
Evidence of Placental Aging in Late SGA, Fetal Growth Restriction and Stillbirth—A Systematic Review

 , , , , , , ,
, , , , , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Inclusion and Exclusion Criteria
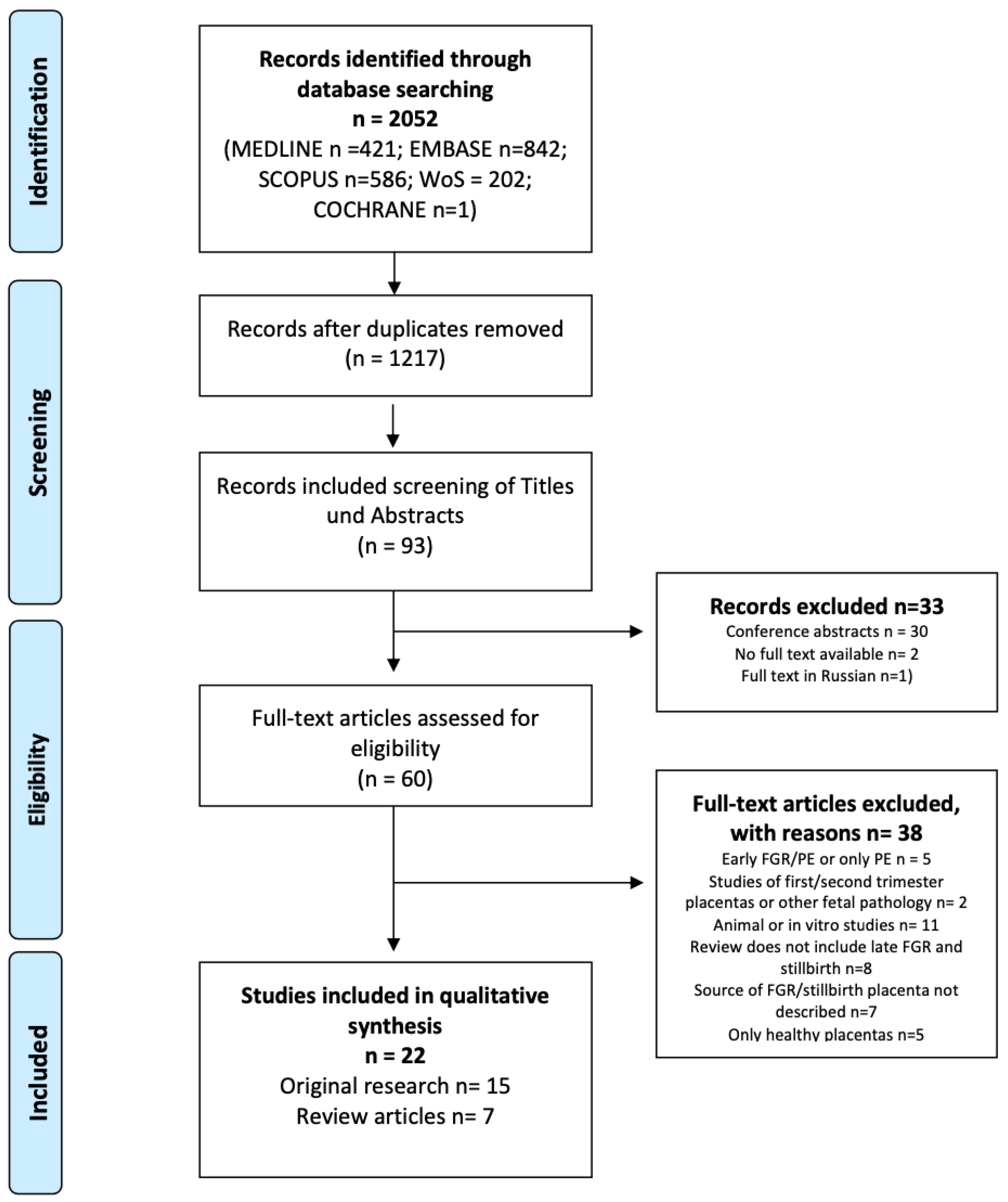
2.3. Study Selection
2.4. Process of Data Collection
2.5. Evaluation of Risk of Bias
2.6. Synthesis Methods
3. Results
3.1. Characteristics of the Studies
3.2. Synthesis of the Results
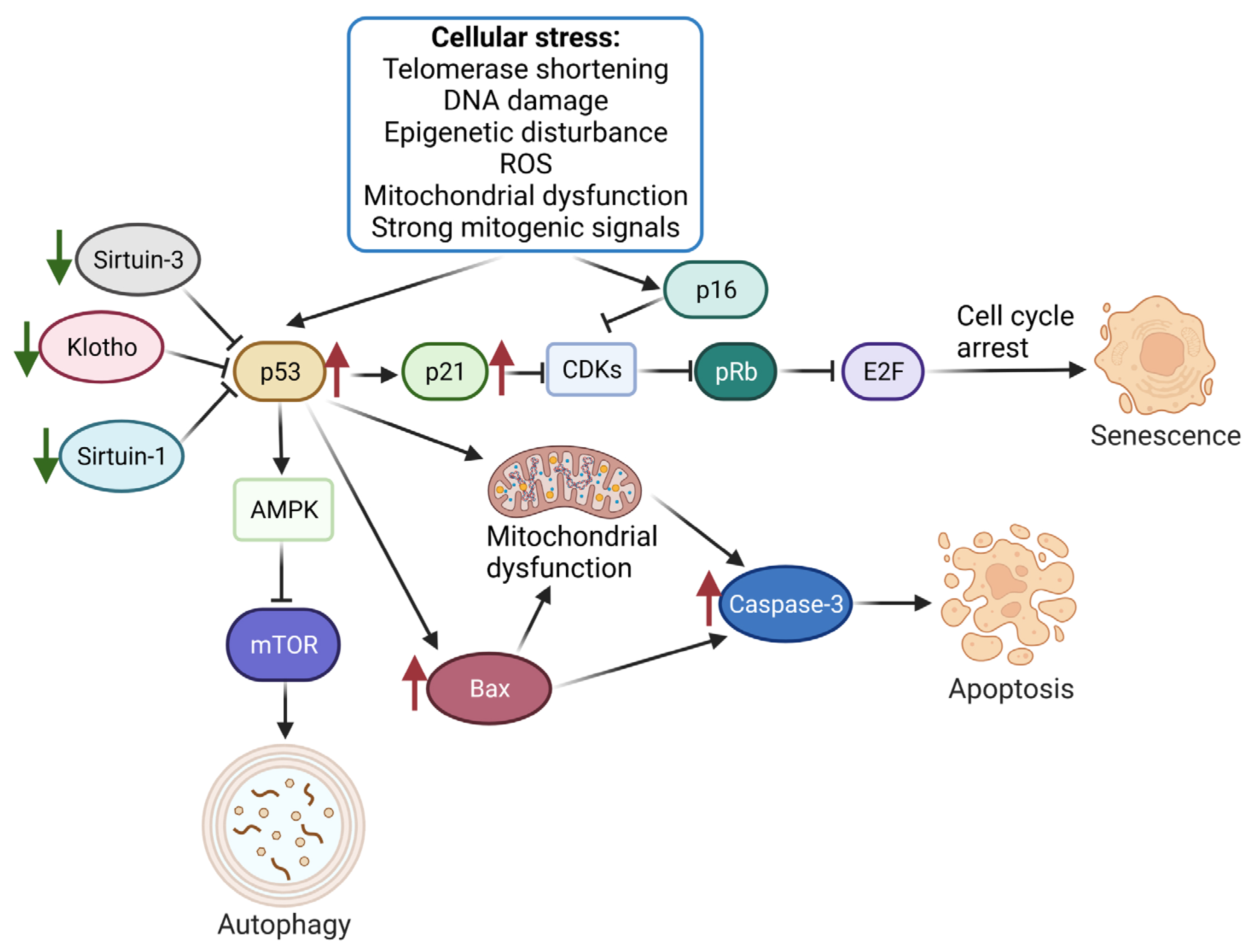
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sultana, Z.; Maiti, K.; Dedman, L.; Smith, R. Is There a Role for Placental Senescence in the Genesis of Obstetric Complications and Fetal Growth Restriction? Am. J. Obstet. Gynecol. 2018, 218, S762–S773. [Google Scholar] [CrossRef] [PubMed]
- Maiti, K.; Sultana, Z.; Aitken, R.J.; Morris, J.; Park, F.; Andrew, B.; Riley, S.C.; Smith, R. Evidence That Fetal Death Is Associated with Placental Aging. Am. J. Obstet. Gynecol. 2017, 217, 441.e1–441.e14. [Google Scholar] [CrossRef] [PubMed]
- Paules, C.; Dantas, A.P.; Miranda, J.; Crovetto, F.; Eixarch, E.; Rodriguez-Sureda, V.; Dominguez, C.; Casu, G.; Rovira, C.; Nadal, A.; et al. Premature Placental Aging in Term Small-for-Gestational-Age and Growth-Restricted Fetuses. Ultrasound Obstet. Gynecol. 2019, 53, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Figueras, F.; Caradeux, J.; Crispi, F.; Eixarch, E.; Peguero, A.; Gratacos, E. Diagnosis and Surveillance of Late-Onset Fetal Growth Restriction. Am. J. Obstet. Gynecol. 2018, 218, S790–S802.e1. [Google Scholar] [CrossRef] [PubMed]
- Gordijn, S.J.; Beune, I.M.; Thilaganathan, B.; Papageorghiou, A.; Baschat, A.A.; Baker, P.N.; Silver, R.M.; Wynia, K.; Ganzevoort, W. Consensus Definition of Fetal Growth Restriction: A Delphi Procedure. Ultrasound Obstet. Gynecol. 2016, 48, 333–339. [Google Scholar] [CrossRef]
- Coutinho, C.M.; Melchiorre, K.; Thilaganathan, B. Stillbirth at Term: Does Size Really Matter? Int. J. Gynaecol. Obstet. 2020, 150, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Man, J.; Hutchinson, J.C.; Ashworth, M.; Heazell, A.E.; Levine, S.; Sebire, N.J. Effects of Intrauterine Retention and Postmortem Interval on Body Weight Following Intrauterine Death: Implications for Assessment of Fetal Growth Restriction at Autopsy. Ultrasound Obstet. Gynecol. 2016, 48, 574–578. [Google Scholar] [CrossRef]
- Gardosi, J. Counterpoint. Am. J. Obstet. Gynecol. 2019, 220, 74–82. [Google Scholar] [CrossRef]
- Poon, L.C.Y.; Volpe, N.; Muto, B.; Syngelaki, A.; Nicolaides, K.H. Birthweight with Gestation and Maternal Characteristics in Live Births and Stillbirths. FDT 2012, 32, 156–165. [Google Scholar] [CrossRef]
- Poon, L.C.Y.; Tan, M.Y.; Yerlikaya, G.; Syngelaki, A.; Nicolaides, K.H. Birth Weight in Live Births and Stillbirths. Ultrasound Obstet. Gynecol. 2016, 48, 602–606. [Google Scholar] [CrossRef]
- Gardosi, J.; Kady, S.M.; McGeown, P.; Francis, A.; Tonks, A. Classification of Stillbirth by Relevant Condition at Death (ReCoDe): Population Based Cohort Study. BMJ 2005, 331, 1113–1117. [Google Scholar] [CrossRef] [PubMed]
- McCowan, L.M.; Figueras, F.; Anderson, N.H. Evidence-Based National Guidelines for the Management of Suspected Fetal Growth Restriction: Comparison, Consensus, and Controversy. Am. J. Obstet. Gynecol. 2018, 218, S855–S868. [Google Scholar] [CrossRef] [PubMed]
