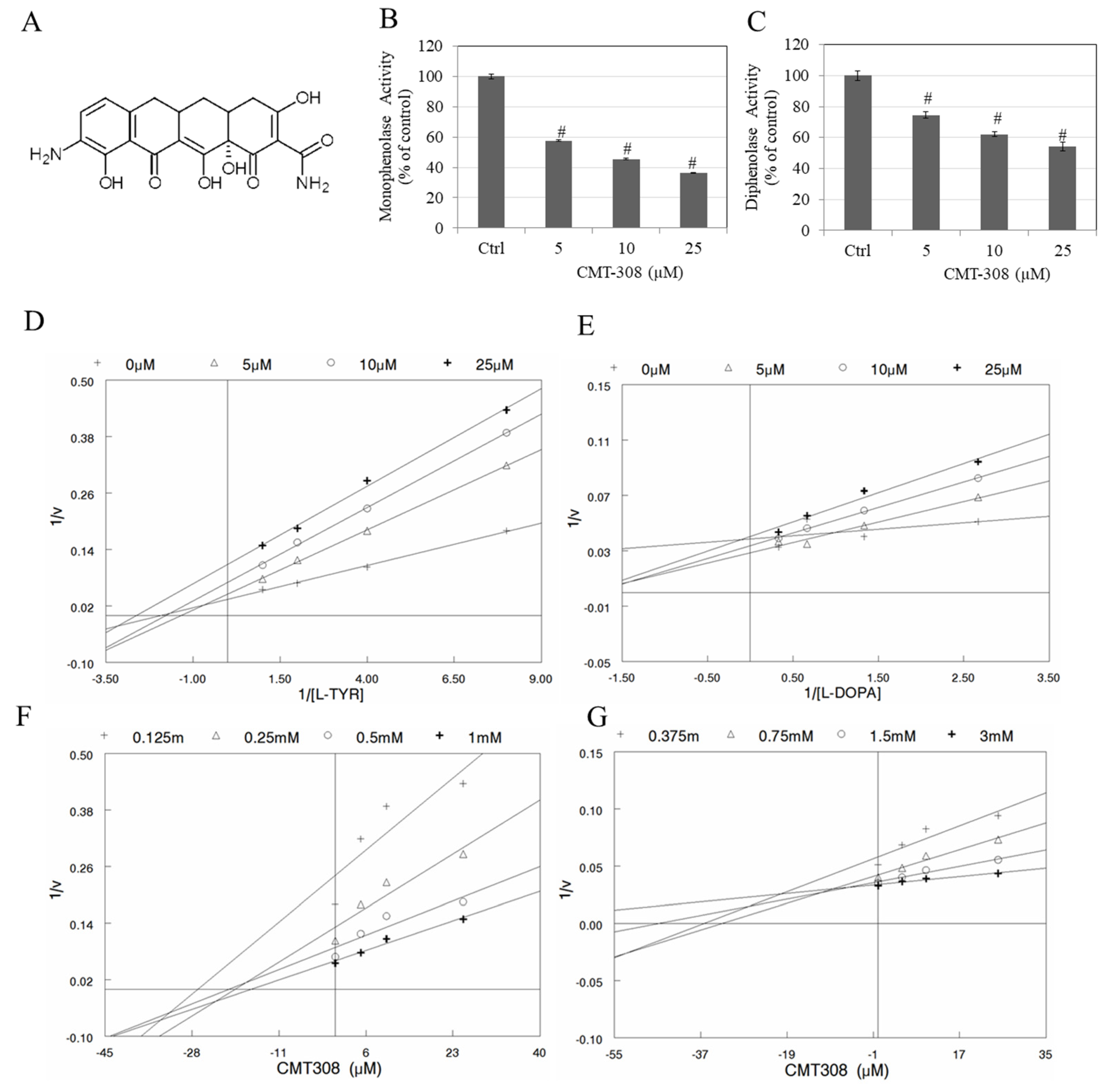
3.1. Effects of CMT-308 on the Activity of Mushroom Tyrosinase Enzyme for l-TYR and l-DOPA Substrates
3.2. Kinetic Analysis of Mushroom Tyrosinase Inhibition by CMT-308
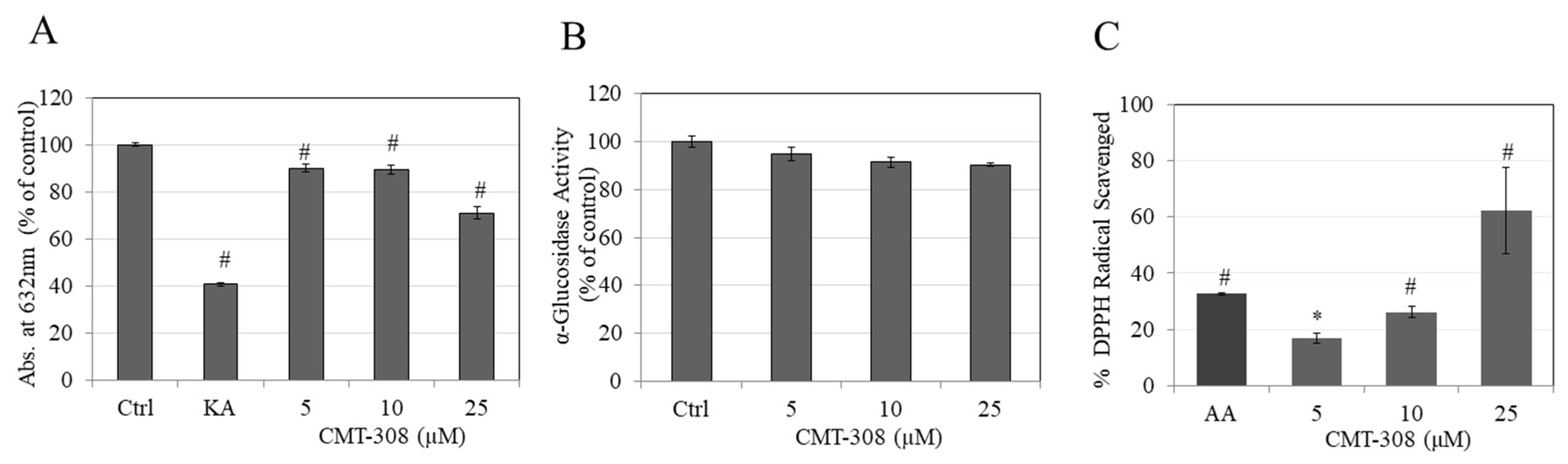
3.3. Effects of CMT-308 on Copper Chelating Capacity
3.4. Effects of CMT-308 on α-Glucosidase Activity
3.5. Effects of CMT-308 on DPPH Radical Scavenging Activity
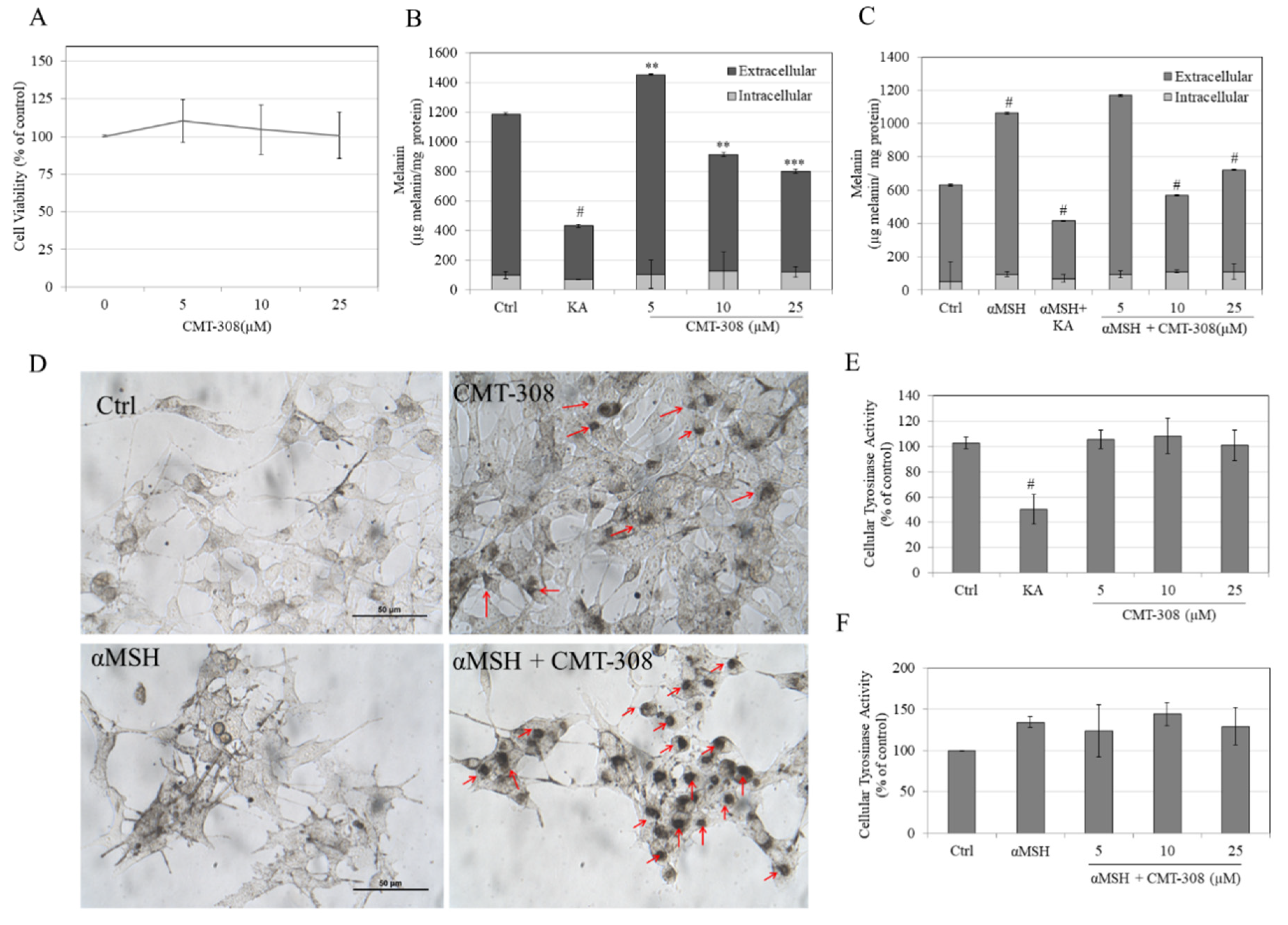
3.6. Effects of CMT-308 on Total Melanin in B16F10 Cell Cultures under Both Basal and Hormone-Stimulated Conditions
3.7. Effects of CMT-308 on α-Glucosidase Activity in B16F10 Cells
3.8. Effects of CMT-308 on Intracellular ROS Generation in B16F10 Cells
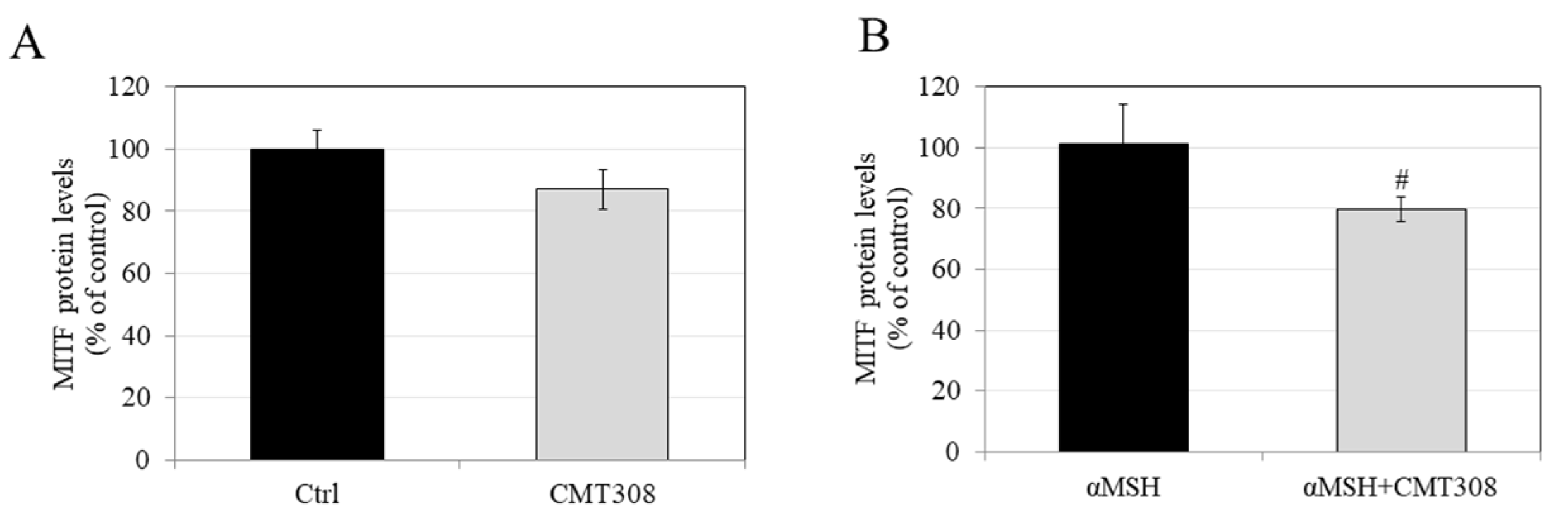
3.9. Effects of CMT-308 on MITF Protein Expression in B16F10 Cells under Hormone-Stimulated Conditions
3.10. Effects of CMT-308 on Melanin Synthesis and Cellular Tyrosinase Activity in HEMn-DP Cells
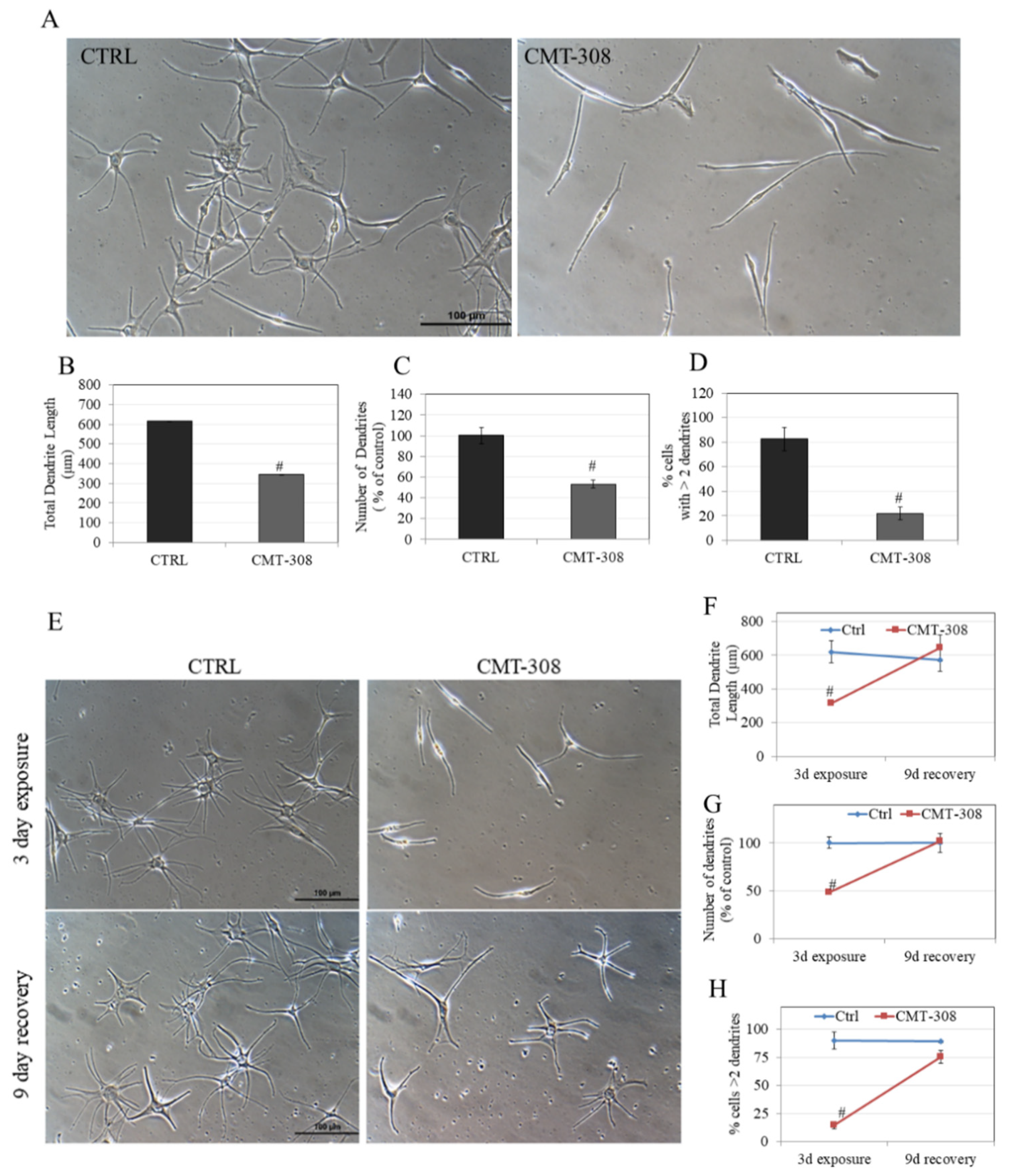
3.11. Effects of CMT-308 on Melanocyte Dendricity
3.12. Reversibility Study of Melanocyte Dendricity by CMT-308
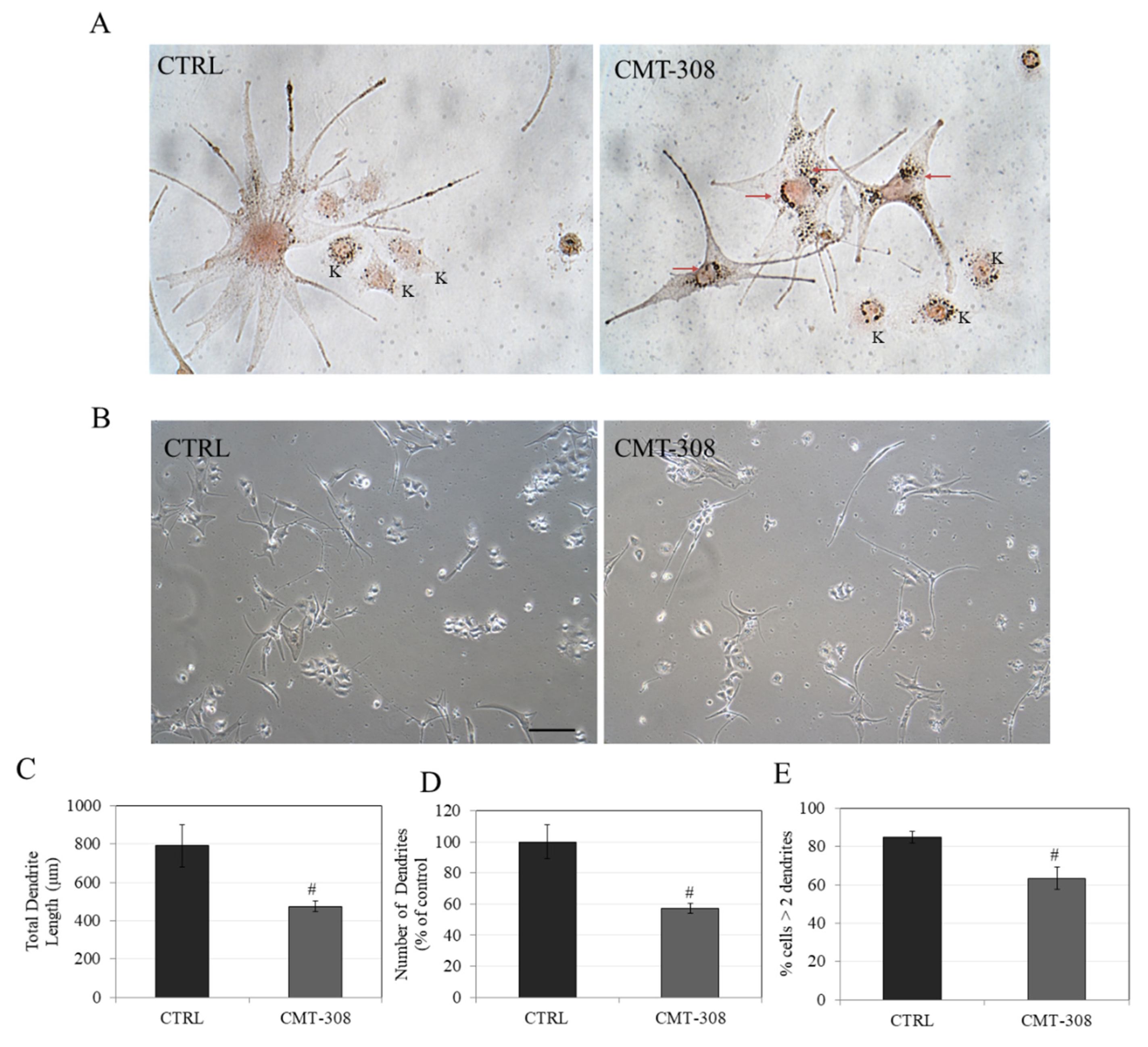
3.13. Effects of CMT-308 on Dendricity in Melanocyte Cocultures
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PIH | Post-Inflammatory Hyperpigmentation |
| LS | Lentigo Senilis |
| KA | Kojic Acid |
| HQ | Hydroquinone |
| l-DOPA | l-3,4-dihydroxyphenylalanine |
| CMT | Chemically-Modified Tetracycline |
| MMP | Matrix Metalloproteinase |
| DMEM | Dulbecco’s Modified Eagle Medium |
| HI-FBS | Heat-Inactivated Fetal Bovine Serum |
| DMSO | Dimethyl Sulfoxide |
| MSH | Melanocyte Stimulating Hormone |
| HMGS | Human Melanocyte Growth Supplement |
| HEMn-DP | Human-Epidermal Melanocytes-Darkly Pigmented |
| TDL | Total Dendrite Length |
| DCF | Dichlorofluorescein |
| ANOVA | Analysis of Variance |
| MITF | Microphthalmia Transcription Factor |
| HBSS | Hank’s Balanced Salt Solution |
| ROS | Reactive Oxygen Species |
| ELISA | Enzyme Linked Immunosorbent Assay |
| FM | Fontana-Masson |
| Ctr | Control |
| IC50 | Half-Maximal Inhibitory Concentration |
References
- Seiji, M.; Shimao, K.; Birbeck, M.S.; Fitzpatrick, T.B. Subcellular Localization of Melanin Biosynthesis. Ann. N. Y. Acad. Sci. 1963, 100, 497–533. [Google Scholar] [CrossRef]
- Brenner, M.; Hearing, V.J. The Protective Role of Melanin against UV Damage in Human Skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar]
- Solano, F. Melanins: Skin Pigments and Much More—Types, Structural Models, Biological Functions, and Formation Routes. New J. Sci. 2014, 2014, 1–28. [Google Scholar]
- Okazaki, K.; Uzuka, M.; Morikawa, F.; Toda, K.; Seiji, M. Transfer Mechanism of Melanosomes in Epidermal Cell Culture. J. Investig. Dermatol. 1976, 67, 541–547. [Google Scholar] [CrossRef]
- Millington, G.W. Proopiomelanocortin (Pomc): The Cutaneous Roles of Its Melanocortin Products and Receptors. Clin. Exp. Dermatol. 2006, 31, 407–412. [Google Scholar] [CrossRef]
- Abdel-Malek, Z.; Swope, V.B.; Suzuki, I.; Akcali, C.; Harriger, M.D.; Boyce, S.T.; Urabe, K.; Hearing, V.J. Mitogenic and Melanogenic Stimulation of Normal Human Melanocytes by Melanotropic Peptides. Proc. Natl. Acad. Sci. USA 1995, 92, 1789–1793. [Google Scholar] [CrossRef]
