Abstract
Parkinson’s Disease is the second most prevalent neurological disorder globally, and its cause is still largely unknown. Likewise, there is no cure, and existing treatments do little more than subdue symptoms before becoming ineffective. It is increasingly important to understand the factors contributing to Parkinson’s Disease aetiology so that new and more effective pharmacotherapies can be established. In recent years, there has been an emergence of research linking gut dysbiosis to Parkinson’s Disease via the gut–brain axis. Advancements in microbial profiling have led to characterisation of a Parkinson’s-specific microbial signature, where novel treatments that leverage and correct gut dysbiosis are beginning to emerge for the safe and effective treatment of Parkinson’s Disease. Preliminary clinical studies investigating microbiome-targeted therapeutics for Parkinson’s Disease have revealed promising outcomes, and as such, the aim of this review is to provide a timely and comprehensive update of the most recent advances in this field. Faecal microbiota transplantation has emerged as a novel and potential frontrunner for microbial-based therapies due to their efficacy in alleviating Parkinson’s Disease symptomology through modulation of the gut–brain axis. However, more rigorous clinical investigation, along with technological advancements in diagnostic and in vitro testing tools, are critically required to facilitate the widespread clinical translation of microbiome-targeting Parkinson’s Disease therapeutics.
1. Introduction
Parkinson’s Disease (PD) is the second most prevalent neurological disorder, affecting approximately 3% of people aged over 65 globally [1]. One of the greatest risk factors for PD is age, and with the global trend towards aging populations, the number of PD sufferers is expected to double by 2030 [2]. Only 5–10% of PD is thought to be familial PD, which is heritable and caused by genetic mutations [3]. Sporadic (or idiopathic) parkinsonism is the most common form of PD that manifests as progressive protein aggregation through the formation of misfolded α-synuclein clusters, known as Lewy bodies, which accumulate in the neurons of the central nervous system (CNS), as well as the enteric nervous system (ENS) of the gastrointestinal tract (GIT) [4]. Some environmental toxins, such as certain pollutants and pesticides (such as rotenone and paraquat) or drugs (MPTP, a toxic by-product of the opioid analgesic desmethylprodine, MPPP), have been identified as agents that can cause PD [5]; however, for the vast majority of those affected by PD, the catalyst is unknown [6].
PD is characterised by motor dysfunction, such as tremor, rigidity, postural instability, and bradykinesia [7,8]. These hallmarks of PD are caused by damage to the substantia nigra pars compacta (SNc). The motor symptoms start to appear when a majority of the dopaminergic neurons are already damaged in the SNc [9], and approximately 80% of dopamine levels are depleted in the nigrostriatal terminals [10]. Unfortunately, this means PD diagnosis and treatment are delayed until substantial neurological damage has already occurred, resulting in suboptimal treatment and reduced quality of life in PD sufferers [11].
In addition to motor dysfunction, patients experience non-motor symptoms, often including cognitive or behavioural abnormalities, anxiety, depression, sleep disturbances, hyposmia/anosmia, and gastrointestinal (GI) symptoms, such as constipation and delayed gastric emptying [1,8,12]. Importantly, evidence shows that GI symptoms, especially constipation and intestinal inflammation, occur up to decades before motor symptoms [13,14]. As these symptoms are experienced by virtually all PD patients (gastric motility disorders: 70–100%, constipation: up to 90% of PD patients) [13], these symptoms are important indicators of early-stage PD and may even be implicated in the pathogenesis of the disease. The pathogenesis of PD is still poorly understood, and existing treatments for PD are inadequate, often only alleviating symptoms for a few years before becoming ineffective or causing side effects, such as dyskinesia [15]. There is currently no cure for Parkinson’s, nor any way to slow its progression. Thus, understanding PD aetiology and developing new pharmacological treatments for the disease is vital.
Recently, there has been an emergence of research into the bi-directional interaction between the brain and gut microbiome, known as the “gut–brain axis” [16]. The gut–brain axis is a complex communication network mediated by the CNS, the ENS, and the various microbe populations that inhabit the gut microbiome. Communication is maintained through the vagus nerve, as well as the immune system, tryptophan metabolism, and microbial metabolites, such as short-chain fatty acids, branched-chain amino acids, and peptidoglycans [16,17]. As research into this area expands, evidence points to the role of the gut–brain axis in neurological diseases, including PD [18]. This review updates recent findings that outline the link between gut microbiome disturbances (commonly referred to as gut dysbiosis) and PD development, progression, and severity, providing clear rationale for the development of novel microbiome-targeted therapies for the effective prevention and treatment of PD. Here, we summarise recent microbiome-targeted PD therapies in light of the role of gut dysbiosis in PD, building on other quality reviews on this topic [10,14,16,19,20,21,22]. It is envisioned that insights derived from this timely updated review will provide future directions for researchers to engineer new therapies that enable improved management of this debilitating disease with few current effective treatment options.
2. The Gut Microbiome as a Key Driver of Parkinson’s Disease
In 1913, Dr. Friederich Lewy put forth the notion of the vagus nerve as a centre of brain pathology [4]. By 2003, Professor Heiko Braak elaborated on this in the context of PD by presenting the hypothesis that an unknown pathogen in the gut is the culprit behind sporadic PD. Braak’s “dual-hit theory” suggests that this pathogen may enter the gastrointestinal tract (GIT) from the nasal passage and affects neurons in the gut and nasal cavity [6]. This pathogen leads to α-synuclein protein aggregation in the GIT, which then travels to the CNS and higher cortical regions via the vagal nerve and the olfactory tract. Subsequent studies have shown that α-synuclein deposits can be detected in GIT biopsies prior to the onset of PD, and post-mortem examinations have shown similar accumulation of these proteins inside the olfactory bulb of the CNS [23]. The pioneering work performed by Lewy and Braak has served as the foundation for the wealth of subsequent studies that explore the role of the gut in PD, with emerging evidence linking the gut microbiome with disease progression and severity. Aside from the vagal communication method highlighted by Braak’s theory, the gut microbiome may also communicate with the central nervous system in other ways, including hormonal and immune signalling. For a detailed outline of the other mechanisms, the reader is referred to Cryan et al., 2019 [17]. The purpose of this section was to review the evidence that links the gut microbiome with PD, providing an overview for the microbial signatures of PD and the mechanisms by which gut dysbiosis triggers neuroinflammation and degeneration, ultimately highlighting the rationale for gut microbiome-targeted therapies for treating PD.
3. Gut Dysbiosis as a Microbial Signature of PD
The human gut microbiome is composed of more than 100 trillion microorganisms, including bacteria, yeasts, and viruses, along with their genomes and metabolome [22]. The gut microbiome differs between individuals, and is affected by diet, antibiotics, environment, physical activity, and age [18,23]. Early life also impacts microbial composition, with birth delivery type, infant feeding practices (i.e., breast milk versus formula), and gestational age playing important roles [21,24]. In humans, Bacillota (formerly Firmicutes), Bacteroidota (formerly Bacteroidetes), Actinomycetota (formerly Actinobacteria), Pseudomonadota (formerly Proteobacteria), Fusobacteria, and Verrucomicrobia are the main phyla found in the GIT. Bacillota (formerly Firmicutes) and Bacteroidota (formerly Bacteroidetes) are typically dominant, contributing to 90% of the microbial population [22]. A healthy gut microbiome is integral to host wellbeing, with important roles in maintaining intestinal barrier integrity, function, and metabolism, as well as immune system function and regulation of the gut–brain axis [21]. Additionally, gut microbes play an important role in digestion, degrading indigestible dietary fibres via fermentation, and producing short-chain fatty acids (SCFAs) and other metabolites that are essential for host health [23].
While there is still no consensus on what the ideal microbiome composition is in a healthy human, there are distinct changes in the gut microbial colonies of PD sufferers. For example, small intestinal bacterial overgrowth (SIBO) is present in 25% of PD patients, a prevalence that is significantly higher than that in healthy adults [9]. Since technological advancements in genome sequencing have made the classification of the various microbial populations in the human gut more accurate and accessible, there has been a push to identify characteristic compositional shifts as ‘microbial signatures’ of numerous diseases, including PD. Several studies have analysed human faecal samples to highlight the key changes in taxa at the phyla, family, and genera levels, where the findings indeed indicate that there are specific microbial shifts that characterise the PD gut microbiome (Figure 1). Evidence is emerging that would link various changes in abundance of key taxa at the phyla, family, and genera levels to the onset, progression, and severity of PD. These data are summarised in Table 1, which outlines shifts in the relative abundance of microbes by taxa in PD patients compared with healthy controls. However, it is noted that many studies do not consider the impacts of age and sex on microbial composition, whether in patients or healthy controls. While differences exist in microbiota composition in PD patients across each study, there are consistent results across key genera: numerous studies show an increase in the relative abundance of Akkermansia and Bifidobacterium, and decreases in Blautia, Faecalibacterium, and Roseburia in PD faecal samples.
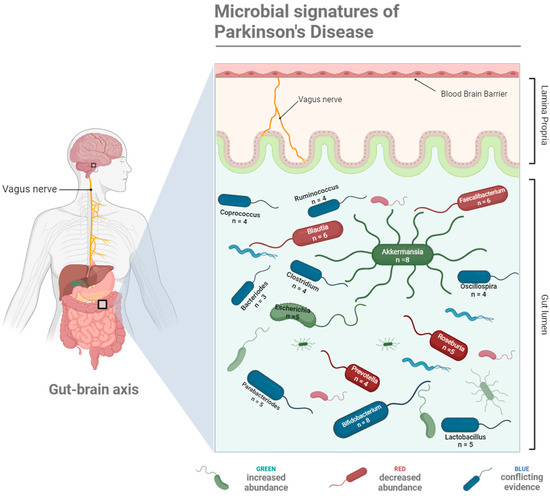
Figure 1.
A schematic representation of the key microbial taxa linked with Parkinson’s Disease onset, progression, and severity via the gut–brain axis, serving as microbial signatures of Parkinson’s Disease. Taxa coloured green have been observed to increase in abundance in PD patients, taxa coloured red decrease in abundance in PD patients, and conflicting evidence relating to their relative abundance in PD patients exists for taxa coloured blue. The number of studies providing evidence for each microbial shift are provided in the labels. Created with BioRender.

