

Generation and Characterization of a Human-Derived and Induced Pluripotent Stem Cell (iPSC) Line from an Alzheimer’s Disease Patient with Neuropsychiatric Symptoms

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
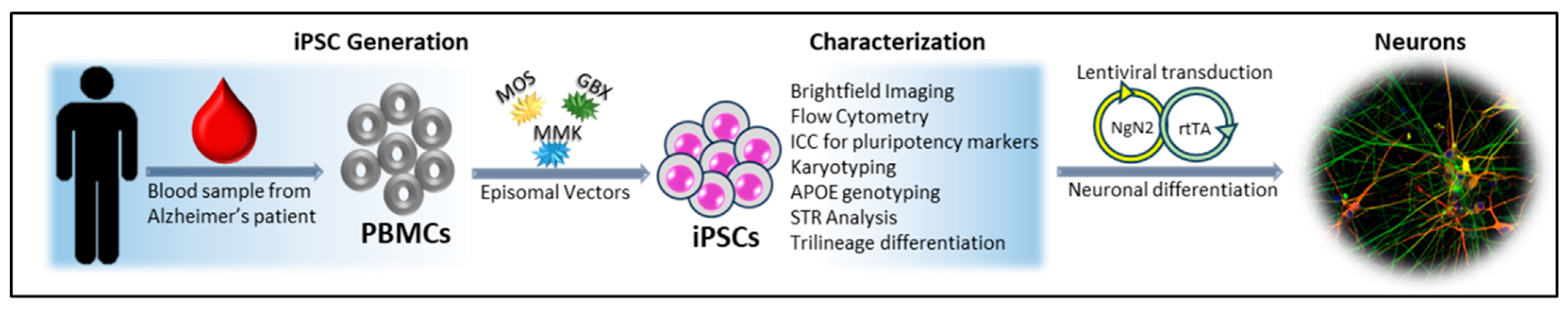
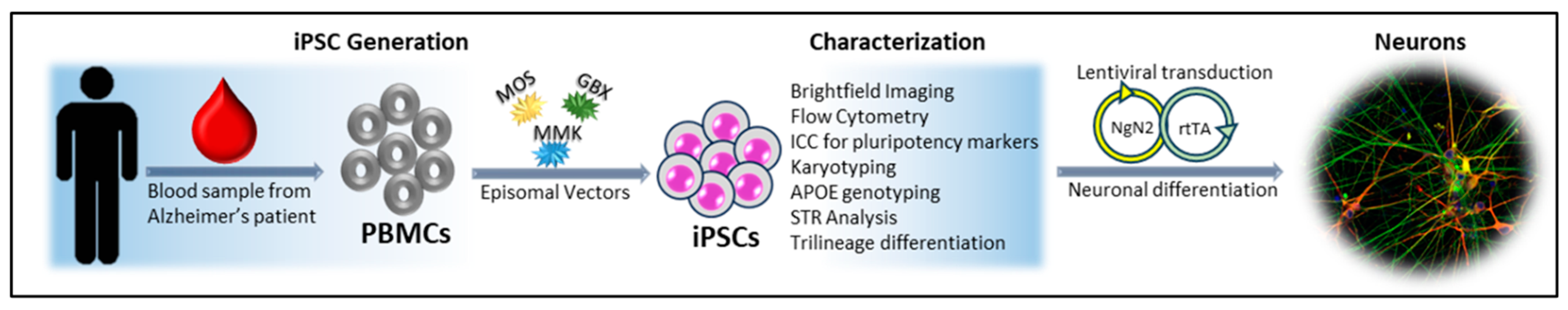
2.1. Human Induced Pluripotent Stem Cell Generation
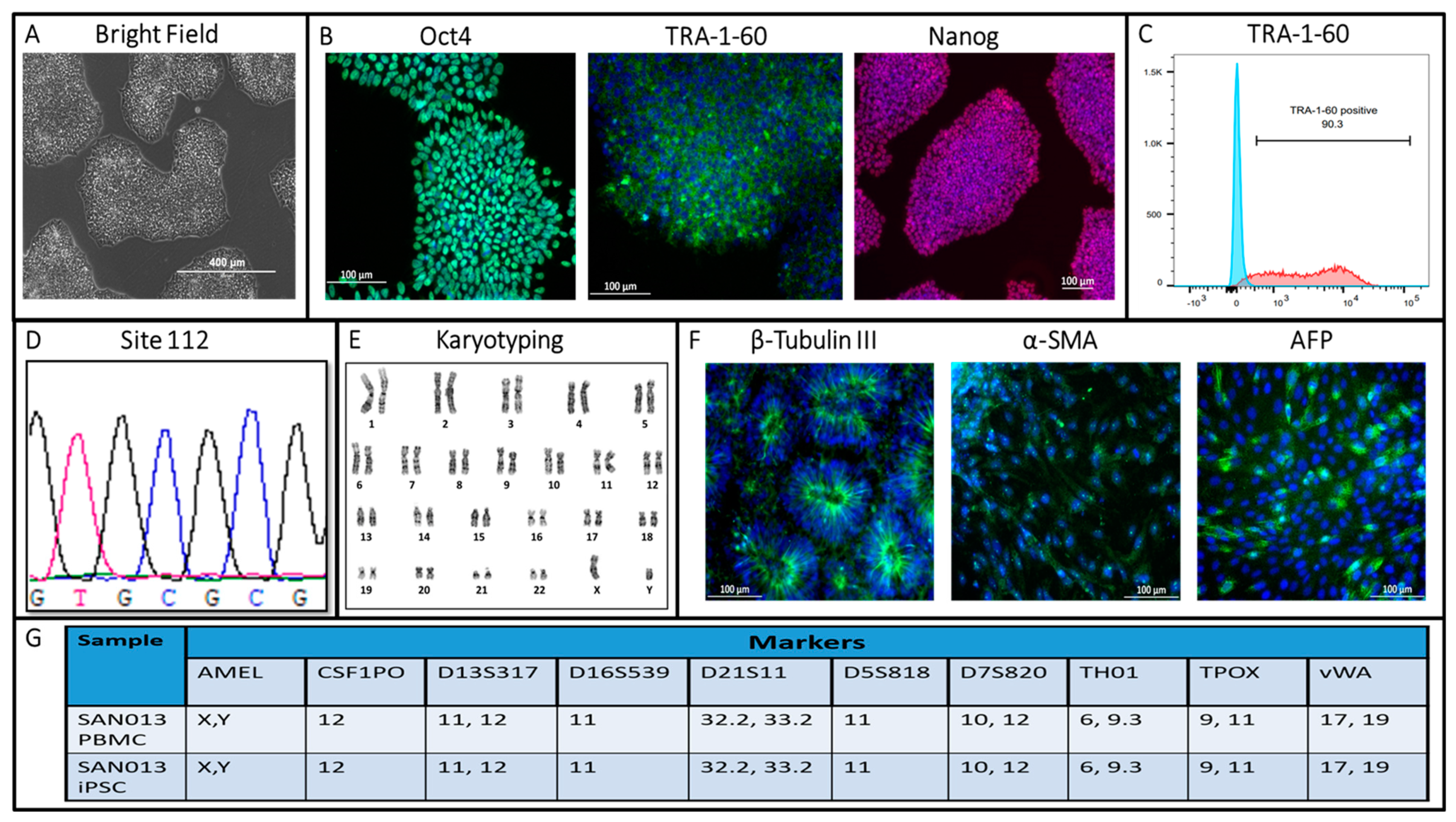
2.2. Flow Cytometric Analysis
2.3. APOE Genotyping
2.4. Immunocytochemistry Staining for Pluripotency Markers
2.5. In Vitro Trilineage Differentiation
2.6. Karyotyping Analysis
2.7. Short Tandem Repeats (STRs) Analysis
2.8. Mycoplasma Detection
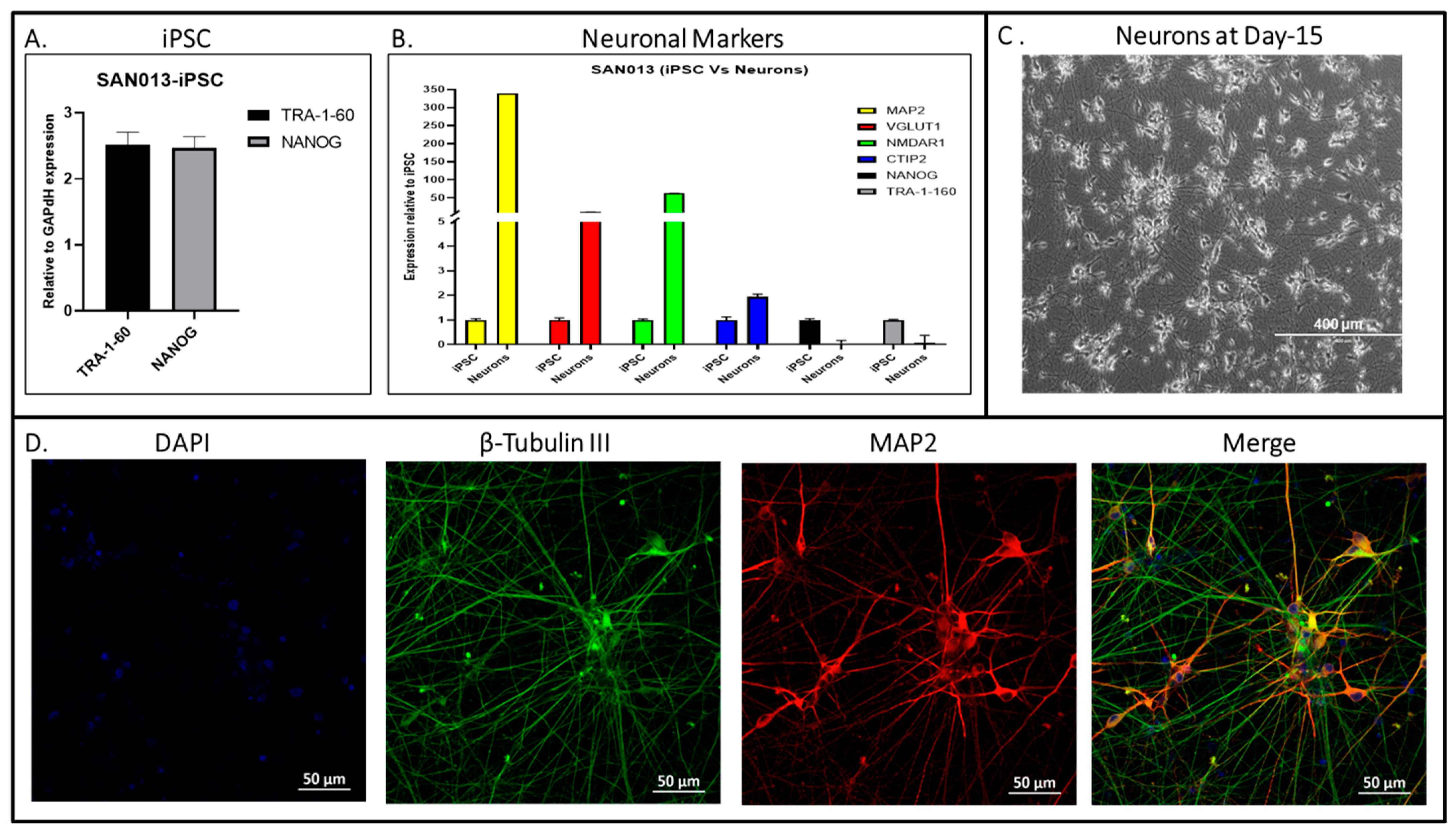
2.9. Lentiviral Transduction and Neuronal Differentiation
2.10. Immunocytochemistry Staining for Neuronal Markers
2.11. Quantitative RT-PCR
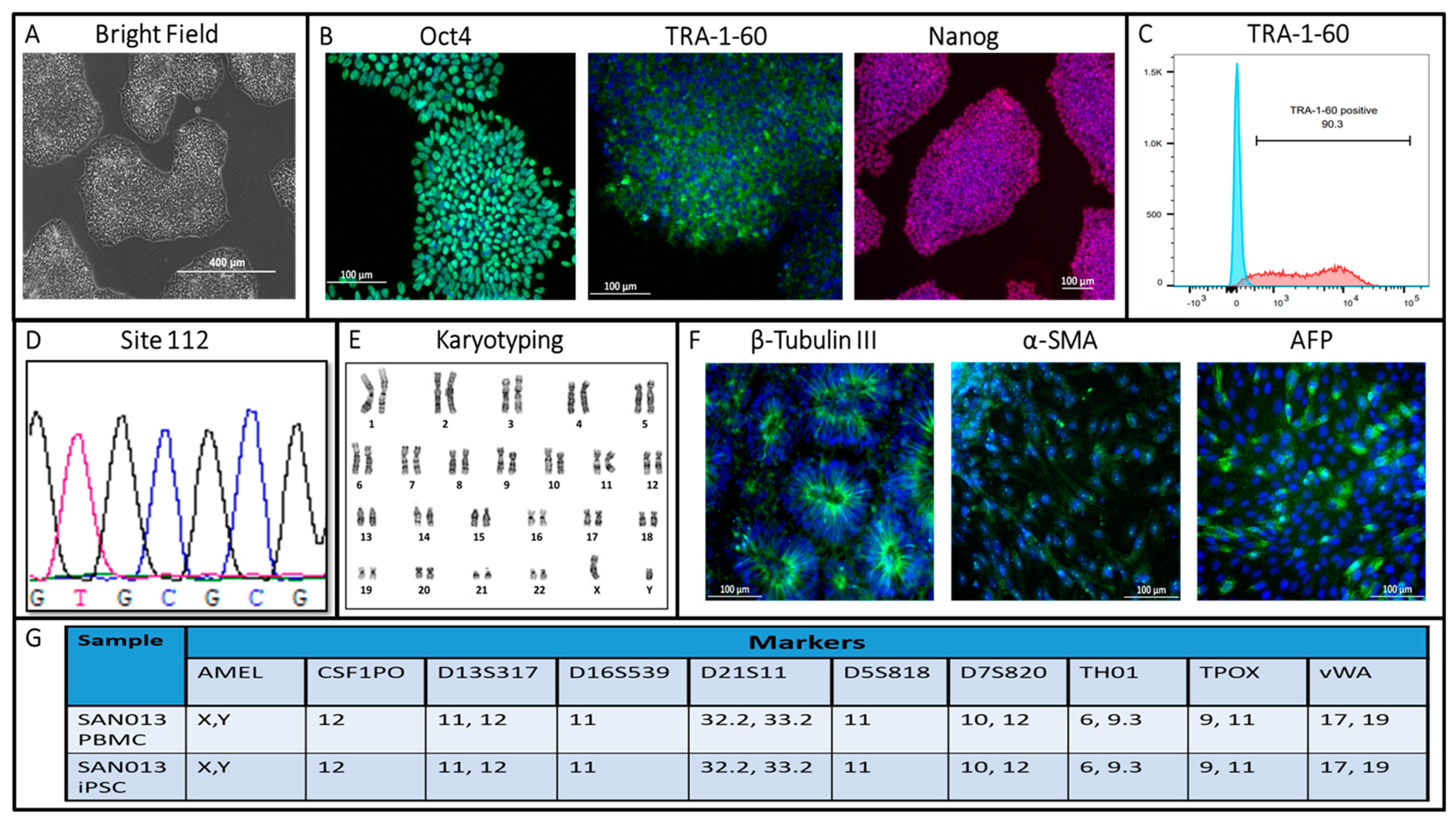
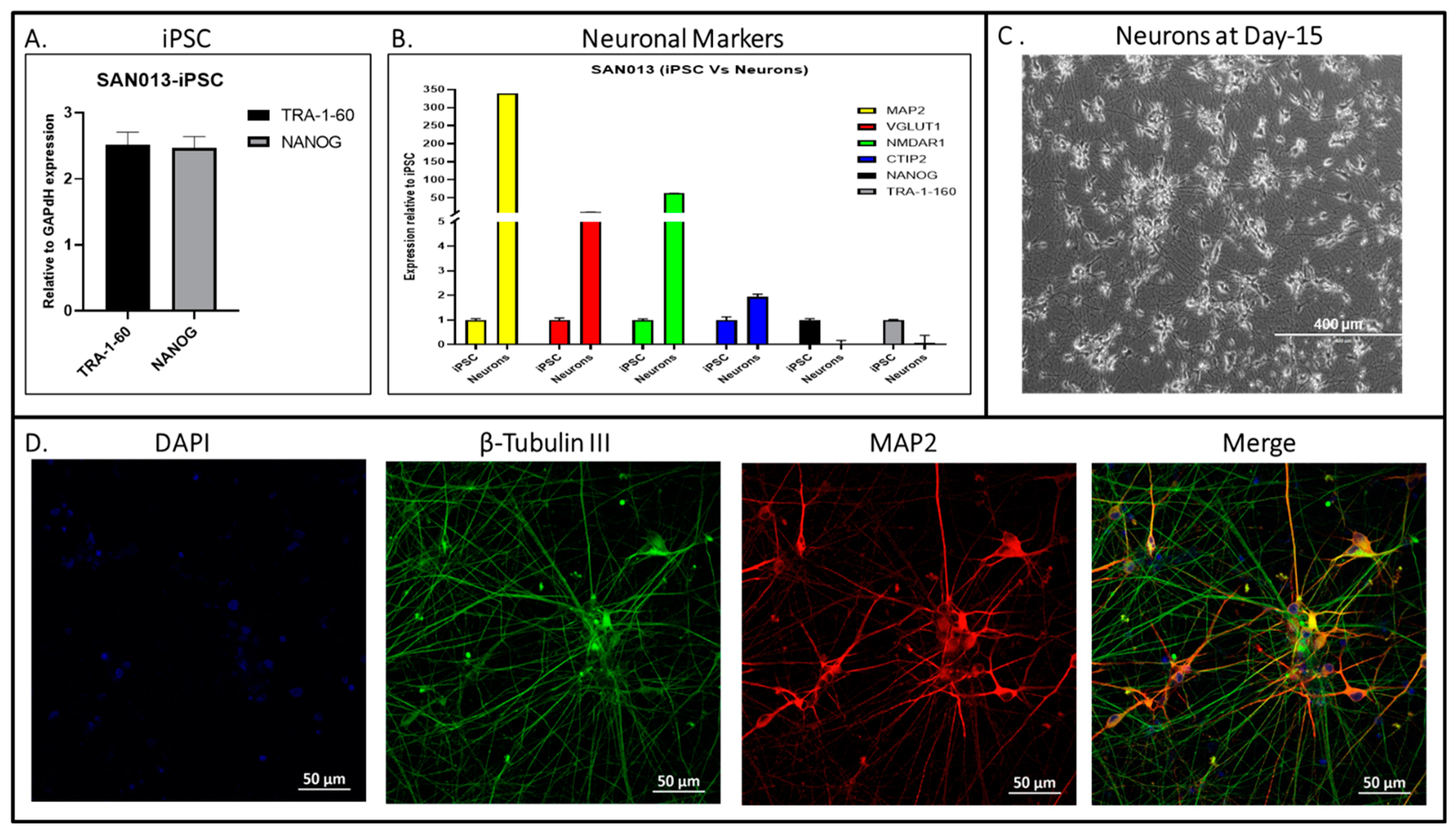
3. Results and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lyketsos, C.G.; Carrillo, M.C.; Ryan, J.M.; Khachaturian, A.S.; Trzepacz, P.; Amatniek, J.; Cedarbaum, J.; Brashear, R.; Miller, D.S. Neuropsychiatric symptoms in Alzheimer’s disease. Alzheimer’s Dement. 2011, 7, 532–539. [Google Scholar] [CrossRef]
- Soto, M.; Andrieu, S.; Nourhashemi, F.; Ousset, P.J.; Ballard, C.; Robert, P.; Vellas, B.; Lyketsos, C.G.; Rosenberg, P.B. Medication development for agitation and aggression in Alzheimer disease: Review and discussion of recent randomized clinical trial design. Int. Psychogeriatr. 2014, 27, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Lanctôt, K.L.; Amatniek, J.; Ancoli-Israel, S.; Arnold, S.E.; Ballard, C.; Cohen-Mansfield, J.; Ismail, Z.; Lyketsos, C.; Miller, D.S.; Musiek, E.; et al. Neuropsychiatric signs and symptoms of Alzheimer’s disease: New treatment paradigms. Alzheimer’s Dement. 