
The Generation of Dual-Targeting Fusion Protein PD-L1/CD47 for the Inhibition of Triple-Negative Breast Cancer


Abstract
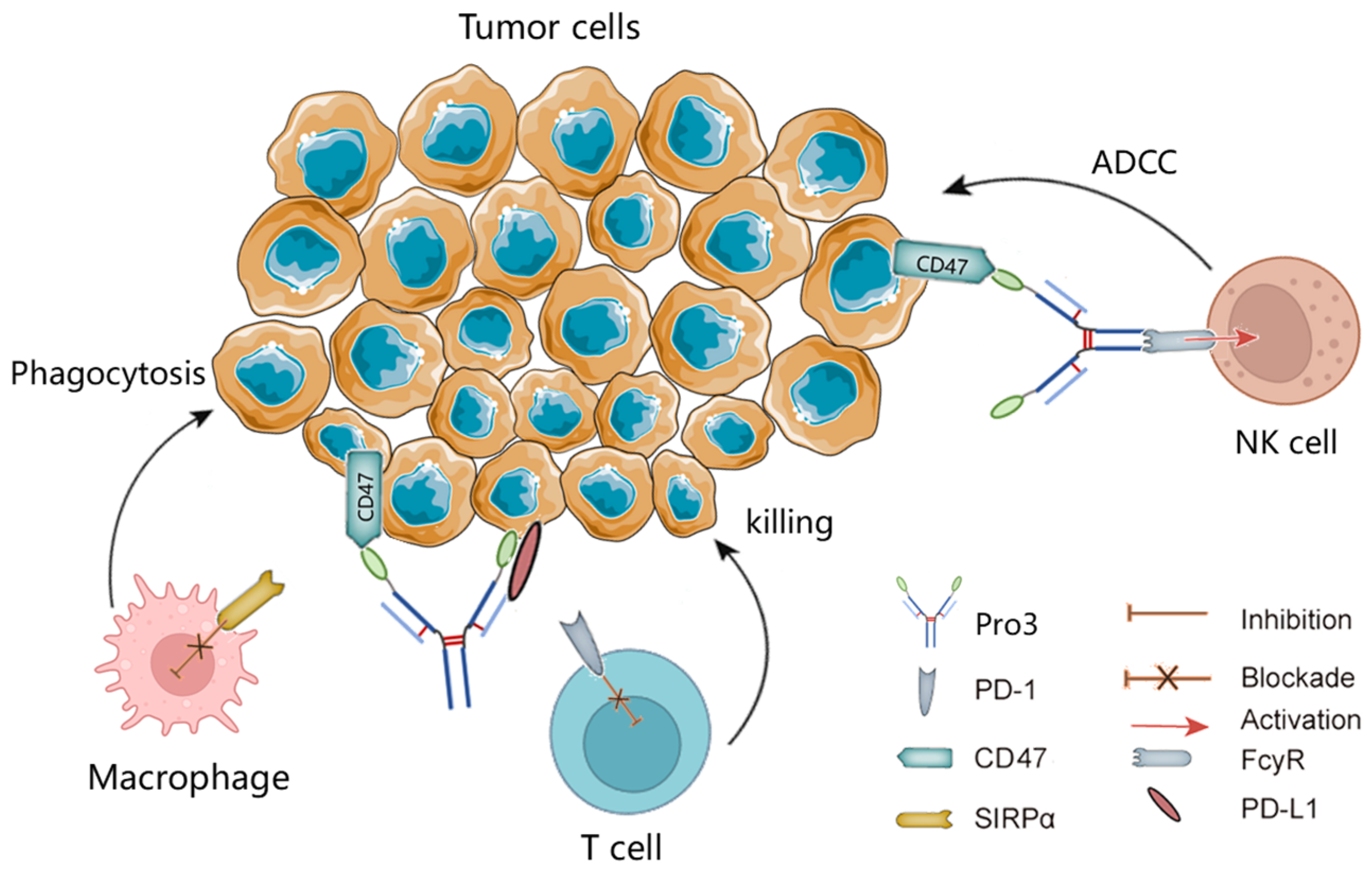
1. Introduction
2. Materials and Methods
2.1. Strains, Cell Lines, and Other Reagents
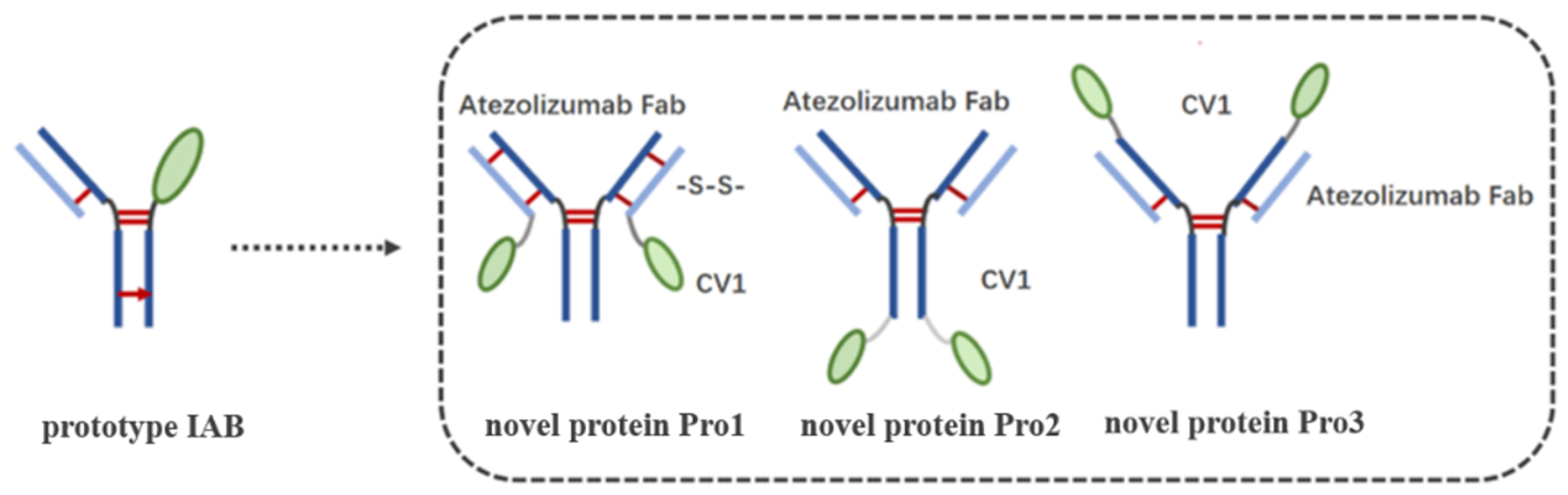
2.2. Molecular Design and General Construction of Dual-Targeting Proteins
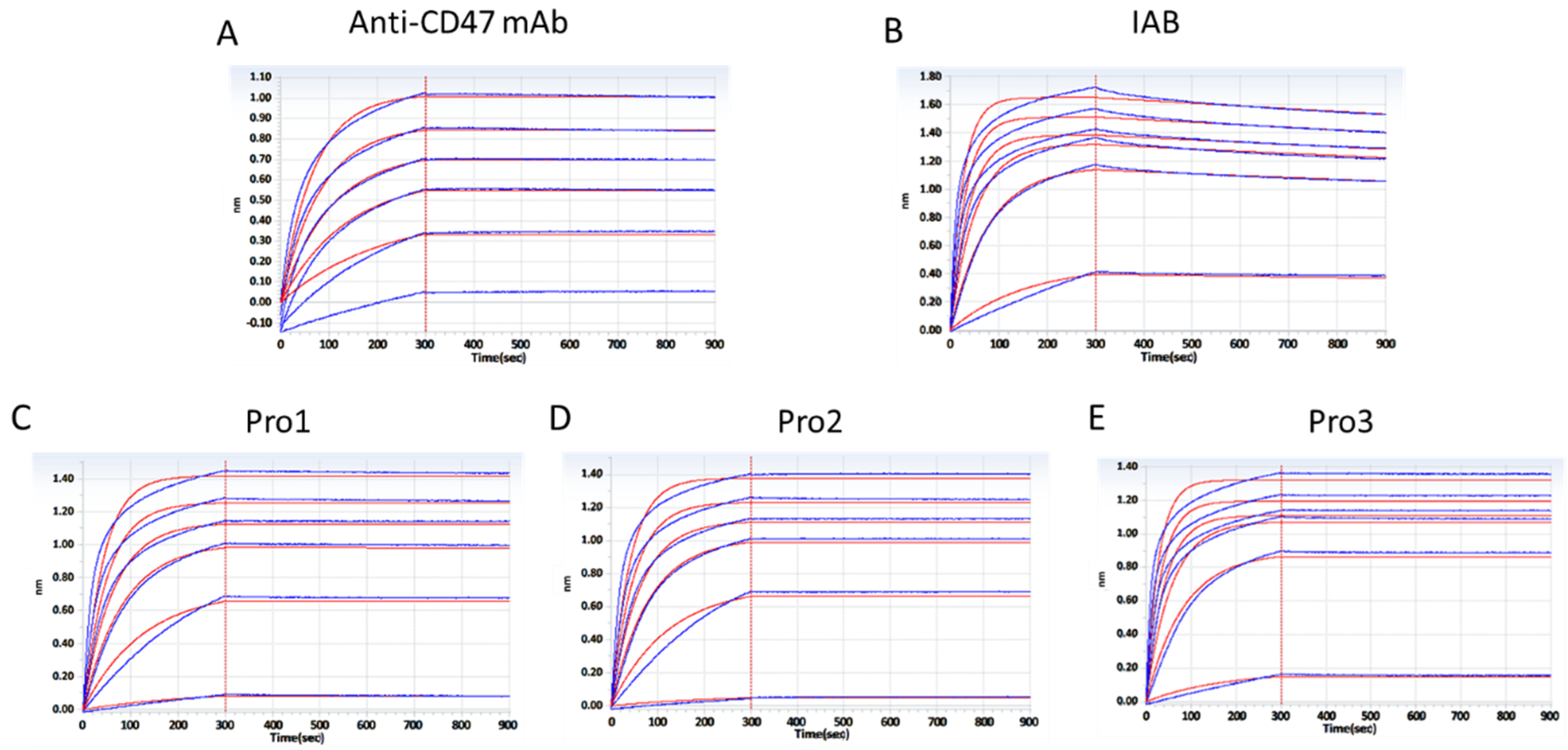
2.3. Binding Analysis In Vitro
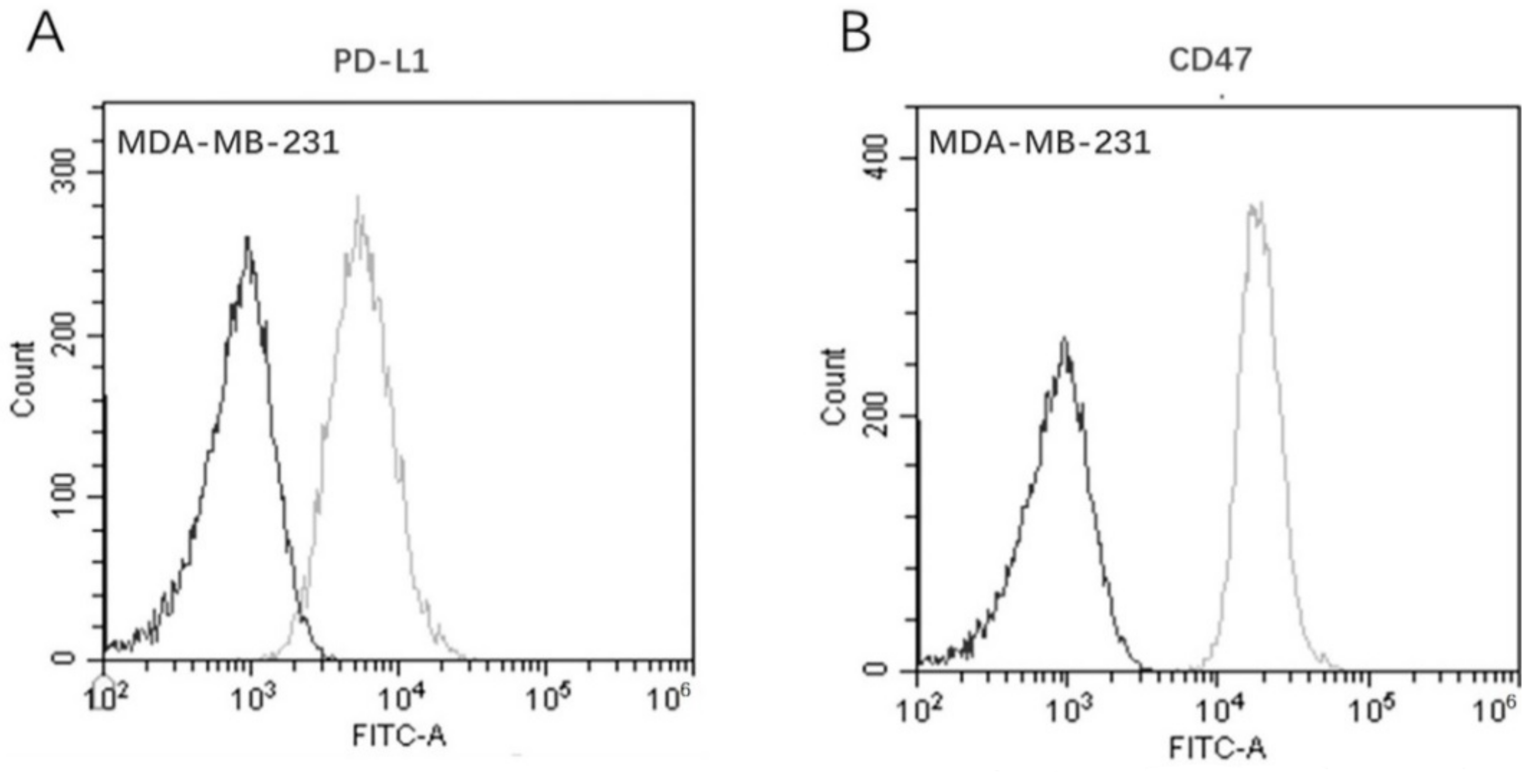
2.4. PD-L1 and CD47 Co-Expression on MDA-MB-231 Cells
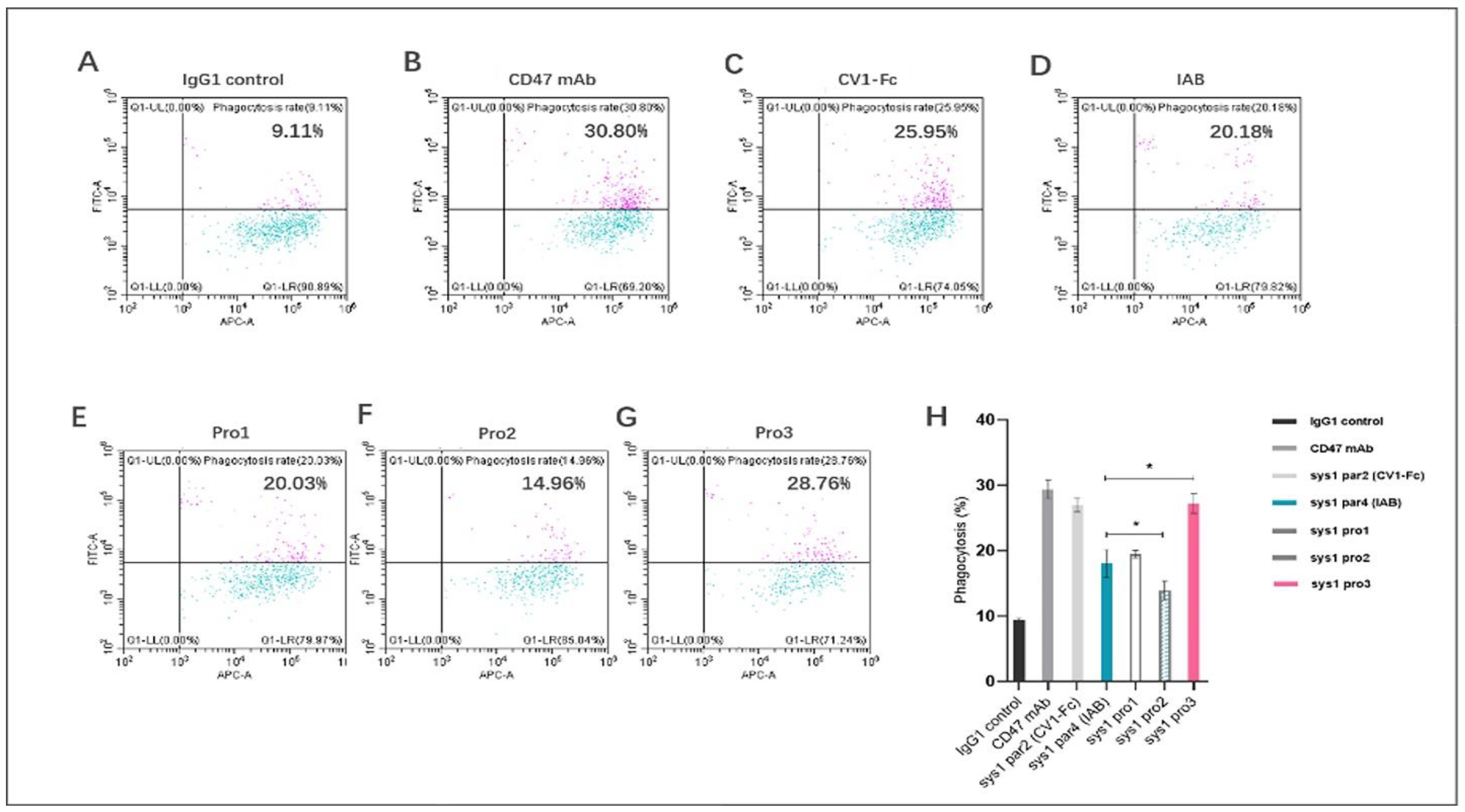
2.5. In Vitro Human Macrophage Activation
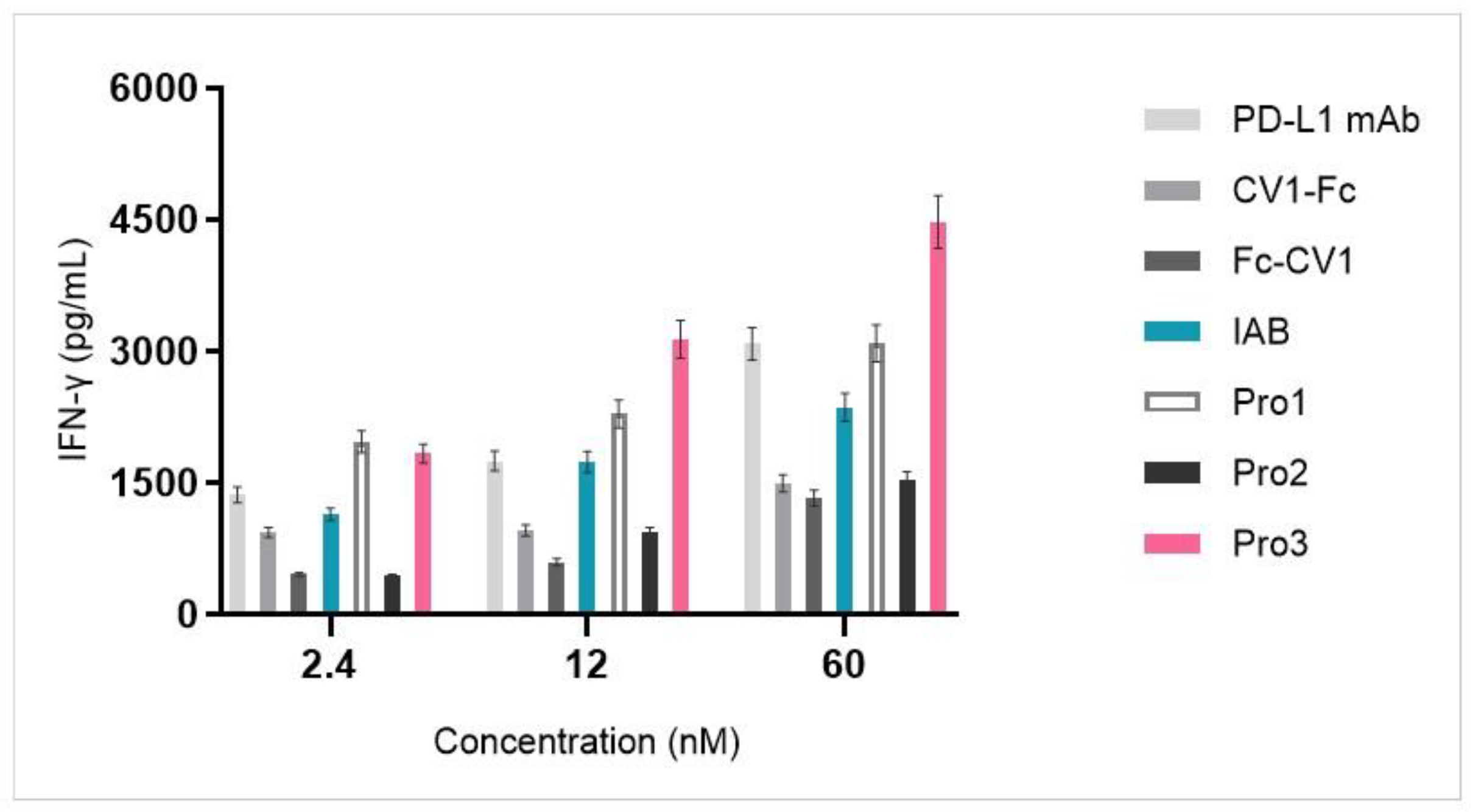
2.6. Mixed Lymphocyte Reaction (MLR)
2.7. Antibody-Dependent Cell-Mediated Cytotoxicity (ADCC)
3. Results
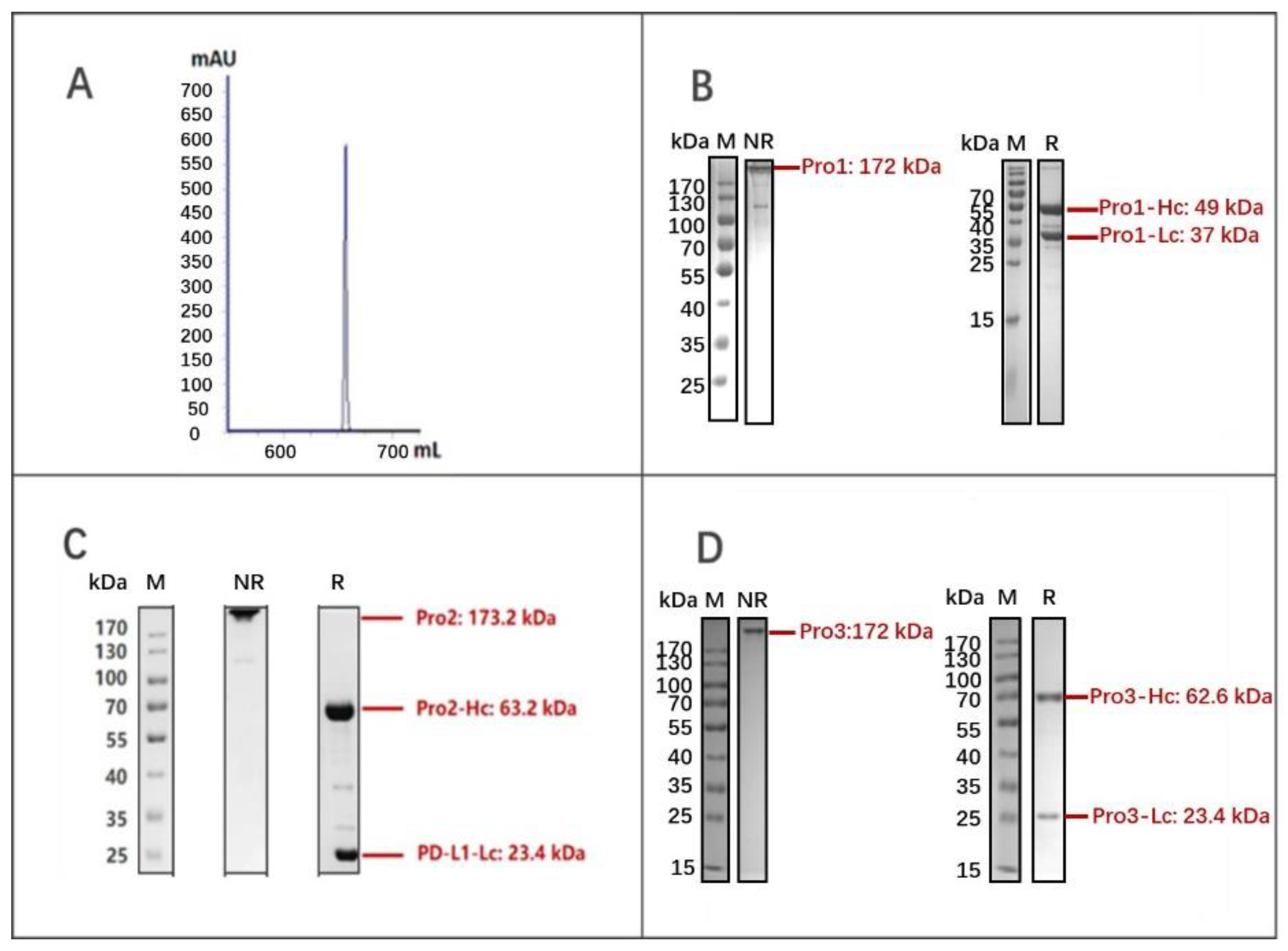
3.1. Molecular Construction and Preparation of the Dual-Targeting Proteins
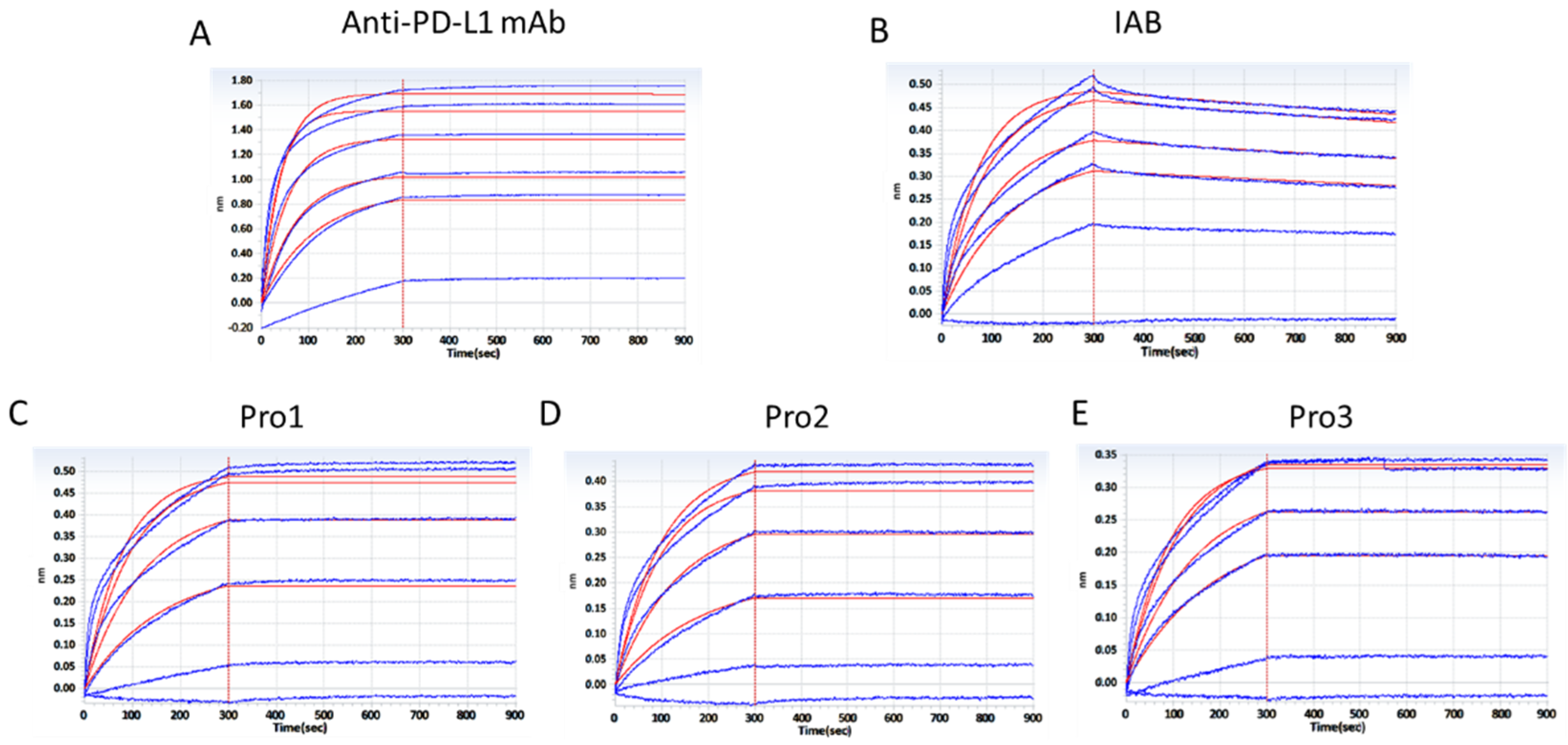
3.2. Binding Characterization In Vitro
3.3. Co–Expression of PD–L1 and CD47 on MDA–MB–231
3.4. In Vitro Human Macrophage Activation
3.5. Dual-Targeting Proteins Exhibited Superior Activity in a T Cell Activation Assay
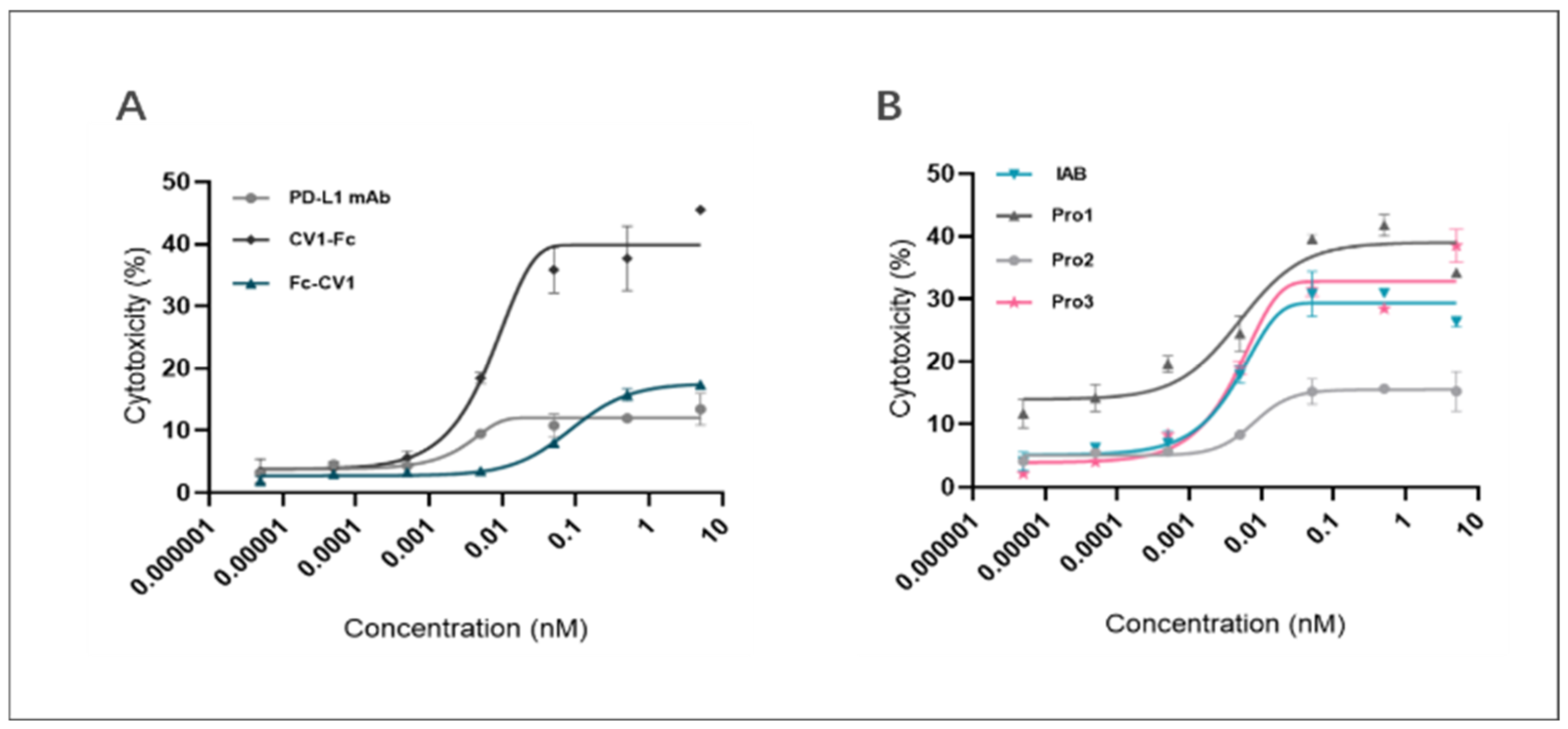
3.6. Dual-Targeting Proteins Retained the ADCC-Mediated Capacity of IAB
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. The DNA Sequence of Anti-PD-L1 mAb (Atezolizumab)
- Variable Domain of the Heavy Chain
- Variable Domain of the Light Chain
Appendix A.2. The DNA Sequence of Anti-CD47 mAb (B6H12)
- Variable Domain of the Heavy Chain
- Variable Domain of the Light Chain
Appendix A.3. The Nucleic Acid Sequence of CV1
References
- Sorlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef] [PubMed]
- Abkevich, V.; Timms, K.M.; Hennessy, B.T.; Potter, J.; Carey, M.S.; Meyer, L.A.; Smith-McCune, K.; Broaddus, R.; Lu, K.H.; Chen, J.; et al. Patterns of genomic loss of heterozygosity predict homologous recombination repair defects in epithelial ovarian cancer. Br. J. Cancer 2012, 107, 1776–1782. [Google Scholar] [CrossRef]
- Mittendorf, E.A.; Philips, A.V.; Meric-Bernstam, F.; Qiao, N.; Wu, Y.; Harrington, S.; Su, X.P.; Wang, Y.; Gonzalez-Angulo, A.M.; Akcakanat, A.; et al. PD-L1 Expression in Triple-Negative Breast Cancer. Cancer Immunol. Res. 2014, 2, 361–370. [Google Scholar] [CrossRef]
