
A Scoping Review of Rodent Studies Investigating the Epigenetic Mechanisms in the Brain Underlying the Effects of Diet on Depressive-like Behaviour

Abstract
1. Introduction
2. Methods
2.1. Search Strategy
2.2. Eligibility Criteria
2.3. Extraction and Synthesis of Data
2.4. Risk of Bias
3. Results
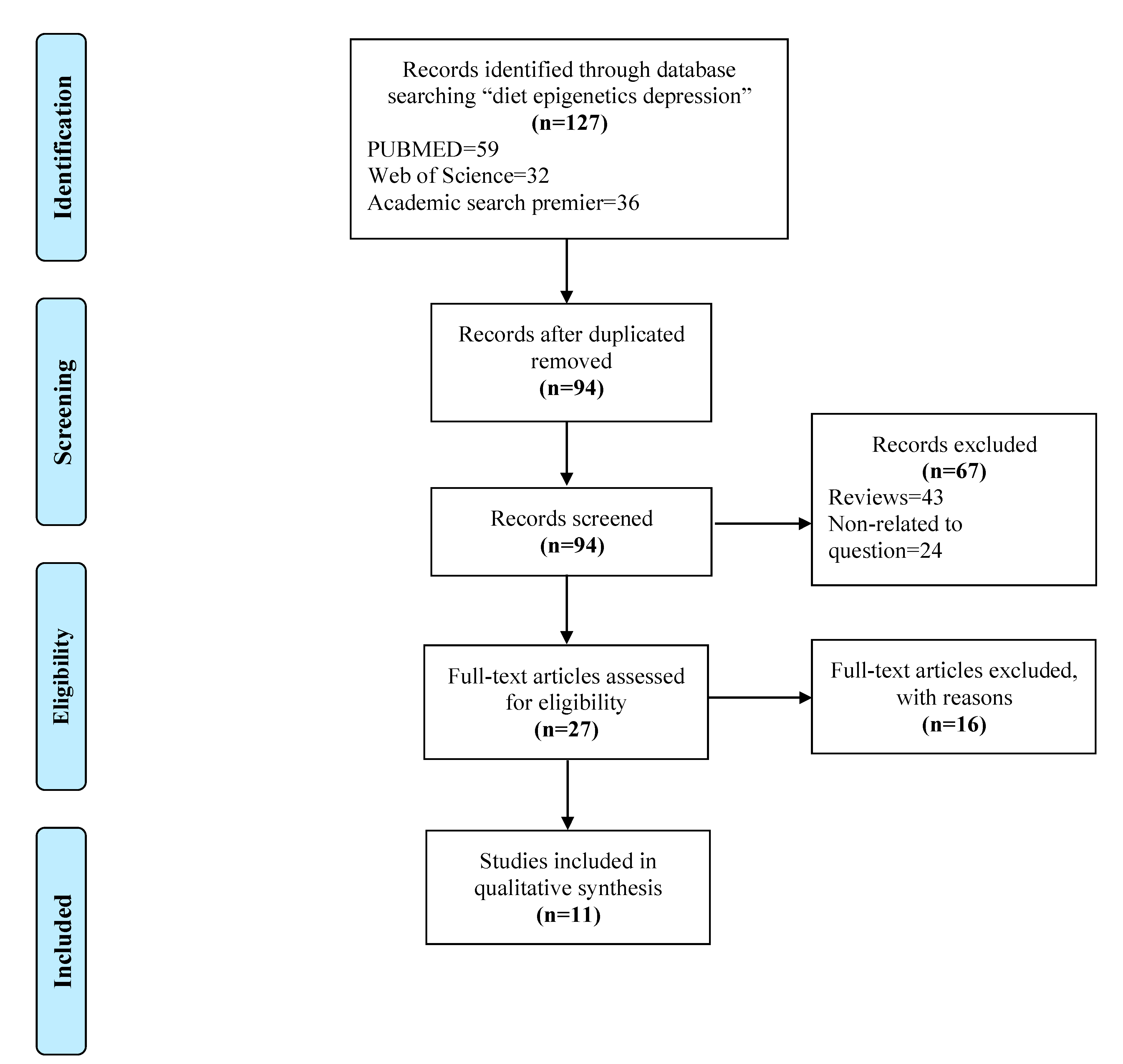
3.1. Selection
3.2. Study Characteristics
3.3. Study Quality and Risk of Bias
3.4. Study of Diet
3.5. Behavioural Measures
3.6. Epigenetic Mechanisms
3.7. Effects of Diet on Mood
3.8. Epigenetic Mechanisms behind the Effects of Diet on Mood
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Friedrich, M.J. Depression Is the Leading Cause of Disability Around the World. JAMA 2017, 317, 1517. [Google Scholar] [CrossRef]
- Li, Y.; Scherer, N.; Felix, L.; Kuper, H. Prevalence of depression, anxiety and post-traumatic stress disorder in health care workers during the COVID-19 pandemic: A systematic review and meta-analysis. PLoS ONE 2021, 16, e0246454. [Google Scholar] [CrossRef]
- McCormack, J.; Korownyk, C. Effectiveness of antidepressants. BMJ 2018, 360, k1073. [Google Scholar] [CrossRef]
- Santarsieri, D.; Schwartz, T.L. Antidepressant efficacy and side-effect burden: A quick guide for clinicians. Drugs Context 2015, 4, 212290. [Google Scholar] [CrossRef]
- Zarate, C.A., Jr.; Singh, J.B.; Carlson, P.J.; Brutsche, N.E.; Ameli, R.; Luckenbaugh, D.A.; Charney, D.S.; Manji, H.K. A randomized trial of an N-methyl-D-aspartate antagonist in treatment-resistant major depression. Arch. Gen. Psychiatry 2006, 63, 856–864. [Google Scholar] [CrossRef]
- Hashimoto, K. Molecular mechanisms of the rapid-acting and long-lasting antidepressant actions of (R)-ketamine. Biochem. Pharmacol. 2020, 177, 113935. [Google Scholar] [CrossRef]
- Otte, C.; Gold, S.M.; Penninx, B.W.; Pariante, C.M.; Etkin, A.; Fava, M.; Mohr, D.C.; Schatzberg, A.F. Major depressive disorder. Nat. Rev. Dis. 2016, 2, 16065. [Google Scholar] [CrossRef]
- Peña, C.J.; Bagot, R.C.; Labonté, B.; Nestler, E.J. Epigenetic Signaling in Psychiatric Disorders. J. Mol. Biol. 2014, 426, 3389–3412. [Google Scholar] [CrossRef]
- Zhang, L.; Lu, Q.; Chang, C. Epigenetics in Health and Disease. Adv. Exp. Med. Biol. 2020, 1253, 3–55. [Google Scholar]
- Asif, M. The prevention and control the type-2 diabetes by changing lifestyle and dietary pattern. J. Educ. Health Promot. 2014, 3, 1. [Google Scholar] [CrossRef]
- Atakan, M.; Koşar, Ş.; Güzel, Y.; Tin, H.; Yan, X. The Role of Exercise, Diet, and Cytokines in Preventing Obesity and Improving Adipose Tissue. Nutrients 2021, 13, 1459. [Google Scholar] [CrossRef]
- Theodoratou, E.; Timofeeva, M.; Li, X.; Meng, X.; Ioannidis, J.P. Nature, Nurture, and Cancer Risks: Genetic and Nutritional Contributions to Cancer. Annu. Rev. Nutr. 2017, 37, 293–320. [Google Scholar] [CrossRef] [PubMed]