2017, 3, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Geda, Y.E.; Schneider, L.S.; Gitlin, L.N.; Miller, D.S.; Smith, G.S.; Bell, J.; Evans, J.; Lee, M.; Porsteinsson, A.; Lanctôt, K.L.; et al. Neuropsychiatric symptoms in Alzheimer’s disease: Past progress and anticipation of the future. Alzheimer’s Dement. 2013, 9, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Rüb, U.; Del Tredici, K.; Schultz, C.; Thal, D.R.; Braak, E.; Braak, H. The evolution of Alzheimer’s disease-related cytoskeletal pathology in the human raphe nuclei. Neuropathol. Appl. Neurobiol. 2000, 26, 553–567. [Google Scholar] [CrossRef] [PubMed]
- Zweig, R.M.; Ross, C.A.; Hedreen, J.C.; Steele, C.; Cardillo, J.E.; Whitehouse, P.J.; Folstein, M.F.; Price, D.L. The neuropathology of aminergic nuclei in Alzheimer’s disease. Ann. Neurol. 1988, 24, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Lyness, S.A.; Zarow, C.; Chui, H.C. Neuron loss in key cholinergic and aminergic nuclei in Alzheimer disease: A meta-analysis. Neurobiol. Aging 2003, 24, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Assal, F.; Alarcón, M.; Solomon, E.C.; Masterman, D.; Geschwind, D.H.; Cummings, J.L. Association of the serotonin transporter and receptor gene polymorphisms in neuropsychiatric symptoms in Alzheimer disease. Arch. Neurol. 2004, 61, 1249–1253. [Google Scholar] [CrossRef]
- Garcia-Alloza, M.; Gil-Bea, F.J.; Diez-Ariza, M.; Chen, C.P.L.-H.; Francis, P.T.; Lasheras, B.; Ramirez, M.J. Cholinergic-serotonergic imbalance contributes to cognitive and behavioral symptoms in Alzheimer’s disease. Neuropsychologia 2005, 43, 442–449. [Google Scholar] [CrossRef]
