
Biomechanical Properties of the Aortic Wall: Changes during Vascular Calcification

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Aorta Harvest
2.2. Ex Vivo Stimulation of Aortic Rings
2.3. Passive Response
2.4. Calcium Content
2.5. Gene Expression
2.6. Statistical Analysis
3. Results
3.1. Biomechanical Properties of Rat Aortic Tissue: Proof-of-Concept
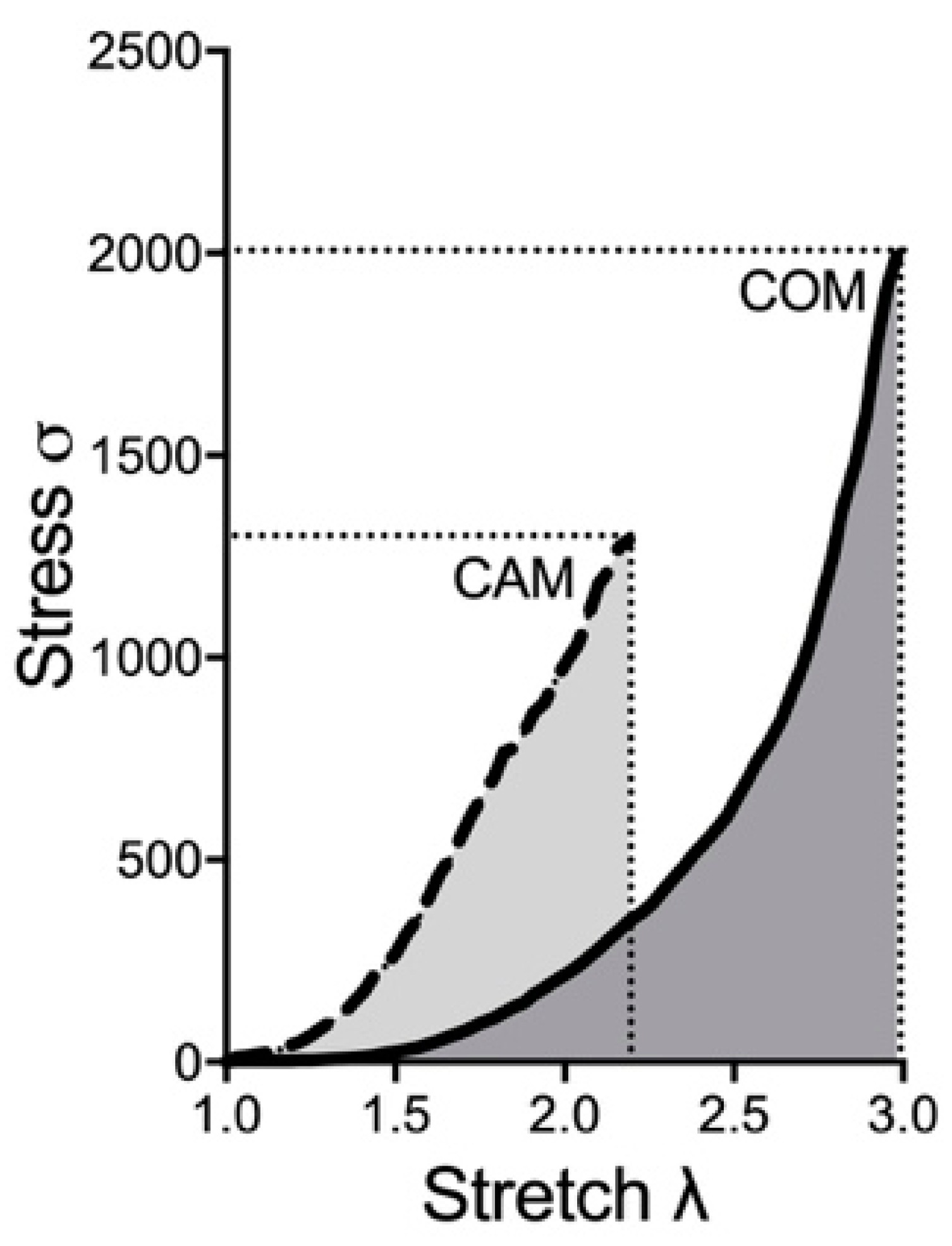
3.2. Biomechanical Properties: COM vs. CAM
3.3. Changes in Vascular Elastin and Collagen
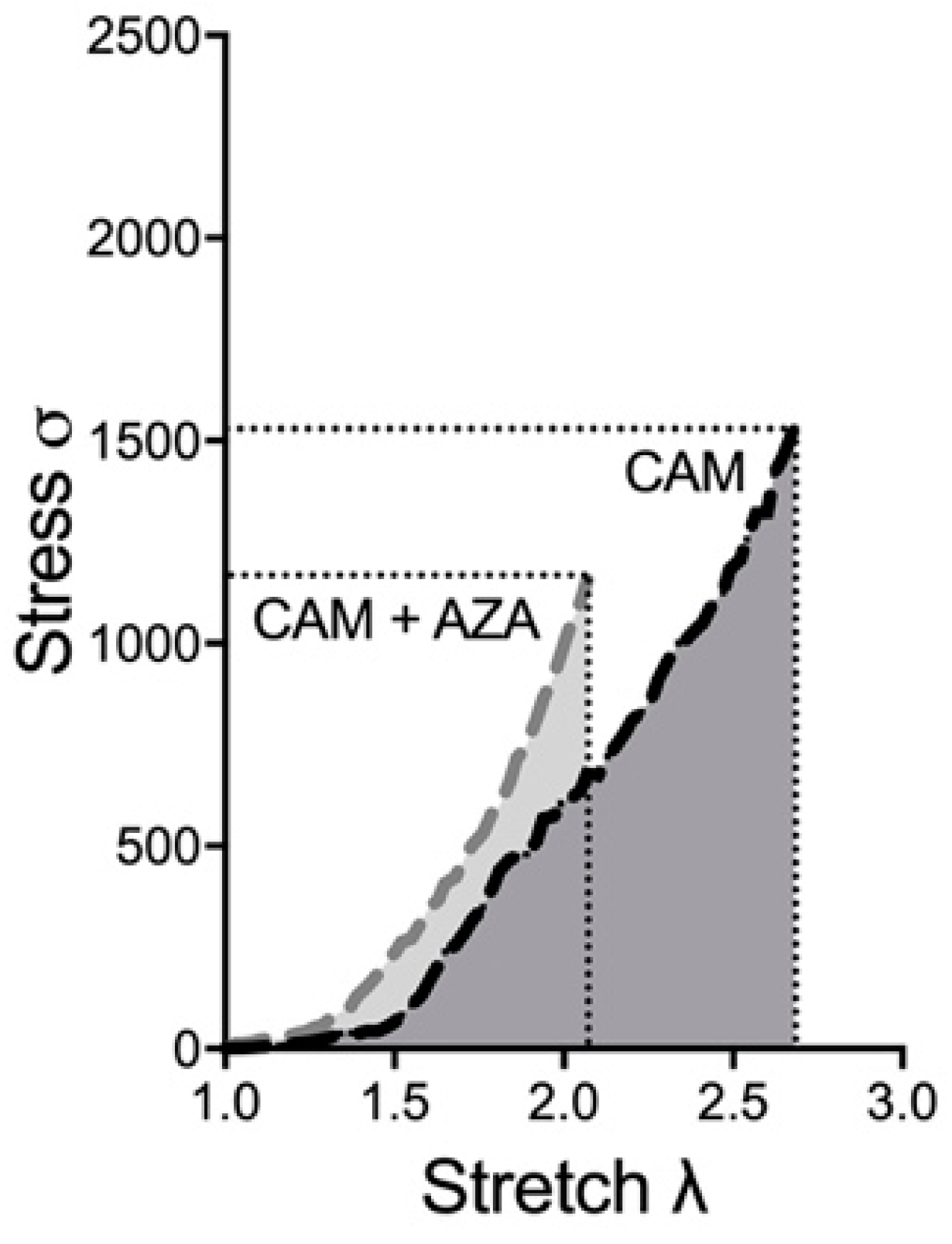
3.4. Method Validation by a Known Inductor of Vascular Calcification: Azathioprine
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vos, T.; Lim, S.S.; Abbafati, C.; Abbas, K.M.; Abbasi, M.; Abbasifard, M.; Abbasi-Kangevari, M.; Abbastabar, H.; Abd-Allah, F.; Abdelalim, A.; et al. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar] [CrossRef] [PubMed]
- Lanzer, P.; Hannan, F.M.; Lanzer, J.D.; Janzen, J.; Raggi, P.; Furniss, D.; Schuchardt, M.; Thakker, R.; Fok, P.W.; Saez-Rodriguez, J.; et al. Medial Arterial Calcification: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2021, 78, 1145–1165. [Google Scholar] [CrossRef]
- Voelkl, J.; Cejka, D.; Alesutan, I. An overview of the mechanisms in vascular calcification during chronic kidney disease. Curr. Opin. Nephrol. Hypertens. 2019, 28, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Thubrikar, M.J. Vascular Mechanics and Pathology; Springer: Berlin/Heidelberg, Germany, 2007; Volume 494. [Google Scholar]
- Henaut, L.; Mary, A.; Chillon, J.M.; Kamel, S.; Massy, Z.A. The Impact of Uremic Toxins on Vascular Smooth Muscle Cell Function. Toxins 2018, 10, 218. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, J.; Gummi, M.R.; Xia, M.; van der Giet, M.; Tolle, M.; Schuchardt, M. Vascular Calcification in Rodent Models-Keeping Track with an Extented Method Assortment. Biology 2021, 10, 459. [Google Scholar] [CrossRef]
- Prufer, J.; Schuchardt, M.; Tolle, M.; Prufer, N.; Hohne, M.; Zidek, W.; van der Giet, M. Harmful effects of the azathioprine metabolite 6-mercaptopurine in vascular cells: Induction of mineralization. PLoS ONE 2014, 9, e101709. [Google Scholar] [CrossRef] [PubMed]