- Yu, C.; Zhen, L.; Li, H.; Zhang, Y.; Zhang, Q. The correlations between expression of tumor associated macrophages and the prognostic significances in triple negative breast cancer. Pract. Oncol. J. 2017, 31, 1–6. [Google Scholar]
- Edris, B.; Weiskopf, K.; Volkmer, A.K.; Volkmer, J.P.; Willingham, S.B.; Contreras-Trujillo, H.; Liu, J.; Majeti, R.; West, R.B.; Fletcher, J.A.; et al. Antibody therapy targeting the CD47 protein is effective in a model of aggressive metastatic leiomyosarcoma. Proc. Natl. Acad. Sci. USA 2012, 109, 6656–6661. [Google Scholar] [CrossRef]
- Korman, A.J.; Peggs, K.S.; Allison, J.P. Checkpoint blockade in cancer immunotherapy. In Advances in Immunology: Cancer Immunotherapy; Allison, J.P., Franoff, G., Eds.; Elsevier Academic Press Inc.: San Diego, CA, USA, 2006; Volume 90, pp. 297–339. [Google Scholar]
- Freeman, G.J.; Long, A.J.; Iwai, Y.; Bourque, K.; Chernova, T.; Nishimura, H.; Fitz, L.J.; Malenkovich, N.; Okazaki, T.; Byrne, M.C.; et al. Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation. J. Exp. Med. 2000, 192, 1027–1034. [Google Scholar] [CrossRef]
- Brown, J.A.; Dorfman, D.M.; Ma, F.R.; Sullivan, E.L.; Munoz, O.; Wood, C.R.; Greenfield, E.A.; Freeman, G.J. Blockade of programmed death-1 ligands on dendritic cells enhances T cell activation and cytokine production. J. Immunol. 2003, 170, 1257–1266. [Google Scholar] [CrossRef]
- Liu, B.; Guo, H.; Xu, J.; Qin, T.; Guo, Q.; Gu, N.; Zhang, D.; Qian, W.; Dai, J.; Hou, S.; et al. Elimination of tumor by CD47/PD-L1 dual-targeting fusion protein that engages innate and adaptive immune responses. MAbs 2018, 10, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Boyerinas, B.; Jochems, C.; Fantini, M.; Heery, C.R.; Gulley, J.L.; Tsang, K.Y.; Schlom, J. Antibody-Dependent Cellular Cytotoxicity Activity of a Novel Anti-PD-L1 Antibody Avelumab (MSB0010718C) on Human Tumor Cells. Cancer Immunol. Res. 2015, 3, 1148–1157. [Google Scholar] [CrossRef]