- Vermeiren, Y.; Van Dam, D.; Aerts, T.; Engelborghs, S.; De Deyn, P.P. Brain Region-Specific Monoaminergic Correlates of Neuropsychiatric Symptoms in Alzheimer’s Disease. J. Alzheimer’s Dis. 2014, 41, 819–833. [Google Scholar] [CrossRef]
- Palmer, A.M. Neurochemical studies of Alzheimer’s disease. Neurodegeneration 1996, 5, 381–391. [Google Scholar] [CrossRef]
- Schneider, L.S.; Frangakis, C.; Drye, L.T.; Devanand, D.P.; Marano, C.M.; Mintzer, J.; Mulsant, B.H.; Munro, C.A.; Newell, J.A.; Pawluczyk, S.; et al. Heterogeneity of Treatment Response to Citalopram for Patients With Alzheimer’s Disease With Aggression or Agitation: The CitAD Randomized Clinical Trial. Am. J. Psychiatry 2016, 173, 465–472. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef]
- Yu, J.; Hu, K.; Smuga-Otto, K.; Tian, S.; Stewart, R.; Slukvin, I.I.; Thomson, J.A. Human Induced Pluripotent Stem Cells Free of Vector and Transgene Sequences. Science 2009, 324, 797–801. [Google Scholar] [CrossRef]
- Nishikawa, S.; Goldstein, R.A.; Nierras, C.R. The promise of human induced pluripotent stem cells for research and therapy. Nat. Rev. Mol. Cell Biol. 2008, 9, 725–729. [Google Scholar] [CrossRef]
- Liu, G.; David, B.T.; Trawczynski, M.; Fessler, R.G. Advances in Pluripotent Stem Cells: History, Mechanisms, Technologies, and Applications. Stem. Cell Rev. Rep. 2020, 16, 3–32. [Google Scholar] [CrossRef]
- Das, D.; Li, J.; Liu, S.; Oh, E.; Cheng, L.; Lyketsos, C.; Mahairaki, V. Generation and characterization of a novel human iPSC line from a resilient Alzheimer’s disease patient. Stem Cell Res. 2020, 48, 101979. [Google Scholar] [CrossRef]
- Chou, B.-K.; Mali, P.; Huang, X.; Ye, Z.; Dowey, S.N.; Resar, L.M.; Zou, C.; Zhang, Y.A.; Tong, J.; Cheng, L. Efficient human iPS cell derivation by a non-integrating plasmid from blood cells with unique epigenetic and gene expression signatures. Cell Res. 2011, 21, 518–529. [Google Scholar] [CrossRef] [PubMed]
