Capacity of Pseudomonas Strains to Degrade Hydrocarbons, Produce Auxins and Maintain Plant Growth under Normal Conditions and in the Presence of Petroleum Contaminants



Abstract
1. Introduction
2. Results and Discussion
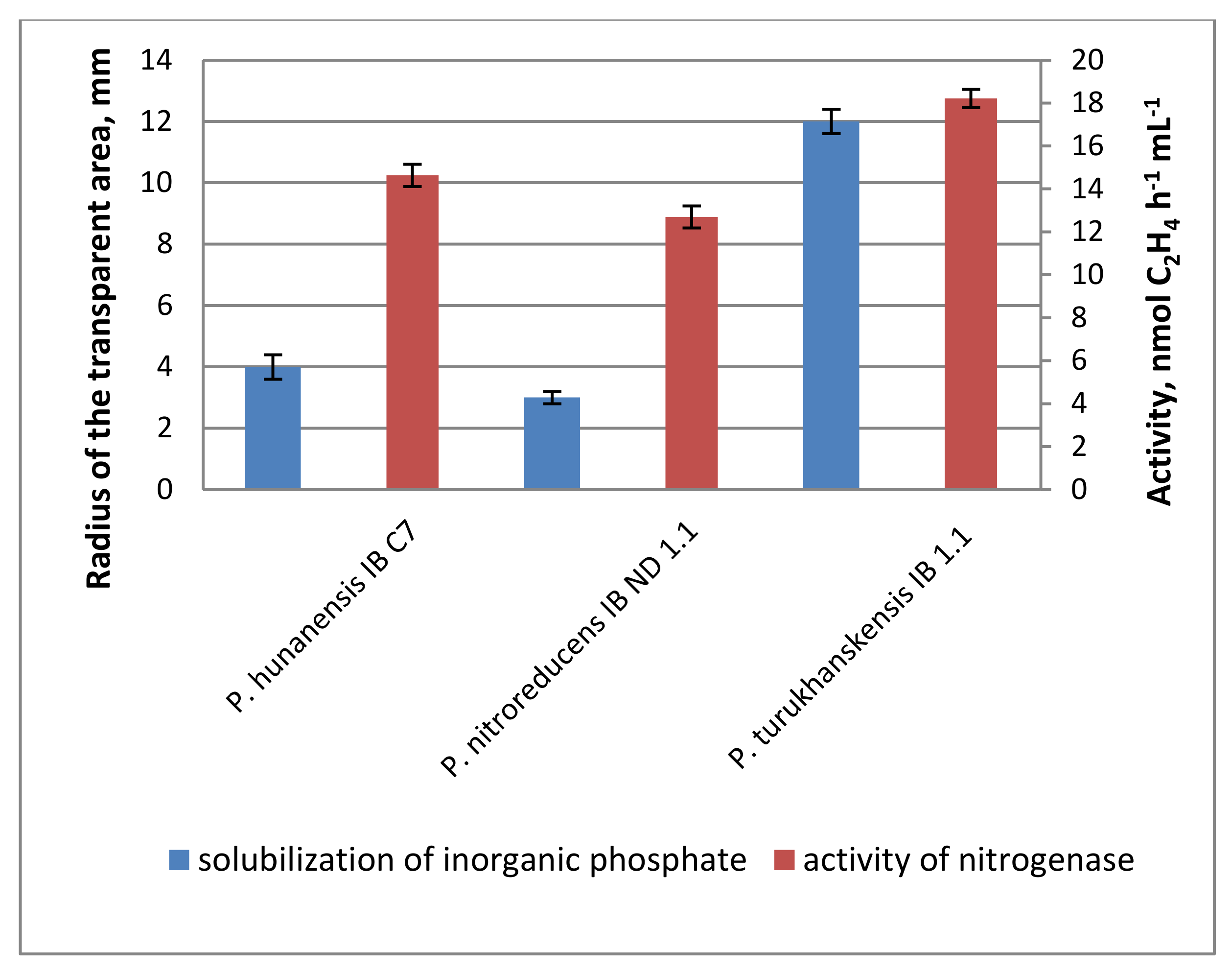