- Akbaraly, T.N.; Brunner, E.J.; Ferrie, J.E.; Marmot, M.G.; Kivimaki, M.; Singh-Manoux, A. Dietary pattern and depressive symptoms in middle age. Br. J. Psychiatry 2009, 195, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Lecorguillé, M.; Teo, S.; Phillips, C.M. Maternal Dietary Quality and Dietary Inflammation Associations with Offspring Growth, Placental Development, and DNA Methylation. Nutrients 2021, 13, 3130. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Timberlake, M.A.; Prall, K.; Dwivedi, Y. The recent progress in animal models of depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 77, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Zhang, L.; Wang, D.; Guan, S.; Lu, P.; Xu, X.; Xu, H. Influence of early life stress on depression: From the perspective of neuroendocrine to the participation of gut microbiota. Aging 2021, 13, 25588–25601. [Google Scholar] [CrossRef]
- Murthy, S.; Gould, E. Early life stress in rodents: Animal models of illness or resilience? Front. Behav. Neurosci. 2018, 12, 157. [Google Scholar] [CrossRef]
- Plotsky, P.M.; Thrivikraman, K.V.; Nemeroff, C.B.; Caldji, C.; Sharma, S.; Meaney, M.J. Long-Term Consequences of Neonatal Rearing on Central Corticotropin-Releasing Factor Systems in Adult Male Rat Offspring. Neuropsychopharmacology 2005, 30, 2192–2204. [Google Scholar] [CrossRef]
- Franklin, T.B.; Russig, H.; Weiss, I.C.; Gräff, J.; Linder, N.; Michalon, A.; Vizi, S.; Mansuy, I.M. Epigenetic Transmission of the Impact of Early Stress Across Generations. Biol. Psychiatry 2010, 68, 408–415. [Google Scholar] [CrossRef]
- Gudsnuk, K.; Champagne, F.A. Epigenetic Influence of Stress and the Social Environment. ILAR J. 2012, 53, 279–288. [Google Scholar] [CrossRef]
- Dimitratos, S.M.; Hercules, M.; Stephensen, C.B.; Cervantes, E.; Laugero, K.D. Association between physiological stress load and diet quality patterns differs between male and female adults. Physiol. Behav. 2021, 240, 113538. [Google Scholar] [CrossRef] [PubMed]
- Lutter, M.; Krishnan, V.; Russo, S.; Jung, S.; McClung, C.A.; Nestler, E.J. Orexin Signaling Mediates the Antidepressant-Like Effect of Calorie Restriction. J. Neurosci. 2008, 28, 3071–3075. [Google Scholar] [CrossRef] [PubMed]
- Reddy, B.R.; Maitra, S.; Jhelum, P.; Kumar, K.P.; Bagul, P.K.; Kaur, G.; Banerjee, S.K.; Kumar, A.; Chakravarty, S. Sirtuin 1 and 7 mediate resveratrol-induced recovery from hyper-anxiety in high-fructose-fed prediabetic rats. J. Biosci. 2016, 41, 407–417. [Google Scholar] [CrossRef]
- McCoy, C.R.; Jackson, N.L.; Day, J.; Clinton, S.M. Genetic predisposition to high anxiety- and depression-like behavior coincides with diminished DNA methylation in the adult rat amygdala. Behav. Brain Res. 2017, 320, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Nätt, D.; Barchiesi, R.; Murad, J.; Feng, J.; Nestler, E.J.; Champagne, F.A.; Thorsell, A. Perinatal Malnutrition Leads to Sexually Dimorphic Behavioral Responses with Associated Epigenetic Changes in the Mouse Brain. Sci. Rep. 2017, 7, 11082. [Google Scholar] [CrossRef] [PubMed]
