Abstract
Polymorphonuclear neutrophils (PMNs) are the most abundant white blood cells in the circulation. These cells act as the fast and powerful defenders against environmental pathogenic microbes to protect the body. In addition, these innate inflammatory cells can produce a number of cytokines/chemokines/growth factors for actively participating in the immune network and immune homeostasis. Many novel biological functions including mitogen-induced cell-mediated cytotoxicity (MICC) and antibody-dependent cell-mediated cytotoxicity (ADCC), exocytosis of microvesicles (ectosomes and exosomes), trogocytosis (plasma membrane exchange) and release of neutrophil extracellular traps (NETs) have been successively discovered. Furthermore, recent investigations unveiled that PMNs act as a double-edged sword to exhibit paradoxical activities on pro-inflammation/anti-inflammation, antibacteria/autoimmunity, pro-cancer/anticancer, antiviral infection/COVID-19-induced immunothrombotic dysregulation. The NETs released from PMNs are believed to play a pivotal role in these paradoxical activities, especially in the cytokine storm and immunothrombotic dysregulation in the recent SARS-CoV-2 pandemic. In this review, we would like to discuss in detail the molecular basis for these strange activities of PMNs.
1. Introduction
Human polymorphonuclear neutrophils (PMNs) are the most abundant leukocytes in the circulation, carrying out phagocytosis and killing of the invading pathogens. Traditionally, PMNs are considered as a terminally differentiated and homogeneous population with a short lifespan and low transcriptional capacity [1,2]. Functionally, these cells are oriented to as the first-line responders and powerful guardians of the body’s defense against invaders. They protect the body by phagocytosis, intra-phagolysosomal killing, release of proteolytic enzymes through degranulation, production of reactive oxygen species (ROS), and formation of neutrophil extracellular traps (NETs). Recently, many investigators have revealed the heterogeneity [3] and various novel biological functions of PMNs. This heterogeneity has been particularly identified in certain disease entities [3,4]. For instance, low-density granulocytes (LDGs) in SLE [5,6,7,8,9], myeloid-derived suppressor cells (MDSCs) in specific inflammatory diseases [10], and the N1 neutrophils (N1) with anticancer capacity have been found [11,12]. Besides, in the last decade, scientists have further understood that PMNs are much more complex cells, participating in modulating adaptive immune responses, anti-inflammatory, antiviral, pro-cancerous and anticancerous immunity, as well as an inducer of severe COVID-19 (SARS-CoV-II) complications including hyperinflammation/necroinflammation of the lung, cytokine storm, immunothrombosis, and cardiovascular disease (CVD). These findings have suggested that PMNs not only work at the crossroads of innate and adaptive immunity [13], but act as a double-edged sword with various paradoxical activities. They can either protect the body against infections and cancers, or become an unpredictable rebeldom in the COVID-19 pandemic. We are going to discuss in detail the PMNs regarding their granulopoietic regulation, novel biological/immunological functions, and paradoxical activities in the present review.
2. Regulation of Granulopoiesis, Response to Environmental Factors, and Destinies of PMNs in the Body
PMNs are professional phagocytes growing and differentiating in the bone marrow. Autophagy machineries are highly conserved in these cells [14]. Autophagy is an intracellular homeostatic mechanism of eukaryotic cells essential for the cellular response to starvation or other types of stress such as hypoxia/oxidative burst, DNA damage and infections [15]. The importance of autophagy for efficient differentiation has been demonstrated in lymphocytes [16,17,18], monocytes [19], dendritic cells [20], as well as reticulocytes [21,22], but has been overlooked in neutrophils. However, autophagic regulation is crucial for the effector functions of PMNs, as shown in the following aspects.
2.1. Regulatory Roles of Autophagy in Neutrophil Effector Functions
Rozman et al. [23] disclosed that autophagy is not essential for neutrophil granulopoiesis. Instead, the autophagic activity correlates inversely to the rate of neutrophil differentiation. Thus, a reciprocal relationship between autophagy and PMN differentiation is present, which is modulated by the p38-mTORC1. Nevertheless, autophagy is required for many PMN-mediated effector functions such as granule formation, degranulation, release of neutrophil extracellular traps (NETs), cytokine production, bacterial killing and/or inflammation control [24,25,26,27,28]. Figure 1 depicts the regulatory roles of autophagy in many immune cell functions including those of PMNs but not granulopoiesis.
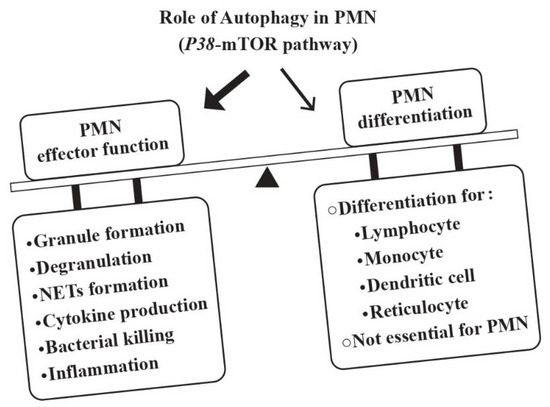
Figure 1.
The roles of autophagy in the development and effector functions of PMNs. Although autophagy is essential for the ontogenetic development of lymphocytes, monocytes/dendritic cells and reticulocytes, it is not essential for neutrophils. Instead, the p38-mTOR-induced autophagy is pivotal for developing effector functions of PMNs including granule formation, degranulation, NETs formation, cytokine production, microbial killing, and inflammation.
2.2. Rapid Sensing and Effective Response of PMNs to Environmental Factors
PMNs can detect different pathogens by using an array of surface-expressed innate immune receptors. Toll-like receptors (TLRs) can sense and bind the pathogen-associated molecular pattern (PAMPs) and damage-associated molecular patterns (DAMPs) from variant origins [29]. Complement receptors (CRs) can identify and attach the complement-fixed immune complexes [30,31]. Low- and high-affinity IgG Fc receptors can recognize and grasp antibody-attached antigens [32]. Thereby, PMNs act as powerful effectors for TLR-, complement- and antibody-mediated inflammation. In addition, PMNs also play a pivotal role in the resolution of tissue inflammation via secretion of lipid anti-inflammatory molecules, resolvins, to inhibit inflammatory reactions [33]. In short conclusion, PMNs can be affected by the environmental factors to which they are exposed and partake in coordinating inflammation and/or anti-inflammation processes for immune adaptation and homeostasis, as shown in Figure 2.
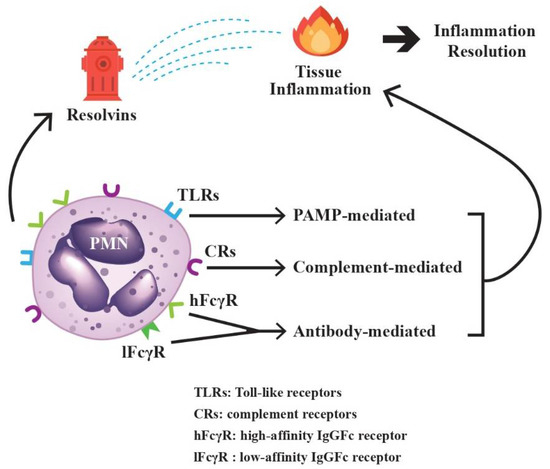
Figure 2.
The paradoxical activities of PMNs on tissue inflammation and inflammation resolution. The surface-expressed Toll-like receptors (TLRs) bind to pathogen-associated molecular patterns (PAMPs), complement receptors (CRs) bind to antigen-antibody-complement immune complexes, and high-affinity (hFcγR) and low-affinity IgG Fc receptors (lFcγR) bind to IgG antibodies, which then can induce tissue inflammation. On the other hand, resolvins, the metabolic products of omega-3 polyunsaturated fatty acid synthesized during the initial phases of acute inflammatory responses, can promote the resolution of inflammation. Both inflammation and its resolution can be mediated by PMNs for immune homeostasis.
2.3. Factors Influencing the Destinies of PMNs
Under physiological conditions, the mature circulating PMNs have a very short half-life of no more than one day in vivo [34,35]. However, Pillay et al. [36] conducted an in vivo incubation of human PMNs with 2H2O and revealed that the lifespan of normal PMNs is around 5.4 days. Ordinarily, the lifespan of PMNs can be lengthened during the course of inflammatory response by signals in the inflammatory milieu [37]. In general, these cells constitutively undergo spontaneous apoptosis, but their short-time survival can be extended by certain growth factors, proinflammatory cytokines or bacterial products including G-CSF, GM-CSF, IFN-γ, TNF-α, IL-2, IL-6, or bacterial lipopolysaccharides (LPS) [38,39,40,41,42]. Hsieh et al. [43] have elucidated the molecular basis for spontaneous PMN apoptosis. The group found that both Fas and Fas ligand (FasL) molecules are simultaneously expressed on the PMN surface. In addition, other apoptosis-related molecules, p53 and BCL-2 but not c-myc, are also expressed in PMN cytoplasm. Interestingly, the FasL molecules were found rapidly disappearing after 24 h of incubation. The authors concluded that the Fas-mediated pathway after Fas–FasL interaction is one of the molecular mechanisms in inducing spontaneous PMN apoptosis after PMN–PMN interactions. It has also been found that PMN apoptosis is tightly regulated by a complex network of signaling pathways controlling the key protein molecules of the BCL-2 family via activation of MAP kinases, NF-κB, and caspase-degraded BCL-2 homolog, as well as Myeloid Cell Leukemia-1 (MCL-1) [44,45]. MCL-1, as a survival molecule [46,47], can sustain PMN survival via heterodimerization with and neutralization of proapoptotic BCL-2 family members, Bim or Bak, in the mitochondrial outer membrane [48,49,50]. On the contrary, the FasL, by bridging extracellular domains of TNF or TRAIL to membrane “death receptors”, TNF-R1 or TRAIL-R1/R2, can activate cytoplasmic “death domains”, FADD or FADD/TRADD. The activation then triggers caspase-8 and -3 to induce apoptosis [51,52]. It is worthy to note that the influence of microbes in alteration of the evolving routes of PMN is highly variable [53]. It could be microbe-specific, ranging from prolongation of PMN lifespan to rapid PMN breakdown after microbe phagocytosis. The molecular basis of the factors implicated in the differentiation of PMNs is shown in Figure 3.
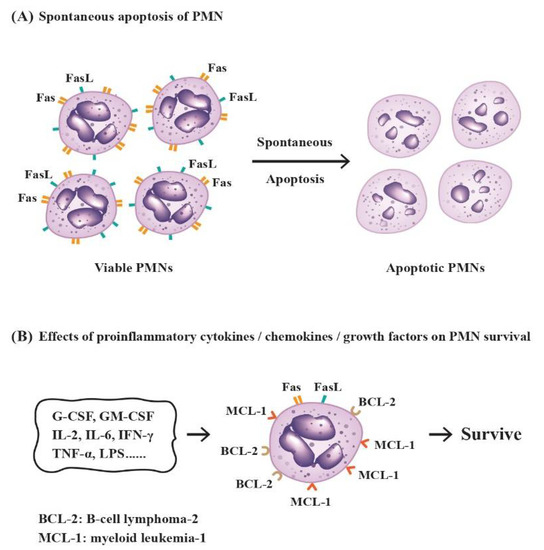
Figure 3.
The molecular basis for spontaneous apoptosis and survival prolongation of PMNs by proinflammatory cytokines/chemokines/growth factors in the physiological or inflammatory environment. (A) Induction of spontaneous PMN apoptosis by interactions of Fas ligand (FasL) and Fas receptor (Fas, CD95) expressed on the cell surface of neighboring PMNs in normal condition; (B) The lifespan of PMNs can be prolonged by inflammation-related factors in the environment via increased expression of survival molecules BCL-2 and MCL-1.
3. Novel Biological/Immunological Functions of PMNs
PMNs are traditionally regarded as the first-line defending cells against microbial invasion by the way of phagocytosis, intracellular proteolytic killing and eradication of the microbes by reactive oxygen species (ROS). Nevertheless, a complete deletion of PMN (<0.5%) in rats with monoclonal anti-granulocyte antibody RP-3 that did not deplete innate and adaptive immune-related cells could alter the adaptive immune responses [54,55,56]. Yue et al. [57] and Dallegre et al. [58], in their in vitro studies, demonstrated that PMNs could exert cytotoxic effect in the presence of mitogen via a mitogen-induced cell-mediated cytotoxicity (MICC). Besides, other investigators discovered that antibody-dependent cell-mediated cytotoxicity (ADCC) is a universal immune activity mediated by IgG-Fc receptor-bearing cells including T cells, B cells, monocytes/macrophages and PMNs [59]. These results indicate that PMNs can actively participate in the effective immune responses in the body beyond the pathogen engulfing and killing functions. In this section, we will discuss more novel biological functions of PMNs involved in the immune network and immune homeostasis. Table 1 lists these new biological and immunological functions of PMN.

Table 1.
Novel biological/immunological functions of PMN.