- Schuchardt, M.; Herrmann, J.; Henkel, C.; Babic, M.; van der Giet, M.; Tolle, M. Long-Term Treatment of Azathioprine in Rats Induces Vessel Mineralization. Biomedicines 2021, 9, 327. [Google Scholar] [CrossRef]
- Spiers, A.; Padmanabhan, N. A guide to wire myography. Methods Mol. Med. 2005, 108, 91–104. [Google Scholar] [CrossRef]
- Gequelim, G.C.; da Luz Veronez, D.A.; Lenci Marques, G.; Tabushi, C.H.; Loures Bueno, R.D.R. Thoracic aorta thickness and histological changes with aging: An experimental rat model. J. Geriatr. Cardiol. 2019, 16, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Canas, D.; Herrera, E.A.; Garcia-Herrera, C.; Celentano, D.; Krause, B.J. Fetal Growth Restriction Induces Heterogeneous Effects on Vascular Biomechanical and Functional Properties in Guinea Pigs (Cavia porcellus). Front. Physiol. 2017, 8, 144. [Google Scholar] [CrossRef]
- Ameer, O.Z.; Salman, I.M.; Avolio, A.P.; Phillips, J.K.; Butlin, M. Opposing changes in thoracic and abdominal aortic biomechanical properties in rodent models of vascular calcification and hypertension. Am. J. Physiol. Heart Circ. Physiol. 2014, 307, H143–H151. [Google Scholar] [CrossRef]
- Russell, W.M.S.; Bruch, R.L. The Principles of Human Experimental Technique; Methuen: London, UK; Johns Hopkins University: Baltimore, MD, USA, 1959. [Google Scholar]
- Giudici, A.; Wilkinson, I.B.; Khir, A.W. Review of the Techniques Used for Investigating the Role Elastin and Collagen Play in Arterial Wall Mechanics. IEEE Rev. Biomed. Eng. 2021, 14, 256–269. [Google Scholar] [CrossRef] [PubMed]
- Mazurek, R.; Dave, J.M.; Chandran, R.R.; Misra, A.; Sheikh, A.Q.; Greif, D.M. Vascular Cells in Blood Vessel Wall Development and Disease. Adv. Pharmacol. 2017, 78, 323–350. [Google Scholar] [CrossRef] [PubMed]
- Wagenseil, J.E.; Mecham, R.P. Vascular extracellular matrix and arterial mechanics. Physiol. Rev. 2009, 89, 957–989. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.T.; Kamm, R.D. Vascular mechanics for the cardiologist. J. Am. Coll. Cardiol. 1994, 23, 1289–1295. [Google Scholar] [CrossRef] [PubMed]