- Chou, B.-K.; Gu, H.; Gao, Y.; Dowey, S.N.; Wang, Y.; Shi, J.; Li, Y.; Ye, Z.; Cheng, T.; Cheng, L. A facile method to establish human induced pluripotent stem cells from adult blood cells under feeder-free and xeno-free culture conditions: A clinically compliant approach. Stem Cells Transl. Med. 2015, 4, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Sagar, R.; Azoidis, I.; Zivko, C.; Xydia, A.; Oh, E.S.; Rosenberg, P.B.; Lyketsos, C.G.; Mahairaki, V.; Avramopoulos, D. Excitatory Neurons Derived from Human-Induced Pluripotent Stem Cells Show Transcriptomic Differences in Alzheimer’s Patients from Controls. Cells 2023, 12, 1990. [Google Scholar] [CrossRef] [PubMed]
- Finkel, S.I.; Costa e Silva, J.; Cohen, G.; Miller, S.; Sartorius, N. Behavioral and psychological signs and symptoms of dementia: A consensus statement on current knowledge and implications for research and treatment. Int. Psychogeriatr. 1996, 8 (Suppl. 3), 497–500. [Google Scholar] [CrossRef]
- O’Donnell, B.F.; Drachman, D.A.; Barnes, H.J.; Peterson, K.E.; Swearer, J.M.; Lew, R.A. Incontinence and troublesome behaviors predict institutionalization in dementia. J. Geriatr. Psychiatry Neurol. 1992, 5, 45–52. [Google Scholar] [CrossRef]
- Scarmeas, N.; Brandt, J.; Albert, M.; Hadjigeorgiou, G.; Papadimitriou, A.; Dubois, B.; Sarazin, M.; Devanand, D.; Honig, L.; Marder, K.; et al. Delusions and hallucinations are associated with worse outcome in Alzheimer disease. Arch. Neurol. 2005, 62, 1601–1608. [Google Scholar] [CrossRef]
- Aalten, P.; de Vugt, M.E.; Lousberg, R.; Korten, E.; Jaspers, N.; Senden, B.; Jolles, J.; Verhey, F.R.J. Behavioral Problems in Dementia: A Factor Analysis of the Neuropsychiatric Inventory. Dement. Geriatr. Cogn. Disord. 2003, 15, 99–105. [Google Scholar] [CrossRef]