3.1. Biosynthesis and Secretion of Complement Component 3 (C3) and Factor B
Okuda T [60] firstly discovered that murine PMN can synthesize complement component C3 and factor B. Botto et al. [61] and Yu et al. [62] have subsequently confirmed that human PMN can synthesize and secrete functional C3 either spontaneously [62] or after activation by LPS or TNF-ɑ [61]. This PMN function would become an important mechanism for host defense at sites of inflammation in addition to granule proteins and ROS generation.
3.2. Release of Granule Proteins, Cytokines, Chemokines, and Growth Factors from PMNs for Cell–Cell Communication and Immune Modulation
PMNs not only play a pivotal role in the inflammatory reaction, but act as an important modulator in immune network by releasing a number of mediators. These mediators include granule proteins released through degranulation, and cytokines/chemokines/growth factors to actively take part in the immune system. The detailed steps of these functions will be discussed in the following sections.
3.2.1. Degranulation to Liberate Azurophilic and Specific Granules
After activations by different stimuli, granule contents are released into the intracellular phagosomes or out to the extracellular space. This is called degranulation. During this process, membrane-attached granules fuse with phagosomes and plasma membrane, allowing a new biomarker expression on the cell surface [63]. Usually, the neutrophil granule proteins can be classified into azurophilic (primary) and specific (secondary) proteins. It should be noted that an excess of granule proteins release can lead to tissue damage, chronic inflammation and immune dysfunction [64,104]. Li et al. [65] found that the surface-expressed lactoferrin on PMN, a specific granule protein, could modulate Th1/Th2 cytokine production. In contrast, myeloperoxidase, an azurophilic granule protein, can induce chronic inflammation and NET formation [64].
3.2.2. The Production of Cytokines/Chemokines/Growth Factors from PMNs for Immune Modulation
Bazzoni et al. [66] have firstly demonstrated that the phagocytosis of IgG opsonized-yeast particles by human PMNs can result in the expression and release of TNF-ɑ, but not IL-6. LPS stimulation on PMNs also releases TNF-ɑ. Since then, a bunch of cytokines/chemokines/growth factors have been successively discovered to be also produced and emancipated by PMNs. Compared to that produced by other innate or adaptive immune cells, the amount of cytokine production by individual PMN is relatively low. Nevertheless, this drawback can be partially compensated by the huge number of PMNs in the blood. In conjunction with the secreted cytokines/chemokines/growth factors [67,68,69,70,71] and the released granule proteins, NET formation as well as trogocytosis can render PMNs to intimately interact with the other immune-related cells, contributing essentially to the homeostasis of the immune system [105,106,107,108,109,110]. Table 2 summarizes the cytokine/chemokine/growth factor expressions in PMNs in vitro and in vivo, which is adapted from those reported by Cassatella et al. and Tsai et al. [110] with permission.
Table 2.
The cytokines/chemokines/growth factors expression in normal human PMNs in vitro and in vivo *.
3.2.3. Liberation of Ectosomes and Exosomes from PMNs to Affect the Biological Functions of the Remote Cells or Tissues
In addition to the release of granule proteins/cytokines/chemokines/growth factors as mentioned in the Section 3.2.1, PMNs can also free extracellular vesicle (EV) to affect the biology/physiology of remote cells or tissues. The EVs released from PMNs contain two microvesicles, ectosomes (Ect) and exosomes (Exo), with a vesicle size ranging from 50 to 1000 nm in diameter. These PMN-derived EVs exhibit many important characteristics in the body defense, inflammatory responses and wound healing [72,73,74].
Stein et al. [111] have found that exocytosis of human PMNs can be elicited in the presence of a small amount of autologous complements. Later, Hess et al. [112] observed that N-formyl-methionyl-leucyl-phenylalanine (fMLP) or C5a can induce ectosome release from PMNs within a few minutes after stimulation. These extruded ectosomes contain a selective set of proteins originating both from the cell membrane and from the intracellular granule molecules such as neutrophil elastase (NE), myeloperoxidase (MPO), and proteinase 3 (Pr3) packed in the plasma membrane. However, a unique property of ectosomes is the exposure of phosphatidylserine (PS) in the outer leaflet of membrane [112], which is different from that of exosomes. In contrast, exosomes are defined as small membrane vesicles formed by inward budding of the endosomal membrane with little PS expression [113]. Functionally, ectosomes exhibit generic functions to downregulate inflammation and immunity, whereas exosomes potentiate the immune responses [114].
Suppressive Effects and the Signaling Pathways of PMN-Derived Ectosomes (PMN-Ect) on Macrophage Maturation
Gasser et al. [73] have shown that PMN-Ect could suppress the release of TNF-α, IL-8, and IL-10 from activated human monocyte-derived macrophages. Eken et al. [115] further disclosed that PMN-Ect could interfere with the LPS-stimulated dendritic cells maturation including morphological changes, phagocytic activity, surface molecule expression, cytokine release, and the capacity to induce T-cell proliferation. The same group then elucidated the signaling pathways of PMN-Ect-induced events against inflammation in macrophages/dendritic cells. These inhibitory processes induced by PMN-Ect include the MerTK pathway, Ca2+-flux and the release of stored TGF-β1 in the macrophages [116,117]. Expectedly, these PMN-derived granule cargos were also found to pave the way for tumor growth and progression [118], an untoward effect for the host.
The Modulatory Roles of PMN-Derived Exosomes (PMN-Exo) on the Immune Responses
Exosomes are the endosomal-derived microvesicles with 30–100 nm in diameter. These vesicles contain lipids, proteins and nucleic acids (DNA, mRNA and microRNA [miRNA]) inside. Many normal cells including immune-related cells (T cells, B cells, dendritic cells, mast cells and PMNs), neuron cells, astrocytes, epithelial cells and various tumor cells can release Exo. Once Exo are extruded into the extracellular milieu, they can be found in many biological fluids including plasma, urine, effusion, synovial fluid, saliva, or breast milk [119]. These microvesicles can transfer their cargo into the remote recipient cells to modify their biological activities.
It is worthy to note that miRNAs in the Exo can modify the biological processes of the recipient cells, leading to autoimmune diseases [120,121] including SLE [122,123], RA [124], and systemic sclerosis (SSc) [125]. In particular, PMN-Exo can be involved in different pathological processes [126] in SSc [127] and dermatomyositis [128] via deranging various signaling pathways. These pieces of evidence demonstrate again that PMNs act as a double-edged sword to exert paradoxical effects in clinical medicine.
3.3. Induction of MHC-II Expression on PMNs by T-Cell-Derived Cytokines, Rendering PMN Mimicking Antigen-Presenting Cells (APC)
PMNs express a wide range of pattern recognition receptors [29], IgG-Fc receptors, and complement receptors [2] in resting state for mediating innate immune responses, as mentioned in Section 2.2. Interestingly, Oehler et al. [129] demonstrated that 9-day culture of PMNs in the presence of GM-CSF, IL-4 and IFN-α can augment the allogeneic stimulatory activity of the specific tetanus toxoid antigen after mixing with autologous memory T cell. Later, many authors found that PMNs activated by the memory T-cell-derived cytokines can express MHC-II and co-stimulatory molecules of CD80 and CD86, behaving like antigen-presenting cells [75,76,130]. Vono et al. [77] found that the cognate antigens of cytomegalovirus pp65- or influenza hemagglutinin-pulsed PMNs can present these two antigens to the autologous antigen-specific CD4+T cells in an MHC-II-dependent manner. These data support that PMNs acquire the capacity for antigen presentation to memory type CD4+ T cells in vitro and ex vivo. In addition, Meinderts et al. [78] demonstrated that human PMNs acquired antigen-presenting phenotype with expression of MHC class-II and co-stimulatory molecules, CD40 and CD80, following engulfment of IgG-opsonized erythrocytes. Besides, Polak et al. [79,80], by investigating IgE-mediated allergy, found that HLA-DR(+)-PMNs, and allergen-specific T cells accumulate in the sites of allergic late-phase reaction (LPR). In an in vitro experiment, the group further found that in the presence of a cocktail of GM-CSF, IFN-γ, and IL-3, PMNs internalized, processed, and then presented the allergen via HLA-DR loci to elicit the proliferation and cytokine production of the allergen-specific T cells. These results can support the potential antigen-processing capacity of PMNs in the stimulation of T cells or in the production of T-cell-derived cytokines.
3.4. Trogocytosis (Plasma Membrane Transfer) among PMN, Non-Immune, and Immune-Related Cells
Trogocytosis (trogo means gnaw) is characterized by the transfer of plasma membrane fragments between two cells in contact after forming an immunological synapse. It has been demonstrated that trogocytosis is an active energy-consuming rapid transfer process after a conjugation between two homogeneous or heterogeneous living cells. The energy-requiring processes in trogocytosis include actin polymerization, membrane remodeling, signaling transfer, and finally the plasma membrane merges in the cell surface. This kind of membrane transfer has been demonstrated among immune, non-immune and even microbial cells [81]. The biological significance of trogocytosis may include: (1) cell–cell information exchange, (2) growth during embryonic development, (3) “nibbled to death” of infectious microbes, (4) immunoregulation, and (5) cancer immunity [82]. The following sections will discuss in detail the biological significance of trogocytosis mediated by PMNs.
3.4.1. Elimination of the Intracellular Parasites or Unwanted Cells by PMN-Mediated Trogocytosis
Mercer et al. [131] have found that human PMNs can kill Trichomonas vaginalis in a dose-dependent, contact-dependent, and NET-independent manner via “bites” of the parasites until death. Both trogocytosis and parasite killing are dependent on the presence of PMN’s serine proteinase and human serum factors. Furthermore, Olivera-Valle et al. [132] found that PMNs attacked and killed excessive exogenous immobile sperms in the vagina via trogocytosis with high efficiency after contact with these sperms without inducing vaginal mucosa damage or infertility.
Taylor et al. [83] are the first authors to propose a specialized form of trogocytosis mediated by Fcγ receptors (FcγR) on effector cells in cancer immunotherapy by using anticancer monoclonal antibodies. The hypothesis is further supported by Valgardsdottir et al. [84] that PMNs can carry out mostly trogocytosis rather than phagocytosis of the anti-CD20-opsonized chronic lymphocytic leukemia cells in autoantibody-based anticancer therapies.
3.4.2. Trogocytosis among PMNs and Other Immune-Related Cells for Immune Modulation
Poupot et al. [85] have found that spontaneous membrane transfer occurs among homotypical leukemia cell lines without stimulation of the prolongation of cell survival. Honer et al. [86] demonstrated the occurrence of trogocytosis between PMNs and tumor cells in the presence of antitumor antibodies. Li et al. [87] have observed many differences in the mechanisms and biological significance of trogocytosis between normal human PMNs and mononuclear cells (MNCs). The group disclosed that membrane transfer from MNCs to PMNs occurred at the site of immunological synapse for transducing survival and activation signals. The membrane transfer from MNCs to PMNs enhances PMN functions, which is dependent on actin polymerization, clathrin activation and the presence of Fcγ receptors. On the other hand, membrane transfer from PMNs to MNCs depends on MAP kinase and PKC signaling pathways for cell–cell communication and immune modulation.
To date, trogocytosis has been found among various types of immune cells including T cells [88], B cells [89], NK cells [133], macrophages/DCs [134], basophils [135], and innate lymphoid cells [136]. In a biological sense, trogocytosis participates in the elimination of invading pathogens and tumor cells to protect the body. On the other hand, in an immunological sense, trogocytosis is implicated in the activation or inhibition for the immune homeostasis [137,138,139].
3.5. Biological and Pathobiological Roles of NET Formation from PMNs
The most powerful antimicrobial mechanism by PMNs is the extrusion of the intracellular structures in the form of NETs into the surrounding environment [140]. NETs can not only effectively trap the invading pathogens for preventing their spread, but also rapidly kill the pathogens by the chromatin-attached granule molecules such as proteinase, elastase, myeloperoxidase and LL-37.
NETs are large web-like structures composed of cytosolic and granule proteins assembled on a scaphoid of decondensed chromatins derived mostly from nucleus and less from mitochondria. However, once the clearance of this exposed web-like structure in the blood vessels or tissues is insufficient, NETs may induce immune-mediated diseases such as rheumatoid arthritis [141]. This is because DNA-containing NETs per se can stimulate proinflammatory cytokine production via TLR9 on the innate immune cells [90]. In a clinical sense, the excessive NET formation in the blood vessels may induce a wide range of pathological conditions including vascular thrombosis/atherosclerosis [142,143,144], autoimmune diseases [145,146] and tumor progression/metastasis [143,147]. Conversely, impaired NET formation may be found in aged individuals due to defective innate immunity [148]. Recently, Lu et al. [149] found that FcγRIII engagement can augment PMA-stimulated NET formation partially via cross talk between Syk-ERK-NF-κB and PKC-ROS signaling pathways. Farrera et al. [150] reported that the clearance of NETs depends on complement components C1q, DNase 1, C-reactive protein and macrophage engulfment.