- Stulberg, D.L.; Clark, N.; Tovey, D. Common Hyperpigmentation Disorders in Adults: Part I. Diagnostic Approach, Cafe Au Lait Macules, Diffuse Hyperpigmentation, Sun Exposure, and Phototoxic Reactions. Am. Fam. Physician 2003, 68, 1955–1960. [Google Scholar]
- Slominski, A.; Kim, T.-K.; Brożyna, A.; Janjetovic, Z.; Brooks, D.; Schwab, L.; Skobowiat, C.; Jóźwicki, W.; Seagroves, T. The Role of Melanogenesis in Regulation of Melanoma Behavior: Melanogenesis Leads to Stimulation of Hif-1α Expression and Hif-Dependent Attendant Pathways. Arch. Biochem. Biophys. 2014, 563, 79–93. [Google Scholar] [CrossRef]
- Slominski, A.; Zbytek, B.; Slominski, R. Inhibitors of Melanogenesis Increase Toxicity of Cyclophosphamide and Lymphocytes against Melanoma Cells. Int. J. Cancer 2009, 124, 1470–1477. [Google Scholar] [CrossRef]
- Brozyna, A.A.; Jozwicki, W.; Roszkowski, K.; Filipiak, J.; Slominski, A.T. Melanin Content in Melanoma Metastases Affects the Outcome of Radiotherapy. Oncotarget 2016, 7, 17844–17853. [Google Scholar] [CrossRef]
- Brożyna, A.A.; Vanmiddlesworth, L.; Slominski, A.T. Inhibition of Melanogenesis as a Radiation Sensitizer for Melanoma Therapy. Int. J. Cancer 2008, 123, 1448–1456. [Google Scholar] [CrossRef]
- Chang, T.S. An Updated Review of Tyrosinase Inhibitors. Int. J. Mol. Sci. 2009, 10, 2440–2475. [Google Scholar] [CrossRef]
- Bae-Harboe, Y.-S.C.; Park, H.-Y. Tyrosinase: A Central Regulatory Protein for Cutaneous Pigmentation. J. Investig. Dermatol. 2012, 132, 2678–2680. [Google Scholar] [CrossRef]
- Okajima, S.; Hamamoto, A.; Asano, M.; Isogawa, K.; Ito, H.; Kato, S.; Hirata, Y.; Furuta, K.; Takemori, H. Azepine Derivative T4fat, a New Copper Chelator, Inhibits Tyrosinase. Biochem. Biophys. Res. Commun. 2019, 509, 209–215. [Google Scholar] [CrossRef]
- Song, H.; Hwang, Y.J.; Ha, J.W.; Boo, Y.C. Screening of An Epigenetic Drug Library Identifies 4-((Hydroxyamino) Carbonyl)-N-(2-Hydroxyethyl)-N-Phenyl-Benzeneacetamide That Reduces Melanin Synthesis by Inhibiting Tyrosinase Activity Independently of Epigenetic Mechanisms. Int. J. Mol. Sci. 2020, 21, 4589. [Google Scholar] [CrossRef]
- Park, Y.D.; Lyou, Y.J.; Hahn, H.S.; Hahn, M.J.; Yang, J.M. Complex Inhibition of Tyrosinase by Thiol-Composed Cu2+ Chelators: A Clue for Designing Whitening Agents. J. Biomol. Struct. Dyn. 2006, 24, 131–138. [Google Scholar] [CrossRef]
- Dolinska, M.B.; Sergeev, Y.V. The Consequences of Deglycosylation of Recombinant Intra-Melanosomal Domain of Human Tyrosinase. Biol. Chem. 2017, 399, 73–77. [Google Scholar] [CrossRef]
- Mikami, M.; Sonoki, T.; Ito, M.; Funasaka, Y.; Suzuki, T.; Katagata, Y. Glycosylation of Tyrosinase Is a Determinant of Melanin Production in Cultured Melanoma Cells. Mol. Med. Rep. 2013, 8, 818–822. [Google Scholar] [CrossRef]
- Wang, N.; Hebert, D.N. Tyrosinase Maturation through the Mammalian Secretory Pathway: Bringing Color to Life. Pigment. Cell Res. 2006, 19, 3–18. [Google Scholar]
- Bin, B.H.; Seo, J.; Yang, S.H.; Lee, E.; Choi, H.; Kim, K.H.; Cho, E.G.; Lee, T.R. Novel Inhibitory Effect of the Antidiabetic Drug Voglibose on Melanogenesis. Exp. Dermatol. 2013, 22, 541–546. [Google Scholar] [CrossRef]
- Bin, B.H.; Kim, S.T.; Bhin, J.; Byoun, K.; Lee, T.R.; Cho, E.G. The Synergistic Effect of Maltose Enhances the Anti-Melanogenic Activity of Acarbose. Arch. Dermatol. Res. 2017, 309, 217–223. [Google Scholar] [CrossRef]