- Ego, A.; Zeitlin, J.; Batailler, P.; Cornec, S.; Fondeur, A.; Baran-Marszak, M.; Jouk, P.-S.; Debillon, T.; Cans, C. Stillbirth Classification in Population-Based Data and Role of Fetal Growth Restriction: The Example of RECODE. BMC Pregnancy Childbirth 2013, 13, 182. [Google Scholar] [CrossRef] [PubMed]
- American College of Obstetricians and Gynecologists. ACOG Practice Bulletin No. 134: Fetal Growth Restriction. Obstet. Gynecol. 2013, 121, 1122–1133. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowski, S.; Torbe, A.; Borowski, D.; Breborowicz, G.; Czajkowski, K.; Huras, H.; Kajdy, A.; Kalinka, J.; Kosinska-Kaczynska, K.; Leszczynska-Gorzelak, B.; et al. Polish Society of Gynecologists and Obstetricians Recommendations on Diagnosis and Management of Fetal Growth Restriction. Ginekol. Pol. 2020, 91, 10. [Google Scholar] [CrossRef]
- McLaughlin, E.J.; Hiscock, R.J.; Robinson, A.J.; Hui, L.; Tong, S.; Dane, K.M.; Middleton, A.L.; Walker, S.P.; MacDonald, T.M. Appropriate-for-Gestational-Age Infants Who Exhibit Reduced Antenatal Growth Velocity Display Postnatal Catch-up Growth. PLoS ONE 2020, 15, e0238700. [Google Scholar] [CrossRef]
- Fedarko, N.S. The Biology of Aging and Frailty. Clin. Geriatr. Med. 2011, 27, 27–37. [Google Scholar] [CrossRef]
- Burton, D.G.A. Cellular Senescence, Ageing and Disease. AGE 2009, 31, 1–9. [Google Scholar] [CrossRef]
- Kajdy, A.; Modzelewski, J.; Cymbaluk-Płoska, A.; Kwiatkowska, E.; Bednarek-Jędrzejek, M.; Borowski, D.; Stefańska, K.; Rabijewski, M.; Torbé, A.; Kwiatkowski, S. Molecular Pathways of Cellular Senescence and Placental Aging in Late Fetal Growth Restriction and Stillbirth. Int. J. Mol. Sci. 2021, 22, 4186. [Google Scholar] [CrossRef]
- Ruffaner-Hanson, C.; Noor, S.; Sun, M.S.; Solomon, E.; Marquez, L.E.; Rodriguez, D.E.; Allan, A.M.; Caldwell, K.K.; Bakhireva, L.N.; Milligan, E.D. The Maternal-Placental-Fetal Interface: Adaptations of the HPA Axis and Immune Mediators Following Maternal Stress and Prenatal Alcohol Exposure. Exp. Neurol. 2022, 355, 114121. [Google Scholar] [CrossRef]
- Papargyri, P.; Zapanti, E.; Salakos, N.; Papargyris, L.; Bargiota, A.; Mastorakos, G. Links between HPA Axis and Adipokines: Clinical Implications in Paradigms of Stress-Related Disorders. Expert. Rev. Endocrinol. Metab. 2018, 13, 317–332. [Google Scholar] [CrossRef]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.A.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA Statement for Reporting Systematic Reviews and Meta-Analyses of Studies That Evaluate Health Care Interventions: Explanation and Elaboration. PLoS Med. 2009, 6, e1000100. [Google Scholar] [CrossRef]
- Lo, C.K.-L.; Mertz, D.; Loeb, M. Newcastle-Ottawa Scale: Comparing Reviewers’ to Authors’ Assessments. BMC Med. Res. Methodol. 2014, 14, 45. [Google Scholar] [CrossRef] [PubMed]
- Baethge, C.; Goldbeck-Wood, S.; Mertens, S. SANRA—A Scale for the Quality Assessment of Narrative Review Articles. Res. Integr. Peer Rev. 2019, 4, 5. [Google Scholar] [CrossRef]
- Beard, S.; Pritchard, N.; Binder, N.; Schindler, K.; De Alwis, N.; Kaitu’u-Lino, T.J.; Tong, S.; Hannan, N.J. Aurora Kinase MRNA Expression Is Reduced with Increasing Gestational Age and in Severe Early Onset Fetal Growth Restriction. Placenta 2020, 95, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Naha, R.; Anees, A.; Chakrabarty, S.; Naik, P.S.; Pandove, M.; Pandey, D.; Satyamoorthy, K. Placental Mitochondrial DNA Mutations and Copy Numbers in Intrauterine Growth Restricted (IUGR) Pregnancy. Mitochondrion 2020, 55, 85–94. [Google Scholar] [CrossRef]