Table 1.
Variations in human gut microbiome composition, grouped by taxa, between healthy controls and Parkinson’s Disease patients, based on faecal matter and mucosal sample analysis [4,13,25,26,27,28,29,30,31,32].
As highlighted in Table 1, it is not possible to pinpoint any one responsible bacterium or microbial taxa in the pathogenesis of PD, since it is not necessarily the presence or the absence of certain microbes, but rather overall compositional shifts, known as dysbiosis, that contribute to PD. Dysbiosis refers to alterations in the number and composition of gut microbes, the gut microbiome environment, and microbial metabolites [21]. These alterations in microbial communities are frequently measured and quantified via alpha (i.e., microbial richness) and beta diversity (i.e., variations between microbiota within two or more hosts). In their systematic review of gut microbiome studies between PD patients and healthy controls, Boertien et al. [35] found that alpha diversity indices were reported in eleven of thirteen assessed studies; of these, three found higher alpha diversity in PD, six reported no difference, one reported lower alpha diversity, and one reported a difference without suggesting whether it was increased or decreased, further highlighting the inter-study variability in microbial signatures of PD. However, the authors noted that in all studies that performed 16S rRNA gene sequencing or metagenomic sequencing, beta diversity showed differences between PD patients and healthy controls, indicating changes are present [35].
4. Key Microbial Alterations in PD at the Genus Taxonomical Level
4.1. Akkermansia
Akkermansia are some of the most important bacteria under the Verrucomicrobiaceae family, which degrade mucous for energy. While these mucin-degrading bacteria are vital for intestinal barrier homeostasis, their over-abundance leads to degradation of the intestinal mucous barrier and contributes to increased intestinal permeability [8], which may expose the intestinal neural plexus directly to oxidative stress or toxins, such as lipopolysaccharide (LPS) and pesticides [20,36]. This can lead to aggregation of α-synuclein fibrils and the subsequent generation of Lewy bodies and neuroinflammation in the ENS [14,37]. Numerous studies have shown that increased abundance of Akkermansia increases the risk of PD, as well as accelerating disease progression [37,38].
4.2. Bifidobacterium
While the literature shows conflicting information about Bifidobacteria, when considering statistically significant results from Table 1, PD patients appeared to have a relative increase in abundance of this genus, compared with healthy controls. Interestingly, Bifidobacteria are typically considered commensal bacteria, and may be protective against PD symptoms. For example, Dogra et al. [9] suggested that low counts of Bifidobacteria are associated with worsening of hallucinations. The main role of Bifidobacteria, and other bacteria in the Bifidobacteriaceae family, is to prevent the overgrowth of harmful bacteria and regulate the GI immune system. Bifidobacterium is also an important producer of the SCFA acetate [37]. Thus, it is suggested that relative over-abundances may indicate potential over-compensation in an attempt to reconstruct gut homeostasis [24].
4.3. Blautia
Blautia, and other relevant genera implicated in PD pathology, such as Coprococcus, Dorea, Lachnospira, Roseburia, and Ruminococcus, fall under the Lachnospiraceae family. This family is important for its role in hydrolysing diet-derived polysaccharides to produce butyrate and other SCFAs [39]. As shown in Table 1, bacteria from the Lachnospiraceae family are often depleted in PD patients, and numerous studies have found Blautia to be consistently decreased in PD. SCFAs, such as butyrate, modulate the activity of the ENS and thereby increase GI motility. Therefore, the reduction in Blautia, and other SCFA-producing bacteria, likely leads to the gastrointestinal dysmotility exhibited in PD [40]. Additionally, butyrate can suppress colon inflammation [39], which is another symptom implicated in PD pathology.
4.4. Coprococcus
Bacteria in the Coprococcus genus are key SCFA-producers under the Bacillota (formerly Firmicutes) phylum. These Gram-positive, obligate anaerobic cocci ferment carbohydrates to produce butyric and acetic acids. The relative abundance of this genus is associated with a reduced risk of many neuropsychological and neurodegenerative disorders. Conversely, the species Coprococcus eutactus, of the Coprococcus genus, is depleted in adults with PD [41]. Coprococcus and other genera from the Lachnospiraceae family, such as Blautia and Roseburia, are associated with anti-inflammatory properties [21]. It is noted that one study reviewed in Table 1 showed conflicting evidence: Li et al. [27] found that Coprococcus was increased rather than decreased, which necessitates further studies to elucidate the role of Coprococcus in PD.
4.5. Faecalibacterium
Faecalibacterium facilitate the degradation of cellulose and starch, fermenting these indigestible fibres into SCFAs, including butyrate [13,21]. The resultant butyrate and other anti-inflammatory metabolites support GI health, so the reduction in Faecalibacterium may impair gut barrier function. This can increase the risk of pathogenic invasion and α-synuclein formation in the ENS [24]. Additionally, Faecalibacterium, similar to other SCFA-producing bacteria, have been associated with anti-inflammatory properties [42]. SCFAs have also been shown to protect against dopamine and tyrosine hydroxylase depletion in the SNc [42]. Decreases in Faecalibacterium abundance are noted in PD faecal samples, and this change is likely accelerated with disease progression [43]. Furthermore, Faecalibacterium is also decreased in both Chron’s disease and ulcerative colitis; thus, the reduction in PD patients may cause similar symptoms to inflammatory bowel diseases, which could then lead to PD pathology [38].
4.6. Roseburia
Roseburia is another important butyrate producer [13] that plays an important role in strengthening the intestinal barrier, as butyrate supports the maintenance of tight junctions and mucin production by enterocytes. This is important for preventing microorganisms from crossing into the lamina propria [13,44]. Reductions in SCFA-producing bacteria may lead to increased intestinal permeability, also known as “leaky gut” [9]. Additionally, butyrate exhibits anti-inflammatory effects via the induction of regulatory T cells and downregulation of pro-inflammatory cytokines and Toll-like receptor (TLR) 4 receptors [39]. Some species in the Roseburia genus also produce acetate. This SCFA is beneficial in inhibiting the growth of entero-pathogens, reducing luminal pH, and increasing the absorption of dietary nutrients [39].
As well as the consistent depletion of Roseburia evidenced in PD patient faecal samples, studies have shown significant correlations between lower abundance of Roseburia and worse clinical progression of motor and non-motor symptoms [37]. According to Vacca et al. [39], Blautia and Roseburia are the genera most involved in the control of gut inflammatory processes, which is mediated through their metabolism of butyrate.
5. Factors Contributing to Inconsistent and Variable Microbial Signatures of PD
Despite the consistent changes in key taxa highlighted above, the findings summarised in Table 1 also demonstrated distinct inconsistencies for changes in the relative abundance of other key taxa in PD patients. For example, Petrov et al. [33] reported a decrease in Bacteroides in PD patients, while Keshavarzian et al. [25] reported an increase, with both findings being statistically significant (in faecal samples). The inconsistencies may be due to a multitude of factors, including the following:
- Stage of disease: Multiple studies have demonstrated that the gut microbiome composition varies considerably for various stages of PD patients [1,9,22]. For example, one study [25] took samples from both medication-naive PD subjects as well as treated PD patients of various disease durations. They noticed significant differences in microbiota composition between patients of different disease stages and highlighted that the significance was maintained when the medication-naive samples were omitted from the analysis. Thus, they concluded that while medications may have an effect, disease duration significantly impacts microbial communities in PD patients.
- Patient variations, including ethnicity, age, and dietary habits: Some patient variables are well-known determinants of microbiome composition, for example, age and sex [35]. There are also differences found between ethnicities, for example, bacteria from the Lactobacillaceae family are typically enriched in Western PD cohorts, but the same has not been found in Chinese PD studies [45]. Aside from racial differences, this variability between populations could be due to geographic location, and related dietary and/or environmental factors [27]. For instance, one study, which took samples from patients in three locations across the United States of America, identified geographical location as a confounding factor [26]. Dietary habits are also an important confounder, as the types of food consumed are directly related to the survival of various microbial populations. Of particular relevance is fruit and vegetable consumption, as these foods supply the dietary fibres that many microbes use as energy.
- Sampling method: Sample collection, transport, and storage, laboratory procedures, and sequencing methods can all impact upon the microbial composition of samples. For example, the “gold standard” for transport and storage of stool samples is to freeze samples at −80 °C immediately upon collection, which will preserve the samples for up to two years. Alternatively, samples can be stored at −20 °C for a few months [35]. Of the 11 studies reviewed in Table 1, only 5 mentioned freezing samples. Two of these [29,32] immediately froze stool samples to −80 °C and −35 °C, respectively. A further two stated that samples were collected at home before being frozen (at −80 °C and −20 °C, respectively) but provided no information on the time of collection and freezing [13,31]. Finally, Li et al. [27] noted that subjects were advised to store their own samples at −20 °C in a freezer until collection (within three days) when the samples were frozen at −80 °C. However, it is unlikely that subjects were able to adhere to this protocol in their home freezer. Thus, storage and transportation conditions can be a potential source for both intra- and inter-study differences [35]. Likewise, there are composition differences between faecal and mucosal samplings. For example, there are environmental differences between the colonic mucosa and the lumen, which lead to distinct differences in biodiversity and bacterial taxa [37]. Most studies characterise the microbiome through faecal extraction and microbial sequencing. However, one utilised both faecal and mucosal sampling in their study. They found that as well as the expected differences between microbiota collected from PD patients compared with healthy subjects, significant differences in α-diversity were observed between mucosal and faecal microbiota within both groups [25]. Finally, differences in sample DNA extraction methods and subsequent sequencing methods are also contributing factors to the differences found between these studies.
- Other contributing factors, including medication usage: While it can be difficult to extricate the effect of medications from the impact of disease duration on the gut microbiome composition of PD patients, it is apparent that medications and pharmaceutical formulations can affect microbiota composition [46,47]. For example, catechol-o-methyltransferase (COMT) inhibitors and anticholinergics led to significant differences in gut microbiome composition in PD patients [26]. Furthermore, both classes of drug led to GI side effects, which can further contribute to gut dysbiosis. While Hill-Burns et al. [26] only found a borderline significant effect of levodopa on gut microbiome composition, some strains of Lactobacillus produce enzymes that degrade levodopa into dopamine. This implies that levodopa use in PD patients may increase the abundance of certain Lactobacillus strains [45].