4. Heterogeneity of PMN in Facilitating or Deterring Tumorigenesis
Evidence has shown that the functional heterogeneity of PMNs depends on the tumor microenvironment (TME) for determining either pro- or antitumor effect. PMNs exhibit an antagonizing effect at the early stage of tumorigenesis [151], whereas they exhibit a facilitating effect [11] at the late stage of tumorigenesis. It has been reported that G-CSF [152] and IL-8 [153] are produced from tumor cells and tumor-surrounding cells for skewing the number and nature of neutrophils in the TME. Thereby, tumor-associated PMNs (TANs) contain heterogeneous populations with different functional capacities ranging from effector to myeloid suppressor cells [154,155]. Arbitrarily, the TANs can be classified into antitumor neutrophils (N1) upon stimulation by IFN-β [156] and tumor-promoting neutrophils (N2) upon stimulation by TGF-β [157]. The antitumor N1 are characterized by high expression levels of TNF-α, Fas, and ICAM, but low expression level of arginase [158]. In addition, N1 cells can release ROS, proteolytic enzymes, NETs and exhibit high adherence capacity to destroy the tumor cells. They can also promote T-cell immunity by recruitment and activation of CD8+ T cells via IFN-α production to induce cell-mediated cytotoxicity [159].
The possible mechanisms of N1 cells to kill tumor cells are through (1) ADCC [160], (2) antibody-opsonized FcγR-mediated trogocytosis [161], and (3) FcαR-mediated antibody-dependent ADCC that can be potentiated by CD47-SIRPA checkpoint blockade [161,162]. On the contrary, FcγRIIIb (as a decoy receptor), which is different from FcγRIIa, may restrict antibody-dependent destruction of cancer cells by human PMNs [163]. Recently, NETs have been found to exert the pro-tumor effects on the malignant tumor progression via mechanisms involving the establishment of an inflammatory microenvironment in association with other pro-tumor mechanisms such as inflammasomes or autophagy [164,165,166,167]. The potential antitumor mechanisms by N1 neutrophils are illustrated in Figure 4.
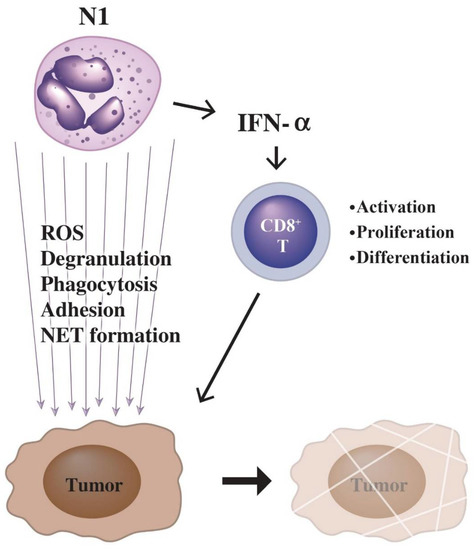
Figure 4.
Killing of tumor cells by N1-type PMNs is through two pathways: (1) enhanced tumor-killing activity of cytotoxic CD8+ T-cell by IFN-α released from PMNs; (2) tumor-killing molecules released from PMNs including ROS, granule proteins, and NETs in association with phagocytosis and adhesion of tumor cells by PMNs.
In addition, a particular group of granulocytic myeloid-derived suppressor cells (MDSCs) is widely described as an immature subset by their ability to impede both innate and adaptive immunity. Accordingly, this subset of cells have been demonstrated both in vitro and in vivo to inhibit T-cell immunity as well as to promote the growth and spread of cancers [168,169]. The neutrophil heterogeneity in facilitating and deterring tumorigenesis is summarized in Table 3.
Table 3.
The functions and characterization of different heterogeneity of neutrophil subpopulations including N1, N2, and granulocytic MDSCs, relevant to the tumorigenesis.
5. Impact of PMNs on Cardiovascular Disease (CVD)
Besides the role of PMN-NETs on vascular thrombosis and atherosclerosis [142,143,144], as mentioned in Section 3.5, inflammation is also regarded as a risk factor for CVD in apparently healthy people [170], independent of dyslipidemia [171]. PMNs have been found to destroy the endothelial cells (ECs) by releasing Pr3, NE [172] and MPO [173]. These proteolytic enzymes can break down basement membranes and induce endothelial apoptosis. Indeed, the serum levels of Pr3 and NE are found to increase in patients with acute myocardial infarction (AMI) [174,175]. In addition, release of ROS from PMN is another potent EC-damage factor in CVD [176]. Recently, Sreejit et al. [177] discovered that a PMN-derived alarmin (i.e., S100A8/A9) can induce inflammation and cardiac injury after myocardial infarction. Critical reviews for the impact of CVDs on PMNs have been reported by Bonaventura et al. [178] and Silvertre-Roig et al. [179].
7. Overwhelming Immune Responses Relevant to PMNs in Pandemic Coronavirus Disease
Coronavirus disease 2019 (COVID-19 or SARS-CoV-II) is a virus-infected respiratory disease that can potentially progress to acute respiratory distress syndrome (ARDS), cytokine storm, and multiple-organ failure. This hyper-inflammatory status is originated from dysregulatory immune responses as a result of impaired T-cell suppression and excessive innate immune activation [182,183,184]. Some of the patients with COVID-19 infection displayed severe lymphopenia (decrease in CD4+ T, CD8+ T, and B cells) and delayed lymphocyte activation [185]. In contrast, elevated serum concentration of IP-10 (IFN-γ-induced protein 10) and GM-CSF in COVID-19 disease is compatible with enhanced T-cell and monocyte recruitments, increased proinflammatory cytokine production and excessive PMN chemotaxis [185]. Accordingly, the overwhelming innate and adaptive immune responses inevitably elicit ARDS in some patients with COVID-19 disease [186]. Yao et al. [187], by analyzing transcriptomic profiles of immune-related cells, revealed that defective antigen presenting and IFN responsiveness of monocytes were present in patients with COVID-19-induced ARDS, in contrast to higher lymphocyte responsiveness. Besides, the gene expression involved in cytotoxic activity was decreased in NK, CD8+T and B cells in these patients. Delayed viral clearance is also found in severely ill COVID-19 patients. de Candia et al. [188] found that the mortality of SARS-CoV-2 infection was higher in the elderly than in younger adults and apparently rare in children, which seemed attributable to the innate and adaptive immune status. Although PMNs can facilitate virus clearance as mentioned in Section 6, neutrophilic inflammation may conversely contribute to the higher mortality of COVID-19 patients with underlying comorbidities such as diabetes or CVD. The molecular mechanisms of PMNs in triggering severe complications such as cytokine storm, hyperinflammation/necroinflammation, immunothrombotic dysregulation, and multi-organ failure in relation to NETs formation in COVID-19 disease will be discussed in the following sections.
7.1. The role of PMN-Derived NETs in Inducing Hyperinflammation, Lung Cell Death, Cytokine Storm, ARDS, and Immunothrombotic Dysregulation in COVID-19 Disease
Scientists have observed that an increased number of circulating PMNs may become an indicator for the severity of respiratory symptoms and poor prognosis in COVID-19 infection [189]. PMN-derived NETs are considered one of the potent inducers in lung inflammation. Veras et al. [190] unraveled that viable SARS-CoV-2 can trigger NETs release from PMNs, which is dependent on the activation of angiotensin-converting enzyme receptor 2 (ACE-R2), serine proteinase, and peptidylarginine deiminase 4 (PAD4) in addition to viral replication per se. The activation of these molecules can be reflected by elevated NET concentration in plasma, tracheal aspirate and autopsied lung tissues in severely ill patients. Moreover, these NETs derived from PMNs can concomitantly cause death of the pulmonary epithelial cells and hyperinflammation of the lungs [190,191,192,193].
Cytokine storms are usually driven by the unrestrained activation of leukocytes to produce a huge amount of proinflammatory cytokines including IL-1β, IL-6, TNF-α, IL-17, IFN-α, along with PMN-NETs release. These pathological factors elicit severe damage on the inflamed blood vessels and infiltrated tissues, especially in patients with diabetes and hypertension [194,195,196].
7.2. The Interactions of NETs, Complements, Coagulation Factors, and Platelets in the Immunothrombotic Dysregulation in COVID-19 Infection
Nicolai et al. [197] examined the autopsied cases of COVID-19 disease and found extensive inflammatory microvascular thrombus formation in the lung, kidney and heart tissues containing NETs in association with platelets and fibrin. Besides, patients with COVID-19 disease also exhibited PMN-platelet aggregates with distinct PMN and platelet activation markers in blood. Middleton et al. [198] have also confirmed NET-containing microthrombi with PMN and platelet infiltration in autopsied lungs. Radermecker et al. [199] disclosed that NETs could contribute to inflammation-associated lung damage, thrombosis, and fibrosis in severely ill patients. Regarding the molecular basis of NET-induced immunothrombus formation in COVID-19-infected patients, Skendros et al. [200] found increased plasma NETs, tissue factor (TF) activity, and soluble complements sC5b-9 in COVID-19 patients. In addition, PMNs from those patients released high amounts of TF and NET-carrying active TF. These results have indicated a pivotal role of complements and NETs in immunothrombus formation in COVID-19 patients. Busch et al. [201] have explored the role of intrinsic pathway of coagulation cascades including plasma kallikrein and bradykinin in COVID-19-induced immune thrombosis. They concluded that the hypercoagulability and thrombotic events in COVID-19 patients are driven by PMN-NETs, complement activation, and contact activation. The use of C5a blockers, the plasma kallikrein and blood coagulation factor XIa (FXIa) inhibitors, and agents neutralizing extracellular histones may be beneficial for the control of COVID-19-induced immunothrombotic dysregulation.
Recently, Ercan et al. [202] analyzed phenotypic changes of platelets in COVID-19 patients and found a decrease in the total amount of integrin αIIb (ITGA2B), a subunit of αIIbβ3, in the patients compared to healthy controls. Higher consumption of fibrin-stabilizing factor, i.e., coagulation factor XIIIA (F13A1), in platelets was found in COVID-19 patients. Conversely, increased amounts of annexin A5, eukaryotic initiation factor 4A-1 (ElF4A1) and transaldolase 1 (TALDO1) in platelets were correlated to nasopharyngeal COVID-19 viral load. Furthermore, the levels of 2 protein disulfide isomerase, P4HB and PD1A6, which facilitate thrombosis, were found to be increased in COVID-19 disease. The crucial role of PMNs in the immunothrombosis in COVID-19-infected patients has been critically reviewed by Iliadi et al. [203] and Bautista-Becerril et al. [204].
A proposed molecular basis for PMN-derived NETs in the pathogenesis of lung cell death, hyperinflammation, cytokine storm, multiple-organ failure, vascular damage, and immunothrombotic dysregulation in SARS-CoV-infected patients is provided in Figure 5.
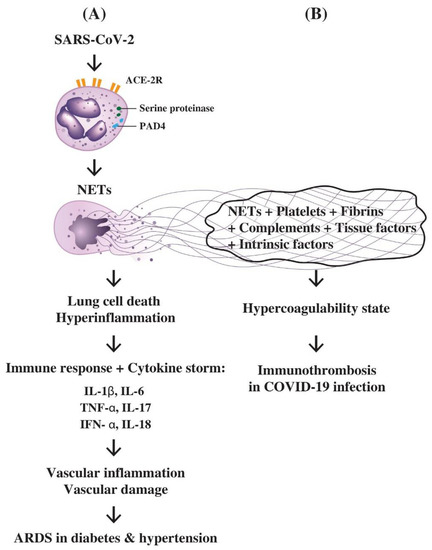
Figure 5.
The molecular basis of PMNs in inducing hyperinflammation of lung, cytokine storm and immunothrombosis during SARS-CoV-2 infection in patients with diabetes or hypertension. (A) Attachment of SARS-CoV-2 virus to ACE2 receptors on PMN results in the enzymatic activation of serine proteinase and PAD4 to induce NETs formation. The NETs can cause pulmonary epithelial cell death and hyperinflammation in the lung. The overwhelming immune responses with profound proinflammatory cytokine production including IL-1β, IL-6, IL-8, IL-17, IFN-α, and TNF-α elicit cytokine storm, extensive vascular inflammation and damage leading to acute respiratory distress syndrome (ARDS); (B) The trapping of platelets, fibrins, complements, tissue factors, intrinsic coagulation factors, and tissue debris by NETs released from PMNs lead to a hypercoagulability state of the body. Finally, immunothrombosis in the lung, heart, brain, kidney or gastrointestinal tract occurs in COVID-19 disease.
8. Conclusions
A number of novel biological/immunological functions of PMNs have been successively found. Some of the unique functions including NET formation, N1/N2 neutrophil and granulocytic MDSCs heterogeneity, MHC-II antigen expression, trogocytosis, extrusion of ectosomes and exosomes, and antiviral activity can act as a fast and effective defender to protect the body. On the contrary, PMNs may also play a dual role to exhibit paradoxical activities to favor or to oppose inflammation, to exert antimicrobial effect or autoimmunity, to exhibit pro-cancer or anticancer behaviors, to present antiviral effect or facilitate SARS-CoV-II-induced immunothrombotic dysregulation in clinical medicine. The NETs derived from activated PMNs play a pivotal role in these paradoxical activities. It is quite important to further elucidate more molecular evidence for this strange phenomenon and to design an effective therapeutic strategy for overcoming these untoward effects of PMNs in clinical medicine as shown in Table 4. Accordingly, we propose several tips to take advantages of these paradoxical characteristics of PMNs for SARS-CoV-2 therapy, as shown below:
Table 4.