- Franklin, A.D.; Saqibuddin, J.; Stephens, K.; Birkett, R.; Marsden, L.; Ernst, L.M.; Mestan, K.K. Cord Blood Alpha Klotho Is Decreased in Small for Gestational Age Preterm Infants with Placental Lesions of Accelerated Aging. Placenta 2019, 87, 1–7. [Google Scholar] [CrossRef]
- Ferrari, F.; Facchinetti, F.; Saade, G.; Menon, R. Placental Telomere Shortening in Stillbirth: A Sign of Premature Senescence? J. Matern.-Fetal Neonatal Med. 2016, 29, 1283–1288. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Rosario, F.J.; Shehab, M.A.; Powell, T.L.; Gupta, M.B.; Jansson, T. Increased Ubiquitination and Reduced Plasma Membrane Trafficking of Placental Amino Acid Transporter SNAT-2 in Human IUGR. Clin. Sci. 2015, 129, 1131–1141. [Google Scholar] [CrossRef] [PubMed]
- Biron-Shental, T.; Sukenik-Halevy, R.; Sharon, Y.; Laish, I.; Fejgin, M.D.; Amiel, A. Telomere Shortening in Intra Uterine Growth Restriction Placentas. Early Hum. Dev. 2014, 90, 465–469. [Google Scholar] [CrossRef]
- Seidmann, L.; Suhan, T.; Unger, R.; Gerein, V.; Kirkpatrick, C.J. Imbalance of Expression of BFGF and PK1 Is Associated with Defective Maturation and Antenatal Placental Insufficiency. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 170, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Biron-Shental, T.; Kidron, D.; Sukenik-Halevy, R.; Goldberg-Bittman, L.; Sharony, R.; Fejgin, M.D.; Amiel, A. TERC Telomerase Subunit Gene Copy Number in Placentas from Pregnancies Complicated with Intrauterine Growth Restriction. Early Hum. Dev. 2011, 87, 73–75. [Google Scholar] [CrossRef] [PubMed]
- Heazell, A.E.P.; Sharp, A.N.; Baker, P.N.; Crocker, I.P. Intra-Uterine Growth Restriction Is Associated with Increased Apoptosis and Altered Expression of Proteins in the P53 Pathway in Villous Trophoblast. Apoptosis 2011, 16, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Biron-Shental, T.; Sukenik-Halevy, R.; Sharon, Y.; Goldberg-Bittman, L.; Kidron, D.; Fejgin, M.D.; Amiel, A. Short Telomeres May Play a Role in Placental Dysfunction in Preeclampsia and Intrauterine Growth Restriction. Am. J. Obstet. Gynecol. 2010, 202, 381.e1–381.e7. [Google Scholar] [CrossRef]
- Biron-Shental, T.; Sukenik Halevy, R.; Goldberg-Bittman, L.; Kidron, D.; Fejgin, M.D.; Amiel, A. Telomeres Are Shorter in Placental Trophoblasts of Pregnancies Complicated with Intrauterine Growth Restriction (IUGR). Early Hum. Dev. 2010, 86, 451–456. [Google Scholar] [CrossRef]
- Davy, P.; Nagata, M.; Bullard, P.; Fogelson, N.S.; Allsopp, R. Fetal Growth Restriction Is Associated with Accelerated Telomere Shortening and Increased Expression of Cell Senescence Markers in the Placenta. Placenta 2009, 30, 539–542. [Google Scholar] [CrossRef]
- Kudo, T.; Izutsu, T.; Sato, T. Telomerase Activity and Apoptosis as Indicators of Ageing in Placenta with and without Intrauterine Growth Retardation. Placenta 2000, 21, 493–500. [Google Scholar] [CrossRef]
- Kohlrausch, F.B.; Keefe, D.L. Telomere Erosion as a Placental Clock: From Placental Pathologies to Adverse Pregnancy Outcomes. Placenta 2020, 97, 101–107. [Google Scholar] [CrossRef]
- Silva, E.; Rosario, F.J.; Powell, T.L.; Jansson, T. Mechanistic Target of Rapamycin Is a Novel Molecular Mechanism Linking Folate Availability and Cell Function. J. Nutr. 2017, 147, 1237–1242. [Google Scholar] [CrossRef]
- Biron-Shental, T.; Sadeh-Mestechkin, D.; Amiel, A. Telomere Homeostasis in IUGR Placentas—A Review. Placenta 2016, 39, 21–23. [Google Scholar] [CrossRef]