- Cohen-Mansfield, J.; Werner, P. Longitudinal predictors of non-aggressive agitated behaviors in the elderly. Int. J. Geriatr. Psychiatry 1999, 14, 831–844. [Google Scholar] [CrossRef]
- Defrancesco, M.; Marksteiner, J.; Kemmler, G.; Dal-Bianco, P.; Ransmayr, G.; Benke, T.; Mosbacher, J.; Höller, Y.; Schmidt, R. Specific Neuropsychiatric Symptoms Are Associated with Faster Progression in Alzheimer’s Disease: Results of the Prospective Dementia Registry (PRODEM-Austria). J. Alzheimer’s Dis. 2020, 73, 125–133. [Google Scholar] [CrossRef]
- Koppel, J.; Goldberg, T.E.; Gordon, M.L.; Huey, E.; Davies, P.; Keehlisen, L.; Huet, S.; Christen, E.; Greenwald, B.S. Relationships between behavioral syndromes and cognitive domains in Alzheimer disease: The impact of mood and psychosis. Am. J. Geriatr. Psychiatry 2012, 20, 994–1000. [Google Scholar] [CrossRef]
- Cacabelos, R.; Rodríguez, B.; Carrera, C.; Beyer, K.; Lao, J.I.; Sellers, M.A. Behavioral changes associated with different apolipoprotein E genotypes in dementia. Alzheimer Dis. Assoc. Disord. 1997, 11 (Suppl. 4), S27–S34. [Google Scholar] [PubMed]
- Murphy, G.M.; Taylor, J.; Tinklenberg, J.R.; Yesavage, J.A. The apolipoprotein E epsilon 4 allele is associated with increased behavioral disturbance in Alzheimer’s disease. Am. J. Geriatr. Psychiatry 1997, 5, 88–89. [Google Scholar] [CrossRef]
- Craig, D.; Hart, D.J.; McCool, K.; McIlroy, S.P.; Passmore, A.P. Apolipoprotein E e4 allele influences aggressive behaviour in Alzheimer’s disease. J. Neurol. Neurosurg. Psychiatry 2004, 75, 1327–1330. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.; Mintzer, J.; Brodaty, H.; Sano, M.; Banerjee, S.; Devanand, D.P.; Gauthier, S.; Howard, R.; Lanctôt, K.; Lyketsos, C.G.; et al. Agitation in cognitive disorders: International Psychogeriatric Association provisional consensus clinical and research definition. Int. Psychogeriatr. 2015, 27, 7–17. [Google Scholar] [CrossRef]