Untoward effects of deranged PMN functions in clinical practice.
- (1)
- Adequate use of the PMN-released defensins (HNP1, HD5) or antimicrobial peptide retrocyclin-101 (RC101) to block SARS-CoV-2 entry.
- (2)
- Facilitating NET release initially to trap and kill the virus followed by adding of complement C1q to rapidly clear the accumulated NETs.
- (3)
- Application of monoclonal anti-IL-17 antibody to suppress SARS-CoV-2 virus-induced hyperinflammation.
- (4)
- Induction of granulocytic MDSCs by TGF-β for anti-inflammation effect and immunosuppression of the cytokine storm.
Author Contributions
Conceptualization, C.-L.Y. and C.-Y.T.; original writing—draft preparation, T.-H.W. and S.-C.H.; final writing—review and editing, T.-H.L., C.-H.L., H.-T.L., C.-Y.S., K.-J.L., C.-H.W. and Y.-M.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the Ministry of Science and Technology (MOST-107-2314-B075-051-MY3), Executive Yuan, Taiwan.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank all of the individuals who participated in this investigation.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ACE2 | Angiotensin-converting enzyme 2 |
| ADCC | antibody-dependent cellular cytotoxicity |
| Bak | Bcl-2 homologous antagonist killer |
| Bim | Bcl-2-like protein 11, commonly called BIM |
| Bcl-2 | B cell lymphoma 2 protein family (for regulation of apoptosis) |
| C | complement component |
| c-myc | human homologue of virus Myelocytomatosis, cytoplasmic domain |
| CVD | cardiovascular disease |
| C1q | q fragment of complement C1 |
| DAMP | Damage-associated molecular pattern |
| DNA | deoxyribonucleic acid |
| Ect | ectosome |
| ERK | extracellular signal-regulated kinase |
| EV | extracellular vesicle |
| Exo | exosome |
| FADD | Fas-associated protein with death domain |
| Fas | apoptosis antigen 1 (cluster of differentiation 95) |
| FasL | Fas ligand |
| FcγR | receptor for fragment C of immunoglobulin G |
| fMLP | formyl-methionyl-leucyl-phenylalanine |
| FXIa | coagulation factor 11, ‘a’ component |
| G-CSF | granulocyte colony stimulating factor |
| GM-CSF | granulocyte-macrophage colony stimulating factor |
| ICAM | intercellular adhesion molecule |
| IFN | interferon |
| IgG-Fc | Fragment C of immunoglobulin G |
| IL | interleukin |
| LDG | Low-density granulocyte |
| LPS | lipopolysaccharide |
| MCL-1 | Induced myeloid leukemia cell differentiation protein 1 |
| Mer-TK | proto-oncogene receptor tyrosine kinase |
| MDSC | myeloid-derived suppressor cell |
| MHC-II | class 2 major histocompatibility complex |
| MICC | mitogen-induced cellular cytotoxicity |
| mi-RNA | micro-ribonucleic acid |
| MPO | myeloperoxidase |
| N1 | type 1 neutrophil |
| N2 | type 2 neutrophil |
| NE | neutrophil elastase |
| NET | neutrophil extracellular trap |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| PAD-4 | protein arginine deiminase 4 |
| PAMP | pathogen-associated molecular pattern |
| PKC | protein kinase C |
| PMN | polymorphonuclear neutrophil |
| Pr3 | proteinase 3 |
| PS | phosphotidylserine |
| p53 | cellular tumor phosphoprotein 53 |
| ROS | reactive oxygen species |
| SARS-CoV-II | Severe acute respiratory syndrome-related coronavirus type II or coronavirus found in 2019 (COVID-19) |
| sC5b-9 | soluble complement fragments 5b to 9 complex |
| SIRPA | Signal regulatory protein alpha |
| SLE | systemic lupus erythematosus |
| SSc | systemic sclerosis |
| Syk | spleen tyrosine kinase |
| TAN | tumor-associated neutrophil |
| TF | tissue factor |
| TME | tumor microenvironment |
| TNF | tumor necrosis factor |
| TGF | transforming growth factor |
| TRADD | tumor necrosis factor receptor type 1-associated death domain |
| TRAIL-R | TNF-related apoptosis-inducing ligand receptor |
| TLR | Toll-like receptor |
References
- Summers, C.; Rankin, S.M.; Condliffe, A.M.; Singh, N.; Peters, A.M.; Chilvers, E.R. Neutrophil kinetics in health and disease. Trends Immunol. 2010, 31, 318–324. [Google Scholar] [CrossRef] [Green Version]
- Cassatella, M.A. Neutrophil-derived proteins: Selling cytokines by the pound. Adv. Immunol. 1999, 73, 369–509. [Google Scholar] [CrossRef]
- Yang, P.; Li, Y.; Xie, Y.; Liu, Y. Different faces for different places: Heterogeneity of neutrophil phenotype and function. J. Immunol. Res. 2019, 2019, 8016254. [Google Scholar] [CrossRef] [Green Version]
- Fine, N.; Tasevski, N.; McCulloch, C.A.; Tenenbaum, H.C.; Glogauer, M. The neutrophil: Constant defender and first responder. Front. Immunol. 2020, 11, 571085. [Google Scholar] [CrossRef]
- Hacbarth, E.; Kajdacsy-Balla, A. Low density neutrophils in patients with systemic lupus erythematosus, rheumatoid arthritis, and acute rheumatic fever. Arthritis Rheum. 1986, 29, 1334–1342. [Google Scholar] [CrossRef]
- Bennett, L.; Palucka, A.K.; Arce, E.; Cantrell, V.; Borvak, J.; Banchereau, J.; Pascual, V. Interferon and granulopoiesis signatures in systemic lupus erythematosus blood. J. Exp. Med. 2003, 197, 711–723. [Google Scholar] [CrossRef] [Green Version]
- Carmona-Rivera, C.; Kaplan, M.J. Low-density granulocytes: A distinct class of neutrophils in systemic autoimmunity. Sem. Immunopathol. 2013, 35, 455–463. [Google Scholar] [CrossRef]
- Fu, J.; Tobin, M.C.; Thomas, L.L. Neutrophil-like low-density granulocytes are elevated in patients with moderate to severe persistent asthma. Ann. Allergy Asthma Immunol. 2014, 113, 635–640.e2. [Google Scholar] [CrossRef]
- Grayson, P.C.; Carmona-Rivera, C.; Xu, L.; Lim, N.; Gao, Z.; Asare, A.L.; Specks, U.; Stone, J.H.; Seo, P.; Spiera, R.F.; et al. Neutrophil-related gene expression and low-density granulocytes associated with disease activity and response to treatment in antineutrophil cytoplasmic antibody-associated vasculitis. Arthritis Rheumatol. 2015, 57, 1922–1932. [Google Scholar] [CrossRef]
- Waight, J.D.; Hu, Q.; Miller, A.; Liu, S.; Abrams, S.I. Tumor-derived G-CSF facilitates neoplastic growth through a granulocytic myeloid-derived suppressor cell-dependent mechanism. PLoS ONE 2011, 6, e27690. [Google Scholar] [CrossRef]
- Sagiv, J.Y.; Michaeli, J.; Assi, S.; Mishalian, I.; Kisos, H.; Levy, L.; Damti, P.; Lumbroso, D.; Polyansky, L.; Sionov, R.V.; et al. Phenotypic diversity and plasticity in circulating neutrophil subpopulations in cancer. Cell Rep. 2015, 10, 562–573. [Google Scholar] [CrossRef] [Green Version]
- Granot, Z.; Fridlender, Z.G. Plasticity beyond cancer cells and the “immunosuppressive switch”. Cancer Res. 2015, 75, 4441–4445. [Google Scholar] [CrossRef] [Green Version]
- Rosales, C. Neutrophils at the crossroads of innate and adaptive immunity. J. Leukoc. Biol. 2020, 108, 377–396. [Google Scholar] [CrossRef]
- Mitroulis, I.; Kourtzelis, I.; Kambas, K.; Rafail, S.; Chrysanthopoulou, A.; Speletas, M.; Ritis, K. Regulation of the autophagic machinery in human neutrophils. Eur. J. Immunol. 2010, 40, 1461–1472. [Google Scholar] [CrossRef]
- Levine, B.; Mizushima, N.; Virgin, H.W. Autophagy in immunity and inflammation. Nature 2011, 469, 323–335. [Google Scholar] [CrossRef]
- Stephenson, L.M.; Miller, B.C.; Ng, A.; Eisenberg, J.; Zhao, Z.; Cadwell, K.; Graham, D.B.; Mizushima, N.N.; Xavier, R.; Virgin, H.W.; et al. Identification of Atg5-dependent transcriptional changes and increases in mitochondrial mass in Atg5-deficient T lymphocyte. Autophagy 2009, 5, 625–635. [Google Scholar] [CrossRef] [Green Version]
- Pua, H.H.; Dzhagalov, I.; Chuck, M.; Mizushima, N.; He, Y.W. A critical role for the autophagy gene Atg5 in T cell survival and proliferation. J. Exp. Med. 2007, 204, 25–31. [Google Scholar] [CrossRef]
- Miller, B.C.; Zhao, Z.; Stephenson, L.M.; Cadwell, K.; Pua, H.H.; Lee, H.K.; Mizushima, N.N.; Iwasaki, A.; He, Y.-W.; Swat, W.; et al. The autophagy gene ATG5 plays an essential role in B lymphocyte development. Autophagy 2008, 4, 309–314. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Morgan, M.J.; Chen, K.; Choksi, S.; Liu, Z.G. Induction of autophagy is essential for monocyte-macrophage differentiation. Blood 2012, 119, 2895–2905. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.K.; Mattei, L.M.; Steinberg, B.E.; Alberts, P.; Lee, Y.H.; Chervonsky, A.; Mizushima, N.; Grinstein, S.; Iwasaki, A. In vivo requirement for Atg5 in antigen presentation by dendritic cells. Immunity 2010, 32, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Kundu, M.; Lindsten, T.; Yang, C.-Y.; Wu, J.; Zhao, F.; Zhang, J.; Selak, M.A.; Ney, P.A.; Thompson, C.B. Ulk 1 plays a critical role in the autophagic clearance of mitochondria and ribosomes during reticulocyte maturation. Blood 2008, 112, 1493–1502. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, M.; Ferguson, D.J.P.; Edelmann, M.; Kessler, B.; Morten, K.J.; Komatsu, M.; Simon, A.K. Loss of autophagy in erythroid cells leads to defective removal of mitochondria and severe anemia in vivo. Proc. Natl. Acad. Sci. USA 2010, 107, 832–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozman, S.; Yousefi, S.; Oberson, K.; Kaufmann, T.; Benarafa, C.; Simon, H.U. The generation of neutrophils in the bone marrow is controlled by autophagy. Cell Death Diff. 2015, 22, 445–456. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, A.; Wei, Q.; Shin, J.N.; Fattah, E.A.; Bonilla, D.L.; Xiang, Q.; Eissa, N.T. Autophagy is required for neutrophil-mediated inflammation. Cell Rep. 2015, 12, 1731–1739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, I.; Ritchie, N.D.; Evans, T.J. The interrelationship between phagocytosis, autophagy and formation of neutrophil extracellular traps following infection of human neutrophils by Streptococcus pneumoniae. Innate Immun. 2017, 23, 413–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Zhang, C.; Zou, Z.; Fan, E.K.Y.; Chen, L.; Li, Y.; Billiar, T.R.; Wilson, M.A.; Shi, X.; Fan, J. Aging-related Atg5 defect impairs neutrophil extracellular traps formation. Immunology 2017, 151, 417–432. [Google Scholar] [CrossRef] [Green Version]
- Skendros, P.; Mitroulis, I.; Ritis, K. Autophagy in neutrophils: From granulopoiesis to neutrophil extracellular traps. Front. Cell Dev. Biol. 2018, 6, 109. [Google Scholar] [CrossRef]
- Shrestha, S.; Lee, J.M.; Hong, C.-W. Autophagy in neutrophils. Korean J. Physiol. Pharmacol. 2020, 24, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.J.; Schroder, K. Pattern recognition receptor function in neutrophils. Trends Immunol. 2013, 34, 317–328. [Google Scholar] [CrossRef]
- Cumpelik, A.; Ankli, B.; Zecher, D.; Schifferli, J.A. Neutrophil microvesicles resolve gout by inhibiting C5a-mediated priming of the inflammasome. Ann. Rheum. Dis. 2016, 75, 1236–1245. [Google Scholar] [CrossRef] [Green Version]
- Miyabe, Y.; Miyabe, C.; Mani, V.; Mempel, T.R.; Luster, A.D. Atypical complement receptor C5aR2 transports C5a to initiate neutrophil adhesion and inflammation. Sci. Immunol. 2019, 4, eaav5951. [Google Scholar] [CrossRef] [PubMed]