- Sultana, Z.; Maiti, K.; Aitken, J.; Morris, J.; Dedman, L.; Smith, R. Oxidative Stress, Placental Ageing-Related Pathologies and Adverse Pregnancy Outcomes. Am. J. Reprod. Immunol. 2017, 77, e12653. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.; Maiti, K.; Aitken, R.J. Unexplained Antepartum Stillbirth: A Consequence of Placental Aging? Placenta 2013, 34, 310–313. [Google Scholar] [CrossRef] [PubMed]
- Heazell, A.E.P.; Crocker, I.P. Live and Let Die—Regulation of Villous Trophoblast Apoptosis in Normal and Abnormal Pregnancies. Placenta 2008, 29, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Crispi, F.; Miranda, J.; Gratacós, E. Long-Term Cardiovascular Consequences of Fetal Growth Restriction: Biology, Clinical Implications, and Opportunities for Prevention of Adult Disease. Am. J. Obstet. Gynecol. 2018, 218, S869–S879. [Google Scholar] [CrossRef] [PubMed]
- Johnson, F.B.; Sinclair, D.A.; Guarente, L. Molecular Biology of Aging. Cell 1999, 96, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Muglu, J.; Rather, H.; Arroyo-Manzano, D.; Bhattacharya, S.; Balchin, I.; Khalil, A.; Thilaganathan, B.; Khan, K.S.; Zamora, J.; Thangaratinam, S. Risks of Stillbirth and Neonatal Death with Advancing Gestation at Term: A Systematic Review and Meta-Analysis of Cohort Studies of 15 Million Pregnancies. PLoS Med. 2019, 16, e1002838. [Google Scholar] [CrossRef]
- Crispi, F.; Figueras, F.; Cruz-Lemini, M.; Bartrons, J.; Bijnens, B.; Gratacos, E. Cardiovascular Programming in Children Born Small for Gestational Age and Relationship with Prenatal Signs of Severity. Am. J. Obstet. Gynecol. 2012, 207, 121.e1–121.e9. [Google Scholar] [CrossRef]
- Burton, G.J.; Yung, H.W.; Murray, A.J. Mitochondrial—Endoplasmic Reticulum Interactions in the Trophoblast: Stress and Senescence. Placenta 2017, 52, 146–155. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Rosario, F.J.; Powell, T.L.; Gupta, M.B.; Jansson, T. Inhibition of Placental Mammalian Target of Rapamycin Complex 2 (MTORC2) Signaling in Human IUGR. Reprod. Sci. 2015, 22, 376A. [Google Scholar]
- Rosario, F.J.; Dimasuay, K.G.; Kanai, Y.; Powell, T.L.; Jansson, T. Regulation of Amino Acid Transporter Trafficking by MTORC1 in Primary Human Trophoblast Cells Is Mediated by the Ubiquitin Ligase Nedd4-2. Clin. Sci. 2016, 130, 499–512. [Google Scholar] [CrossRef]
- Dimasuay, G.; Glazier, J.; Rogerson, S.; Jansson, T.; Boeuf, P. Inhibition of Placental MTORC1 Signalling May Explain Decreased Placental Nutrient Transport and Contribute to Restricted Fetal Growth in Placental Malaria. Placenta 2014, 35, A62. [Google Scholar] [CrossRef]
- Cox, L.S.; Redman, C. The Role of Cellular Senescence in Ageing of the Placenta. Placenta 2017, 52, 139–145. [Google Scholar] [CrossRef]
- Bonney, E.A.; Krebs, K.; Saade, G.; Kechichian, T.; Trivedi, J.; Huaizhi, Y.; Menon, R. Differential Senescence in Feto-Maternal Tissues during Mouse Pregnancy. Placenta 2016, 43, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Menon, R.; Behnia, F.; Polettini, J.; Saade, G.R.; Campisi, J.; Velarde, M. Placental Membrane Aging and HMGB1 Signaling Associated with Human Parturition. Aging 2016, 8, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Bowen, J.M.; Chamley, L.; Keelan, J.A.; Mitchell, M.D. Cytokines of the Placenta and Extra-Placental Membranes: Roles and Regulation During Human Pregnancy and Parturition. Placenta 2002, 23, 257–273. [Google Scholar] [CrossRef] [PubMed]