- De Los Angeles, A.; Fernando, M.B.; Hall, N.A.L.; Brennand, K.J.; Harrison, P.J.; Maher, B.J.; Weinberger, D.R.; Tunbridge, E.M. Induced Pluripotent Stem Cells in Psychiatry: An Overview and Critical Perspective. Biol. Psychiatry 2021, 90, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-T.; Bendriem, R.M.; Wu, W.W.; Shen, R.-F. 3D brain Organoids derived from pluripotent stem cells: Promising experimental models for brain development and neurodegenerative disorders. J. Biomed. Sci. 2017, 24, 59. [Google Scholar] [CrossRef]
- Mesquita, F.C.P.; Hochman-Mendez, C.; Taylor, D.A. Chapter 8—Strategies for iPSC expansion: From feeder cells to laminin. In Recent Advances in iPSC Technology; Birbrair, A., Ed.; Advances in Stem Cell Biology; Academic Press: Cambridge, MA, USA, 2021; Volume 5, pp. 209–229. ISBN 978-0-12-822231-7. [Google Scholar] [CrossRef]
- Hendricksen, M.; Thomas, A.J.; Ferrier, I.N.; Ince, P.; O’Brien, J.T. Neuropathological study of the dorsal raphe nuclei in late-life depression and Alzheimer’s disease with and without depression. Am. J. Psychiatry 2004, 161, 1096–1102. [Google Scholar] [CrossRef]
- Šimić, G.; Leko, M.B.; Wray, S.; Harrington, C.; Delalle, I.; Jovanov-Milošević, N.; Bažadona, D.; Buée, L.; de Silva, R.; Di Giovanni, G.; et al. Monoaminergic Neuropathology in Alzheimer’s disease. Prog. Neurobiol. 2017, 151, 101–138. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zhong, X.; Liu, H.; Hao, L.; Huang, C.T.-L.; Sherafat, M.A.; Jones, J.; Ayala, M.; Li, L.; Zhang, S.-C. Generation of serotonin neurons from human pluripotent stem cells. Nat. Biotechnol. 2016, 34, 89–94. [Google Scholar] [CrossRef]