- Mann, T.; Gerwat, W.; Batzer, J.; Eggers, K.; Scherner, C.; Wenck, H.; Stäb, F.; Hearing, V.J.; Röhm, K.-H.; Kolbe, L. Inhibition of Human Tyrosinase Requires Molecular Motifs Distinctively Different from Mushroom Tyrosinase. J. Investig. Dermatol. 2018, 138, 1601–1608. [Google Scholar] [CrossRef]
- Jow, T.; Hantash, B.M. Hydroquinone-Induced Depigmentation: Case Report and Review of the Literature. Dermatitis 2014, 25, E1–E5. [Google Scholar] [CrossRef]
- García-Gavín, J.; González-Vilas, D.; Fernández-Redondo, V.; Toribio, J. Pigmented Contact Dermatitis Due to Kojic Acid. A Paradoxical Side Effect of a Skin Lightener. Contact Derm. 2010, 62, 63–64. [Google Scholar] [CrossRef]
- Ando, H.; Niki, Y.; Ito, M.; Akiyama, K.; Matsui, M.S.; Yarosh, D.B.; Ichihashi, M. Melanosomes Are Transferred from Melanocytes to Keratinocytes through the Processes of Packaging, Release, Uptake, and Dispersion. J. Investig. Dermatol. 2012, 132, 1222–1229. [Google Scholar] [CrossRef]
- Tada, A.; Kanamaru, A.; Ito, Y. Control of Melanosome Transfer by Promoting Shrinkage or Expansion of Melanocyte Dendrites. Int. J. Cosmet. Sci. 2006, 28, 148. [Google Scholar] [CrossRef]
- Wu, X.; Hammer, J.A. Melanosome Transfer: It Is Best to Give and Receive. Curr. Opin. Cell Biol. 2014, 29, 1–7. [Google Scholar]
- Scott, G.A. Melanosome Trafficking and Transfer. Pigment. Syst. Physiol. Pathophysiol. 2006, 171–180. [Google Scholar]
- Kang, M.C.; Lee, J.W.; Lee, T.H.; Subedi, L.; Wahedi, H.M.; Do, S.G.; Shin, E.; Moon, E.Y.; Kim, S.Y. Up256 Inhibits Hyperpigmentation by Tyrosinase Expression/Dendrite Formation via Rho-Dependent Signaling and by Primary Cilium Formation in Melanocytes. Int. J. Mol. Sci. 2020, 21, 5341. [Google Scholar] [CrossRef]
- Lv, J.; Fu, Y.; Cao, Y.; Jiang, S.; Yang, Y.; Song, G.; Yun, C.; Gao, R. Isoliquiritigenin Inhibits Melanogenesis, Melanocyte Dendricity and Melanosome Transport by Regulating Erk-Mediated Mitf Degradation. Exp. Dermatol. 2020, 29, 149–157. [Google Scholar] [CrossRef]
- Ito, Y.; Kanamaru, A.; Tada, A. Centaureidin Promotes Dendrite Retraction of Melanocytes by Activating Rho. Biochim. Biophys. Acta 2006, 1760, 487–494. [Google Scholar] [CrossRef]
- Goenka, S.; R Simon, S. Asoprisnil, a Selective Progesterone Receptor Modulator (Sprm), Inhibits Melanosome Export in B16f10 Cells and Hemn-Dp Melanocytes. Molecules 2020, 25, 3581. [Google Scholar] [CrossRef]
- Goenka, S.; R Simon, S. Inhibitory Effects of the Bioactive Thermorubin Isolated from the Fungus Thermoactinomyces Antibioticus on Melanogenesis. Cosmetics 2020, 7, 61. [Google Scholar] [CrossRef]
- Tachibana, M. Mitf: A Stream Flowing for Pigment Cells. Pigment. Cell Res. 2000, 13, 230–240. [Google Scholar] [CrossRef]
- Yasumoto, K.; Yokoyama, K.; Takahashi, K.; Tomita, Y.; Shibahara, S. Functional Analysis of Microphthalmia-Associated Transcription Factor in Pigment Cell-Specific Transcription of the Human Tyrosinase Family Genes. J. Biol. Chem. 1997, 272, 503–509. [Google Scholar] [CrossRef]
- Chiaverini, C.; Beuret, L.; Flori, E.; Busca, R.; Abbe, P.; Bille, K.; Bahadoran, P.; Ortonne, J.P.; Bertolotto, C.; Ballotti, R. Microphthalmia-Associated Transcription Factor Regulates Rab27a Gene Expression and Controls Melanosome Transport. J. Biol. Chem. 2008, 283, 12635–12642. [Google Scholar] [CrossRef]
- Swamy, D.N.; Sanivarapu, S.; Moogla, S.; Kapalavai, V. Chemically Modified Tetracyclines: The Novel Host Modulating Agents. J. Indian Soc. Periodontol. 2015, 19, 370–374. [Google Scholar] [CrossRef]