- Gosling, R.G.; Budge, M.M. Terminology for describing the elastic behavior of arteries. Hypertension 2003, 41, 1180–1182. [Google Scholar] [CrossRef] [PubMed]
- Camasao, D.B.; Mantovani, D. The mechanical characterization of blood vessels and their substitutes in the continuous quest for physiological-relevant performances. A critical review. Mater. Today Bio 2021, 10, 100106. [Google Scholar] [CrossRef]
- Mahutga, R.R.; Schoephoerster, C.T.; Barocas, V.H. The Ring-Pull Assay for Mechanical Properties of Fibrous Soft Tissues—An Analysis of the Uniaxial Approximation and a Correction for Nonlinear Thick-Walled Tissues. Exp. Mech. 2021, 61, 53–66. [Google Scholar] [CrossRef]
- Holmar, J.; Noels, H.; Bohm, M.; Bhargava, S.; Jankowski, J.; Orth-Alampour, S. Development, establishment and validation of in vitro and ex vivo assays of vascular calcification. Biochem. Biophys. Res. Commun. 2020, 530, 462–470. [Google Scholar] [CrossRef]
- Herrmann, J.; Xia, M.; Gummi, M.R.; Greco, A.; Schacke, A.; van der Giet, M.; Tolle, M.; Schuchardt, M. Stressor-Induced “Inflammaging” of Vascular Smooth Muscle Cells via Nlrp3-Mediated Pro-inflammatory Auto-Loop. Front. Cardiovasc Med. 2021, 8, 752305. [Google Scholar] [CrossRef]
- Vrhovski, B.; Weiss, A.S. Biochemistry of tropoelastin. Eur. J. Biochem. 1998, 258, 1–18. [Google Scholar] [CrossRef]
- Wang, K.; Meng, X.; Guo, Z. Elastin Structure, Synthesis, Regulatory Mechanism and Relationship with Cardiovascular Diseases. Front. Cell Dev. Biol. 2021, 9, 596702. [Google Scholar] [CrossRef] [PubMed]
- Aikawa, E.; Aikawa, M.; Libby, P.; Figueiredo, J.L.; Rusanescu, G.; Iwamoto, Y.; Fukuda, D.; Kohler, R.H.; Shi, G.P.; Jaffer, F.A.; et al. Arterial and aortic valve calcification abolished by elastolytic cathepsin S deficiency in chronic renal disease. Circulation 2009, 119, 1785–1794. [Google Scholar] [CrossRef]
- Burton, A.C. Relation of structure to function of the tissues of the wall of blood vessels. Physiol. Rev. 1954, 34, 619–642. [Google Scholar] [CrossRef] [PubMed]