- Menon, R.; Bonney, E.A.; Condon, J.; Mesiano, S.; Taylor, R.N. Novel Concepts on Pregnancy Clocks and Alarms: Redundancy and Synergy in Human Parturition. Hum. Reprod. Update 2016, 22, 535–560. [Google Scholar] [CrossRef]
- Carter, E.B.; Stockburger, J.; Tuuli, M.G.; Macones, G.A.; Odibo, A.O.; Trudell, A.S. Large for Gestational Age and Stillbirth: Is There a Role for Antenatal Testing? Ultrasound Obstet. Gynecol. 2019, 54, 334–337. [Google Scholar] [CrossRef]
- National Collaborating Centre for Women’s and Children’s Health (UK). Hypertension in Pregnancy: The Management of Hypertensive Disorders During Pregnancy; National Institute for Health and Clinical Excellence: Guidance; RCOG Press: London, UK, 2010. [Google Scholar]
- American College of Obstetricians and Gynecologists. ACOG Practice Bulletin No. 102: Management of Stillbirth. Obstet. Gynecol. 2009, 113, 748–761. [Google Scholar] [CrossRef]
- Reddy, U.M.; Goldenberg, R.; Silver, R.; Smith, G.C.S.; Pauli, R.M.; Wapner, R.J.; Gardosi, J.; Pinar, H.; Grafe, M.; Kupferminc, M.; et al. Stillbirth Classification—Developing an International Consensus for Research: Executive Summary of a National Institute of Child Health and Human Development Workshop. Obstet. Gynecol. 2009, 114, 901–914. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Database | Searching Quote |
|---|---|
| Medline | (“markers of placental aging” OR “cellular senescence” OR “aging” OR “maturation” OR (p53 AND p21) OR (p16 AND pRB) OR “SA-β-gal” OR “mitochondrial dysfunction” OR “mTORC1” OR “telomere shortening” OR “short telomere” OR “telomere attrition”) AND (“placenta” OR “trophoblast” OR “syncytiotrophoblast”) AND (“fetal growth restriction” OR “FGR” OR “intrauterine growth restriction” OR “IUGR” OR “small for gestational age” OR “SGA” OR “stillbirth” OR “Fetal Growth Retardation” [MeSH]) |
| Web of Science | TS = ((“markers of placental aging” OR “cellular senescence” OR “aging” OR “maturation” OR (“p53” AND “p21”) OR (“p16” AND “pRB”) OR “SA-β-gal” OR “mitochondrial dysfunction” OR “mTORC1” OR “telomere shortening” OR “short telomere” OR “telomere attrition”) AND (“placenta” OR “trophoblast” OR “syncytiotrophoblast”) AND (“fetal growth restriction” OR “FGR” OR “intrauterine growth restriction” OR “IUGR” OR “small for gestational age” OR “SGA” OR “stillbirth” OR “Fetal Growth Retardation”)) |
| Scopus and Embase | (TITLE-ABS-KEY(“markers of placental aging”) OR TITLE-ABS-KEY(“cellular senescence”) OR TITLE-ABS-KEY(aging) OR TITLE-ABS-KEY(maturation) OR (TITLE-ABS-KEY(p53) AND TITLE-ABS-KEY(p21)) OR (TITLE-ABS-KEY(p16) AND TITLE-ABS-KEY(pRB)) OR TITLE-ABS-KEY(SA-β-gal) OR TITLE-ABS-KEY(“mitochondrial dysfunction”) OR TITLE-ABS-KEY(mTORC1) OR TITLE-ABS-KEY(“telomere shortening”) OR TITLE-ABS-KEY(“short telomere”) OR TITLE-ABS-KEY(“telomere attrition”)) AND (TITLE-ABS-KEY(placenta) OR TITLE-ABS-KEY(trophoblast) OR TITLE-ABS-KEY(syncytiotrophoblast)) AND (TITLE-ABS-KEY(“fetal growth restriction”) OR TITLE-ABS-KEY(FGR) OR TITLE-ABS-KEY(“intrauterine growth restriction”) OR TITLE-ABS-KEY(IUGR) OR TITLE-ABS-KEY(“small for gestational age”) OR TITLE-ABS-KEY(SGA) OR TITLE-ABS-KEY(stillbirth) OR TITLE-ABS-KEY(“Fetal Growth Retardation”)) |
| Author | Year | Population | Key Points Relevant to Late SGA, FGR, and Stillbirth Research |
|---|---|---|---|