- Tseng, D.; Volkmer, J.P.; Willingham, S.B.; Contreras-Trujillo, H.; Fathman, J.W.; Fernhoff, N.B.; Seita, J.; Inlay, M.A.; Weiskopf, K.; Miyanishi, M.; et al. Anti-CD47 antibody-mediated phagocytosis of cancer by macrophages primes an effective antitumor T-cell response. Proc. Natl. Acad. Sci. USA 2013, 110, 11103–11108. [Google Scholar] [CrossRef]
- Orcutt, K.D.; Ackerman, M.E.; Cieslewicz, M.; Quiroz, E.; Slusarczyk, A.L.; Frangioni, J.V.; Wittrup, K.D. A modular IgG-scFv bispecific antibody topology. Protein Eng. Des. Sel. 2010, 23, 221–228. [Google Scholar] [CrossRef]
- Reiter, Y.; Brinkmann, U.; Lee, B.; Pastan, I. Engineering antibody Fv fragments for cancer detection and therapy: Bisulfide-stabilized Fv fragments. Nat. Biotechnol. 1996, 14, 7. [Google Scholar] [CrossRef]
- McDermott, D.F.; Sosman, J.A.; Sznol, M.; Massard, C.; Gordon, M.S.; Hamid, O.; Powderly, J.D.; Infante, J.R.; Fasso, M.; Wang, Y.V.; et al. Atezolizumab, an Anti-Programmed Death-Ligand 1 Antibody, in Metastatic Renal Cell Carcinoma: Long-Term Safety, Clinical Activity, and Immune Correlates From a Phase Ia Study. J. Clin. Oncol. 2016, 34, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Weiskopf, K.; Ring, A.M.; Ho, C.C.; Volkmer, J.P.; Levin, A.M.; Volkmer, A.K.; Ozkan, E.; Fernhoff, N.B.; van de Rijn, M.; Weissman, I.L.; et al. Engineered SIRPα variants as immunotherapeutic adjuvants to anticancer antibodies. Science 2013, 341, 88–91. [Google Scholar] [CrossRef]
- Zhu, J.; Kahn, C.R. Analysis of a peptide hormone-receptor interaction in the yeast two-hybrid system. Proc. Natl. Acad. Sci. USA 1997, 94, 13063–13068. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Chen, J.S.; Ding, K.; Zong, H.F.; Xie, Y.Q.; Jiang, H.; Zhang, B.H.; Lu, H.L.; Yin, W.H.; Gilly, J.; et al. Efficient generation of bispecific IgG antibodies by split intein mediated protein trans-splicing system. Sci. Rep. 2017, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Zong, H.F.; Zhou, Y.X.; Pan, Z.D.; Chen, J.; Ding, K.; Xie, Y.Q.; Jiang, H.; Zhang, B.H.; Lu, H.L.; et al. Naturally split intein Npu DnaE mediated rapid generation of bispecific IgG antibodies. Methods 2019, 154, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Zong, H.F.; Han, L.; Chen, J.; Pan, Z.D.; Wang, L.; Sun, R.; Ding, K.; Xie, Y.Q.; Jiang, H.; Lu, H.L.; et al. Kinetics study of the natural split Npu DnaE intein in the generation of bispecific IgG antibodies. Appl. Microbiol. Biotechnol. 