- Cocciolone, A.J.; Hawes, J.Z.; Staiculescu, M.C.; Johnson, E.O.; Murshed, M.; Wagenseil, J.E. Elastin, arterial mechanics, and cardiovascular disease. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H189–H205. [Google Scholar] [CrossRef] [PubMed]
- Gabriela Espinosa, M.; Catalin Staiculescu, M.; Kim, J.; Marin, E.; Wagenseil, J.E. Elastic Fibers and Large Artery Mechanics in Animal Models of Development and Disease. J. Biomech. Eng. 2018, 140, 0208031–02080313. [Google Scholar] [CrossRef]
- Tsamis, A.; Krawiec, J.T.; Vorp, D.A. Elastin and collagen fibre microstructure of the human aorta in ageing and disease: A review. J. R. Soc. Interface 2013, 10, 20121004. [Google Scholar] [CrossRef]
- Molinari-Tosatti, M.P.; Galzigna, L.; Moret, V.; Gotte, L. Some features of the binding of calcium ions to elastin. Calcif. Tissue Res. 1968, 6 (Suppl. 88), 329–334. [Google Scholar] [CrossRef]
- Yu, S.Y.; Blumenthal, H.T. The calcification of elastic fibers. I. Biochemical studies. J. Gerontol. 1963, 18, 119–126. [Google Scholar] [CrossRef]
- Schiffmann, E.; Lavender, D.R.; Miller, E.J.; Corcoran, B.A. Amino acids at the nucleating site in mineralizing elastic tissue. Calcif. Tissue Res. 1969, 3, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Contri, M.B.; Boraldi, F.; Taparelli, F.; De Paepe, A.; Ronchetti, I.P. Matrix proteins with high affinity for calcium ions are associated with mineralization within the elastic fibers of pseudoxanthoma elasticum dermis. Am. J. Pathol. 1996, 148, 569–577. [Google Scholar] [PubMed]
- Gheduzzi, D.; Boraldi, F.; Annovi, G.; DeVincenzi, C.P.; Schurgers, L.J.; Vermeer, C.; Quaglino, D.; Ronchetti, I.P. Matrix Gla protein is involved in elastic fiber calcification in the dermis of pseudoxanthoma elasticum patients. Lab. Investig. 2007, 87, 998–1008. [Google Scholar] [CrossRef] [PubMed]
- Fornieri, C.; Baccarani-Contri, M.; Quaglino, D., Jr.; Pasquali-Ronchetti, I. Lysyl oxidase activity and elastin/glycosaminoglycan interactions in growing chick and rat aortas. J. Cell Biol. 1987, 105, 1463–1469. [Google Scholar] [CrossRef]