- Cui, P.; Ma, T.; Tamadon, A.; Han, S.; Li, B.; Chen, Z.; An, X.; Shao, L.R.; Wang, Y.; Feng, Y. Hypothalamic DNA methylation in rats with dihydrotestosterone-induced polycystic ovary syndrome: Effects of low-frequency electro-acupuncture. Exp. Physiol. 2018, 103, 1618–1632. [Google Scholar] [CrossRef]
- Cinquina, V.; Calvigioni, D.; Farlik, M.; Halbritter, F.; Fife-Gernedl, V.; Shirran, S.L.; Fuszard, M.A.; Botting, C.H.; Poullet, P.; Piscitelli, F.; et al. Life-long epigenetic programming of cortical architecture by maternal ‘Western’ diet during pregnancy. Mol. Psychiatry 2020, 25, 22–36. [Google Scholar] [CrossRef]
- Ghosh, S.; Sinha, J.K.; Khandelwal, N.; Chakravarty, S.; Kumar, A.; Raghunath, M. Increased stress and altered expression of histone modifying enzymes in brain are associated with aberrant behaviour in vitamin B12 deficient female mice. Nutr. Neurosci. 2020, 23, 714–723. [Google Scholar] [CrossRef]
- Isac, S.; Panaitescu, A.M.; Iesanu, M.I.; Zeca, V.; Cucu, N.; Zagrean, L.; Peltecu, G.; Zagrean, A.-M. Maternal Citicoline-Supplemented Diet Improves the Response of the Immature Hippocampus to Perinatal Asphyxia in Rats. Neonatology 2020, 117, 729–735. [Google Scholar] [CrossRef]
- Gawlińska, K.; Gawliński, D.; Korostyński, M.; Borczyk, M.; Frankowska, M.; Piechota, M.; Filip, M.; Przegaliński, E. Maternal dietary patterns are associated with susceptibility to a depressive-like phenotype in rat offspring. Dev. Cogn. Neurosci. 2021, 47, 100879. [Google Scholar] [CrossRef]
- Xu, P.; Pang, D.; Zhou, J.; Li, S.; Chen, D.; Yu, B. Behavioral changes and brain epigenetic alterations induced by maternal deficiencies of B vitamins in a mouse model. Psychopharmacology 2021, 238, 1213–1222. [Google Scholar] [CrossRef] [PubMed]
- Gawliński, D.; Gawlińska, K.; Smaga, I. Maternal High-Fat Diet Modulates Cnr1 Gene Expression in Male Rat Offspring. Nutrients 2021, 13, 2885. [Google Scholar] [CrossRef] [PubMed]
- Groen, R.N.; Ryan, O.; Wigman, J.T.W.; Riese, H.; Penninx, B.W.J.H.; Giltay, E.J.; Wichers, M.; Hartman, C.A. Comorbidity between depression and anxiety: Assessing the role of bridge mental states in dynamic psychological networks. BMC Med. 2020, 18, 1–17. [Google Scholar] [CrossRef]
- Maugeri, A.; Barchitta, M. How Dietary Factors Affect DNA Methylation: Lesson from Epidemiological Studies. Medicina 2020, 56, 374. [Google Scholar] [CrossRef]
- Gao, Y.-N.; Zhang, Y.-Q.; Wang, H.; Deng, Y.-L.; Li, N.-M. A New Player in Depression: MiRNAs as Modulators of Altered Synaptic Plasticity. Int. J. Mol. Sci. 2022, 23, 4555. [Google Scholar] [CrossRef]
- Wanet, A.; Tacheny, A.; Arnould, T.; Renard, P. miR-212/132 expression and functions: Within and beyond the neuronal compartment. Nucleic Acids Res. 2012, 40, 4742. [Google Scholar] [CrossRef]