2022, 106, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Chen, J.; Xiao, X.; Xie, Y.; Jiang, H.; Zhang, B.; Lu, H.; Yuan, Y.; Han, L.; Zhou, Y.; et al. Characterization of a novel bispecific antibody targeting tissue factor-positive tumors with T cell engagement. Acta Pharm. Sin. B 2022, 12, 1928–1942. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Pan, Z.D.; Han, L.; Zhou, Y.X.; Zong, H.F.; Wang, L.; Sun, R.; Jiang, H.; Xie, Y.Q.; Yuan, Y.S.; et al. A Novel Bispecific Antibody Targeting CD3 and Lewis Y with Potent Therapeutic Efficacy against Gastric Cancer. Biomedicines 2021, 9, 17. [Google Scholar] [CrossRef]
- Ding, K.; Han, L.; Zong, H.F.; Chen, J.S.; Zhang, B.H.; Zhu, J.W. Production process reproducibility and product quality consistency of transient gene expression in HEK293 cells with anti-PD1 antibody as the model protein. Appl. Microbiol. Biotechnol. 2017, 101, 1889–1898. [Google Scholar] [CrossRef]
- Wallner, J.; Lhota, G.; Jeschek, D.; Mader, A.; Vorauer-Uhl, K. Application of Bio-Layer Interferometry for the analysis of protein/liposome interactions. J. Pharm. Biomed. Anal. 2013, 72, 150–154. [Google Scholar] [CrossRef]
- Katsamba, P.S.; Navratilova, I.; Calderon-Cacia, M.; Fan, L.; Thornton, K.; Zhu, M.D.; Vanden Bos, T.; Forte, C.; Friend, D.; Laird-Offringa, I.; et al. Kinetic analysis of a high-affinity antibody/antigen interaction performed by multiple Biacore users. Anal. Biochem. 2006, 352, 208–221. [Google Scholar] [CrossRef]
- Sakakura, K.; Takahashi, H.; Kaira, K.; Toyoda, M.; Murata, T.; Ohnishi, H.; Oyama, T.; Chikamatsu, K. Relationship between tumor-associated macrophage subsets and CD47 expression in squamous cell carcinoma of the head and neck in the tumor microenvironment. Lab. Investig. 2016, 96, 994–1003. [Google Scholar] [CrossRef]
- Broussas, M.; Broyer, L.; Goetsch, L. Evaluation of antibody-dependent cell cytotoxicity using lactate dehydrogenase (LDH) measurement. Methods Mol. Biol. 2013, 988, 305–317. [Google Scholar] [CrossRef]