| Beard et al. [25] | 2020 | 13 FGR (8 late FGR) 51 PE 14 PE/FGR 17 term controls | Placental AURK A, B, and C expression decreased with gestational age. Circulating AURKB mRNA reduced at term in comparison to pregnancies <34 weeks. AURKC reduced significantly in placentas from pregnancies complicated by severe early-onset FGR. No change in placental AURKC expression in FGR at later gestations. |
| Naha et al. [26] | 2020 | 29 controls 18 FGR 25 SGA | Significant increase in mtDNA copy number in both FGR and SGA compared to control. Sirtuin-3 (Sirt3) protein expression was significantly downregulated in FGR placenta, but there was no significant difference in Nrf1 expression compared to the control group. |
| Franklin et al. [27] | 2019 | 22 SGA 32 AGA | Alpha klotho levels in cord blood were found to be lower in cases of SGA and maternal vascular malperfusion. These findings suggest a potential link between alpha klotho and the accelerated maturation of placental villi, which is often associated with increased levels of angiopoietin 2. This indicated that alpha klotho may contribute to vascular-mediated placental aging, which can lead to fetal growth restriction. |
| Maiti et al. [2] | 2017 | 34 GA 37–39 28 late term 4 stillbirth | Aldehyde oxidase 1 and G-protein-coupled estrogen receptor 1 mediate aging of the placenta in stillbirth. |
| Ferrari et al. [28] | 2015 | 42 stillbirths 43 live term births 15 live preterm births | Telomere length attrition was an essential feature in unexplained SB. |
| Chen et al. [29] | 2015 | 25 FGR 19 AGA | The activity of mTORC1 and mTORC2 was decreased, and the protein expression of the ubiquitin ligase NEDD4-2 and the ubiquitination of SNAT-2 were increased in FGR placentas. FGR was associated with decreased system A-amino acid transport activity and SNAT-1 and SNAT-2 protein expression in MVM. |
| Biron-Shental et al. [30] | 2014 | 15 controls 15 FGR | Significant increase in telomere capture in FGR trophoblasts compared to controls. Compensatory response of the FGR placentas to shortened telomeres to maintain homeostasis. Demonstrated the presence of senescence-associated heterochromatin bodies by exposing the involvement of hTERT mRNA and telomere capture mechanisms. |
| Seidmann et al. [31] | 2013 | 45 term placentas 6 FGR 3 stillbirths | Defective placental maturation was associated with an imbalance of expression of bFGF and PK1. |
| Paules et al. [3] | 2013 | 21 controls 18 SGA 18 FGR | Both SGA and FGR cases exhibited accelerated placental aging, lower telomerase activity, shorter telomeres, decreased SIRT1 RNA expression, and increased P53 RNA expression. In FGR cases, there was also evidence of increased apoptosis, as evidenced by elevated CASP3 RNA levels. Furthermore, there was a linear trend between the severity of the condition and the levels of SIRT1 RNA expression, telomerase activity, telomere length, and Caspase-3 activity. |
| Biron-Shental et al. [32] | 2011 | 5 controls 5 FGR | The TERC gene copy number was decreased in FGR trophoblasts. |
| Heazell et al. [33] | 2011 | 6 normal 6 FGR | In cases of FGR, there was an observed increase in the expression of P53 mRNA and protein, which were found to be localized in the syncytiotrophoblast. Similar changes were also observed in the expression of P21 and BAX. However, no significant changes were noted in the expression of Mdm2, Bak, and Bcl2. The increased expression of p53 was thought to be associated with altered trophoblast cell turnover, which is similar to the effects of hypoxia exposure. |