- Van Rees, D.J.; Szilagyi, K.; Kuijpers, T.W.; Matlung, H.L.; van den Berg, T.K. Immunoreceptors on neutrophils. Sem. Immunol. 2016, 28, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N. Pro-resolving lipid mediators are leads for resolution physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dancey, J.T.; Deubelbeiss, K.A.; Harker, L.A.; Finch, C.A. Neutrophil kinetics in man. J. Clin. Investig. 1976, 58, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.; Hodgson, G.; Katz, M.; Dunn, A.R. Evaluation of role of G-CSF in the production, survival, and release of neutrophils from bone marrow into circulation. Blood 2002, 100, 854–861. [Google Scholar] [CrossRef] [Green Version]
- Pillay, J.; den Braber, I.; Vrisekoop, N.; Kwast, L.M.; de Boer, R.J.; Borghans, J.A.M.; Tesselaar, K.; Koenderman, L. In vivo labeling with 2H2O reveals a human neutrophil lifespan of 5.4 days. Blood 2010, 116, 625–627. [Google Scholar] [CrossRef]
- Simon, H.-U. Neutrophil apoptosis pathways and their modifications in inflammation. Immunol. Rev. 2003, 193, 101–110. [Google Scholar] [CrossRef]
- Lopez, A.F.; Williamson, D.J.; Gamble, J.R.; Begley, C.G.; Harlan, J.M.; Klebanoff, S.J.; Waltersdorph, A.; Wong, G.; Clark, S.C.; Vadas, M.A. Recombinant human granulocyte-macrophage colony-stimulating factor stimulates in vitro mature human neutrophil and eosinophil function, surface receptor expression, and survival. J. Clin. Investig. 1986, 78, 1220–1228. [Google Scholar] [CrossRef]
- Begley, C.G.; Lopez, A.F.; Nicola, N.A.; Warren, D.J.; Vadas, M.A.; Sanderson, C.J.; Metcalf, D. Purified colony-stimulating factors enhance the survival of human neutrophils and eosinophils in vitro: A rapid and sensitive microassay for colony-stimulating factors. Blood 1986, 68, 162–166. [Google Scholar] [CrossRef] [Green Version]
- Brach, M.A.; de Vos, S.; Gruss, H.J.; Herrmann, F. Prolongation of survival of human polymorphonuclear neutrophils by granulocyte-macrophage colony-stimulating factor is caused by inhibition of programmed cell death. Blood 1992, 80, 2920–2924. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, C.; Yoshida, S.; Taniguchi, H.; Qin, M.H.; Miyamoto, H.; Mizuguchi, Y. Lipopolysaccharide and granulocyte colony-stimulating factor delay neutrophil apoptosis and ingestion by guinea pig macrophages. Infect. Immun. 1993, 61, 1972–1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pericle, F.; Liu, J.H.; Diaz, J.I.; Blanchard, D.K.; Wei, S.; Forni, G.; Djeu, J.Y. Interleukin-2 prevention of apoptosis in human neutrophils. Eur. J. Immunol. 1994, 24, 440–444. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, S.C.; Huang, M.-H.; Tsai, C.-Y.; Tsai, Y.-Y.; Tsai, S.-T.; Sun, K.-H.; Yu, H.-S.; Han, S.-H.; Yu, C.-L. The expression of genes modulating programmed cell death in normal human polymorphonuclear neutrophils. Biochem. Biophys. Res. Commun. 1997, 233, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Savill, J.; Dransfield, I.; Gregory, C.; Haslett, C. A blast from the past: Clearance of apoptotic cells regulates immune responses. Nat. Rev. Immunol. 2002, 2, 965–975. [Google Scholar] [CrossRef]
- Luo, H.R.; Loison, F. Constitutive neutrophil apoptosis: Mechanisms and regulation. Am. J. Hematol. 2008, 83, 288–295. [Google Scholar] [CrossRef]
- Edwards, S.W.; Derouet, M.; Howse, M.; Moots, R.J. Regulation of neutrophil apoptosis by Mcl-1. Biochem. Soc. Transac. 2004, 32, 489–492. [Google Scholar] [CrossRef] [Green Version]
- Dzhagalov, I.; St. John, A.; He, Y.-W. The antiapoptotic protein Mcl-1 is essential for the survival of neutrophils but not macrophages. Blood 2007, 109, 1620–1626. [Google Scholar] [CrossRef] [Green Version]
- Reed, J.C. Proapoptotic multidomain Bcl-2/Bax-family proteins: Mechanisms, physiological roles, and therapeutic opportunities. Cell Death Differ. 2006, 13, 1378–1386. [Google Scholar] [CrossRef] [Green Version]
- Brenner, D.; Mak, T.W. Mitochondrial cell death effectors. Curr. Opin. Cell Biol. 2009, 21, 871–877. [Google Scholar] [CrossRef]
- Milot, E.; Filep, J.G. Regulation of neutrophil survival/apoptosis by Mcl-1. Sci. World J. 2011, 11, 1948–1962. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, A.D.; DeLeo, F.R. Neutrophil apoptosis and the resolution of infection. Immunol. Res. 2009, 43, 25–61. [Google Scholar] [CrossRef] [PubMed]
- Noseykina, E.M.; Schepetkin, I.A.; Atochin, D.N. Molecular mechanisms for regulation of neutrophil apoptosis under normal and pathological conditions. J. Evol. Biochem. Physiol. 2021, 57, 429–450. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.D.; Malachowa, N.; DeLeo, F.R. Influence of microbes on neutrophil life and death. Front. Cell. Inf. Microbiol. 2017, 7, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudo, C.; Yamashita, T.; Araki, A.; Terashita, M.; Watanabe, T.; Atsumi, M.; Tamura, M.; Sendo, F. Modulation of in vivo immune response by selective depletion of neutrophils using a monoclonal antibody, RP-3, I. Inhibition by RP-3 treatment of the priming and effector phases of delayed-type hypersensitivity to sheep red cells in rats. J. Immunol. 1993, 150, 3728–3738. [Google Scholar]
- Tamura, M.; Sekiya, S.; Terashita, M.; Sendo, F. Modulation of the in vivo immune response by selective depletion of neutrophils using a monoclonal antibody, RP-3, III. Enhancement by RP-3 treatment of the anti-sheep red blood cell plaque-forming cell response in rats. J. Immunol. 1994, 153, 1301–1308. [Google Scholar]
- Matsuzaki, J.; Tsuji, T.; Chamoto, K.; Takeshima, T.; Sendo, F.; Nishimura, T. Successful elimination of memory-type CD8+ T cell subsets by the administration of anti-Gr-1 monoclonal antibody in vivo. Cell. Immunol. 2003, 224, 98–105. [Google Scholar] [CrossRef]
- Yue, C.L.; Tanimoto, K.; Horiuchi, Y. Characterization and possible mechanisms of mitogen-induced cell-mediated cytotoxicity. Scand. J. Immunol. 1981, 14, 397–408. [Google Scholar] [CrossRef]
- Dallegri, F.; Frumento, G.; Maggi, A.; Patrone, F. PHA-induced neutrophil-mediated cytotoxicity. J. Clin. Lab. Immunol. 1983, 11, 203–206. [Google Scholar]
- Stiehm, E.R.; Roberts, R.L.; Ank, B.J.; Plaeger-Marshall, S.; Salman, N.; Shen, L.; Fanger, M.W. Comparison of cytotoxic properties of neonatal and adult neutrophils and monocytes and enhancement by cytokines. Clin. Diagn. Lab. Immunol. 1994, 1, 342–347. [Google Scholar] [CrossRef]
- Okuda, T. Murine polymorphonuclear leukocytes synthesize and secrete the third component and factor B of complement. Int. Immunol. 1991, 3, 293–296. [Google Scholar] [CrossRef]
- Botto, M.; Lissandrini, D.; Sorio, C.; Walport, M.J. Biosynthesis and secretion of complement component (C3) by activated human polymorphonuclear leukocytes. J. Immunol. 1992, 149, 1348–1355. [Google Scholar] [PubMed]
- Yu, C.L.; Tsai, C.Y.; Hsieh, S.C.; Tsai, Y.Y.; Tsai, S.T.; Sun, K.H.; Yu, H.S.; Han, S.H. Production of the third component of complement (C3) by peripheral polymorphonuclear neutrophils of the patients with rheumatoid arthritis. Proc. Natl. Sci. Counc. China B 1995, 19, 225–232. [Google Scholar]
- Bedouhène, S.; Dang, P.M.-C.; Hurtado-Nedelec, M.; El-Benna, J. Neutrophil degranulation of azurophil and specific granules. Methods Mol. Biol. 2020, 2087, 215–222. [Google Scholar] [PubMed]
- Gigon, L.; Yousefi, S.; Karaulov, A.; Simon, H.-U. Mechanisms of toxicity mediated by neutrophil and eosinophil granule proteins. Allergol. Int. 2021, 70, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Li, K.-J.; Lu, M.-C.; Hsieh, S.-C.; Wu, C.-H.; Yu, H.-S.; Tsai, C.-Y.; Yu, C.-L. Release of surface-expressed lactoferrin from polymorphonuclear neutrophils after contact with CD4+ T cells and its modulation on Th1/Th2 cytokine production. J. Leukoc. Biol. 2006, 80, 350–358. [Google Scholar] [CrossRef]
- Bazzoni, F.; Cassatella, M.A.; Laudanna, C.; Rossi, F. Phagocytosis of opsonized yeast induces tumor necrosis factor-alpha mRNA accumulation and protein release by human polymorphonuclear leukocytes. J. Leukoc. Biol. 1991, 50, 223–228. [Google Scholar] [CrossRef]
- Cassatella, M.A. The production of cytokines by polymorphonuclear neutrophils. Immunol. Today 1995, 16, 21–26. [Google Scholar] [CrossRef]
- Yoshimura, A.; Hara, Y.; Kaneko, T.; Kato, I. Secretion of IL-1β, TNF-α, IL-8, and IL-1ra by human polymorphonuclear leukocytes in response to lipopolysaccharides from periodontopathic bacteria. J. Periodont. Res. 1997, 32, 279–286. [Google Scholar] [CrossRef]
- Tecchio, C.; Micheletti, A.; Cassatella, M.A. Neutrophil-derived cytokines: Facts beyond expression. Front. Immunol. 2014, 5, 508. [Google Scholar] [CrossRef] [Green Version]
- Tecchio, C.; Cassatella, M.A. Neutrophil-derived chemokines on the road to immunity. Semin. Immunol. 2016, 28, 119–128. [Google Scholar] [CrossRef]
- Tamassia, N.; Bianchetto-Aguilera, F.; Arruda-Silva, F.; Gardiman, E.; Gasperini, S.; Calzetti, F.; Cassatella, M.A. Cytokine production by human neutrophils: Revisiting the “dark side of the moon”. Eur. J. Clin. Investig. 2018, 48 (Suppl. S2), e12952. [Google Scholar] [CrossRef] [PubMed]
- Butin-Israeli, V.; Houser, M.C.; Feng, M.; Thorp, E.B.; Nusrat, A.; Parkos, C.A.; Sumagin, R. Deposition of microparticles by neutrophils onto inflamed epithelium: A new mechanism to disrupt epithelial intercellular adhesions and promote transepithelial migration. FASEB J. 2016, 30, 4007–4020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasser, O.; Schifferli, J.A. Activated polymorphonuclear neutrophils disseminate anti-inflammatory microparticles by ectocytosis. Blood 2004, 104, 2543–2548. [Google Scholar] [CrossRef] [PubMed]
- Finkielsztein, A.; Mascarenhas, L.; Butin-Israeli, V.; Sumagin, R. Isolation and characterization of neutrophil-derived microparticles for functional studies. J. Vis. Exp. 2018, 133, e56949. [Google Scholar] [CrossRef]
- Takashima, A.; Yao, Y. Neutrophil plasticity: Acquisition of phenotype and functionality of antigen-presenting cell. J. Leukoc. Biol. 2015, 98, 489–496. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.; Loré, K. Granulocytes: New members of the antigen presenting cell family. Front. Immunol. 2017, 8, 1781. [Google Scholar] [CrossRef] [Green Version]
- Vono, M.; Lin, A.; Norrby-Teglund, A.; Koup, R.A.; Liang, F.; Loré, K. Neutrophils acquire the capacity for antigen presentation to memory CD4+ T cells in vitro and ex vivo. Blood 2017, 129, 1991–2001. [Google Scholar] [CrossRef] [Green Version]
- Meinderts, S.M.; Baker, G.; van Wijk, S.; Beuger, B.M.; Geissler, J.; Jansen, M.H.; Saris, A.; Brinke, A.T.; Kuijpers, T.W.; van den Berg, T.K.; et al. Neutrophils acquire antigen-presenting cell features after phagocytosis of IgG-opsonized erythrocytes. Blood Adv. 2019, 3, 1761–1773. [Google Scholar] [CrossRef]