Ultimately, the inconsistencies and variations in microbial profiles of PD patients provide further evidence for the complexity of microbiome analyses and pinpointing a microbial signature for disease states, such as PD, considering the multifaceted factors that contribute to microbiome ecology, which introduces challenges in the design and implementation of gut microbiota-targeted therapies for widespread use across all PD patients.
6. Mechanisms Linking Gut Dysbiosis with PD Pathology
The altered abundance of various microbes is linked to PD pathology through a multitude of mechanisms, since all microbial genera/families exert different roles within the GIT. For example, an increase in pathogenic bacteria can lead to the production of toxins, tissue damage, and inflammation, while the relative increase in typically commensal bacteria can crowd other bacteria, dysregulate homeostatic functions, and/or alter the metabolome. In this way, dysbiosis refers to both the changes in relative abundance of microbes, at different taxonomical levels, and in the microbiome environment (including mucin layers, epithelial barriers, TLR, etc.) and metabolome. These factors all interact to induce and accelerate disease pathology via various factors, schematically summarised in Figure 2. Specifically, the mechanisms that link gut dysbiosis to PD pathology are detailed in Table 2, which provides key examples of compositional changes in the gut microbiome that are known to induce such mechanisms in PD patients. The focus of this review was too ambitious to provide a complete understanding of all the mechanistic actions involved in the pathogenesis of PD, which have been covered extensively in prior literature [8,9,16,20,22,24] (Dogra et al., 2022; Huang et al., 2021; Liang et al., 2021; Lorente-Picón and Laguna, 2021; Rani and Mondal, 2021; Shen et al., 2021; Wang et al., 2021). Thus, these key mechanisms and outcomes of dysbiosis have been highlighted, as they can potentially be corrected by microbiome-targeting therapies.
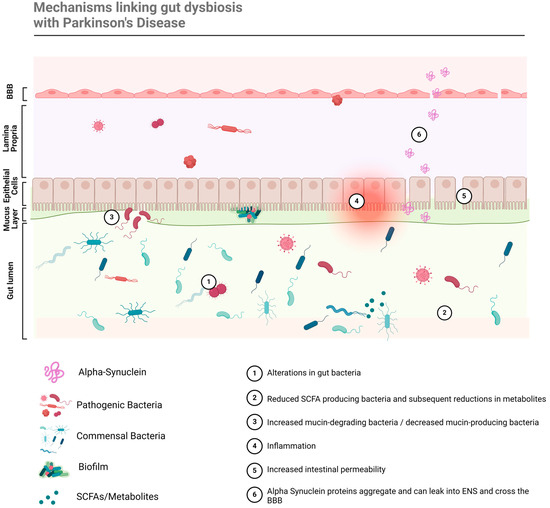
Figure 2.
Gut dysbiosis is linked with PD pathogenesis through a multitude of mechanisms that lead to neuroinflammation and neurodegeneration.
Table 2.
A summary of the various outcomes of gut dysbiosis, caused by specific composition changes in the gut microbiome, that are linked with Parkinson’s Disease pathogenesis.
7. Emerging Gut Microbiome-Targeted Therapies for Treating Parkinson’s Disease
As evidence continues to emerge linking gut dysbiosis with idiopathic PD, burgeoning research also highlights potential therapies that can effectively and safely mitigate symptoms and slow PD progression by targeting the gut microbiome. These include antibiotics, probiotics, prebiotics, postbiotics, and faecal microbiota transplantation (or faecal matter transplantation, FMT; Figure 3). A summary of the research and most recent evidence in the field of novel PD treatments follows.
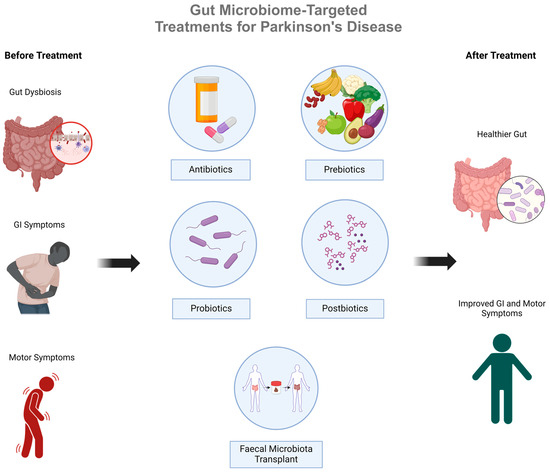
Figure 3.
An overview of gut microbiome-targeted therapies for Parkinson’s Disease that aim to overcome gut dysbiosis by restoring the gut microbiome in PD patients.
7.1. Antibiotics
Antibiotics, as well as clearing pathogenic microbes, can have other biological actions in the CNS, such as anti-inflammatory, immunomodulatory, and antioxidant effects, while also demonstrating potential to prevent abnormal protein aggregation [20]. Additionally, the repurposing of existing drugs (e.g., conventional antibiotics) can be beneficial, as safety profiling has already been completed and they are approved for use in humans, which accelerates the process of approval for clinical trials and their use for treating a new indication (e.g., PD). Numerous studies, including clinical trials and animal models, have examined the effects of various antibiotics on neurological disorders. Current evidence shows that antibiotics may exert neuroprotection by modulating gut microbiota and ameliorating PD-like pathophysiology [16]. Table 3 highlights some of the key antibiotic drugs that have demonstrated positive indications in PD models.
Table 3.
A summary of antibiotic agents that have been employed to exert neuroprotective effects or alleviate Parkinson’s Disease symptoms through defined mechanisms of action. Adapted from [16,20,21].
In addition to those listed in Table 3, combinations of antibiotics may also be useful to correct microbial imbalances. Additionally, there are established antibiotic treatments for H. pylori infection and SIBO, which have been linked with PD [37].
While some antibiotics show promise in helping to rectify microbial imbalances, the use of antibiotics as a therapy should be approached with an abundance of caution. Antibiotics can lead to dysbiosis through the disappearance of commensal bacteria, and the proliferation of new bacterial species. Thus, it is important to understand the mechanisms of various antibiotics, and their interactions between not only pathogenic bacteria, but also the wider microbiome population, ensuring that the beneficial bacteria are preserved and protected [20]. For example, minocycline showed promise as a potential drug candidate to treat PD and transitioned into a Phase II clinical trial in 2006 and 2008. However, the results of these trials did not show beneficial results and highlighted that long-term use of minocycline may be of concern [20]. Thus, more investigations are required to demonstrate their clinical potential in treating or alleviating PD symptomology.
7.2. Probiotics
The International Scientific Association for Probiotics and Prebiotics defines probiotics as “live microorganisms that when administered in adequate amounts confer a health benefit on the host” [51]. Recent studies using various probiotics have shown promising results in patients with PD. For example, administration of fermented milk containing Lactobacillus casei Shirota has shown therapeutic potential towards the GI symptoms of PD [52]. Indeed, probiotic use, in conjunction with prebiotics, has been shown to alleviate constipation [53]. Likewise, beneficial effects have been shown using probiotic strains of Lactobacillus and Bifidobacterium [16]. Probiotics may also be used during the management of Heliobacter Pylori infections to help restore the balance of commensal bacteria after antibiotic treatment. In PD patients, this helps to improve the absorption of levodopa [37].
In a 12-week randomised, double-blind, placebo-controlled clinical trial, PD patients who received a probiotic (containing Lactobacillus acidophilus, Bifidobacterium bifidum, Lactobacillus reuteri, and Lactobacillus fermentum) had better scores on the Movement Disorders Society–Unified Parkinson’s Disease Rating Scale (MDS-UPDRS), compared to the control group [54]. Reductions in oxidative stress biomarkers (such as C-reactive protein and malondialdehyde) and increased glutathione were also recorded. In other studies, L. salivarius LS01 and L. acidophilus LA02 significantly decreased oxidative stress and pro-inflammatory cytokines and promoted the production of anti-inflammatory cytokines in PD patients [16]. This may be because probiotics can upregulate the production of anti-inflammatory vitamins (for example, vitamins E, D3, B6, riboflavin, biotin, cobalamin, folates, pantothenic acid, nicotinic acid, and pyridoxin) and other bioactive molecules [37]. Finally, there is an indication that most probiotic strains also influence GI membrane integrity and inhibit potentially pathogenic bacterial overgrowth [16].
Animal models of PD have also shown that probiotic treatment improves PD outcomes. For example, L. plantarum DP189 has been shown to reduce α-synuclein aggravation by reducing oxidative damage, inflammation, and gut microbiota dysfunction [14]. Likewise, oral probiotic administration over a period of 16 weeks had neuroprotective effects in a transgenic PD mouse model, via the preservation of dopaminergic neurons [50]. The progressive deterioration of motor functions was also arrested: quantitative assessments showed that the treatment mitigated balance, coordination, and gait impairments. Probiotics also regulate the microbiome and impact upon serotonin metabolism, and other microbiome-targeting interventions can modulate the function of neurotransmitters, including dopamine and serotonin. However, further studies are needed to understand the impact of probiotics on neurotransmitters in PD patients.
While studies have produced exciting results, the use of probiotics for the treatment of illnesses is contentious. This is due in part to issues surrounding the processing and storage of probiotic supplements that use live bacteria. For example, changes in nomenclature and the lack of regulation in this space make it difficult to determine what probiotic products contain [55]. Additionally, there can be issues with whether storage materials (including capsules and jars) and storage environments (for example, temperature) are conducive to maintaining live bacteria [56]. Further to storage challenges, evidence suggests that traditional probiotics rarely survive the passage through the acidic environment of the stomach and/or effectively colonise the GIT without being excreted. In fact, recent evidence suggests that rather than live bacteria, bacterial spores are not only more likely to survive the stomach, but they are also more stable during processing and storage [41]. Alternatively, an emerging technique that can help to maintain the survivability of probiotics through processing and storage, as well as consumption and transit through the GIT, is through microencapsulation, which refers to incorporating probiotics into biopolymer- or emulsion-based delivery systems. The reader is referred to the review by Yoha et al. [57], which discusses the importance and various methods for encapsulation of probiotics. Encapsulation can also improve the stability and adhesion of probiotics, via the use of different pH, enzymatic, or immune responsive systems, which allow for targeted delivery of probiotics to specific areas within the GIT [58].