- Golub, L.M.; Mcnamara, T.F.; D’angelo, G.; Greenwald, R.A.; Ramamurthy, N.S. A Non-Antibacterial Chemically-Modified Tetracycline Inhibits Mammalian Collagenase Activity. J. Dent. Res. 1987, 66, 1310–1314. [Google Scholar] [CrossRef]
- Golub, L.M.; Lee, H.M. Periodontal Therapeutics: Current Host-Modulation Agents and Future Directions. Periodontology 2000 2020, 82, 186–204. [Google Scholar] [CrossRef]
- Kocer, S.S.; Walker, S.G.; Zerler, B.; Golub, L.M.; Simon, S.R. Metalloproteinase Inhibitors, Nonantimicrobial Chemically Modified Tetracyclines, and Ilomastat Block Bacillus Anthracis Lethal Factor Activity in Viable Cells. Infect. Immun. 2005, 73, 7548–7557. [Google Scholar] [CrossRef]
- Simon, S.R.; Roemer, E.J.; Golub, L.M.; Ramamurthy, N.S. Serine Proteinase Inhibitory Activity by Hydrophobic Tetracycline. U.S. Patent 5,773,430, 30 June 1998. [Google Scholar]
- Lokeshwar, B.L.; Escatel, E.; Zhu, B. Cytotoxic Activity and Inhibition of Tumor Cell Invasion by Derivatives of a Chemically Modified Tetracycline Cmt-3 (Col-3). Curr. Med. Chem. 2001, 8, 271–279. [Google Scholar] [CrossRef]
- Lokeshwar, B.L. Chemically Modified Non-Antimicrobial Tetracyclines Are Multifunctional Drugs Against Advanced Cancers. Pharmacol. Res. 2011, 63, 146–150. [Google Scholar] [CrossRef]
- Sandler, C.; Nurmi, K.; Lindstedt, K.A.; Sorsa, T.; Golub, L.M.; Kovanen, P.T.; Eklund, K.K. Chemically Modified Tetracyclines Induce Apoptosis in Cultured Mast Cells. Int. Immunopharmacol. 2005, 5, 1611–1621. [Google Scholar] [CrossRef]
- Adisen, E.; Gurer, M.A.; Erdem, O. Tetracycline/Doxycycline-Induced Cutaneous Depressed Pigmentation. Int. J. Dermatol. 2006, 45, 1245–1247. [Google Scholar] [CrossRef]
- Bohm, M.; Schmidt, P.F.; Lodding, B.; Uphoff, H.; Westermann, G.; Luger, T.A.; Bonsmann, G.; Metze, D. Cutaneous Hyperpigmentation Induced by Doxycycline: Histochemical and Ultrastructural Examination, Laser Microprobe Mass Analysis, and Cathodoluminescence. Am. J. Dermatopathol. 2002, 24, 345–350. [Google Scholar] [CrossRef]
- Lee, H.; Zerler, B.; Liu, Y.; Roemer, E.; Simon, S.; Ryan, M.; Golub, L. Biologic Efficacy of a New Chemically-Modified Non-Antimicrobial Tetracycline: Cmt-308. J. Dent. Res. 2002, 81, A126. [Google Scholar]
- Zhang, Y.; Gu, Y.; Lee, H.M.; Hambardjieva, E.; Vrankova, K.; Golub, L.M.; Johnson, F. Design, Synthesis And Biological Activity of New Polyenolic Inhibitors of Matrix Metalloproteinases: A Focus on Chemically-Modified Curcumins. Curr. Med. Chem. 2012, 19, 4348–4358. [Google Scholar] [CrossRef]
- Zhang, Y.; M Golub, L.; Johnson, F.; Wishnia, A. Pka, Zinc-And Serum Albumin-Binding of Curcumin and Two Novel Biologically-Active Chemically-Modified Curcumins. Curr. Med. Chem. 2012, 19, 4367–4375. [Google Scholar] [CrossRef]
- Burk, D.; Lineweaver, H.; Horner, C.K. The Specific Influence of Acidity on the Mechanism of Nitrogen Fixation by Azotobacter. J. Bacteriol. 1934, 27, 325–340. [Google Scholar] [CrossRef]
- Goenka, S.; Ceccoli, J.; Simon, S.R. Anti-Melanogenic Activity of Ellagitannin Casuarictin in B16f10 Mouse Melanoma Cells. Nat. Prod. Res. 2019, 1–6. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The Chemistry Behind Antioxidant Capacity Assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Chan, Y.Y.; Kim, K.H.; Cheah, S.H. Inhibitory Effects of Sargassum Polycystum on Tyrosinase Activity and Melanin Formation in B16f10 Murine Melanoma Cells. J. Ethnopharmacol. 2011, 137, 1183–1188. [Google Scholar] [CrossRef]