- Polak, D.; Hafner, C.; Briza, P.; Kitzmüller, C.; Elbe-Bürger, A.; Samadi, N.; Gschwandtner, M.; Pfützner, W.; Zlabinger, G.J.; Jahn-Schmid, B.; et al. A novel role for neutrophils in IgE-mediated allergy: Evidence for antigen presentation in late-phase reactions. J. Allergy Clin. Immunol. 2019, 143, 1143–1152.e4. [Google Scholar] [CrossRef] [Green Version]
- Samadi, N.; Polak, D.; Kitzmüller, C.; Steinberger, P.; Zlabinger, G.J.; Jahn-Schmid, B.; Bohle, B. T-cell-derived cytokines enhance the antigen-presenting capacity of human neutrophils. Eur. J. Immunol. 2019, 49, 1441–1443. [Google Scholar] [CrossRef]
- Li, K.-J.; Wu, C.-H.; Lu, C.-H.; Shen, C.-Y.; Kuo, Y.-M.; Tsai, C.-Y.; Hsieh, S.-C.; Yu, C.-L. Trogocytosis between non-immune cells for cell clearance, and among immune-related cells for modulating immune responses and autoimmunity. Int. J. Mol. Sci. 2021, 22, 2236. [Google Scholar] [CrossRef] [PubMed]
- Dance, A. Core concept: Cells nibble one another via the under-appreciated process of trogocytosis. Proc. Natl. Acad. Sci. USA 2019, 116, 17608–17610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, R.P.; Lindorfer, M.A. Fcγ-receptor-mediated trogocytosis impacts mAb-based therapies: Historical precedence and recent developments. Blood 2015, 125, 762–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valgardsdottir, R.; Cattaneo, I.; Klein, C.; Introna, M.; Figliuzzi, M.; Golay, J. Human neutrophils mediate trogocytosis rather than phagocytosis of CLL B cells opsonized with anti-CD20 antibodies. Blood 2017, 129, 2636–2644. [Google Scholar] [CrossRef] [Green Version]
- Poupot, M.; Fournié, J.-J. Spontaneous membrane transfer through homotypic synapses between lymphoma cells. J. Immunol. 2003, 171, 2517–2523. [Google Scholar] [CrossRef] [Green Version]
- Honer, H.; Frank, C.; Dechant, C.; Repp, R.; Glennie, M.; Herrmann, M. Intimate cell conjugate formation and exchange of membrane lipids procede apoptosis induction in target cells during antibody-dependent, granulocyte-mediated cytotoxicity. J. Immunol. 2007, 179, 337–345. [Google Scholar] [CrossRef] [Green Version]
- Li, K.-J.; Wu, C.-H.; Shen, C.-Y.; Kuo, Y.-M.; Yu, C.-L.; Hsieh, S.-C. Membrane transfer from mononuclear cells to polymorphonuclear neutrophils transduces cell survival and activation signals in the recipient cells via anti-extrinsic apoptotic and MAP kinase signaling pathways. PLoS ONE 2016, 11, e0156262. [Google Scholar] [CrossRef] [Green Version]
- Hudrisier, D.; Riond, J.; Mazarguil, H.; Gairin, J.E.; Joly, E. Cutting edge: CTLs rapidly capture membrane fragments from target cells in a TCR signaling-dependent manner. J. Immunol. 2001, 166, 3645–3649. [Google Scholar] [CrossRef] [Green Version]
- Batista, F.D.; Iber, D.; Neuberger, M.S. B cells acquire antigen from target cells after synapse formation. Nature 2001, 411, 489–494. [Google Scholar] [CrossRef]
- Dömer, D.; Walther, T.; Möller, S.; Behnen, M.; Laskay, T. Neutrophil extracellular traps activate proinflammatory functions of human neutrophils. Front. Immunol. 2021, 12, 636954. [Google Scholar] [CrossRef]
- Douglas, R.G.; Alford, R.H.; Cate, T.R.; Couch, R.B. The leukocyte response during viral respiratory illness in man. Ann. Intern. Med. 1966, 64, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Faden, H.; Hong, J.J.; Ogra, P.L. Interaction of polymorphonuclear leukocytes and viruses in humans: Adherence of polymorphonuclear leukocytes to respiratory syncytial virus-infected cells. J. Virol. 1984, 52, 16–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacGregor, R.R.; Friedman, H.M.; Macarak, E.J.; Kefalides, N.A. Virus infection of endothelial cells increases granulocyte adherence. J. Clin. Investig. 1980, 65, 1469–1477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratcliffe, D.R.; Nolin, S.L.; Cramer, E.B. Neutrophil interaction with influenza-infected epithelial cells. Blood 1988, 72, 142–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouse, B.T.; Babiuk, L.A.; Henson, P.M. Neutrophils in antiviral immunity: Inhibition of virus replication by a mediator produced by bovine neutrophils. J. Infect. Dis. 1980, 141, 223–232. [Google Scholar] [CrossRef]
- Tumpey, T.M.; Chen, S.-H.; Oakes, J.E.; Lausch, R.N. Neutrophil-mediated suppression of virus replication after herpes simplex virus type 1 infection of the murine cornea. J. Virol. 1996, 70, 898–904. [Google Scholar] [CrossRef] [Green Version]
- Tumpey, T.M.; García-Sastre, A.; Taubenberger, J.K.; Palese, P.; Swayne, D.E.; Pantin-Jackwood, M.J.; Schultz-Cherry, S.; Solórzano, A.; Van Rooijen, N.; Katz, J.M.; et al. Pathogenicity of influenza viruses with genes from the 1918 pandemic virus: Functional roles of alveolar macrophages and neutrophils in limiting virus replication and mortality in mice. J. Virol. 2005, 79, 14933–14944. [Google Scholar] [CrossRef] [Green Version]
- Tate, M.D.; Brooks, A.G.; Reading, P.C. The role of neutrophils in the upper and lower respiratory tract during influenza virus infection of mice. Respir. Res. 2008, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Wojtasiak, M.; Pickett, D.L.; Tate, M.D.; Londrigan, S.L.; Bedoui, S.; Brooks, A.G.; Reading, P.C. Depletion of Gr-1+, but not Ly6G+, immune cells exacerbates virus replication and disease in an intranasal model of herpes simplex virus type 1 infection. J. Gen. Virol. 2010, 91, 2158–2166. [Google Scholar] [CrossRef]
- Tate, M.D.; Brooks, A.G.; Reading, P.C.; Mintern, J.D. Neutrophils sustain effective CD8+ T-cell responses in the respiratory tract following influenza infection. Immunol. Cell Biol. 2012, 90, 197–205. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, Y.; Zhu, L. Role of neutrophils in acute viral infection. Immun. Inflamm. Dis. 2021, 9, 1186–1196. [Google Scholar] [CrossRef] [PubMed]
- Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Al-hussaniy, H.A.; Al-Harcan, N.A.H.; Alexiou, A.; Batiha, G.E.-S. Neutrophil extracellular traps (NETs) and COVID-19: A new frontiers for therapeutic modality. Int. Immunopharmacol. 2022, 104, 108516. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, F.; Means, T.K.; Luster, A.D. Toll-like receptors stimulate human neutrophil function. Blood 2003, 102, 2660–2669. [Google Scholar] [CrossRef] [PubMed]
- Witko-Sarsat, V.; Rieu, P.; Descamps-Latscha, B.; Lesavre, P.; Halbwachs-Mecarelli, L. Neutrophils: Molecules, functions and pathophysiological aspects. Lab. Investig. 2000, 80, 617–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, S.C.; Tsai, C.Y.; Sun, K.H.; Yu, H.S.; Tsai, S.T.; Wang, J.C.; Tsai, Y.Y.; Han, S.H.; Yu, C.L. Decreased spontaneous and lipopolysaccharide stimulated production of interleukin 8 by polymorphonuclear neutrophils of patients with active systemic lupus erythematosus. Clin. Exp. Rheumatol. 1994, 12, 627–633. [Google Scholar] [PubMed]
- Hsieh, S.C.; Wu, T.-H.; Tsai, C.-Y.; Li, K.-J.; Lu, M.-C.; Wu, C.-H.; Yu, C.-L. Abnormal in vitro CXCR2 modulation and defective cationic ion transporter expression on polymorphonuclear neutrophils responsible for hyporesponsiveness to IL-8 stimulation in patients with active systemic lupus erythematosus. Rheumatology 2008, 47, 150–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, S.C.; Tsai, C.Y.; Sun, K.H.; Tsai, Y.Y.; Tsai, S.T.; Han, S.H.; Yu, H.S.; Yu, C.L. Defective spontaneous and bacterial lipopolysaccharide-stimulated production of interleukin-1 receptor antagonist by polymorphonuclear neutrophils of patients with active systemic lupus erythematosus. Br. J. Rheumatol. 1995, 34, 107–112. [Google Scholar] [CrossRef]
- Yu, C.-L.; Sun, K.-H.; Tsai, C.Y.; Tsai, Y.-Y.; Tsai, S.-T.; Huang, D.-F.; Han, S.-H.; Yu, H.-S. Expression of Th1/Th2 cytokine mRNA in peritoneal exudative polymorphonuclear neutrophils and their effects on mononuclear Th1/Th2 cytokine production in MRL-lpr/lpr mice. Immunology 1998, 95, 480–487. [Google Scholar] [CrossRef]
- Tsai, C.-Y.; Wu, T.-H.; Yu, C.-L.; Tsai, Y.-Y.; Chou, C.-T. Decreased IL-12 production by polymorphonuclear leukocytes in patients with active systemic lupus erythematosus. Immunol. Investig. 2002, 31, 177–189. [Google Scholar] [CrossRef]
- Tsai, C.-Y.; Li, K.-J.; Hsieh, S.-C.; Liao, H.-T.; Yu, C.-L. What’s wrong with neutrophils in lupus? Clin. Exp. Rheumatol. 2019, 37, 684–693. [Google Scholar]
- Stein, J.M.; Luzio, J.P. Ectocytosis caused by sublytic autologous complement attack on human neutrophils. The sorting of endogenous plasma-membrane proteins and lipids into shed vesicles. Biochem. J. 1991, 274 Pt 2, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Hess, C.; Sadallah, S.; Hefti, A.; Landmann, R.; Schifferli, J.A. Ectosomes released by human neutrophils are specialized functional units. J. Immunol. 1999, 163, 4564–4573. [Google Scholar] [CrossRef]
- Johnstone, R.M. Exosomes biological significance: A concise review. Blood Cells Mol. Dis. 2006, 36, 315–321. [Google Scholar] [CrossRef]
- Sadallah, S.; Eken, C.; Schifferli, J.A. Ectosomes as modulators of inflammation and immunity. Clin. Exp. Immunol. 2011, 163, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Eken, C.; Gasser, O.; Zenhaeusern, G.; Oehri, I.; Hess, C.; Schifferli, J.A. Polymorphonuclear neutrophil-derived ectosomes interfere with the maturation of monocyte-derived dendritic cells. J. Immunol. 2008, 180, 817–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eken, C.; Martin, P.J.; Sadallah, S.; Treves, S.; Schaller, M.; Schifferli, J.A. Ectosomes released by polymorphonuclear neutrophils induce a MerTK-dependent anti-inflammatory pathway in macrophages. J. Biol. Chem. 2010, 285, 39914–39921. [Google Scholar] [CrossRef] [Green Version]
- Eken, C.; Sadallah, S.; Martin, P.J.; Treves, S.; Schifferli, J.A. Ectosomes of polymorphonuclear neutrophils activate multiple signaling pathways in macrophages. Immunobiology 2013, 218, 382–392. [Google Scholar] [CrossRef]
- Rawat, K.; Syeda, S.; Shrivastava, A. Neutrophil-derived granule cargoes: Paving the way for tumor growth and progression. Cancer Metastasis Rev. 2021, 40, 221–244. [Google Scholar] [CrossRef]
- Zhu, T.; Wang, Y.; Jin, H.; Li, L. The role of exosome in autoimmune connective tissue disease. Ann. Med. 2019, 51, 101–108. [Google Scholar] [CrossRef]
- Mirzaei, R.; Zamani, F.; Hajibaba, M.; Rasouli-Saravani, A.; Noroozbeygi, M.; Gorgani, M.; Hosseini-Fard, S.R.; Jalalifar, S.; Ajdarkosh, H.; Abedi, S.H.; et al. The pathogenic therapeutic and diagnostic role of exosomal microRNA in the autoimmune diseases. J. Neuroimmunol. 2021, 358, 577640. [Google Scholar] [CrossRef]