Finally, if the bacteria do make it alive to the intestines, sufficient fuel sources (e.g., prebiotic fibres) are needed to support their subsequent colonisation of the GIT. In this regard, the use of a combination of probiotics with prebiotic fibres, known as synbiotics, has shown to support the survival of probiotics [16]. Another important consideration about the use of probiotics as a treatment is that some strains of bacteria that are typically seen as beneficial or commensal can lead to SIBO or disease if they become over-abundant. For example, Akkermansia, increasingly implicated as a catalyst of PD, is often utilised in probiotics to improve host health. Thus, supplementation without knowledge of patients’ gut microbiome composition could potentially cause undue harm.
7.3. Prebiotics
Prebiotics are beneficial to human health, as they stimulate the growth and activity of health-promoting bacteria, in particular favouring the growth of SCFA-producing bacteria [37,39]. Prebiotics are indigestible dietary components, including fructo-oligosaccharides (FOS), inulin, starches, lactulose, galacto-oligosaccharides (GOS), etc., which are fermented by gut bacteria in the colon for energy [20,39,41]. This fermentation results in the production of beneficial SCFAs as by-products. As SCFAs and other by-products of fermentation are typically acidic, prebiotics can modify the gut via changes in pH. For example, pH alteration can change the population of acid-sensitive species, such as Bacteroides, and promote butyrate formation by Bacillota (formerly Firmicutes), in a process known as the butyrogenic effect [56]. Prebiotic-related increases in SCFA reduce intestinal barrier permeability, inhibit endotoxin crossing the mucosal barrier, and reduce inflammation. Prebiotic fibres are also beneficial in promoting GI motility, stool quality, immune function, and alleviating constipation [20,22]. Animal studies have also indicated that supplementation with prebiotics can increase the levels of brain-derived neurotrophic factor (BDNF) in the dentate gyrus of the hippocampus. These effects result in decreased neuroinflammation and reduced dopaminergic neuron loss [37].
While supplementation with prebiotic fibres has positive results, simple dietary changes are also an effective way to increase prebiotic consumption. For example, the Western diet, which consists of limited dietary fibre and increased amounts of animal protein, sugars, and saturated fatty acids, is increasingly implicated in chronic disease, including PD. A recent systematic review found that Western dietary patterns not only increased the incidence of PD, but also exacerbated disease severity in patients [14]. This is because the Western diet leads to a reduction in carbohydrate-fermenting bacteria, and thus, reduced butyrate, propionate, and acetate [20,22]. Conversely, numerous studies have concluded that the Mediterranean dietary pattern, with its high consumption of vegetables, fruits, nuts, and fish, and thus fibre, polyphenols, and omega-3 fatty acids, lowers the risk of PD, and may exert neuroprotective effects in PD patients [14,41]. This may be, in part, due to the higher total SCFAs found in those with greater plant-food consumption. Increased consumption of omega-3 fatty acids, vitamins, and polyphenols in this dietary pattern is also a contributing factor. Clinical studies have shown that supplementation with omega-3 in conjunction with vitamin E has beneficial effects in PD patients [59,60]. Supplementation with omega-3 also supports the proliferation of SCFA-producing bacteria from the Lachnospiraceae family (such as Blautia and Coprococcus genera, and Roseburia spp. species) [39]. By limiting ultra-processed foods, and increasing consumption of high-fibre foods, which include fruits and vegetables, nuts, legumes and beans, and wholegrain carbohydrates, prebiotic intake is naturally increased. The increase in dietary prebiotics and other beneficial nutrients will support the proliferation of commensal, SCFA-producing bacteria.
Although prebiotic fibres are typically found in a balanced diet, chitosan oligosaccharide (COS) is emerging as another promising prebiotic. Unlike other prebiotics, COS is not a dietary fibre—it is obtained from the shells of certain crustaceans. COS has been shown to reduce oxidative stress and oxidative damage, while exerting an anti-inflammatory response by reducing the release of pro-inflammatory cytokines [53]. Furthermore, COS is beneficial when utilised for probiotic encapsulation, and it has strong adhesion properties, which support the colonisation of probiotics in the colon [36,57]. Despite the promising results emerging in recent research for the use of prebiotics across a range of disease states linked with gut dysbiosis, limited evidence exists for their use as a PD pharmacotherapy. A recent study assessed the impact of different types of prebiotic fibres on SCFA production in PD patients compared with healthy controls. The authors concluded that all fibre types stimulated SCFA production. However, they observed that butyrate production remained lower in PD patients than healthy humans, regardless of prebiotic supplementation [61]. Thus, while prebiotics may exert beneficial results in PD patients, they are best treated as an adjunct therapy to support microbiome health by providing energy to commensal bacteria and improving GI function.
7.4. Postbiotics
SCFAs and other metabolites, including bioactive peptides, tryptophan degradation metabolites, bile acids, and vitamins (such as vitamins B1, B3, B9, B12, K, and A), that are produced by probiotic microbes are considered postbiotics and may be supplemented without the need for administering the live bacteria [62]. The importance of SCFAs and other by-products of microbial fermentation cannot be overstated. Covered in detail previously, SCFAs have several functions that impact host wellbeing and homeostasis. Some of these important impacts include maintaining the integrity of the intestinal barrier, promoting normal microglial development, anti-inflammatory effects, and mitigating neuronal damage [37]. In recent years, postbiotic supplementation has been explored due to increasing evidence showing the immunomodulatory, anti-inflammatory, anti-proliferative, and antioxidant effects of SCFAs. Studies have shown that butyrate administration alleviates motor symptoms, improves dopamine deficiency, and reduces neuroinflammation in PD animal models [14]. Animal models have also shown that oral supplementation of butyrate increases plasma concentrations, indicating that oral administration may have direct actions in the brain [20].
A recent study investigated the effects of sodium butyrate treatment on PD-induced mice [63]. This study utilised ceftriaxone as a pre-treatment to induce dysbiosis and gut alterations, followed by a 6-hydroxydopamine interstitial injection to catalyse dopaminergic damage in order to mimic Braak’s ‘dual-hit theory’ of PD in the mice. The mice were subsequently treated with sodium butyrate. This study showed that butyrate treatment repaired gut damage by significantly increasing goblet cell numbers and Muc2 expression in the intestines of the mice, reducing intestinal inflammation and gut permeability. Treatment also led to improvements in motor coordination and neuro-behavioural deficits, which the authors associated with reduced pathogenic factors and inflammation in the striatum, as well as reduced systemic inflammation. The authors concluded that butyrate treatment led to improvements in microbiome composition, helping to establish a new microbial balance with positive effects on the PD phenotype.
Tryptophan is also metabolised by gut microbiota, as well as via the kynurenine pathway or the serotonin pathway [64]. The subsequent metabolites produced can be either neuroprotective (such as kynurenic acid (KYNA), picolinic acid, and nicotinamide adenine dinucleotide), or neurotoxic (such as quinolinic acid (QA) and 3-hydroxykynurenine (3-HK)). In PD patients’ brains, the balance is shifted in favour of neurotoxic tryptophan metabolites: KYNA is decreased in the putamen, SNc, and the frontal cortex, while QA in plasma and 3-HK in the SNc and putamen are increased. The accumulation of these metabolites increases neurotoxicity and oxidative stress. Melatonin, which is another metabolite of the serotonin pathway, decreases oxidative stress and the expression of mitochondrial-dependent apoptotic pathways. This prevents the destruction of dopaminergic neurons and relieves the non-motor symptoms of PD [64]. A recent meta-analysis of randomised controlled trials utilising melatonin supplementation in PD patients suggested that melatonin supplementation reduces motor symptoms and sleep disturbances in patients [65]. However, the five included studies each had small sample sizes, with the meta-analysis only including a total of 155 patients. Furthermore, the authors suggested that more promising results might be observed if melatonin was administered in early-stage PD patients.
Despite indications that postbiotic supplementation could offer hope to PD patients, there have been few studies that utilise postbiotic supplementation in humans. Furthermore, while supplementation with postbiotics is an exciting area of research in the treatment of neurological disorders, it is noted that altering the microbial composition of the gut microbiome would lead to increased microbially derived metabolites. Microbially produced metabolites act both locally and systemically via the gut–brain axis. Thus, treatments that work to shift a patient’s microbiome, from a PD microbial signature to that of a healthy human, would likely be a more effective long-term solution.
7.5. Faecal Microbiota Transplantation
FMT uses stool from a healthy donor to recolonise the gut microbiome in a patient. Faecal matter is transferred into the patient via colonoscopy, nasal-jejunal tube, or orally via capsules [36,66]. FMT is perhaps the most advanced microbiome-targeted therapy for a wide range of disorders linked with gut dysbiosis, in large part due to its holistic approach, whereby native microbiomes (including microbes, fibre, and metabolites) are transplanted into the patient’s GIT. As such, FMT is already an established treatment for Clostridium difficile infection and IBD and has shown promising results in patients with various neurological disorders [2,20,40,67,68]. FMT is also associated with beneficial changes in microbial composition, such as increased relative abundance of Blautia (Genus) and Lachnospiraceae (family), and significantly reduced abundance of Escherichia-Shigella (Genus) [37]. The use of FMT in PD patients is still under-researched; however, multiple clinical trials (and one case study) have revealed promising results, as outlined in Table 4.
Table 4.
A summary of faecal microbiota transplantation (FMT) clinical study designs and outcomes for the treatment of Parkinson’s Disease [2,36,40,42,67,69,70].
The most statistically powered FMT clinical trial in PD patients to date is the recent study by Cheng et al. [70], which included 54 patients (27 received treatment and 27 a placebo). In this study, improvements were seen in roughly half of the treatment group, termed “FMT responders” (FMT.R). The responders showed statistically significant improvements in MDS-UPDRS total scores and non-motor symptoms, including GI symptoms. Interestingly, the FMT treatment participants showed improvements in cognitive function, regardless of their responses in PD symptoms. The authors noted that a high placebo response was observed with regard to GI symptoms at weeks 4 and 8. They suggest that this may be due to dietary recommendations given to study participants. Regardless, by week 12, the FMT arm showed significant improvements, which were not matched in the placebo group.