- Sim, M.-O.; Choi, I.-Y.; Cho, J.-H.; Shin, H.-M.; Cho, H.-W. Anti-Melanogenesis And Anti-Oxidant of Salix Pseudo-Lasiogyne Water Extract in A-Msh-Induced B16f10 Melanoma Cells. Food Agric. Immunol. 2017, 28, 1003–1016. [Google Scholar] [CrossRef]
- Mclennan, H.R.; Degli Esposti, M. The Contribution of Mitochondrial Respiratory Complexes to the Production of Reactive Oxygen Species. J. Bioenerg. Biomembr. 2000, 32, 153–162. [Google Scholar] [CrossRef]
- Chang, C.H.; Tsai, R.K.; Tsai, M.H.; Lin, Y.H.; Hirobe, T. The Roles of Frizzled-3 and Wnt3a on Melanocyte Development: In Vitro Studies on Neural Crest Cells and Melanocyte Precursor Cell Lines. J. Dermatol. Sci. 2014, 75, 100–108. [Google Scholar] [CrossRef]
- Ohbayashi, N.; Yatsu, A.; Tamura, K.; Fukuda, M. The Rab21-Gef Activity of Varp, But Not Its Rab32/38 Effector Function, Is Required for Dendrite Formation in Melanocytes. Mol. Biol. Cell 2012, 23, 669–678. [Google Scholar]
- Scott, G.; Fricke, A.; Fender, A.; Mcclelland, L.; Jacobs, S. Prostaglandin E2 Regulates Melanocyte Dendrite Formation through Activation of Pkcζ. Exp. Cell Res. 2007, 313, 3840–3850. [Google Scholar] [CrossRef]
- Joshi, P.G.; Nair, N.; Begum, G.; Joshi, N.B.; Sinkar, V.P.; Vora, S. Melanocyte-Keratinocyte Interaction Induces Calcium Signalling and Melanin Transfer to Keratinocytes. Pigment Cell Res. 2007, 20, 380–384. [Google Scholar] [CrossRef]
- Ando, H.; Niki, Y.; Yoshida, M.; Ito, M.; Akiyama, K.; Kim, J.H.; Yoon, T.J.; Lee, J.H.; Matsui, M.S.; Ichihashi, M. Keratinocytes in Culture Accumulate Phagocytosed Melanosomes in the Perinuclear Area. Pigment Cell Melanoma Res. 2010, 23, 129–133. [Google Scholar] [CrossRef]
- Diwakar, G.; Klump, V.; Lazova, R.; Pawelek, J. Evidence for Glycosylation as a Regulator of the Pigmentary System: Key Roles of Sialyl (A2-6) Gal/Galnac-Terminated Glycans in Melanin Synthesis and Transfer. Glycoconj. J. 2015, 32, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.L. Chemical and Biological Dynamics of Tetracyclines. Adv. Dent. Res. 1998, 12, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Golub, L.; Suomalainen, K.; Sorsa, T. Host Modulation with Tetracyclines and Their Chemically Modified Analogues. Curr. Opin. Dent. 1992, 2, 80. [Google Scholar] [PubMed]
- Sorsa, T.; Ramamurthy, N.; Vernillo, A.; Zhang, X.; Konttinen, Y.T.; Rifkin, B.; Golub, L. Functional Sites of Chemically Modified Tetracyclines: Inhibition of the Oxidative Activation of Human Neutrophil and Chicken Osteoclast Pro-Matrix Metalloproteinases. J. Rheumatol. 1998, 25, 975–982. [Google Scholar] [PubMed]
- Kraus, R.L.; Pasieczny, R.; Lariosa-Willingham, K.; Turner, M.S.; Jiang, A.; Trauger, J.W. Antioxidant Properties of Minocycline: Neuroprotection in an Oxidative Stress Assay and Direct Radical-Scavenging Activity. J. Neurochem. 2005, 94, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Jastreboff, P.; Pawelek, J. l-Tyrosine Stimulates Induction of Tyrosinase Activity by Msh and Reduces Cooperative Interactions between Msh Receptors in Hamster Melanoma Cells. Biosci. Rep. 1989, 9, 579–586. [Google Scholar] [CrossRef]