- Liu, Y.-P.; Wu, X.; Meng, J.-H.; Yao, J.; Wang, B.-J. Functional Analysis of the 3ʹ Untranslated Region of the Human GRIN1 Gene in Regulating Gene Expression in vitro. Neuropsychiatr. Dis. Treat. 2020, 16, 2361–2370. [Google Scholar] [CrossRef]
- González-Becerra, K.; Ramos-Lopez, O.; Barrón-Cabrera, E.; Riezu-Boj, J.I.; Milagro, F.I.; Martinez-Lopez, E.; Martínez, J.A. Fatty acids, epigenetic mechanisms and chronic diseases: A systematic review. Lipids Health Dis. 2019, 18, 178. [Google Scholar] [CrossRef]
- Haskell, C.F.; Scholey, A.B.; Jackson, P.A.; Elliott, J.M.; Defeyter, M.A.; Greer, J.; Robertson, B.C.; Buchanan, T.; Tiplady, B.; Kennedy, D.O. Cognitive and mood effects in healthy children during 12 weeks’ supplementation with multi-vitamin/minerals. Br. J. Nutr. 2008, 100, 1086–1096. [Google Scholar] [CrossRef]
- White, D.J.; Cox, K.H.M.; Peters, R.; Pipingas, A.; Scholey, A.B. Effects of Four-Week Supplementation with a Multi-Vitamin/Mineral Preparation on Mood and Blood Biomarkers in Young Adults: A Randomised, Double-Blind, Placebo-Controlled Trial. Nutrients 2015, 7, 9005–9017. [Google Scholar] [CrossRef]
- Covington, H.E.; Maze, I.; Sun, H.; Bomze, H.M.; DeMaio, K.D.; Wu, E.Y.; Dietz, D.M.; Lobo, M.K.; Ghose, S.; Mouzon, E.; et al. A Role for Repressive Histone Methylation in Cocaine-Induced Vulnerability to Stress. Neuron 2011, 71, 656–670. [Google Scholar] [CrossRef]
- Wang, Z.; Qin, G.; Zhao, T.C. HDAC4: Mechanism of regulation and biological functions. Epigenomics 2014, 6, 139–150. [Google Scholar] [CrossRef]
- Kennedy, D.O. B Vitamins and the Brain: Mechanisms, Dose and Efficacy—A Review. Nutrients 2016, 8, 68. [Google Scholar] [CrossRef]
- Jahng, J.W.; Kim, J.G.; Kim, H.J.; Kim, B.-T.; Kang, D.-W.; Lee, J.-H. Chronic food restriction in young rats results in depression- and anxiety-like behaviors with decreased expression of serotonin reuptake transporter. Brain Res. 2007, 1150, 100–107. [Google Scholar] [CrossRef]
- Martin, C.K.; Heilbronn, L.; de Jonge, L.; DeLany, J.; Volaufova, J.; Anton, S.D.; Redman, L.M.; Smith, S.R.; Ravussin, E. Effect of Calorie Restriction on Resting Metabolic Rate and Spontaneous Physical Activity. Obesity 2007, 15, 2964–2973. [Google Scholar] [CrossRef]
- Kawashima, H.; Numakawa, T.; Kumamaru, E.; Adachi, N.; Mizuno, H.; Ninomiya, M.; Kunugi, H.; Hashido, K. Glucocorticoid attenuates brain-derived neurotrophic factor-dependent upregulation of glutamate receptors via the suppression of microRNA-132 expression. Neuroscience 2010, 165, 1301–1311. [Google Scholar] [CrossRef]
- Nudelman, A.S.; DiRocco, D.P.; Lambert, T.; Garelick, M.G.; Le, J.; Nathanson, N.; Storm, D.R. Neuronal activity rapidly induces transcription of the CREB-regulated microRNA-132, in vivo. Hippocampus 2010, 20, 492–498. [Google Scholar] [CrossRef]
- Santarelli, D.M.; Beveridge, N.J.; Tooney, P.A.; Cairns, M.J. Upregulation of Dicer and MicroRNA Expression in the Dorsolateral Prefrontal Cortex Brodmann Area 46 in Schizophrenia. Biol. Psychiatry 2011, 69, 180–187. [Google Scholar] [CrossRef]