To understand why some patients responded to FMT while others did not, the study utilised metagenomic sequencing to analyse microbial species in the FMT.R and FMT non-responder (FMT.NR) subgroups. At baseline, there were no significant differences in microbiota diversity between either subgroup and taxa were found with significantly different abundance. By the 12th week, there was still no significant microbiota diversity between the two subgroups, although 20 microbial species had notably altered levels in FMT.R individuals compared with FMT.NR. Furthermore, some of the altered species had strong correlations with improvements in patient clinical scores. The authors highlighted Eubacterium eligens, Eubacterium ventriosum, Clostridiales bacterium 42_27, uncultured Blautia sp., Clostridioides difficile, uncultured Clostridium sp., and Roseburia hominis as key species that were positively correlated with GI and PD symptoms. Additionally, they found that the two subgroups had different traits in gut microbial functional pathways, which also positively correlated with the improvement in both GI and PD symptoms. Unfortunately, the follow-up time of this trial was short, at just 12 weeks. Future studies with longer observation periods and more detailed supporting dietary interventions will complement these findings.
Beyond Cheng et al.’s study, Table 4 highlights the capacity for FMT to alleviate gastric symptoms, such as constipation, in PD patients, with some studies observing positive responses in motor symptoms. However, these studies are not without limitations. Firstly, the sample sizes are very small, with 4 of the 7 studies having sample sizes of between 6 and 15 subjects. Additionally, three of the seven clinical trials did not analyse patients’ microbiome composition before or after FMT through longitudinal studies. Furthermore, in most of the studies, it does not appear as though patients were encouraged to eat diets that would support the colonisation of the new microbiota, post-treatment. Where this recommendation was made, the implementation of dietary measures was lacking. Kuai et al. [40] suggested that “all the patients basically followed the traditional Chinese Food structure (containing mainly grains and vegetables, small amounts of meat) before and after the FMT treatment”. However, no further mention of dietary methods is made in the article. Likewise, Cheng et al. [70] suggested that participants were advised to follow a “specific regular and healthy diet, with limited protein intake and high fibre”; however, the authors also noted that no dietary information was recorded.
In animal models of PD, FMT has been evidenced to increase dopamine levels in the SN, reduce neuroinflammation, modulate immune responses, decrease α-synuclein expression, and reduce motor symptoms [20,67]. Animal studies have also shown that FMT is likely also neuroprotective in PD by inhibiting TLR4-NFκB pathway-mediated inflammation [21]. Furthermore, Sun et al. [71] showed that mice receiving gut microbiota from PD mice also displayed motor dysfunctions, as well as decreased striatal dopamine, serotonin, and their metabolites. Conversely, the PD mice receiving faecal microbiota from normal mice displayed significant recovery of dopamine, serotonin, and their metabolites in striatum, as well as recovery of motor function. They also highlighted that FMT can suppress the loss of dopaminergic neurons in the SN in animal models.
8. Current Knowledge Gaps and Future Directions
Recent research has built upon the early works of Lewy and Braak, to highlight the involvement of the gut–brain axis in the pathogenesis and progression of PD. Exciting developments have been made when it comes to determining the ‘PD microbial signature’, as well as leveraging the gut microbiome in treatments for PD. However, more research is vital. One of the biggest limitations to progress is the absence of defined research protocols. Likewise, differences in sampling methods make it difficult to compare results between studies. This affects both studies that determine the composition of the gut microbiome and those that leverage the microbiome for treatment. Another area for improvement is the recruitment of patients and controls for clinical trials. Where clinical trials have shown promise, often a small sample size limited the statistical power of the results. Thus, there is a need for larger studies with high participant numbers to accurately determine the PD microbial signature at various stages of PD, as well as to demonstrate efficacy for promising treatments, such as FMT. A recent conference abstract referred to a study that sought to characterise the PD gut microbiome in a large cohort, including 1009 PD patients and 546 neurologically healthy controls [72]. Unfortunately, at the time of writing, it does not appear as though these data have been published. The results from this analysis, as well as other ongoing studies, will add value to the current knowledge base, and serve as precedent for more clinical trials in the future. Other knowledge gaps and promising future directions are summarised below.
8.1. Microbiome Composition and Non-Motor Symptoms as Diagnostic Markers
As important as determining the PD microbial signature and researching novel treatments for the disease is identifying new diagnostic tools to detect PD and potentially mitigate some of the neurological damage. As it stands, there are currently no reliable biomarkers to predict the onset or early stages of PD. Often the pathway to diagnosis involves medicating patients with motor symptoms to see if they respond to dopamine replacement therapy. However, this review has highlighted numerous symptoms and risk factors of PD that occur decades before the onset of motor symptoms. These include common non-motor symptoms, such as constipation, rapid eye movement (REM) sleep behaviour disorder, excessive daytime sleepiness, and postprandial fullness [73]. Anosmia and taste loss, mood disturbances, excessive sweating, fatigue, and pain can also occur up to ten years prior to motor symptoms. Patients who experience these symptoms should be considered for biological testing, which may indicate potential PD biomarkers.
PD patients often exhibit reduced production of thiamine and folate, which presents as deficiency [16]. Riboflavin metabolism, which is reflective of riboflavin status, is also reduced, and vitamin D deficiency is also common [73,74]. Nutrient levels, and the presence of lipopolysaccharide-binding protein (as a marker for LPS presence), can be determined via serum testing. Likewise, total and active levels of plasma ghrelin are decreased in patients with PD, which can also be determined by a blood test [22]. The presence of H. pylori and SIBO have also been identified as risk factors for PD, both of which are measurable by non-invasive breath tests (carbon and lactulose hydrogen breath tests, respectively). Other microbiome-related PD indications include gut inflammation, reduced SCFAs, GI presence of α-synuclein proteins and LPS, and microbial signatures. For example, reduced abundance of Prevotellaceae in faeces of PD was considered a biomarker of PD [13]. Similarly, reductions in SCFA producers, such as Ruminococcus, Blautia, and Faecalibacterium, might be an indicator of PD. Furthermore, reduced faecal SCFAs and higher faecal calprotectin levels also correlate with PD [49]. Additionally, increased pathobiont species, such as Enterococcus and Escherichia-Shigella can also be indicative of PD [13]. Thus, faecal sampling should be utilised in patients at risk of PD to ascertain the presence of a ‘PD microbial signature’ and identify the presence of calprotectin and other inflammatory markers.
Taken individually, each of the above factors are likely not strong enough to predict PD. However, together, they could form a diagnostic tool that may be utilised in the identification of early-stage PD, or perhaps even to identify and mitigate risk prior to disease onset. For example, a simple symptom checklist could serve as an indication for combined biological testing, including blood and breath testing, and faecal sampling. The development of such a clinical tool will lead to improved and earlier diagnoses of PD, and thus enable quicker and more effective treatment. Likewise, improved diagnostic tools will mitigate the risk of unnecessarily medicating patients with similar presentations.
8.2. Technological Advancements Required for the Development of Next-Generation Diagnostic Tools and Pharmacotherapies
Considering the obvious role of the gut microbiome in the pathogenesis of PD, defining the PD microbial signature for use as a diagnostic tool is pertinent. Data from previous studies can be utilised to determine epidemiological associations between specific microbial taxa and PD. The influence of individual genetics on the abundance of specific taxa is also an important consideration. This should be established in genome-wide association studies (GWAS) [35]. Furthermore, analysis of microbial metabolites and their functions is essential to comprehend the relationship between the gut microbiome and human health, and in doing so it is important to differentiate microbial metabolites from the host or food-derived components [75]. Thus, the use of machine learning (ML) and other new technologies will be vital in the development of diagnostic tools. Emerging technologies in this space are already showing promise, as outlined below.
Machine Learning: Recently, computational approaches have led to greater understanding of the relationship between gut microbes, human genetics, and microbial metabolites, as integration of various ‘omics’ methods (for example, metataxonomic and metagenomic sequencing, and metabolomics) with machine learning is becoming increasingly common. For example, the gutMGene database has linked 332 gut microbes, 223 genes, and 207 microbial metabolites in humans. Furthermore, the relationship between 774 gut microbiota and 221 human diseases has been elucidated in the Amadis database, and the relationship between 579 gut microbiota and 77 intervention measures or 123 disorders in humans according to the gutMDisorder database [75].
In 2020, Pietrucci et al. [34] utilised a machine learning approach to determine microbial shifts in PD patients. They re-analysed data from studies comprising a total of 472 PD patients and 374 controls, randomly selecting 80% of the samples to create the training set, and the remaining 20% to create the test set and evaluate the prediction. They compared efficacy between three different ML algorithms and found that the RF algorithm was the most effective, with an accuracy of 71%. Using RF, they found a subset of 22 bacterial families that were predictive of PD and characterised them by importance. The authors noted that the importance of each family in PD pathology was not directly correlated with its relative abundance in the gut microbiome. For example, similar to the previous studies, the authors found that Verrucomicrobiaceae/Akkermansiaceae and Bifidobacteriaceae are key bacterial taxa associated strongly with PD pathogenesis; however, they noted that these species have a low relative abundance. Interestingly, not all the families identified as important by the algorithm have been previously identified in the literature. For example, two of the ten most important bacterial families, Veillonellaceae and Alcaligenaceae, have never been reported before. These families were both relatively more abundant in PD patient samples.
This work identified ML as a powerful tool to determine the PD microbial signature for diagnostic purposes; however, more work needs to be carried out. The data samples used were a small fraction of the available information, and a larger dataset may help to improve accuracy. Furthermore, inconsistencies in methodologies used in studies may also impact the results. Identification of a standard protocol for microbiome studies and data sharing between researchers will be key to effectively utilising ML in PD microbiome research. Advances in the genetic characterisation of microbes will enable researchers to identify the key species of bacteria (and their metabolites) involved in PD. This will support the development of precision interventions to target the specific bacteria involved in PD aetiology and progression, as opposed to current microbiome-targeting therapies, which are indiscriminate in their approach.