- Slominski, A.; Moellmann, G.; Kuklinska, E.; Bomirski, A.; Pawelek, J. Positive Regulation of Melanin Pigmentation by Two Key Substrates of the Melanogenic Pathway, l-Tyrosine and l-Dopa. J. Cell Sci. 1988, 89, 287–296. [Google Scholar]
- Slominski, A.; Zmijewski, M.A.; Pawelek, J. L-Tyrosine and L-Dihydroxyphenylalanine as Hormone-Like Regulators of Melanocyte Functions. Pigment Cell Melanoma Res. 2012, 25, 14–27. [Google Scholar]
- Liu, Y.; Ramamurthy, N.; Marecek, J.; Lee, H.M.; Chen, J.L.; Ryan, M.E.; Rifkin, B.R.; Golub, L.M. The Lipophilicity, Pharmacokinetics, and Cellular Uptake of Different Chemically-Modified Tetracyclines (Cmts). Curr. Med. Chem. 2001, 8, 243–252. [Google Scholar] [CrossRef]
- Hearing, V.J., Jr.; Ekel, T.M.; Montague, P.M.; Nicholson, J.M. Mammalin Tyrosinase. Stoichiometry and Measurement of Reaction Products. Biochim. Biophys. Acta (BBA)-Enzymol. 1980, 611, 251–268. [Google Scholar] [CrossRef]
- Oki, T.; Matsui, T.; Osajima, Y. Inhibitory Effect of A-Glucosidase Inhibitors Varies According to Its Origin. J. Agric. Food Chem. 1999, 47, 550–553. [Google Scholar] [CrossRef]
- Chung, S.; Lim, G.J.; Lee, J.Y. Quantitative Analysis of Melanin Content in a Three-Dimensional Melanoma Cell Culture. Sci. Rep. 2019, 9, 1–9. [Google Scholar]
- Myung, C.H.; Kim, K.; Park, J.I.; Lee, J.E.; Lee, J.A.; Hong, S.C.; Lim, K.M.; Hwang, J.S. 16-Kauren-2-Beta-18,19-Triol Inhibits Melanosome Transport in Melanocytes by Down-Regulation of Melanophilin Expression. J. Dermatol. Sci. 2020, 97, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Il Park, J.; Lee, H.Y.; Lee, J.E.; Hwan Myung, C.; Hwang, J.S. Inhibitory Effect of 2-Methyl-Naphtho [1, 2, 3-De] Quinolin-8-One on Melanosome Transport and Skin Pigmentation. Sci. Rep. 2016, 6, 1–9. [Google Scholar]
- Kudo, M.; Kobayashi-Nakamura, K.; Tsuji-Naito, K. Bifunctional Effects of O-Methylated Flavones from Scutellaria Baicalensis Georgi on Melanocytes: Inhibition of Melanin Production and Intracellular Melanosome Transport. PLoS ONE 2017, 12, e0171513. [Google Scholar] [CrossRef]
- Barral, D.C.; Seabra, M.C. The Melanosome as a Model to Study Organelle Motility in Mammals. Pigment Cell Res. 2004, 17, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Provance, D.W.; Wei, M.; Ipe, V.; Mercer, J.A. Cultured Melanocytes from Dilute Mutant Mice Exhibit Dendritic Morphology and Altered Melanosome Distribution. Proc. Natl. Acad. Sci. USA 1996, 93, 14554–14558. [Google Scholar] [CrossRef] [PubMed]
- Mayerson, P.L.; Brumbaugh, J.A. Lavender, a Chick Melanocyte Mutant with Defective Melanosome Translocation: A Possible Role for 10 nm Filaments and Microfilaments but not Microtubules. J. Cell Sci. 1981, 51, 25–51. [Google Scholar]
- Westbroek, W.; Lambert, J.; Naeyaert, J.M. The Dilute Locus and Griscelli Syndrome: Gateways towards a Better Understanding of Melanosome Transport. Pigment Cell Res. 2001, 14, 320–327. [Google Scholar] [CrossRef]
- Jung, E.; Hwang, W.; Kim, S.; Kim, Y.S.; Kim, Y.S.; Lee, J.; Park, D. Depigmenting Action of Platycodin D Depends on the Camp/Rho-Dependent Signalling Pathway. Exp. Dermatol. 2011, 20, 986–991. [Google Scholar] [CrossRef]
- Greatens, A.; Hakozaki, T.; Koshoffer, A.; Epstein, H.; Schwemberger, S.; Babcock, G.; Bissett, D.; Takiwaki, H.; Arase, S.; Wickett, R.R. Effective Inhibition of Melanosome Transfer to Keratinocytes by Lectins and Niacinamide Is Reversible. Exp. Dermatol. 2005, 14, 498–508. [Google Scholar] [CrossRef]