- Rong, H.; Liu, T.B.; Yang, K.J.; Yang, H.C.; Wu, D.H.; Liao, C.P.; Hong, F.; Yang, H.Z.; Wan, F.; Ye, X.Y.; et al. MicroRNA-134 plasma levels before and after treatment for bipolar mania. J. Psychiatr. Res. 2011, 45, 92–95. [Google Scholar] [CrossRef]
- Chang, L.; Wei, Y.; Hashimoto, K. Brain–gut–microbiota axis in depression: A historical overview and future directions. Brain Res. Bull. 2022, 182, 44–56. [Google Scholar] [CrossRef]


{kind=link}
| Studies | Journal Name | Peer Reviewed | Subject of Study (for Diet) | Age (Start Experiment) | Sex |
|---|---|---|---|---|---|
| Lutter et al., 2008 [22] | Journal of Neuroscience | Yes | Orexin-deficient C57BL/6 mice | 8–10 weeks old | Male and female |
| Reddy et al., 2016 [23] | Journal of Biosciences | Yes | Sprague–Dawley rats | 7 to 8 weeks old | Male |
| McCoy et al., 2017 [24] | Behavioural Brain Research | Yes | Sprague Dawley rats HR/LR bred line | adult (do not specify age) | Male and female |
| Nätt et al., 2017 [25] | Scientific Reports | Yes | Sv129 mice and C57BL/6 J(JAX)mice for mechanistic study | Offspring (10 weeks old) | Male and female |
| Cui et al., 2018 [26] | Experimental Physiology | Yes | Wistar rats | 21 days old | Female |
| Cinquina et al., 2020 [27] | Molecular Psychiatry | Yes | C57Bl6/J, cholecystokinin (CCK)BAC/DsRed and CCKBAC/DsRed::GAD67gfp/+ transgenic mice | E18.5 fetuses | Male and female |
| Ghosh et al., 2020 [28] | Nutritional Neuroscience | Yes | C57BL/6 mice | 12 weeks old | Female |
| Isac et al., 2020 [29] | Neonatology | Yes | Wistar rats | Offspring (6 day old) | Male and female |
| Gawlińska et al., 2021 [30] | Developmental Cognitive Neuroscience | Yes | Wistar rats | Offspring (28 days old) | Male and female |
| Xu et al., 2021 [31] | Psychopharmacology | Yes | C57BL/6 J mice | Offspring (8 weeks old) | Male and female |
| Gawliński et al., 2021 [32] | Nutrients (MDPI) | Yes | Wistar rats | Offspring (4 and 8 weeks old) | Male and female |
| Studies | Random Sequence Generation (Selection Bias) | Baseline Characteristics (Selection Bias) | Allocation Concealment (Selection Bias) | Random Housing (Performance Bias) | Investigator Blinding (Performance Bias) | Random Outcome Assessment (Detection Bias) | Blinding of Outcome Data (Detection Bias) | Incomplete Outcome Data (Attrition Bias) | Selective Outcome Reporting (Reporting Bias) |
|---|---|---|---|---|---|---|---|---|---|
| Lutter et al., 2008 [22] | |||||||||
| Reddy et al., 2016 [23] | |||||||||
| McCoy et al., 2017 [24] | |||||||||
| Nätt et al., 2017 [25] | |||||||||
| Cui et al., 2018 [26] | |||||||||
| Cinquina et al., 2020 [27] | |||||||||
| Ghosh et al., 2020 [28] | |||||||||
| Isac et al., 2020 [29] | |||||||||
| Gawlińska et al., 2021 [30] | |||||||||
| Xu et al., 2021 [31] | |||||||||
| Gawliński et al., 2021 [32] |