- Valiulahi, P.; Vidyawan, V.; Puspita, L.; Oh, Y.; Juwono, V.B.; Sittipo, P.; Friedlander, G.; Yahalomi, D.; Sohn, J.-W.; Lee, Y.K.; et al. Generation of caudal-type serotonin neurons and hindbrain-fate organoids from hPSCs. Stem Cell Rep. 2021, 16, 1938–1952. [Google Scholar] [CrossRef]
- Wadman, M. FDA no longer has to require animal testing for new drugs. Science 2023, 379, 127–128. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Antibody and Host Species | Dilution | Manufacturer | |
|---|---|---|---|
| Pluripotency markers | TRA-1-60, Mouse IgM | 1:300 | EMD Millipore, MAB4360, RRID:AB_3548341 |
| OCT4, Rabbit IgG | 1:200 | Santa Cruz Biotechnology, sc-9081, RRID:AB_E1011 | |
| NANOG, Mouse IgG | 1:200 | BD Biosciences, 560482, RRID:AB_8240787 | |
| Differentiation markers | β-tubulin III, Mouse IgG | 1:500 | Biolegend, 801201, RRID:AB_B205808 |
| AFP, Mouse IgG | 1:100 | Thermo Fisher Scientific, MA5-14666, RRID:AB_VI308040 | |
| α-SMA, Mouse IgG | 1:50 | EMD Millipore, CBL171, RRID:AB_2223166 | |
| MAP2 (D5G1), Rabbit IgG | 1:400 | Cell Signaling, 8707, D5G1, RRID:AB_2722660 | |
| Secondary antibodies | Alexa Fluor 555 goat anti-mouse IgM | 1:500 | Thermo Fisher Scientific, A-21426, RRID:AB_2128995 |
| Alexa Fluor 488 goat anti-mouse IgG | 1:500 | Thermo Fisher Scientific, A-11001, RRID:AB_1907294 | |
| Alexa Fluor 555 goat anti-Rabbit IgG | 1:500 | Thermo Fisher Scientific, A-21428, RRID:AB_ 2535849 | |
| Alexa Fluor 488 goat anti-rabbit IgG | 1:500 | Thermo Fisher Scientific, A-11034, RRID:AB_1885241 | |
| Primers for APOE Genotyping | Forward/Reverse primer (5′–3′) F: GGCACGGCTGTCCAAGGA; R: GCCCCGGCCTGGTACAC | ||