In Vitro models: Developing biologically relevant in vitro models that simulate the gut microbiome and gut–brain axis and benefit from strong in vitro–in vivo correlations is critical for the testing and optimisation of next-generation PD therapeutics. Human intestinal organoids (HIO) are cellular models of the human intestine established through differentiation of induced pluripotent stem cells. These models can then be ‘reprogrammed’ using patient-derived materials, to create minimally invasive, patient-specific assessments. Patient ENS tissue and gut microbiota sampling can be used to determine significant contributions of gut microbiota to the pathophysiology of PD. These models can also be used to determine the efficacy of novel therapeutic targets that target the microbiome.
Another novel in vitro model is the Mucosal-Simulator of the Human Intestinal Microbial Ecosystem (M-SHIME®), which is a simulator of the human intestinal microbial ecosystem, equipped with a mucosal compartment. Ghyselinck et al. [76] used M-SHIME® to determine the efficacy of probiotic supplementation in restoring bacterial composition in PD patients. While their study was small (6 subjects: 3 PD and 3 control), and although the data covered only a 48 h period, some significant changes in bacterial composition and microbial metabolites were observed. While more studies using these models are needed, HIOs, M-SHIME®, and other new in vitro models will be valuable tools in the development and testing of novel treatments for PD.
8.3. Dietary Interventions to Support Microbiome-Targeted Therapies
Diet is an important consideration when designing microbiome-targeting interventions, since it is well established that many commensal bacteria rely on indigestible dietary fibres for energy sources. While dietary interventions are not recommended as a treatment for PD, ensuring accurate measurement and recording of dietary consumption prior to, and following, microbiome-altering treatments (such as FMT and probiotic supplementation) would strengthen the methodological reporting of these interventions. Moreover, adequate dietary fibre consumption can prevent mucin degradation, as some bacteria will break down and utilise intestinal mucous for energy in the absence of prebiotic fibres. Thus, ensuring that any introduced bacteria have adequate ‘food’ sources will support their subsequent colonisation of the GIT, and prevent damage to the GI lining. Furthermore, healthy diets that incorporate adequate dietary fibre can also help to alleviate some of the GI symptoms involved in PD.
Aside from dietary fibres, other dietary components and nutrients have benefits in supporting the proliferation of commensal bacteria. For example, a pilot human clinical trial confirmed that the relative abundance of Coprococcus spp. increased upon administration of vitamins A, B2, C, and D3. Furthermore, the SCFA concentration also increased according to the type and dose of administered vitamins. Another example is coffee, which is widely regarded as a negative risk factor towards PD. Coffee may also be neuroprotective, likely due to a combination of the caffeine and polyphenols. Coffee consumption can impact the gut microbiome, leading to an increase in anti-inflammatory Bifidobacteria and decreased pathogenic bacteria, such as Clostridium spp. and Escherichia coli. In PD mouse models, coffee regulates the gut microbiota, reducing α-synuclein aggregation and mitigating dopaminergic neuronal loss, as well as improving motor symptoms [14].
Improved understanding of the role of non-bacterial microbes in the human gut microbiome: Despite the surge of interest in the human gut microbiome over recent decades, up to 65% of bacterial species residing in the human GIT remain uncultured, so understanding of their biological roles is lacking [64]. Moreover, most of the literature revolves around the bacterial microbiome, and there is a lack of understanding of the role of the fungal, viral, and eukaryotic microbiomes [17]. There is also a lack of tools for sequencing fungal and viral genomes, or analysing their functions, as well as limited reference databases in this space [77]. Determining the role of viruses in the GIT is vital to understanding the role of the gut microbiome in diseases, including PD, since viruses exert a direct impact on the human host, as well as the other residing microbes within the microbiota. While fungi make up a smaller portion of the gut microbiome compared to viruses, studying their impact on the microbiome is also important, as there appears to be a correlation between increased fungal communities and gut dysbiosis, as seen in IBD [17].
9. Conclusions
It is now clear that the gut microbiome has a role in both the aetiology and treatment of PD. Studies characterising the PD microbial signature have come a long way in recent years to identify the specific microbial imbalances that characterise gut dysbiosis in PD. As more trials are completed, researchers will better understand the complete PD microbial signature, as well as early indications of the disease. With these advancements, there is hope that soon we will be able to detect PD at a much earlier stage, or even identify high-risk individuals before the onset of disease. This will potentially mitigate the severe neurological damage seen in the later stages of PD. Importantly, results from both preclinical and clinical studies have shown that the gut microbiome can be leveraged to alleviate PD symptoms via advanced microbiome-targeting therapies. However, more extensive clinical testing and development is required to optimise microbiome-targeting therapies for the treatment of PD, with specific focus on adequately controlled and high-powered clinical studies that validate treatment efficacy. Additionally, further studies must be completed to determine at which stage of disease microbiome-targeted therapies may be deployed for beneficial results. With future technological advancements in diagnostic tools, in vitro testing, and patient-centric formulation strategies, it is expected that microbiome-targeted therapies may serve as pharmacotherapies for improving the quality of life of PD sufferers, either as standalone treatment or in conjunction with current care practices and future advancements
Author Contributions
R.K., conceptualisation, methodology, investigation, writing—original draft, visualisation; P.J., conceptualisation, writing—review and editing, visualisation, supervision, project administration, funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the University of South Australia Vacation Scholarship Program and The Hospital Research Foundation (THRF) EMCR Fellowship funding (2022-CF-EMCR-004-25314).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Chen, Q.-Q.; Haikal, C.; Li, W.; Li, J.-Y. Gut Inflammation in Association with Pathogenesis of Parkinson’s Disease. Front. Mol. Neurosci. 2019, 12, 218. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Xu, H.; Luo, Q.; He, J.; Li, M.; Chen, H.; Tang, W.; Nie, Y.; Zhou, Y. Fecal microbiota transplantation to treat Parkinson’s disease with constipation: A case report. Medicine 2019, 98, e16163. [Google Scholar] [CrossRef] [PubMed]
- Schmit, K.J.; Garcia, P.; Sciortino, A.; Aho, V.T.E.; Pardo Rodriguez, B.; Thomas, M.H.; Gérardy, J.-J.; Bastero Acha, I.; Halder, R.; Cialini, C.; et al. Fiber deprivation and microbiome-borne curli shift gut bacterial populations and accelerate disease in a mouse model of Parkinson’s disease. Cell Rep. 2023, 42, 113071. [Google Scholar] [CrossRef] [PubMed]
- Bedarf, J.R.; Hildebrand, F.; Coelho, L.P.; Sunagawa, S.; Bahram, M.; Goeser, F.; Bork, P.; Wüllner, U. Functional implications of microbial and viral gut metagenome changes in early stage L-DOPA-naïve Parkinson’s disease patients. Genome Med. 2017, 9, 39. [Google Scholar] [CrossRef] [PubMed]
- Kulcsarova, K.; Bang, C.; Berg, D.; Schaeffer, E. Pesticides and the Microbiome-Gut-Brain Axis: Convergent Pathways in the Pathogenesis of Parkinson’s Disease. J. Park. Dis. 2023, 13, 1079–1106. [Google Scholar] [CrossRef] [PubMed]
- Rietdijk, C.D.; Perez-Pardo, P.; Garssen, J.; van Wezel, R.J.; Kraneveld, A.D. Exploring Braak’s Hypothesis of Parkinson’s Disease. Front. Neurol. 2017, 8, 37. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zhu, G.; Wang, G.; Zhang, F. Oxidative Stress and Neuroinflammation Potentiate Each Other to Promote Progression of Dopamine Neurodegeneration. Oxidative Med. Cell. Longev. 2020, 2020, 6137521. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Liao, J.; Liu, X.; Zhong, Y.; Cai, X.; Long, L. Review: The Role of Intestinal Dysbiosis in Parkinson’s Disease. Front. Cell. Infect. Microbiol. 2021, 11, 615075. [Google Scholar] [CrossRef] [PubMed]