- Miao, C.; Wang, X.; Zhou, W.; Huang, J. The emerging roles of exosomes in autoimmune diseases, with special emphasis on microRNAs in exosomes. Pharmacol. Res. 2021, 169, 105680. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-Y.; Hsieh, S.-C.; Lu, C.-S.; Wu, T.-H.; Liao, H.-T.; Wu, C.-H.; Li, K.-J.; Kuo, Y.-M.; Lee, H.-T.; Shen, C.-Y.; et al. Cross-talk between mitochondrial dysfunction-provoked oxidative stress and aberrant noncoding RNA expression in the pathogenesis and pathophysiology of SLE. Int. J. Mol. Sci. 2019, 20, 5183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.Y.; Shen, C.-Y.; Liu, C.-W.; Hsieh, S.-C.; Liao, H.-T.; Li, K.-J.; Lu, C.-S.; Lee, H.-T.; Lin, C.-S.; Wu, C.-H.; et al. Aberrant non-coding RNA expression in patients with systemic lupus erythematosus: Consequences for immune dysfunctions and tissue damage. Biomolecules 2020, 10, 1641. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-Y.; Hsieh, S.-C.; Liu, C.-W.; Lu, C.-H.; Liao, H.-T.; Chen, M.-H.; Li, K.-J.; Wu, C.-H.; Shen, C.-Y.; Kuo, Y.-M.; et al. The expression of non-coding RNAs and their target molecules in rheumatoid arthritis: A molecular basis for rheumatoid pathogenesis and its potential clinical applications. Int. J. Mol. Sci. 2021, 22, 5689. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-Y.; Hsieh, S.-C.; Wu, T.-H.; Li, K.-J.; Shen, C.-Y.; Liao, H.-T.; Wu, C.H.; Kuo, Y.-M.; Lu, C.-S.; Yu, C.-L. Pathogenic roles of autoantibodies and aberrant epigenetic regulation of immune and connective tissue cells in the tissue fibroosis of patients with systemic sclerosis. Int. J. Mol. Sci. 2020, 21, 3069. [Google Scholar] [CrossRef]
- Tsai, C.-Y.; Hsieh, S.-C.; Liu, C.-W.; Lu, C.-S.; Wu, C.-H.; Liao, H.-T.; Chen, M.-H.; Li, K.-J.; Shen, C.-Y.; Kuo, Y.-M.; et al. Cross-talk among polymorphonuclear neutrophils, immune and non-immune cells via released cytokines, granule proteins, microvesicles, and neutrophil extracellular trap formation: A novel concept of biology and pathobiology for neutrophils. Int. J. Mol. Sci. 2021, 22, 3119. [Google Scholar] [CrossRef]
- Li, L.; Zuo, X.; Xiao, Y.; Liu, D.; Luo, H.; Zhu, H. Neutrophil-derived exosome from systemic sclerosis inhibits the proliferation and migration of endothelial cells. Biochem. Biophys. Res. Commun. 2020, 526, 334–340. [Google Scholar] [CrossRef]
- Li, L.; Zuo, X.; Liu, D.; Luo, H.; Zhu, H. The functional roles of RNAs cargoes released by neutrophil-derived exosomes in dermatomyositis. Front. Pharmacol. 2021, 12, 727901. [Google Scholar] [CrossRef]
- Oehler, L.; Majdic, O.; Pickl, W.F.; Stockl, J.; Riedl, E.; Drach, J.; Rappersberger, K.; Geissler, K.; Knapp, W. Neutrophil granulocyte-committed cells can be driven to acquire dendritic cell characteristics. J. Exp. Med. 1998, 187, 1019–1028. [Google Scholar] [CrossRef] [Green Version]
- Sandilands, G.P.; McCrae, J.; Hill, K.; Perry, M.; Baxter, D. Major histocompatibility complex class II (DR) antigen and costimulatory molecules on in vitro and in vivo activated human polymorphonuclear neutrophils. Immunology 2006, 119, 562–571. [Google Scholar] [CrossRef]
- Mercer, F.; Ng, S.H.; Brown, T.M.; Boatman, G.; Johnson, P.J. Neutrophils kill the parasite Trichomonas vaginalis using trogocytosis. PLoS Biol. 2018, 16, e2003885. [Google Scholar] [CrossRef] [PubMed]
- Olivera-Valle, I.; Latorre, M.C.; Calvo, M.; Gaspar, B.; Gómez-Oro, C.; Collazos, A.; Breton, A.; Caballero-Campo, P.; Ardoy, M.; Asensio, F.; et al. Vaginal neutrophils eliminate sperm by trogocytosis. Hum. Reprod. 2020, 35, 2567–2578. [Google Scholar] [CrossRef] [PubMed]
- Vanherberghen, B.; Andersson, K.; Carlin, L.M.; Nolte-‘t Hoen, E.N.M.; Williams, G.S.; Höglund, P.; Davis, D.M. Human and murine inhibitory natural killer cell receptors transfer from natural killer cells to target cells. Proc. Natl. Acad. Sci. USA 2004, 101, 16873–16878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, T.; Mero, P.; Booth, J.W. Dynamics of macrophage trogocytosis of rituximab-coated B cells. PLoS ONE 2001, 6, e14498. [Google Scholar] [CrossRef] [Green Version]
- Miyake, K.; Shiozawa, N.; Nagao, T.; Yoshikawa, S.; Yamanishi, Y.; Karasuyama, H. Trogocytosis of peptide-MHC class II complexes from dendritic cells confers antigen-presenting ability on basophils. Proc. Natl. Acad. Sci. USA 2017, 114, 1111–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliphant, C.J.; Hwang, Y.Y.; Walker, J.A.; Salimi, M.; Wong, S.H.; Brewer, J.M.; Englezakis, A.; Barlow, J.L.; Hams, E.; Scanlon, S.T.; et al. MHC II-mediated dialog between group 2 innate lymphoid cells and CD4+ T cells potentiates type 2 immunity and promotes parasitic helminth expulsion. Immunity 2014, 41, 283–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyake, K.; Karasuyama, H. The role of trogocytosis in the modulation of immune cell functions. Cells 2021, 10, 1255. [Google Scholar] [CrossRef]
- Uribe-Querol, E.; Rosales, C. The multiple roles of trogocytosis in immunity, the nervous system and development. BioMed. Res. Int. 2021, 2021, 1601565. [Google Scholar] [CrossRef]
- Nakayama, M.; Hori, A.; Toyoura, S.; Yamaguchi, S.-I. Shaping of T cell functions by trogocytosis. Cells 2021, 10, 1155. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Ribon, M.; Seninet, S.; Mussard, J.; Sebbag, M.; Clavel, C.; Serre, G.; Boissier, M.-C.; Semerano, L.; Decker, P. Neutrophil extracellular traps exert both pro- and anti-inflammatory actions in rheumatoid arthritis that are modulated by C1q and LL-37. J. Autoimmun. 2019, 98, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Döring, Y.; Libby, P.; Soehnlein, O. Neutrophil extracellular traps participate in cardiovascular diseases: Recent experimental and clinical insights. Circ. Res. 2020, 126, 1228–1241. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Tao, W.; Shen, F.; Du, W.; Xu, Z.; Liu, Z. The emerging role of neutrophil extracellular traps in arterial, venous and cancer-associated thrombosis. Front. Cardiovasc. Med. 2021, 8, 786387. [Google Scholar] [CrossRef] [PubMed]
- Klopf, J.; Brostjan, C.; Eilenberg, W.; Neumayer, C. Neutrophil extracellular traps and their implications in cardiovascular and inflammatory disease. Int. J. Mol. Sci. 2021, 22, 559. [Google Scholar] [CrossRef]
- Leffler, J.; Martin, M.; Gullstrand, B.; Tydén, H.; Lood, C.; Truedsson, L.; Bengtsson, A.A.; Blom, A.M. Neutrophil extracellular traps that are not degraded in systemic lupus erythematosus activate complement exacerbating the disease. J. Immunol. 2012, 188, 3522–3531. [Google Scholar] [CrossRef] [Green Version]
- Söderberg, D.; Segelmark, M. Neutrophil extracellular traps in ANCA-associated vasculitis. Front. Immunol. 2016, 7, 256. [Google Scholar] [CrossRef] [Green Version]
- Masucci, M.T.; Minopoli, M.; Del Vecchio, S.; Carriero, M.V. The emerging role of neutrophil extracellular traps (NETs) in tumor progression and metastasis. Front. Immunol. 2020, 11, 1749. [Google Scholar] [CrossRef]
- Hazeldine, J.; Harris, P.; Chapple, I.L.; Grant, M.; Greenwood, H.; Livesey, A.; Sapey, E.; Lord, J.M. Impaired neutrophil extracellular trap formation: A novel defect in the innate immune system of aged individuals. Aging Cell 2014, 13, 690–698. [Google Scholar] [CrossRef]
- Lu, C.-H.; Li, K.-J.; Wu, C.-H.; Shen, C.-Y.; Kuo, Y.-M.; Hsieh, S.-C.; Yu, C.-L. The FcγRIII engagement augments PMA-stimulated neutrophil extracellular traps (NETs) formation by granulocytes partially via cross-talk between Syk-ERK-NF-κB and PKC-ROS signaling pathways. Biomedicines 2021, 9, 1127. [Google Scholar] [CrossRef]
- Farrera, C.; Fadeel, B. Macrophage clearance of neutrophil extracellular traps is a silent process. J. Immunol. 2013, 191, 2647–2656. [Google Scholar] [CrossRef] [Green Version]
- Eruslanov, E.B.; Bhojnagarwala, P.S.; Quatromoni, J.G.; Stephen, T.L.; Ranganathan, A.; Deshpande, C.; Akimova, T.; Vachani, A.; Litzky, L.; Hancock, W.W.; et al. Tumor-associated neutrophils stimulate T cell responses in early-stage human lung cancer. J. Clin. Investig. 2014, 124, 5466–5480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Duckworth, C.A.; Zhao, Q.; Pritchard, D.M.; Rhodes, J.M.; Yu, L.-G. Increased circulation of galectin-3 in cancer induces secretion of metastasis-promoting cytokines from blood vascular endothelium. Clin. Cancer Res. 2013, 19, 1693–1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Aparicio, M.; Alfaro, C. Significance of the IL-8 pathway for immunotherapy. Hum. Vaccines Immunother. 2020, 16, 2312–2317. [Google Scholar] [CrossRef] [PubMed]
- Furumaya, C.; Martinez-Sanz, P.; Bouti, P.; Kuijpers, T.W.; Matlung, H.L. Plasticity in pro- and anti-tumor activity of neutrophils: Shifting the balance. Front. Immunol. 2020, 11, 2100. [Google Scholar] [CrossRef] [PubMed]
- Avtenyuk, N.U.; Visser, N.; Bremer, E.; Wiersma, V.R. The neutrophil: The underdog that packs a punch in the fight against cancer. Int. J. Mol. Sci. 2020, 21, 7820. [Google Scholar] [CrossRef]
- Andzinski, L.; Kasnitz, N.; Stahnke, S.; Wu, C.-F.; Gereke, M.; von Köckritz-Blickwede, M.; Schilling, B.; Brandau, S.; Weiss, S.; Jablonska, J. Type I IFNs induce anti-tumor polarization of tumor associated neutrophils in mice and human. Int. J. Cancer 2016, 138, 1982–1993. [Google Scholar] [CrossRef]
- Fridlender, Z.G.; Sun, J.; Kim, S.; Kapoor, V.; Cheng, G.; Ling, L.; Worthen, G.S.; Albelda, S.M. Polarization of tumor-associated neutrophil phenotype by TGF-β: “N1” versus “N2” TAN. Cancer Cell 2009, 16, 183–194. [Google Scholar] [CrossRef] [Green Version]
- Shaul, M.E.; Levy, L.; Sun, J.; Mishalian, I.; Singhal, S.; Kapoor, V.; Horng, W.; Fridlender, G.; Albelda, S.M.; Fridlender, Z.G. Tumor-associated neutrophils display a distinct N1 profile following TGFβ modulation: A transcriptomics analysis of pro- vs. antitumor TANs. Oncoimmunology 2016, 5, e1232221. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.Y.; Tai, J.A.; Li, S.; Nishikawa, T.; Kaneda, Y. Virus-stimulated neutrophils in the tumor microenvironment enhance T cell-mediated anti-tumor immunity. Oncotarget 2016, 7, 42195–42207. [Google Scholar] [CrossRef]
- Heemskerk, N.; van Egmond, M. Monoclonal antibody-mediated killing of tumour cells by neutrophils. Eur. J. Clin. Investig. 2018, 48 (Suppl. S2), e12962. [Google Scholar] [CrossRef] [Green Version]
- Matlung, H.L.; Babes, L.; Zhao, X.W.; van Houdt, M.; Treffers, L.W.; van Rees, D.J.; Franke, K.; Schornagel, K.; Verkuijlen, P.; Janssen, H.; et al. Neutrophils kill antibody-opsonized cancer cells by trogoptosis. Cell Rep. 2018, 23, 3946–3959.e6. [Google Scholar] [CrossRef] [PubMed]
- Brandsma, A.M.; Bondza, S.; Evers, M.; Koutstaal, R.; Nederend, M.; Marco Jansen, J.H.; Rösner, T.; Valerius, T.; Leusen, J.H.W.; Ten Broeke, T. Potent Fc receptor signaling by IgA leads to superior killing of cancer cells by neutrophils compared to IgG. Front. Immunol. 2019, 10, 704. [Google Scholar] [CrossRef] [Green Version]