- Baldassare, J.J.; Henderson, P.A.; Fisher, G.J. Isolation and characterization of one soluble and 2 membrane-associated FORMS of phosphoinositide-specific phospholipase-C from human-platelets. Biochemistry 1989, 28, 6010–6016. [Google Scholar] [CrossRef]
- DiNome, M.L.; Orozco, J.I.J.; Matsuba, C.; Manughian-Peter, A.O.; Ensenyat-Mendez, M.; Chang, S.C.; Jalas, J.R.; Salomon, M.P.; Marzese, D.M. Clinicopathological Features of Triple-Negative Breast Cancer Epigenetic Subtypes. Ann. Surg. Oncol. 2019, 26, 3344–3353. [Google Scholar] [CrossRef]
- Stevenson, G.T. Three major uncertainties in the antibody therapy of cancer. Haematologica 2014, 99, 1538–1546. [Google Scholar] [CrossRef]
- Hamilton, G.; Rath, B. Avelumab: Combining immune checkpoint inhibition and antibody-dependent cytotoxicity. Expert Opin. Biol. Ther. 2017, 17, 515–523. [Google Scholar] [CrossRef]
- Liu, Y.; Tong, Q.; Zhou, Y.; Lee, H.-W.; Yang, J.J.; Bühring, H.-J.; Chen, Y.-T.; Ha, B.; Chen, C.X.J.; Yang, Y.; et al. Functional Elements on SIRPα IgV Domain Mediate Cell Surface Binding to CD47. J. Mol. Biol. 2007, 365, 680–693. [Google Scholar] [CrossRef][Green Version]
- Zhao, Y.R. Construction and Characterization a New Fusion Protein Pro-SIRPα; The Second Military Medical University: Shanghai, China, 2016. [Google Scholar]
- Wang, C.Y.; Thudium, K.B.; Han, M.H.; Wang, X.T.; Huang, H.C.; Feingersh, D.; Garcia, C.; Wu, Y.; Kuhne, M.; Srinivasan, M.; et al. In Vitro Characterization of the Anti-PD-1 Antibody Nivolumab, BMS-936558, and In Vivo Toxicology in Non-Human Primates. Cancer Immunol. Res. 2014, 2, 846–856. [Google Scholar] [CrossRef] [PubMed]
- Soto-Pantoja, D.R.; Terabe, M.; Ghosh, A.; Ridnour, L.A.; DeGraff, W.G.; Wink, D.A.; Berzofsky, J.A.; Roberts, D.D. CD47 in the Tumor Microenvironment Limits Cooperation between Antitumor T-cell Immunity and Radiotherapy. Cancer Res. 2014, 74, 6771–6783. [Google Scholar] [CrossRef] [PubMed]
- Lei, C. Effect of Self-Expression of SIRPα-Fc Fusion Protein on T Cell Function; Zhejiang Sci-Tech University: Hangzhou, China, 2015. [Google Scholar]
- Ying, T.; Ju, T.W.; Wang, Y.; Prabakaran, P.; Dimitrov, D.S. Interactions of IgG1 CH2 and CH3 Domains with FcRn. Front. Immunol. 2014, 5, 146. [Google Scholar] [CrossRef] [PubMed]