- Dogra, N.; Mani, R.J.; Katare, D.P. The Gut-Brain Axis: Two Ways Signaling in Parkinson’s Disease. Cell. Mol. Neurobiol. 2022, 42, 315–332. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.G.; Ventura, K.; Villeneuve, A.; Du Bois, P.; Holahan, M.R. Exploring the Connection Between the Gut Microbiome and Parkinson’s Disease Symptom Progression and Pathology: Implications for Supplementary Treatment Options. J. Park. Dis. 2022, 12, 2339–2352. [Google Scholar] [CrossRef]
- Forsyth, C.B.; Shannon, K.M.; Kordower, J.H.; Voigt, R.M.; Shaikh, M.; Jaglin, J.A.; Estes, J.D.; Dodiya, H.B.; Keshavarzian, A. Increased Intestinal Permeability Correlates with Sigmoid Mucosa alpha-Synuclein Staining and Endotoxin Exposure Markers in Early Parkinson’s Disease. PLoS ONE 2011, 6, e28032. [Google Scholar] [CrossRef] [PubMed]
- Elfil, M.; Kamel, S.; Kandil, M.; Koo, B.B.; Schaefer, S.M. Implications of the Gut Microbiome in Parkinson’s Disease. Mov. Disord. 2020, 35, 921–933. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wu, X.; Hu, X.; Wang, T.; Liang, S.; Duan, Y.; Jin, F.; Qin, B. Structural changes of gut microbiota in Parkinson’s disease and its correlation with clinical features. Sci. China Life Sci. 2017, 60, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Liu, X.; Ye, Y.; Yan, X.; Cheng, Y.; Zhao, L.; Chen, F.; Ling, Z. Gut Microbiota: A Novel Therapeutic Target for Parkinson’s Disease. Front. Immunol. 2022, 13, 937555. [Google Scholar] [CrossRef] [PubMed]
- Hassanzadeh, K.; Rahimmi, A. Oxidative stress and neuroinflammation in the story of Parkinson’s disease: Could targeting these pathways write a good ending? J. Cell. Physiol. 2019, 234, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Rani, L.; Mondal, A.C. Unravelling the role of gut microbiota in Parkinson’s disease progression: Pathogenic and therapeutic implications. Neurosci. Res. 2021, 168, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Balan, Y.; Gaur, A.; Sakthivadivel, V.; Kamble, B.; Sundaramurthy, R. Is the Gut Microbiota a Neglected Aspect of Gut and Brain Disorders? 2021, 13, e19740. [Google Scholar] [CrossRef]
- Dutta, S.K.; Verma, S.; Jain, V.; Surapaneni, B.K.; Vinayek, R.; Phillips, L.E.; Nair, P.P. Parkinson’s Disease: The Emerging Role of Gut Dysbiosis, Antibiotics, Probiotics, and Fecal Microbiota Transplantation. J. Neurogastroenterol. Motil. 2019, 25, 363–376. [Google Scholar] [CrossRef]
- Lorente-Picón, M.; Laguna, A. New Avenues for Parkinson’s Disease Therapeutics: Disease-Modifying Strategies Based on the Gut Microbiota. Biomolecules 2021, 11, 433. [Google Scholar] [CrossRef]
- Sun, M.; Shen, Y.-Q. Dysbiosis of gut microbiota and microbial metabolites in Parkinson’s Disease. Ageing Res. Rev. 2018, 45, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Luo, Y.; Ray Chaudhuri, K.; Reynolds, R.; Tan, E.-K.; Pettersson, S. The role of gut dysbiosis in Parkinson’s disease: Mechanistic insights and therapeutic options. Brain 2021, 144, 2571–2593. [Google Scholar] [CrossRef]
- Berthouzoz, E.; Lazarevic, V.; Zekeridou, A.; Castro, M.; Debove, I.; Aybek, S.; Schrenzel, J.; Burkhard, P.R.; Fleury, V. Oral and intestinal dysbiosis in Parkinson’s disease. Rev. Neurol. 2023, 179, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Yue, Y.; He, T.; Huang, C.; Qu, B.; Lv, W.; Lai, H.Y. The Association Between the Gut Microbiota and Parkinson’s Disease, a Meta-Analysis. Front. Aging Neurosci. 2021, 13, 636545. [Google Scholar] [CrossRef]
- Keshavarzian, A.; Green, S.J.; Engen, P.A.; Voigt, R.M.; Naqib, A.; Forsyth, C.B.; Mutlu, E.; Shannon, K.M. Colonic bacterial composition in Parkinson’s disease. Mov. Disord. 2015, 30, 1351–1360. [Google Scholar] [CrossRef] [PubMed]
- Hill-Burns, E.M.; Debelius, J.W.; Morton, J.T.; Wissemann, W.T.; Lewis, M.R.; Wallen, Z.D.; Peddada, S.D.; Factor, S.A.; Molho, E.; Zabetian, C.P.; et al. Parkinson’s disease and Parkinson’s disease medications have distinct signatures of the gut microbiome. Mov. Disord. 2017, 32, 739–749. [Google Scholar] [CrossRef]
- Li, C.; Cui, L.; Yang, Y.; Miao, J.; Zhao, X.; Zhang, J.; Cui, G.; Zhang, Y. Gut Microbiota Differs Between Parkinson’s Disease Patients and Healthy Controls in Northeast China. Front. Mol. Neurosci. 2019, 12, 171. [Google Scholar] [CrossRef]
- Pietrucci, D.; Cerroni, R.; Unida, V.; Farcomeni, A.; Pierantozzi, M.; Mercuri, N.B.; Biocca, S.; Stefani, A.; Desideri, A. Dysbiosis of gut microbiota in a selected population of Parkinson’s patients. Park. Relat. Disord. 2019, 65, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Unger, M.M.; Spiegel, J.; Dillmann, K.U.; Grundmann, D.; Philippeit, H.; Bürmann, J.; Faßbender, K.; Schwiertz, A.; Schäfer, K.H. Short chain fatty acids and gut microbiota differ between patients with Parkinson’s disease and age-matched controls. Park. Relat. Disord. 2016, 32, 66–72. [Google Scholar] [CrossRef]
- Vascellari, S.; Palmas, V.; Melis, M.; Pisanu, S.; Cusano, R.; Uva, P.; Perra, D.; Madau, V.; Sarchioto, M.; Oppo, V.; et al. Gut Microbiota and Metabolome Alterations Associated with Parkinson’s Disease. mSystems 2020, 5, 10–1128. [Google Scholar] [CrossRef]
- Wallen, Z.D.; Demirkan, A.; Twa, G.; Cohen, G.; Dean, M.N.; Standaert, D.G.; Sampson, T.R.; Payami, H. Metagenomics of Parkinson’s disease implicates the gut microbiome in multiple disease mechanisms. Nat. Commun. 2022, 13, 6958. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Yue, L.; Fang, X.; Wang, G.; Li, C.; Sun, X.; Jia, X.; Yang, J.; Song, J.; Zhang, Y.; et al. Altered gut microbiota in Parkinson’s disease patients/healthy spouses and its association with clinical features. Park. Relat. Disord. 2020, 81, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Petrov, V.A.; Saltykova, I.V.; Zhukova, I.A.; Alifirova, V.M.; Zhukova, N.G.; Dorofeeva, Y.B.; Tyakht, A.V.; Kovarsky, B.A.; Alekseev, D.G.; Kostryukova, E.S.; et al. Analysis of Gut Microbiota in Patients with Parkinson’s Disease. Bull. Exp. Biol. Med. 2017, 162, 734–737. [Google Scholar] [CrossRef] [PubMed]
- Pietrucci, D.; Teofani, A.; Unida, V.; Cerroni, R.; Biocca, S.; Stefani, A.; Desideri, A. Can Gut Microbiota Be a Good Predictor for Parkinson’s Disease? A Machine Learning Approach. Brain Sci. 2020, 10, 242. [Google Scholar] [CrossRef] [PubMed]
- Boertien, J.M.; Pereira, P.A.B.; Aho, V.T.E.; Scheperjans, F. Increasing Comparability and Utility of Gut Microbiome Studies in Parkinson’s Disease: A Systematic Review. J. Park. Dis. 2019, 9, S297–S312. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.J.; Yang, X.Z.; Tong, Q.; Shen, P.; Ma, S.J.; Wu, S.N.; Zheng, J.L.; Wang, H.A.-O. Fecal microbiota transplantation therapy for Parkinson’s disease: A preliminary study. Medicine 2020, 99, e22035. [Google Scholar] [CrossRef] [PubMed]
- Bonvegna, S.; Cilia, R. Chapter 8—Disease Mechanisms as Subtypes: Microbiome. In Handbook of Clinical Neurology; Espay, A.J., Ed.; Elsevier: Amsterdam, The Netherlands, 2023; Volume 193, pp. 107–131. [Google Scholar]
- Nishiwaki, H.; Ito, M.; Ishida, T.; Hamaguchi, T.; Maeda, T.; Kashihara, K.; Tsuboi, Y.; Ueyama, J.; Shimamura, T.; Mori, H.; et al. Meta-Analysis of Gut Dysbiosis in Parkinson’s Disease. Mov. Disord. 2020, 35, 1626–1635. [Google Scholar] [CrossRef]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef]
- Kuai, X.-y.; Yao, X.-h.; Xu, L.-j.; Zhou, Y.-q.; Zhang, L.-p.; Liu, Y.; Pei, S.-f.; Zhou, C.-l. Evaluation of fecal microbiota transplantation in Parkinson’s disease patients with constipation. Microb. Cell Factories 2021, 20, 98. [Google Scholar] [CrossRef]
- Notting, F.; Pirovano, W.; Sybesma, W.; Kort, R. The butyrate-producing and spore-forming bacterial genus Coprococcus as a potential biomarker for neurological disorders. Gut Microbiome 2023, 4, e16. [Google Scholar] [CrossRef]
- Segal, A.; Zlotnik, Y.; Moyal-Atias, K.; Abuhasira, R.; Ifergane, G. Fecal microbiota transplant as a potential treatment for Parkinson’s disease – A case series. Clin. Neurol. Neurosurg. 2021, 207, 106791. [Google Scholar] [CrossRef] [PubMed]
- Nishiwaki, H.; Ito, M.; Hamaguchi, T.; Maeda, T.; Kashihara, K.; Tsuboi, Y.; Ueyama, J.; Yoshida, T.; Hanada, H.; Takeuchi, I.; et al. Short chain fatty acids-producing and mucin-degrading intestinal bacteria predict the progression of early Parkinson’s disease. npj Park. Dis. 2022, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Gieryńska, M.; Szulc-Dąbrowska, L.; Struzik, J.; Mielcarska, M.B.; Gregorczyk-Zboroch, K.P. Integrity of the Intestinal Barrier: The Involvement of Epithelial Cells and Microbiota-A Mutual Relationship. Animals 2022, 12, 145. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Savva, G.M.; Bedarf, J.R.; Charles, I.G.; Hildebrand, F.; Narbad, A. Meta-analysis of the Parkinson’s disease gut microbiome suggests alterations linked to intestinal inflammation. npj Park. Dis. 2021, 7, 27. [Google Scholar] [CrossRef] [PubMed]