| Biron-Shental et al. [34] | 2010 | 20 controls 14 PE/FGR 14 FGR 9 severe PE/FGR | Telomeres were shorter in PE and FGR placentas. Telomere aggregate formation increased in PE but not FGR. This implied different placental stress-related mechanisms in PE with and without FGR. |
| Biron-Shental et al. [35] | 2010 | 13 controls 16 FGR | The number and intensity of telomeres staining and telomerase activity were lower in FGR placentas. Telomeres were shorter in trophoblasts of FGR placentas. |
| Davy et al. [36] | 2009 | 32 FGR 36 controls | Significantly shorter telomeres in FGR placenta samples, but not cord blood samples. There was an association between the suppression of telomerase activity and reduced telomere length in FGR placenta. p21, p16, and EF-1 alpha were significantly elevated in FGR placenta samples. |
| Kudo et al. [37] | 2000 | Group A: 31 first trimester chorionic villi specimens of normal pregnancy undergoing abortion Group B: 32 without FGR Group C: 12 with FGR | TA was detected both in CVS and placentas without FGR. Weak TA was observed in FGR. Significantly higher Bcl-2 immunoreactivity was seen in Group A and Group B than in Group C. TUNEL-positive cells were significantly more present in Group C than in Group A or Group B. |
| Year | Author | Type of Review | SANRA Quality Assessment | Key Points Relevant to Late SGA, FGR, and Stillbirth Research |
|---|---|---|---|---|
| 2020 | Fabiana et al. [38] | Narrative Review | 6/12 | Telomere erosion in the placenta, with concomitant cellular senescence, may have contributed to SB. Studies that compared telomere length in FGR vs. aged-matched control placentas observed shorter telomeres in FGR placentas. No difference in TERC copy numbers between FGR and control, despite reduced telomere length in FGR placentas. |
| 2017 | Silva et al. [39] | Narrative Review | 8/12 | mTOR folate sensing linked folate availability and cell function. Placental mTOR was blocked when there were low maternal folate concentrations and caused a decrease in placental amino acid transport, protein synthesis, and mitochondrial respiration. Fetal growth restriction resulted from the unavailability of nutrients. |
| 2017 | Sultana et al. [1] | Expert Review | 6/12 | FGR coexisted with increased expression of DNA damage biomarkers, reduction of TL and TA, upregulation of p53 and p16, and elevated levels of senescence-associated secretory phenotype and SAHF. Late gestational tissues also showed evidence of placental oxidative DNA damage and premature senescence. |
| 2016 | Biron-Shental et al. [40] | Narrative Review | 6/12 | Telomere homeostasis should be compared in early vs. late-onset FGR. Potential benefits may show oxygen supplementation. Studies of cultured human trophoblasts might help with understanding the mechanisms of placental injury. |
| 2016 | Sultana et al. [41] | Narrative Review | 6/12 | A significantly shorter telomere and/or absent or reduced telomerase activity was observed in the placentas from FGR pregnancies with a reduced expression of hTERT. The expression of telomere-induced senescence markers P21 and P16 was elevated, and the anti-apoptotic protein Bcl-2 was decreased. OS led to placental insufficiency and an inability to meet the growing fetal demands, leading to death. |