- Scott, G. Rac and Rho: The Story behind Melanocyte Dendrite Formation. Pigment Cell Res. 2002, 15, 322–330. [Google Scholar] [PubMed]
- Dutta, S.; Panda, S.; Singh, P.; Tawde, S.; Mishra, M.; Andhale, V.; Athavale, A.; Keswani, S.M. Hypopigmentation in Burns Is Associated with Alterations in the Architecture of the Skin and the Dendricity of the Melanocytes. Burns 2020, 46, 906–917. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhou, L.; Huang, J.; Mei, X.; Wu, Z.; Shi, W. A Preliminary Study of Growth Characteristics of Melanocytes Co-Cultured with Keratinocytes In Vitro. J. Cell Biochem. 2018, 119, 6173–6180. [Google Scholar] [CrossRef]
- Virador, V.M.; Muller, J.; Wu, X.; Abdel-Malek, Z.A.; Yu, Z.X.; Ferrans, V.J.; Kobayashi, N.; Wakamatsu, K.; Ito, S.; Hammer, J.A.; et al. Influence of Alpha-Melanocyte-Stimulating Hormone and Ultraviolet Radiation on the Transfer of Melanosomes to Keratinocytes. FASEB J. 2002, 16, 105–107. [Google Scholar] [CrossRef] [PubMed]
- Hunt, G.; Todd, C.; Thody, A.J. Unresponsiveness of Human Epidermal Melanocytes to Melanocyte-Stimulating Hormone and Its Association with Red Hair. Mol. Cell Endocrinol. 1996, 116, 131–136. [Google Scholar] [CrossRef]
- Branquinho, M.S.; Silva, M.B.; Silva, J.C.; Sales, M.C.; Barros, S.B.; Maria-Engler, S.S.; Campa, A. A 2D and 3D Melanogenesis Model with Human Primary Cells Induced by Tyrosine. J. Biol. Methods 2020, 7, e134. [Google Scholar] [CrossRef]
- Shin, Y.-H.; Seo, Y.-K.; Yoon, H.-H.; Song, K.-Y.; Park, J.-K. Effect of Keratinocytes on Regulation of Melanogenesis in Culture of Melanocytes. Biotechnol. Bioprocess. Eng. 2012, 17, 203–210. [Google Scholar]
- Duval, C.; Regnier, M.; Schmidt, R. Distinct Melanogenic Response of Human Melanocytes in Mono-Culture, in Co-Culture with Keratinocytes and in Reconstructed Epidermis, to UV Exposure. Pigment Cell Res. 2001, 14, 348–355. [Google Scholar] [CrossRef]
- Babiarz-Magee, L.; Chen, N.; Seiberg, M.; Lin, C.B. The Expression and Activation of Protease-Activated Receptor-2 Correlate with Skin Color. Pigment Cell Res. 2004, 17, 241–251. [Google Scholar] [CrossRef]
- Minwalla, L.; Zhao, Y.; Le Poole, I.C.; Wickett, R.R.; Boissy, R.E. Keratinocytes Play A Role in Regulating Distribution Patterns of Recipient Melanosomes In Vitro. J. Investig. Dermatol. 2001, 117, 341–347. [Google Scholar] [CrossRef]
- Yoshida, Y.; Hachiya, A.; Sriwiriyanont, P.; Ohuchi, A.; Kitahara, T.; Takema, Y.; Visscher, M.O.; Boissy, R.E. Functional Analysis of Keratinocytes in Skin Color Using a Human Skin Substitute Model Composed of Cells Derived from Different Skin Pigmentation Types. FASEB J. 2007, 21, 2829–2839. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, G.; Bolasco, G.; Aspite, N.; Lucania, G.; Lotti, L.V.; Torrisi, M.R.; Picardo, M. Melanosome Transfer Promoted by Keratinocyte Growth Factor in Light and Dark Skin-Derived Keratinocytes. J. Investig. Dermatol. 2008, 128, 558–567. [Google Scholar] [CrossRef] [PubMed]
- Hurbain, I.; Romao, M.; Sextius, P.; Bourreau, E.; Marchal, C.; Bernerd, F.; Duval, C.; Raposo, G. Melanosome Distribution in Keratinocytes in Different Skin Types: Melanosome Clusters Are Not Degradative Organelles. J. Investig. Dermatol. 2018, 138, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Boukamp, P.; Petrussevska, R.T.; Breitkreutz, D.; Hornung, J.; Markham, A.; Fusenig, N.E. Normal Keratinization in a Spontaneously Immortalized Aneuploid Human Keratinocyte Cell Line. J. Cell Biol. 1988, 106, 761–771. [Google Scholar]