- Zohora, F.-T.; Nosoudi, N.; Karamched, S.R.; Vyavahare, N. The Role of Elastin Degradation in Vascular Calcification: Possibilities to Repair Elastin and Reverse Calcification. In Cardiovascular Calcification and Bone Mineralization; Springer: Berlin/Heidelberg, Germany, 2020; pp. 441–480. [Google Scholar]
- Pereira, L.; Lee, S.Y.; Gayraud, B.; Andrikopoulos, K.; Shapiro, S.D.; Bunton, T.; Biery, N.J.; Dietz, H.C.; Sakai, L.Y.; Ramirez, F. Pathogenetic sequence for aneurysm revealed in mice underexpressing fibrillin-1. Proc. Natl. Acad. Sci. USA 1999, 96, 3819–3823. [Google Scholar] [CrossRef] [PubMed]
- Basalyga, D.M.; Simionescu, D.T.; Xiong, W.; Baxter, B.T.; Starcher, B.C.; Vyavahare, N.R. Elastin degradation and calcification in an abdominal aorta injury model: Role of matrix metalloproteinases. Circulation 2004, 110, 3480–3487. [Google Scholar] [CrossRef]
- Zhao, Y.; Sun, Z.; Li, L.; Yuan, W.; Wang, Z. Role of Collagen in Vascular Calcification. J. Cardiovasc. Pharmacol. 2022, 80, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Watson, K.E.; Parhami, F.; Shin, V.; Demer, L.L. Fibronectin and collagen I matrixes promote calcification of vascular cells in vitro, whereas collagen IV matrix is inhibitory. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 1964–1971. [Google Scholar] [CrossRef]
- Ratzinger, S.; Grassel, S.; Dowejko, A.; Reichert, T.E.; Bauer, R.J. Induction of type XVI collagen expression facilitates proliferation of oral cancer cells. Matrix Biol. 2011, 30, 118–125. [Google Scholar] [CrossRef]
- Luo, G.; Ducy, P.; McKee, M.D.; Pinero, G.J.; Loyer, E.; Behringer, R.R.; Karsenty, G. Spontaneous calcification of arteries and cartilage in mice lacking matrix GLA protein. Nature 1997, 386, 78–81. [Google Scholar] [CrossRef]
- Sorushanova, A.; Delgado, L.M.; Wu, Z.; Shologu, N.; Kshirsagar, A.; Raghunath, R.; Mullen, A.M.; Bayon, Y.; Pandit, A.; Raghunath, M.; et al. The Collagen Suprafamily: From Biosynthesis to Advanced Biomaterial Development. Adv. Mater. 2019, 31, e1801651. [Google Scholar] [CrossRef]