| Markers | Forward Primer | Reverse Primer |
|---|---|---|
| TRA-1-60 | ACAGGAAACACCCTCTGTGC | GAAGGTGGCTTTGACTGCTC |
| NANOG | ACAACTGGCCGAAGAATAGCA | GGTTCCCAGTCGGGTTCAC |
| MAP2 | TC GAGGCAATGACCTTACC | GTGGTAGGCTCTTGGTCTTT |
| VGLUT1 | CACCATGGAGTTCCGCC | CACTCAGCTCCAGCGTCTC |
| NMDAR1 | ATCTACTCGGACAAGAGCATCC | AGCTCTTTCGCCTCCATCAG |
| CTIP2 | CAGAGCAGCAAGCTCACG | GGTGCTGTAGACGCTGAA GG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sagar, R.; Zivko, C.; Xydia, A.; Weisman, D.C.; Lyketsos, C.G.; Mahairaki, V. Generation and Characterization of a Human-Derived and Induced Pluripotent Stem Cell (iPSC) Line from an Alzheimer’s Disease Patient with Neuropsychiatric Symptoms. Biomedicines 2023, 11, 3313. https://doi.org/10.3390/biomedicines11123313
Sagar R, Zivko C, Xydia A, Weisman DC, Lyketsos CG, Mahairaki V. Generation and Characterization of a Human-Derived and Induced Pluripotent Stem Cell (iPSC) Line from an Alzheimer’s Disease Patient with Neuropsychiatric Symptoms. Biomedicines. 2023; 11(12):3313. https://doi.org/10.3390/biomedicines11123313
Chicago/Turabian StyleSagar, Ram, Cristina Zivko, Ariadni Xydia, David C. Weisman, Constantine G. Lyketsos, and Vasiliki Mahairaki. 2023. "Generation and Characterization of a Human-Derived and Induced Pluripotent Stem Cell (iPSC) Line from an Alzheimer’s Disease Patient with Neuropsychiatric Symptoms" Biomedicines 11, no. 12: 3313. https://doi.org/10.3390/biomedicines11123313
APA StyleSagar, R., Zivko, C., Xydia, A., Weisman, D. C., Lyketsos, C. G., & Mahairaki, V. (2023). Generation and Characterization of a Human-Derived and Induced Pluripotent Stem Cell (iPSC) Line from an Alzheimer’s Disease Patient with Neuropsychiatric Symptoms. Biomedicines, 11(12), 3313. https://doi.org/10.3390/biomedicines11123313