- McCall, A.M.; Shahied, L.; Amoroso, A.R.; Horak, E.M.; Simmons, H.H.; Nielson, U.; Adams, G.P.; Schier, R.; Marks, J.D.; Weiner, L.M. Increasing the affinity for tumor antigen enhances bispecific antibody cytotoxicity. J. Immunol. 2001, 166, 6112–6117. [Google Scholar] [CrossRef]
- Brahmer, J.R.; Tykodi, S.S.; Chow, L.Q.M.; Hwu, W.J.; Topalian, S.L.; Hwu, P.; Drake, C.G.; Camacho, L.H.; Kauh, J.; Odunsi, K.; et al. Safety and Activity of Anti-PD-L1 Antibody in Patients with Advanced Cancer. N. Engl. J. Med. 2012, 366, 2455–2465. [Google Scholar] [CrossRef]
- Hugo, W.; Zaretsky, J.M.; Sun, L.; Song, C.; Moreno, B.H.; Hu-Lieskovan, S.; Berent-Maoz, B.; Pang, J.; Chmielowski, B.; Cherry, G.; et al. Genomic and Transcriptomic Features of Response to Anti-PD-1 Therapy in Metastatic Melanoma. Cell 2016, 165, 35–44. [Google Scholar] [CrossRef]
- Sockolosky, J.T.; Dougan, M.; Ingram, J.R.; Ho, C.C.M.; Kauke, M.J.; Almo, S.C.; Ploegh, H.L.; Garcia, K.C. Durable antitumor responses to CD47 blockade require adaptive immune stimulation. Proc. Natl. Acad. Sci. USA 2016, 113, E2646–E2654. [Google Scholar] [CrossRef]
- Kuo, T.C.; Chen, A.; Harrabi, O.; Sockolosky, J.T.; Zhang, A.; Sangalang, E.; Doyle, L.V.; Kauder, S.E.; Fontaine, D.; Bollini, S.; et al. Targeting the myeloid checkpoint receptor SIRPα potentiates innate and adaptive immune responses to promote anti-tumor activity. J. Hematol. Oncol. 2020, 13, 160. [Google Scholar] [CrossRef]
- Lakhani, N.J.; Chow, L.Q.M.; Gainor, J.F.; LoRusso, P.; Lee, K.-W.; Chung, H.C.; Lee, J.; Bang, Y.-J.; Hodi, F.S.; Kim, W.S.; et al. Evorpacept alone and in combination with pembrolizumab or trastuzumab in patients with advanced solid tumours (ASPEN-01): A first-in-human, open-label, multicentre, phase 1 dose-escalation and dose-expansion study. Lancet Oncol. 2021, 22, 1740–1751. [Google Scholar] [CrossRef]
- Jiang, Z.; Sun, H.; Yu, J.; Tian, W.; Song, Y. Targeting CD47 for cancer immunotherapy. J. Hematol. Oncol. 2021, 14, 180. [Google Scholar] [CrossRef]