- Shekhonin, B.V.; Domogatsky, S.P.; Muzykantov, V.R.; Idelson, G.L.; Rukosuev, V.S. Distribution of type I, III, IV and V collagen in normal and atherosclerotic human arterial wall: Immunomorphological characteristics. Coll. Relat. Res. 1985, 5, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Byers, P.H. Vascular Ehlers-Danlos Syndrome. In GeneReviews (Internet); University of Washington, Seattle: Seattle, WA, USA, 1999. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Basal Mean | Basal SD | CAM Mean | CAM SD | T | p | d |
|---|---|---|---|---|---|---|---|
| Elow | 12.55 | 4.55 | 48.68 | 12.29 | 4.77 | 0.01 | 3.90 |
| Ehigh | 3944.00 | 89.72 | 1439.27 | 90.92 | 33.96 | <0.01 | 27.73 |
| AUC | 734.11 | 12.18 | 770.12 | 210.77 | 0.30 | 0.78 | 0.24 |
| Breaking point (σ) | 1884.74 | 41.54 | 1403.03 | 172.16 | 4.71 | 0.01 | 3.85 |
| Breaking point (λ) | 3.02 | 0.05 | 2.51 | 0.28 | 3.15 | 0.04 | 2.57 |
| Calcium content (µg/mg) | 3.13 | 3.50 | 249.15 | 152.96 | 2.79 | 0.05 | 2.27 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Gummi, M.R.; Greco, A.; Babic, M.; Herrmann, J.; Kandil, F.I.; van der Giet, M.; Tölle, M.; Schuchardt, M. Biomechanical Properties of the Aortic Wall: Changes during Vascular Calcification. Biomedicines 2023, 11, 211. https://doi.org/10.3390/biomedicines11010211
Zhou J, Gummi MR, Greco A, Babic M, Herrmann J, Kandil FI, van der Giet M, Tölle M, Schuchardt M. Biomechanical Properties of the Aortic Wall: Changes during Vascular Calcification. Biomedicines. 2023; 11(1):211. https://doi.org/10.3390/biomedicines11010211
Chicago/Turabian StyleZhou, Jinwen, Manasa Reddy Gummi, Anna Greco, Milen Babic, Jaqueline Herrmann, Farid I. Kandil, Markus van der Giet, Markus Tölle, and Mirjam Schuchardt. 2023. "Biomechanical Properties of the Aortic Wall: Changes during Vascular Calcification" Biomedicines 11, no. 1: 211. https://doi.org/10.3390/biomedicines11010211
APA StyleZhou, J., Gummi, M. R., Greco, A., Babic, M., Herrmann, J., Kandil, F. I., van der Giet, M., Tölle, M., & Schuchardt, M. (2023). Biomechanical Properties of the Aortic Wall: Changes during Vascular Calcification. Biomedicines, 11(1), 211. https://doi.org/10.3390/biomedicines11010211