- Kamath, S.; Stringer, A.M.; Prestidge, C.A.; Joyce, P. Targeting the gut microbiome to control drug pharmacomicrobiomics: The next frontier in oral drug delivery. Expert. Opin. Drug Deliv. 2023, 20, 1315–1331. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, S.; Kamath, S.; Ariaee, A.; Prestidge, C.; Joyce, P. The impact of common pharmaceutical excipients on the gut microbiota. Expert. Opin. Drug Deliv. 2023, 20, 1297–1314. [Google Scholar] [CrossRef] [PubMed]
- Dardiotis, E.; Tsouris, Z.; Mentis, A.-F.A.; Siokas, V.; Michalopoulou, A.; Sokratous, M.; Dastamani, M.; Bogdanos, D.P.; Deretzi, G.; Kountouras, J.H. pylori and Parkinson’s disease: Meta-analyses including clinical severity. Clin. Neurol. Neurosurg. 2018, 175, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Aho, V.T.E.; Houser, M.C.; Pereira, P.A.B.; Chang, J.; Rudi, K.; Paulin, L.; Hertzberg, V.; Auvinen, P.; Tansey, M.G.; Scheperjans, F. Relationships of gut microbiota, short-chain fatty acids, inflammation, and the gut barrier in Parkinson’s disease. Mol. Neurodegener. 2021, 16, 6. [Google Scholar] [CrossRef]
- Hsieh, T.-H.; Kuo, C.-W.; Hsieh, K.-H.; Shieh, M.-J.; Peng, C.-W.; Chen, Y.-C.; Chang, Y.-L.; Huang, Y.-Z.; Chen, C.-C.; Chang, P.-K.; et al. Probiotics Alleviate the Progressive Deterioration of Motor Functions in a Mouse Model of Parkinson’s Disease. Brain Sci. 2020, 10, 206. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Yang, X.; He, X.; Xu, S.; Zhang, Y.; Mo, C.; Lai, Y.; Song, Y.; Yan, Z.; Ai, P.; Qian, Y.; et al. Effect of Lacticaseibacillus paracasei strain Shirota supplementation on clinical responses and gut microbiome in Parkinson’s disease. Food Funct. 2023, 14, 6828–6839. [Google Scholar] [CrossRef] [PubMed]
- Uceda, S.; Echeverry-Alzate, V.; Reiriz-Rojas, M.; Martínez-Miguel, E.; Pérez-Curiel, A.; Gómez-Senent, S.; Beltrán-Velasco, A.I. Gut Microbial Metabolome and Dysbiosis in Neurodegenerative Diseases: Psychobiotics and Fecal Microbiota Transplantation as a Therapeutic Approach—A Comprehensive Narrative Review. Int. J. Mol. Sci. 2023, 24, 13294. [Google Scholar] [CrossRef] [PubMed]
- Tamtaji, O.R.; Taghizadeh, M.; Daneshvar Kakhaki, R.; Kouchaki, E.; Bahmani, F.; Borzabadi, S.; Oryan, S.; Mafi, A.; Asemi, Z. Clinical and metabolic response to probiotic administration in people with Parkinson’s disease: A randomized, double-blind, placebo-controlled trial. Clin. Nutr. 2019, 38, 1031–1035. [Google Scholar] [CrossRef]
- Idrees, M.; Imran, M.; Atiq, N.; Zahra, R.; Abid, R.; Alreshidi, M.; Roberts, T.; Abdelgadir, A.; Tipu, M.K.; Farid, A.; et al. Probiotics, their action modality and the use of multi-omics in metamorphosis of commensal microbiota into target-based probiotics. Front. Nutr. 2022, 9, 959941. [Google Scholar] [CrossRef] [PubMed]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods 2019, 8, 92. [Google Scholar] [CrossRef] [PubMed]
- Yoha, K.S.; Nida, S.; Dutta, S.; Moses, J.A.; Anandharamakrishnan, C. Targeted Delivery of Probiotics: Perspectives on Research and Commercialization. Probiotics Antimicrob. Proteins 2022, 14, 15–48. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Gantumur, M.-A.; Sun, J.; Guo, J.; Ma, J.; Jiang, Z.; Wang, W.; Zhang, J.; Ma, Y.; Hou, J.; et al. Design of probiotic delivery systems for targeted release. Food Hydrocoll. 2024, 149, 109588. [Google Scholar] [CrossRef]
- Pantzaris, M.; Loukaides, G.; Paraskevis, D.; Kostaki, E.-G.; Patrikios, I. Neuroaspis PLP10™, a nutritional formula rich in omega-3 and omega-6 fatty acids with antioxidant vitamins including gamma-tocopherol in early Parkinson’s disease: A randomized, double-blind, placebo-controlled trial. Clin. Neurol. Neurosurg. 2021, 210, 106954. [Google Scholar] [CrossRef] [PubMed]
- Tamtaji, O.R.; Taghizadeh, M.; Aghadavod, E.; Mafi, A.; Dadgostar, E.; Daneshvar Kakhaki, R.; Abolhassani, J.; Asemi, Z. The effects of omega-3 fatty acids and vitamin E co-supplementation on gene expression related to inflammation, insulin and lipid in patients with Parkinson’s disease: A randomized, double-blind, placebo-controlled trial. Clin. Neurol. Neurosurg. 2019, 176, 116–121. [Google Scholar] [CrossRef]
- Baert, F.; Matthys, C.; Maselyne, J.; Van Poucke, C.; Van Coillie, E.; Bergmans, B.; Vlaemynck, G. Parkinson’s disease patients’ short chain fatty acids production capacity after in vitro fecal fiber fermentation. npj Park. Dis. 2021, 7, 72. [Google Scholar] [CrossRef]
- Peluzio, M.d.C.G.; Martinez, J.A.; Milagro, F.I. Postbiotics: Metabolites and mechanisms involved in microbiota-host interactions. Trends Food Sci. Technol. 2021, 108, 11–26. [Google Scholar] [CrossRef]
- Avagliano, C.; Coretti, L.; Lama, A.; Pirozzi, C.; De Caro, C.; De Biase, D.; Turco, L.; Mollica, M.P.; Paciello, O.; Calignano, A.; et al. Dual-Hit Model of Parkinson’s Disease: Impact of Dysbiosis on 6-Hydroxydopamine-Insulted Mice—Neuroprotective and Anti-Inflammatory Effects of Butyrate. Int. J. Mol. Sci. 2022, 23, 6367. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Cui, L.; Gao, J.; Zhu, M.; Zhang, Y.; Zhang, H.L. Gut Microbial Metabolites in Parkinson’s Disease: Implications of Mitochondrial Dysfunction in the Pathogenesis and Treatment. Mol. Neurobiol. 2021, 58, 3745–3758. [Google Scholar] [CrossRef] [PubMed]
- Iftikhar, S.; Sameer, H.M.; Zainab. Significant potential of melatonin therapy in Parkinson’s disease—A meta-analysis of randomized controlled trials. Front. Neurol. 2023, 14, 1265789. [Google Scholar] [CrossRef] [PubMed]
- Vongsavath, T.; Laeeq, T.; Tun, K.M.; Hong, A.S. The Potential Role of Fecal Microbiota Transplantation in Parkinson’s Disease: A Systematic Literature Review. Appl. Microbiol. 2023, 3, 993–1002. [Google Scholar] [CrossRef]
- DuPont, H.L.; Suescun, J.; Jiang, Z.D.; Brown, E.L.; Essigmann, H.T.; Alexander, A.S.; DuPont, A.W.; Iqbal, T.; Utay, N.S.; Newmark, M.; et al. Fecal microbiota transplantation in Parkinson’s disease-A randomized repeat-dose, placebo-controlled clinical pilot study. Front. Neurol. 2023, 14, 1104759. [Google Scholar] [CrossRef] [PubMed]
- Vendrik, K.E.W.; Ooijevaar, R.E.; de Jong, P.R.C.; Laman, J.D.; van Oosten, B.W.; van Hilten, J.J.; Ducarmon, Q.R.; Keller, J.J.; Kuijper, E.J.; Contarino, M.F. Fecal Microbiota Transplantation in Neurological Disorders. Front. Cell. Infect. Microbiol. 2020, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- Bruggeman, A.; Vandendriessche, C.; Hamerlinck, H.; De Looze, D.; Tate, D.J.; Vuylsteke, M.; De Commer, L.; Devolder, L.; Raes, J.; Verhasselt, B.; et al. Safety and efficacy of faecal microbiota transplantation in patients with mild to moderate Parkinson’s disease (GUT-PARFECT): A double-blind, placebo-controlled, randomised, phase 2 trial. eClinicalMedicine 2024, 71, 102563. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Tan, G.; Zhu, Q.; Wang, C.; Ruan, G.; Ying, S.; Qie, J.; Hu, X.; Xiao, Z.; Xu, F.; et al. Efficacy of fecal microbiota transplantation in patients with Parkinson’s disease: Clinical trial results from a randomized, placebo-controlled design. Gut Microbes 2023, 15, 2284247. [Google Scholar] [CrossRef]
- Sun, M.-F.; Zhu, Y.-L.; Zhou, Z.-L.; Jia, X.-B.; Xu, Y.-D.; Yang, Q.; Cui, C.; Shen, Y.-Q. Neuroprotective effects of fecal microbiota transplantation on MPTP-induced Parkinson’s disease mice: Gut microbiota, glial reaction and TLR4/TNF-α signaling pathway. Brain Behav. Immun. 2018, 70, 48–60. [Google Scholar] [CrossRef]
- Payami, H.; Standaert, D.; Wallen, Z.; Sampson, T. Widespread involvement of gut microbiome in Parkinson’s disease. J. Neurol. Sci. 2023, 455, 121077. [Google Scholar] [CrossRef]
- Nair, A.T.; Ramachandran, V.; Joghee, N.M.; Antony, S.; Ramalingam, G. Gut Microbiota Dysfunction as Reliable Non-invasive Early Diagnostic Biomarkers in the Pathophysiology of Parkinson’s Disease: A Critical Review. J. Neurogastroenterol. Motil. 2018, 24, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Rosario, D.; Bidkhori, G.; Lee, S.; Bedarf, J.; Hildebrand, F.; Le Chatelier, E.; Uhlen, M.; Ehrlich, S.D.; Proctor, G.; Wüllner, U.; et al. Systematic analysis of gut microbiome reveals the role of bacterial folate and homocysteine metabolism in Parkinson’s disease. Cell Rep. 2021, 34, 108807. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, A.; Ebrahimi, M.; Kharazmi, M.S.; Jafari, S.M. Effects of microbial-derived biotics (meta/pharma/post-biotics) on the modulation of gut microbiome and metabolome; general aspects and emerging trends. Food Chem. 2023, 411, 135478. [Google Scholar] [CrossRef] [PubMed]
- Ghyselinck, J.; Verstrepen, L.; Moens, F.; Van Den Abbeele, P.; Bruggeman, A.; Said, J.; Smith, B.; Barker, L.A.; Jordan, C.; Leta, V.; et al. Influence of probiotic bacteria on gut microbiota composition and gut wall function in an in-vitro model in patients with Parkinson’s disease. Int. J. Pharm. X 2021, 3, 100087. [Google Scholar] [CrossRef]
- Ratiner, K.; Ciocan, D.; Abdeen, S.K.; Elinav, E. Utilization of the microbiome in personalized medicine. Nat. Rev. Microbiol. 2024, 22, 291–308. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).