| Studies | Diet | Diet Characteristics | Animal Model | Behavioural Test | Epigenetic Mechanism | Epigenetic Measures |
|---|---|---|---|---|---|---|
| Lutter et al., 2008 [22] | Calorie restricted diet | Food pellet equal to 60% of their average daily food intake before onset of the dark phase (7:00 P.M.) | Chronic social defeat stress | FST, SI | Functional histone alteration | Dimethyl lysine 9 H3 (H3K9me2) at orexin promoter |
| Reddy et al., 2016 [23] | High-fructose diet | Control diet (65% corn starch diet), high-fructose diet (65% fructose diet), resveratrol group (65% fructose diet with 10 mg/kg/day of resveratrol) and metformin group (65% fructose diet with300 mg/kg/day of metformin) | Diet-induced prediabetic condition causing depressive phenotype | OFT, BT | Histone deacetylation | Levels of NAD-dependent class III histone deacetylases (sirtuins) |
| McCoy et al., 2017 [24] | Methyl donor rich or depleted diet | Control diet (semisynthetic L-amino acid-complete rodent diet), methyl donors depleted diet (L-amino acid-defined rodent diet lacking 90% of normal requirements of choline, folate, and methionine), methyl donors supplemented diet (chow fortified with increased amounts of folic acid, choline, methionine, and Vitamin B12) | Model of temperamental differences, Low Novelty Responder (LR) and High Novelty Responder (HR) rats. | EPM, OFT, SI, and FST | DNA methylation | DNA methyltransferase (DNMT) expression, global DNA methylation (5-methylcytosine) and methylome sequencing |
| Nätt et al., 2017 [25] | Maternal low protein diet | Low protein diet with 8% protein content (78% carbohydrates; 5% fat) or regular chow with 20% protein content (66% carbohydrates; 5% fat) | Early life stress by maternal low protein diet | EPM, FST, OFT, | Interplay between EGR1 and epigenetic mechanisms like DNA methylation | Transcription of the Egr family genes, DNA-methylation in Npy1r, Interaction of Npy, Npy1/2/5r with EGR1 |
| Cui et al., 2018 [26] | DIO-diet induced obesity model | DIO-60% fat diet for 3 months/control has normal rodent chow | Diet induced obesity | SPT, FST, OFT, EPM | DNA methylation | DNMTs levels, DNA methylation |
| Cinquina et al., 2020 [27] | Maternal ‘Western’ diet | Hypercaloric diet enriched ~15-fold in ω-6 PUFAs while maintaining the ω-3:ω-6 PUFA ratio at 1:8 as in standard chow or to a standard diet before and during pregnancy | Early life stress by maternal hypercaloric enriched diet | OFT, EPM | DNA methylation and chromatin accessibility | Chromatin accessibility of transcription factors and DNA methylation |
| Ghosh et al., 2020 [28] | Vitamin B12 deficient diet | AIN76A control diet with cellulose, AIN-76A diet deficient in vitamin B12 with pectin. Vitamin B12 content in the restricted diets 62.5% lesser than the control diet | Chronic B12 deficiency before and during pregnancy | OFT, EPM, BT, SPT, SI, TS | Histone modifications | Gene expression of class I, class IIa and class IIb HDACs, histone methyltransferases |