| 2013 | Smith et al. [42] | Narrative Review | 6/12 | Unexplained antepartum stillbirths occurred as a consequence of changes in the late gestation placenta. Small numbers of pregnancies continued beyond 40 weeks; therefore, the adverse effects of genes that produce aging affected fewer pregnancies, and the polymorphisms in genes that produced these effects remained in the population. |
| 2008 | Heazell et al. [43] | Narrative review | 10/12 | Increased apoptosis rate resulting from the activation of the extrinsic or intrinsic apoptotic pathways. Activation of the apoptotic pathway caused an alteration in downstream effector proteins, and specific post-translational modifications/evaluation of oncoprotein expression patterns may have identified candidates that induce apoptosis, such as placental hypoxia, reactive oxygen species, or a reduction in growth factors. Understanding cell turnover pathways may have shown molecular targets capable of inhibiting apoptosis and providing a novel therapeutic approach. |
| Selection | Comparability (* or **) d | Assessment of Outcome (⋆) e | Total (6*) | |||
|---|---|---|---|---|---|---|
| Representativeness of Exposed Cohort (⋆) a | Selection of Non-Exposed Cohort (⋆) b | Ascertainment of Exposure (⋆) c | ||||
| Beard et al. [25] | * | * | * | ** | * | ****** (6) |
| Naha et al. [26] | - | * | * | * | * | ***** (5) |
| Franklin et al. [27] | * | * | - | ** | * | ***** (5) |
| Maiti et al. [2] | * | * | * | ** | * | ****** (6) |
| Ferrari et al. [28] | * | * | * | *- | * | ***** (5) |
| Chen et al. [29] | * | * | * | ** | * | ****** (6) |
| Biron-Shental et al. [30] | * | * | * | *- | * | ***** (5) |
| Seidmann et al. [31] | * | * | * | ** | * | ****** (6) |
| Paules et al. [3] | * | * | * | ** | * | ****** (6) |
| Biron-Shental et al. [32] | * | * | * | * | **** (4) | |
| Heazell et al. [33] | * | * | * | *- | * | ***** (5) |
| Biron-Shental et al. [34] | * | * | * | ** | * | ****** (6) |
| Biron-Shental et al. [35] | * | * | ** | * | ***** (5) | |
| Davy et al. [36] | * | * | * | * | **** (4) | |
| Kudo et al. [37] | * | * | *? | *** (3) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kajdy, A.; Sys, D.; Modzelewski, J.; Bogusławska, J.; Cymbaluk-Płoska, A.; Kwiatkowska, E.; Bednarek-Jędrzejek, M.; Borowski, D.; Stefańska, K.; Rabijewski, M.; et al. Evidence of Placental Aging in Late SGA, Fetal Growth Restriction and Stillbirth—A Systematic Review. Biomedicines 2023, 11, 1785. https://doi.org/10.3390/biomedicines11071785
Kajdy A, Sys D, Modzelewski J, Bogusławska J, Cymbaluk-Płoska A, Kwiatkowska E, Bednarek-Jędrzejek M, Borowski D, Stefańska K, Rabijewski M, et al. Evidence of Placental Aging in Late SGA, Fetal Growth Restriction and Stillbirth—A Systematic Review. Biomedicines. 2023; 11(7):1785. https://doi.org/10.3390/biomedicines11071785
Chicago/Turabian StyleKajdy, Anna, Dorota Sys, Jan Modzelewski, Joanna Bogusławska, Aneta Cymbaluk-Płoska, Ewa Kwiatkowska, Magdalena Bednarek-Jędrzejek, Dariusz Borowski, Katarzyna Stefańska, Michał Rabijewski, and et al. 2023. "Evidence of Placental Aging in Late SGA, Fetal Growth Restriction and Stillbirth—A Systematic Review" Biomedicines 11, no. 7: 1785. https://doi.org/10.3390/biomedicines11071785
APA StyleKajdy, A., Sys, D., Modzelewski, J., Bogusławska, J., Cymbaluk-Płoska, A., Kwiatkowska, E., Bednarek-Jędrzejek, M., Borowski, D., Stefańska, K., Rabijewski, M., Baran, A., Torbe, A., Feduniw, S., & Kwiatkowski, S. (2023). Evidence of Placental Aging in Late SGA, Fetal Growth Restriction and Stillbirth—A Systematic Review. Biomedicines, 11(7), 1785. https://doi.org/10.3390/biomedicines11071785