- Colombo, I.; Sangiovanni, E.; Maggio, R.; Mattozzi, C.; Zava, S.; Corbett, Y.; Fumagalli, M.; Carlino, C.; Corsetto, P.A.; Scaccabarozzi, D. Hacat Cells as a Reliable In Vitro Differentiation Model to Dissect the Inflammatory/Repair Response of Human Keratinocytes. Mediat. Inflamm. 2017, 2017, 7435621. [Google Scholar] [CrossRef]
- Deyrieux, A.F.; Wilson, V.G. In Vitro Culture Conditions to Study Keratinocyte Differentiation Using the Hacat Cell Line. Cytotechnology 2007, 54, 77–83. [Google Scholar] [CrossRef]
- Fitzpatrick, T.B.; Breathnach, A. The Epidermal Melanin Unit System. Dermatol. Wochenschr. 1963, 147, 481. [Google Scholar]
- Rok, J.; Buszman, E.; Delijewski, M.; Otreba, M.; Beberok, A.; Wrzesniok, D. Effect of Tetracycline and UV Radiation on Melanization and Antioxidant Status of Melanocytes. J. Photochem. Photobiol. B Biol. 2015, 148, 168–173. [Google Scholar] [CrossRef]
- Rok, J.; Buszman, E.; Beberok, A.; Delijewski, M.; Otreba, M.; Wrzesniok, D. Modulation of Melanogenesis and Antioxidant Status of Melanocytes in Response To Phototoxic Action of Doxycycline. Photochem. Photobiol. 2015, 91, 1429–1434. [Google Scholar] [CrossRef]
- Rok, J.; Wrześniok, D.; Beberok, A.; Otręba, M.; Delijewski, M.; Buszman, E. Phototoxic Effect of Oxytetracycline on Normal Human Melanocytes. Toxicol. In Vitro 2018, 48, 26–32. [Google Scholar] [CrossRef]
- Rok, J.; Rzepka, Z.; Respondek, M.; Beberok, A.; Wrześniok, D. Chlortetracycline and Melanin Biopolymer—The Risk of Accumulation and Implications for Phototoxicity: An In Vitro Study on Normal Human Melanocytes. Chem.-Biol. Interact. 2019, 303, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Banning, T.P.; Heard, C.M. Binding of Doxycycline to Keratin, Melanin and Human Epidermal Tissue. Int. J. Pharm. 2002, 235, 219–227. [Google Scholar] [CrossRef]
- Seftor, R.E.; Seftor, E.A.; De Larco, J.E.; Kleiner, D.E.; Leferson, J.; Stetler-Stevenson, W.G.; Mcnamara, T.F.; Golub, L.M.; Hendrix, M.J. Chemically Modified Tetracyclines Inhibit Human Melanoma Cell Invasion and Metastasis. Clin. Exp. Metastasis 1998, 16, 217–225. [Google Scholar] [CrossRef] [PubMed]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goenka, S.; Simon, S.R. CMT-308, a Nonantimicrobial Chemically-Modified Tetracycline, Exhibits Anti-Melanogenic Activity by Suppression of Melanosome Export. Biomedicines 2020, 8, 411. https://doi.org/10.3390/biomedicines8100411
Goenka S, Simon SR. CMT-308, a Nonantimicrobial Chemically-Modified Tetracycline, Exhibits Anti-Melanogenic Activity by Suppression of Melanosome Export. Biomedicines. 2020; 8(10):411. https://doi.org/10.3390/biomedicines8100411
Chicago/Turabian StyleGoenka, Shilpi, and Sanford R. Simon. 2020. "CMT-308, a Nonantimicrobial Chemically-Modified Tetracycline, Exhibits Anti-Melanogenic Activity by Suppression of Melanosome Export" Biomedicines 8, no. 10: 411. https://doi.org/10.3390/biomedicines8100411
APA StyleGoenka, S., & Simon, S. R. (2020). CMT-308, a Nonantimicrobial Chemically-Modified Tetracycline, Exhibits Anti-Melanogenic Activity by Suppression of Melanosome Export. Biomedicines, 8(10), 411. https://doi.org/10.3390/biomedicines8100411