- Lakhani, N.J.; Patnaik, A.; Liao, J.B.; Moroney, J.W.; Miller, D.S.; Fleming, G.F.; Axt, M.; Wang, Y.V.; Agoram, B.; Volkmer, J.-P.; et al. A phase Ib study of the anti-CD47 antibody magrolimab with the PD-L1 inhibitor avelumab (A) in solid tumor (ST) and ovarian cancer (OC) patients. J. Clin. Oncol. 2020, 38, 18. [Google Scholar] [CrossRef]
- Colevas, A.D.; Park, J.J.; Fang, B.; Shao, J.; U’Ren, L.; Odegard, J.; Lal, I.; Phan, M.; Thein, K.Z.; Adkins, D. A Phase 2 Study of Magrolimab Combination Therapy in Patients with Recurrent or Metastatic Head and Neck Squamous-Cell Carcinoma. Int. J. Radiat. Oncol. Biol. Phys. 2022, 112, e42–e43. [Google Scholar] [CrossRef]
- Chauchet, X.; Pernarrieta, E.; Bosson, N.; Calloud, S.; Hellequin, L.; Legrand, M.; Viandier, A.; Richard, F.; Cons, L.; Malinge, P.; et al. CD47 × PD-L1 bispecific antibodies for cancer therapy. J. Immunother. Cancer 2021, 9, A287. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, Z.; Yang, Y. Potential Role of CD47-Directed Bispecific Antibodies in Cancer Immunotherapy. Front. Immunol. 2021, 12, 686031. [Google Scholar] [CrossRef]
- Chen, S.-H.; Dominik, P.K.; Stanfield, J.; Ding, S.; Yang, W.; Kurd, N.; Llewellyn, R.; Heyen, J.; Wang, C.; Melton, Z.; et al. Dual checkpoint blockade of CD47 and PD-L1 using an affinity-tuned bispecific antibody maximizes antitumor immunity. J. Immunother. Cancer 2021, 9, e003464. [Google Scholar] [CrossRef]
- Roohullah, A.; Ganju, V.; Zhang, F.; Zhang, L.; Yu, T.; Wilkinson, K.; Cooper, A.; de Souza, P. First-in-human phase 1 dose escalation study of HX009, a novel recombinant humanized anti-PD-1 and CD47 bispecific antibody, in patients with advanced malignancies. J. Clin. Oncol. 2021, 39, 2517. [Google Scholar] [CrossRef]
- Wang, J.; Sun, Y.P.; Chu, Q.; Duan, J.C.; Wan, R.; Wang, Z.J.; Zhao, J.; Li, H.Y.; Guo, Y.M.; Chen, Y.L.; et al. Abstract CT513: Phase I study of IBI322 (anti-CD47/PD-L1 bispecific antibody) monotherapy therapy in patients with advanced solid tumors in China. Cancer Res. 2022, 82 (Suppl. 12), CT513. [Google Scholar] [CrossRef]