- Treffers, L.W.; van Houdt, M.; Bruggeman, C.W.; Heineke, M.H.; Zhao, X.W.; van der Heijden, J.; Nagelkerke, S.Q.; Verkuijlen, P.J.J.H.; Geissler, J.; Lissenberg-Thunnissen, S.; et al. FcγRIIIb restricts antibody-dependent destruction of cancer cells by human neutrophils. Front. Immunol. 2019, 9, 3124. [Google Scholar] [CrossRef] [PubMed]
- Albrengues, J.; Shields, M.A.; Ng, D.; Park, C.G.; Ambrico, A.; Poindexter, M.E.; Upadhyay, P.; Uyeminami, D.L.; Pommier, A.; Küttner, V.; et al. Neutrophil extracellular traps produced during inflammation awaken dormant cancer cells in mice. Science 2018, 361, eaao4227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thålin, C.; Hisada, Y.; Lundström, S.; Mackman, N.; Wallén, H. Neutrophil extracellular traps: Villains and targets in arterial, venous, and cancer-associated thrombosis. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1724–1738. [Google Scholar] [CrossRef]
- Jung, H.S.; Gu, J.; Kim, J.-E.; Nam, Y.; Song, J.W.; Kim, H.K. Cancer cell-induced neutrophil extracellular traps promote both hypercoagulability and cancer progression. PLoS ONE 2019, 14, e0216055. [Google Scholar] [CrossRef] [Green Version]
- Shao, B.-Z.; Yao, Y.; Li, J.-P.; Chai, N.-L.; Linghu, E.-Q. The role of neutrophil extracellular traps in cancer. Front. Oncol. 2021, 11, 714357. [Google Scholar] [CrossRef]
- Kramer, E.D.; Abrams, S.I. Granulocytic myeloid-derived suppressor cells as negative regulators of anticancer immunity. Front. Immuunol. 2020, 11, 1963. [Google Scholar] [CrossRef]
- Krishnamoorthy, M.; Gerhardt, L.; Vareki, S.M. Immunosuppressive effects of myeloid-derived suppressor cells in cancer and immunotherapy. Cells 2021, 10, 1170. [Google Scholar] [CrossRef]
- Ridker, P.M.; Cushman, M.; Stampfer, M.J.; Tracy, R.P.; Hennekens, C.H. Inflammation, aspirin, and the risk of cardiovascular disease in apparently healthy men. N. Engl. J. Med. 1997, 336, 973–979. [Google Scholar] [CrossRef]
- Ridker, P.M.; Everett, B.M.; Thuren, T.; MacFadyen, J.G.; Chang, W.H.; Ballantyne, C.; Fonseca, F.; Nicolau, J.; Koenig, W.; Anker, S.D.; et al. Antiinflammatory therapy with canakinumab for atherosclerotic disease. N. Engl. J. Med. 2017, 377, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Hattori, S.; Katsuda, S.; Nakanishi, I.; Nagai, Y. Human neutrophil elastase: Degradation of basement membrane components and immunolocalization in the tissue. J. Biochem. 1990, 108, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, G.; Nakano, M.; Prati, F.; Niccoli, G.; Mallus, M.T.; Ramazzotti, V.; Montone, R.A.; Kolodgie, F.D.; Virmani, R.; Crea, F. High levels of systemic myeloperoxidase are associated with coronary plaque erosion in patients with acute coronary syndromes: A clinicopathological study. Circulation 2010, 122, 2505–2513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naruko, T.; Ueda, M.; Haze, K.; van der Wal, A.C.; van der Loos, C.M.; Itoh, A.; Komatsu, R.; Ikura, Y.; Ogami, M.; Shimada, Y.; et al. Neutrophil infiltration of culprit lesions in acute coronary syndromes. Circulation 2002, 106, 2894–2900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.J.; Kettritz, R.; Falk, R.J.; Jennette, J.C.; Gaido, M.L. Apoptosis of endothelial cells induced by the neutrophil serine proteases proteinase 3 and elastase. Am. J. Pathol. 1996, 149, 1617–1626. [Google Scholar]
- Soehnlein, O. Multiple roles for neutrophils in atherosclerosis. Circ. Res. 2012, 110, 875–888. [Google Scholar] [CrossRef] [Green Version]
- Sreejit, G.; Latif, A.A.; Murphy, A.J.; Nagareddy, P.R. Emerging roles of neutrophil-borne S100A8/A9 in cardiovascular inflammation. Pharmacol. Res. 2020, 161, 105212. [Google Scholar] [CrossRef]
- Bonaventura, A.; Montecucco, F.; Dallegri, F.; Carbone, F.; Lüscher, T.F.; Camici, G.G.; Liberale, L. Novel findings in neutrophil biology and their impact on cardiovascular disease. Cardiovasc. Res. 2019, 115, 1266–1285. [Google Scholar] [CrossRef] [PubMed]
- Silvestre-Roig, C.; Braster, Q.; Ortega-Gomez, A.; Soehnlein, O. Neutrophils as regulators of cardiovascular inflammation. Nat. Rev. Cardiol. 2020, 17, 327–340. [Google Scholar] [CrossRef]
- Gan, T.; Yang, Y.; Hu, F.; Chen, X.; Zhou, J.; Li, Y.; Xu, Y.; Wang, H.; Chen, Y.; Zhang, M. TLR3 regulated poly I:C-induced neutrophil extracellular traps and acute lung injury partly through p38 MAP kinase. Front. Microbiol. 2018, 9, 3174. [Google Scholar] [CrossRef]
- Stegelmeier, A.A.; Darzianiazizi, M.; Hanada, K.; Sharif, S.; Wootton, S.K.; Bridle, B.W.; Karimi, K. Type I interferon-mediated regulation of antiviral capabilities of neutrophils. Int. J. Mol. Sci. 2021, 22, 4726. [Google Scholar] [CrossRef] [PubMed]
- Khosroshahi, L.M.; Rezaei, N. Dysregulation of the immune response in coronavirus disease 2019. Cell Biol. Int. 2021, 45, 702–707. [Google Scholar] [CrossRef] [PubMed]
- Schönrich, G.; Raftery, M.J.; Samstag, Y. Devilishly radical NETwork in COVID-19: Oxidative stress, neutrophil extracellular traps (NETs), and T cell suppression. Adv. Biol. Regul. 2020, 77, 100741. [Google Scholar] [CrossRef] [PubMed]
- Chau, A.S.; Weber, A.G.; Maria, N.I.; Narain, S.; Liu, A.; Hajizadeh, N.; Malhotra, P.; Bloom, O.; Marder, G.; Kaplan, B. The longitudinal immune response to coronavirus disease 2019: Chasing the cytokine storm. Arthritis Rheumatol. 2021, 73, 23–35. [Google Scholar] [CrossRef]
- Matthay, M.A.; Leligdowicz, A.; Liu, K.D. Biological mechanisms of COVID-19 acute respiratory distress syndrome. Am. J. Respir. Crit. Care Med. 2020, 202, 1489–1491. [Google Scholar] [CrossRef]
- Hue, S.; Beldi-Ferchiou, A.; Bendib, I.; Surenaud, M.; Fourati, S.; Frapard, T.; Rivoal, S.; Razazi, K.; Carteaux, G.; Delfau-Larue, M.-H.; et al. Uncontrolled innate and impaired adaptive immune responses in patients with COVID-19 acute respiratory distress syndrome. Am. J. Respir. Crit. Care Med. 2020, 202, 1509–1519. [Google Scholar] [CrossRef]
- Yao, C.; Bora, S.A.; Parimon, T.; Zaman, T.; Friedman, O.A.; Palatinus, J.A.; Surapaneni, N.S.; Matusov, Y.P.; Chiang, G.C.; Kassar, A.G.; et al. Cell-type-specific immune dysregulation in severely ill COVID-19 patients. Cell Rep. 2021, 34, 108590. [Google Scholar] [CrossRef]
- De Candia, P.; Prattichizzo, F.; Garavelli, S.; Matarese, G. T cells: Warriors of SARS-CoV-2 infection. Trends Immunol. 2021, 42, 18–30. [Google Scholar] [CrossRef]
- Guan, W.J.; Ni, Z.-Y.; Hu, Y.; Liang, W.-H.; Ou, C.-Q.; He, J.-X.; Liu, L.; Shan, H.; Lei, C.-L.; Hui, D.S.C.; et al. for the China Medical Treatment Expert Group for Covid-19. Clinical characteristics of coronavirus disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Veras, F.P.; Pontelli, M.C.; Silva, C.M.; Toller-Kawahisa, J.E.; de Lima, M.; Nascimento, D.C.; Schneider, A.H.; Caetité, D.; Tavares, L.A.; Paiva, I.M.; et al. SARS-CoV-2-triggered neutrophil extracellular traps mediate COVID-19 pathology. J. Exp. Med. 2020, 217, e20201129. [Google Scholar] [CrossRef]
- Borges, L.; Pithon-Curi, T.C.; Curi, R.; Hatanaka, E. COVID-19 and neutrophils: The relationship between hyperinflammation and neutrophil extracellular traps. Mediat. Inflamm. 2020, 2020, 8829674. [Google Scholar] [CrossRef] [PubMed]
- Yaqinuddin, A.; Kvietys, P.; Kashir, J. COVID-19: Role of neutrophil extracellular traps in acute lung injury. Respir. Investig. 2020, 58, 419–420. [Google Scholar] [CrossRef] [PubMed]
- Szturmowicz, M.; Demkow, U. Neutrophil extracellular traps (NETs) in severe SARS-CoV-2 lung disease. Int. J. Mol. Sci. 2021, 22, 8854. [Google Scholar] [CrossRef] [PubMed]
- Tomar, B.; Anders, H.-J.; Desai, J.; Mulay, S.R. Neutrophils and neutrophil extracellular traps drive necroinflammation in COVID-19. Cells 2020, 9, 1383. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Karakiulakis, G.; Roth, M. Are patients with hypertension and diabetes mellitus at increased risk for COVID-19 infection? Lancet Respir. Med. 2020, 8, e21. [Google Scholar] [CrossRef]
- Chan, L.; Karimi, N.; Morovati, S.; Alizadeh, K.; Kakish, J.E.; Vanderkamp, S.; Fazel, F.; Napoleoni, C.; Alizadeh, K.; Mehrani, Y.; et al. The roles of neutrophil in cytokine storms. Viruses 2021, 13, 2318. [Google Scholar] [CrossRef]
- Nicolai, L.; Leunig, A.; Brambs, S.; Kaiser, R.; Weinberger, T.; Weigand, M.; Muenchhoff, M.; Hellmuth, J.C.; Ledderose, S.; Schulz, H.; et al. Immunothrombotic dysregulation in COVID-19 pneumonia is associated with respiratory failure and coagulopathy. Circulation 2020, 142, 1176–1189. [Google Scholar] [CrossRef]
- Middleton, E.A.; He, X.-Y.; Denorme, F.; Campbell, R.A.; Ng, D.; Salvatore, S.P.; Mostyka, M.; Baxter-Stoltzfus, A.; Borczuk, A.C.; Loda, M.; et al. Neutrophil extracellular traps contribute to immunothrombosis in COVID-19 acute respiratory distress syndrome. Blood 2020, 136, 1169–1179. [Google Scholar] [CrossRef]
- Radermecker, C.; Detrembleur, N.; Guiot, J.; Cavalier, E.; Henket, M.; d’Emal, C.; Vanwinge, C.; Cataldo, D.; Oury, C.; Delvenne, P.; et al. Neutrophil extracellular traps infiltrate the lung airway, interstitial, and vascular compartments in severe COVID-19. J. Exp. Med. 2020, 217, e20201012. [Google Scholar] [CrossRef]
- Skendros, P.; Mitsios, A.; Chrysanthopoulou, A.; Mastellos, D.C.; Metallidis, S.; Rafailidis, P.; Ntinopoulou, M.; Sertaridou, E.; Tsironidou, V.; Tsigalou, C.; et al. Complement and tissue factor-enriched neutrophil extracellular traps are key drivers in COVID-19 immunothrombosis. J. Clin. Investig. 2020, 130, 6151–6157. [Google Scholar] [CrossRef]
- Busch, M.H.; Timmermans, S.A.M.E.G.; Nagy, M.; Visser, M.; Huckriede, J.; Aendekerk, J.P.; de Vries, F.; Potjewijd, J.; Jallah, B.; Ysermans, R.; et al. Neutrophils and contact activation of coagulation as potential drivers of COVID-19. Circulation 2020, 142, 1787–1790. [Google Scholar] [CrossRef] [PubMed]
- Ercan, H.; Schrottmaier, W.C.; Pirabe, A.; Schmuckenschlager, A.; Pereyra, D.; Santol, J.; Pawelka, E.; Traugott, M.T.; Schörgenhofer, C.; Seitz, T.; et al. Platelet phenotype analysis of COVID-19 patients reveals progressive changes in the activation of integrin αIIbβ3, F13A1, the SARS-CoV-2 target EIF4A1 and annexin A5. Front. Cardiovasc. Med. 2021, 8, 779073. [Google Scholar] [CrossRef] [PubMed]
- Iliadi, V.; Konstantinidou, I.; Aftzoglou, K.; Iliadis, S.; Konstantinidis, T.G.; Tsigalou, C. The emerging role of neutrophils in the pathogenesis of thrombosis in COVID-19. Int. J. Mol. Sci. 2021, 22, 5368. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Becerril, B.; Campi-Caballero, R.; Sevilla-Fuentes, S.; Hernández-Regino, L.M.; Hanono, A.; Flores-Bustamante, A.; González-Flores, J.; García-Ávila, C.A.; Aquino-Gálvez, A.; Castillejos-López, M.; et al. Immunothrombosis in COVID-19: Implications of neutrophil extracellular traps. Biomolecules 2021, 11, 694. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).