| Isac et al., 2020 [29] | Maternal citicoline-supplemented diet | Citicoline 200 mg/kg body weight/day in water (CDS) or standard diet | Perinatal asphyxia | OFT, FST | MicroRNAs | MicroRNA expression levels |
| Gawlińska et al., 2021 [30] | High fat diet, high carbohydrate diet and mixed diet | Standard diet (SD; 65% carbohydrate, 13% fat, 22% protein, 3.4 kcal/g), high-fat diet (HFD; 60% fat, 5.31 kcal/g), high-carbohydrate diet (HCD; 70% carbohydrate: rich in sucrose—40%, 3.77 kcal/g) or mixed diet (MD; 56% carbohydrate, 28% fat, 3.90 kcal/g) | Modified maternal diet | EZM, FST, SPT | Epigenetic profile changes | Gene expression by RNA sequencing (they don’t really look at a specific mechanism) |
| Xu et al., 2021 [31] | Vitamin B6, B9, B12, and folate deficient diets | Control diet (0.07% vitamin B6, 0.02% folate, 0.25% vitamin B12, with an energy content of 17.9% protein, 7.0% fat, and 64.4% carbohydrate) (NOR); vitamin B6 deficient diet (no B6) (DB6); folate-deficient diet (no folate) (DB9); vitamin B12–deficient diet (no B12) (DB12); and vitamin B6–, B9–, and B12–deficient diet (no B6, folate, and B12) (DB6912) | Early life stress by maternal vitamin deficient diet | SPT, OFT, FST, TST, SI | Functional histone alteration | Dimethylated lysine 9 on histone H3 (H3K9me2) |
| Gawliński et al., 2021 [32] | Standard diet and high fat diet | Standard diet (13% fat, 3.4 kcal/g) or High-fat diet (60% fat, 5.31 kcal/g) | Modified maternal diet | FST | DNA methylation and alterations in miRNA | CpG methylation (Cnr1 Promoter) and expression levels of miR-154-3p and miR-212-5p |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Lafuente, C.L.; Reive, B.S.; Kalynchuk, L.E.; Caruncho, H.J. A Scoping Review of Rodent Studies Investigating the Epigenetic Mechanisms in the Brain Underlying the Effects of Diet on Depressive-like Behaviour. Biomedicines 2022, 10, 3213. https://doi.org/10.3390/biomedicines10123213
Sánchez-Lafuente CL, Reive BS, Kalynchuk LE, Caruncho HJ. A Scoping Review of Rodent Studies Investigating the Epigenetic Mechanisms in the Brain Underlying the Effects of Diet on Depressive-like Behaviour. Biomedicines. 2022; 10(12):3213. https://doi.org/10.3390/biomedicines10123213
Chicago/Turabian StyleSánchez-Lafuente, Carla L., Brady S. Reive, Lisa E. Kalynchuk, and Hector J. Caruncho. 2022. "A Scoping Review of Rodent Studies Investigating the Epigenetic Mechanisms in the Brain Underlying the Effects of Diet on Depressive-like Behaviour" Biomedicines 10, no. 12: 3213. https://doi.org/10.3390/biomedicines10123213
APA StyleSánchez-Lafuente, C. L., Reive, B. S., Kalynchuk, L. E., & Caruncho, H. J. (2022). A Scoping Review of Rodent Studies Investigating the Epigenetic Mechanisms in the Brain Underlying the Effects of Diet on Depressive-like Behaviour. Biomedicines, 10(12), 3213. https://doi.org/10.3390/biomedicines10123213