- Stirling, E.; Willey-Shelkey, E.; Wilson, A.; Skardal, A.; Triozzi, P.; Terabe, M.; Miller, L.; Soker, S.; Soto-Pantoja, D. 206 An immune-competent tumor organoid platform to test novel immune checkpoint combinations targeting the receptor CD47 in triple negative breast cancer. J. Immunother. Cancer 2020, 8, A121–A122. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Ka 1/Ms | Kd 1/s | KD M | R2− |
|---|---|---|---|---|
| Anti-PD-L1 mAb | 1.82 × 105 | 4.48 × 10−6 | 2.469 × 10−11 | 0.9834 |
| IAB | 9.50 × 104 | 1.80 × 10−4 | 1.894 × 10−9 | 0.9786 |
| Pro1 | 8.40 × 104 | 1.0 × 10−7 | <1.0 × 10−12 | 0.9683 |
| Pro2 | 7.52 × 104 | 2.85 × 10−7 | 3.794 × 10−12 | 0.9861 |
| Pro3 | 7.21 × 104 | 3.05 × 10−7 | 4.229 × 10−12 | 0.9863 |
| Antibody | Ka 1/Ms | Kd 1/s | KD M | R2 |
|---|---|---|---|---|
| Anti-CD47 mAb | 9.79 × 104 | 3.99 × 10−7 | 4.076 × 10−12 | 0.9930 |
| IAB | 2.57 × 105 | 1.24 × 10−4 | 4.813 × 10−10 | 0.9937 |
| Pro1 | 1.60 × 105 | 1.46 × 10−6 | 9.138 × 10−12 | 0.9954 |
| Pro2 | 1.65 × 105 | 1.98 × 10−6 | 1.202 × 10−11 | 0.9954 |
| Pro3 | 2.42 × 105 | 2.40 × 10−6 | 9.918 × 10−12 | 0.9919 |
| Antibody | EC50 (ng/mL) | Span (%) | R2 |
|---|---|---|---|
| IAB | 0.56 | 23.57 | 0.9666 |
| Pro1 | 0.86 | 25.04 | 0.9194 |
| Pro2 | 1.35 | 10.50 | 0.9468 |
| Pro3 | 0.76 | 34.24 | 0.9599 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bian, Y.; Lin, T.; Jakos, T.; Xiao, X.; Zhu, J. The Generation of Dual-Targeting Fusion Protein PD-L1/CD47 for the Inhibition of Triple-Negative Breast Cancer. Biomedicines 2022, 10, 1843. https://doi.org/10.3390/biomedicines10081843
Bian Y, Lin T, Jakos T, Xiao X, Zhu J. The Generation of Dual-Targeting Fusion Protein PD-L1/CD47 for the Inhibition of Triple-Negative Breast Cancer. Biomedicines. 2022; 10(8):1843. https://doi.org/10.3390/biomedicines10081843
Chicago/Turabian StyleBian, Yanlin, Tong Lin, Tanja Jakos, Xiaodong Xiao, and Jianwei Zhu. 2022. "The Generation of Dual-Targeting Fusion Protein PD-L1/CD47 for the Inhibition of Triple-Negative Breast Cancer" Biomedicines 10, no. 8: 1843. https://doi.org/10.3390/biomedicines10081843
APA StyleBian, Y., Lin, T., Jakos, T., Xiao, X., & Zhu, J. (2022). The Generation of Dual-Targeting Fusion Protein PD-L1/CD47 for the Inhibition of Triple-Negative Breast Cancer. Biomedicines, 10(8), 1843. https://doi.org/10.3390/biomedicines10081843