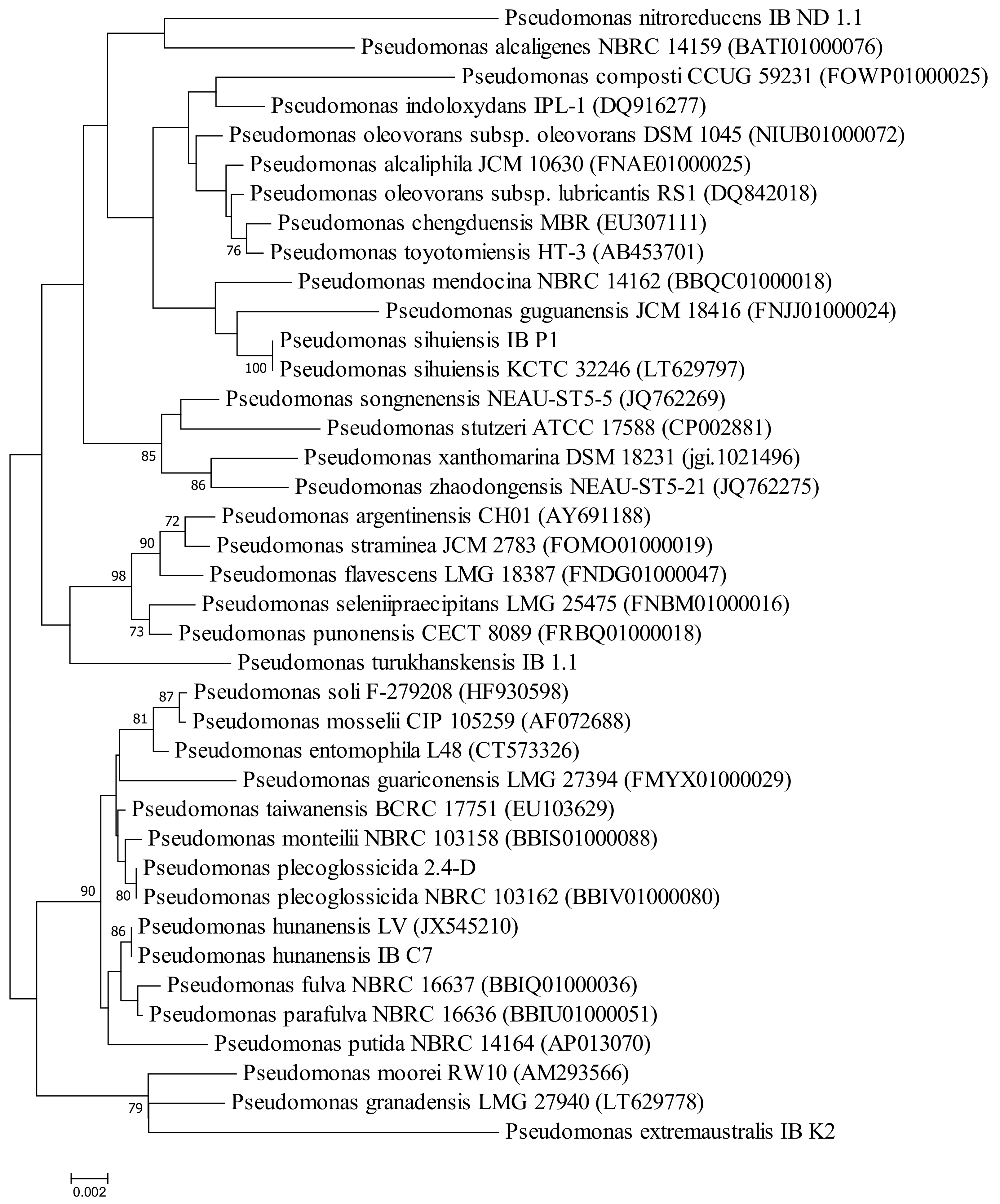
2.1. Properties of Microorganisms
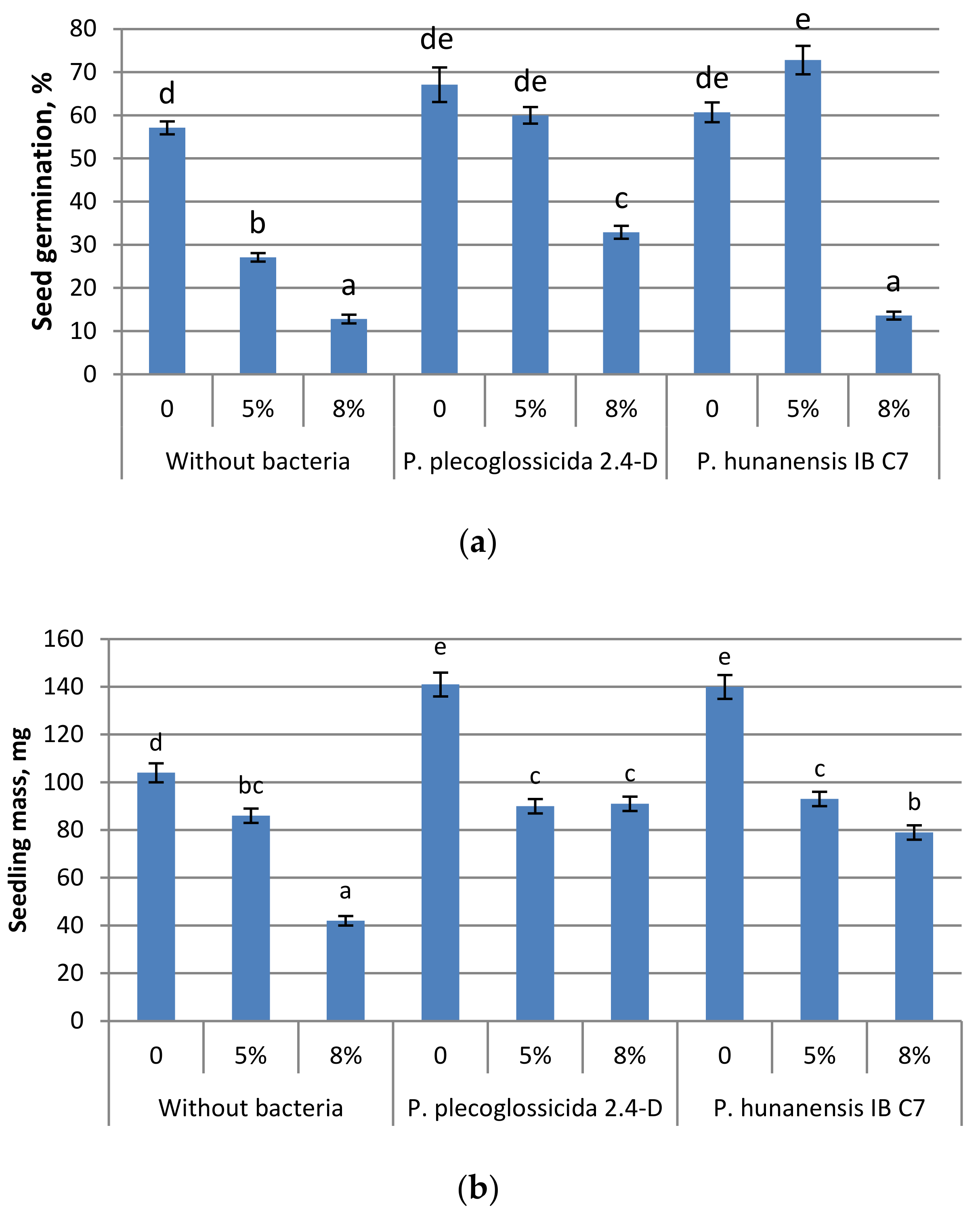
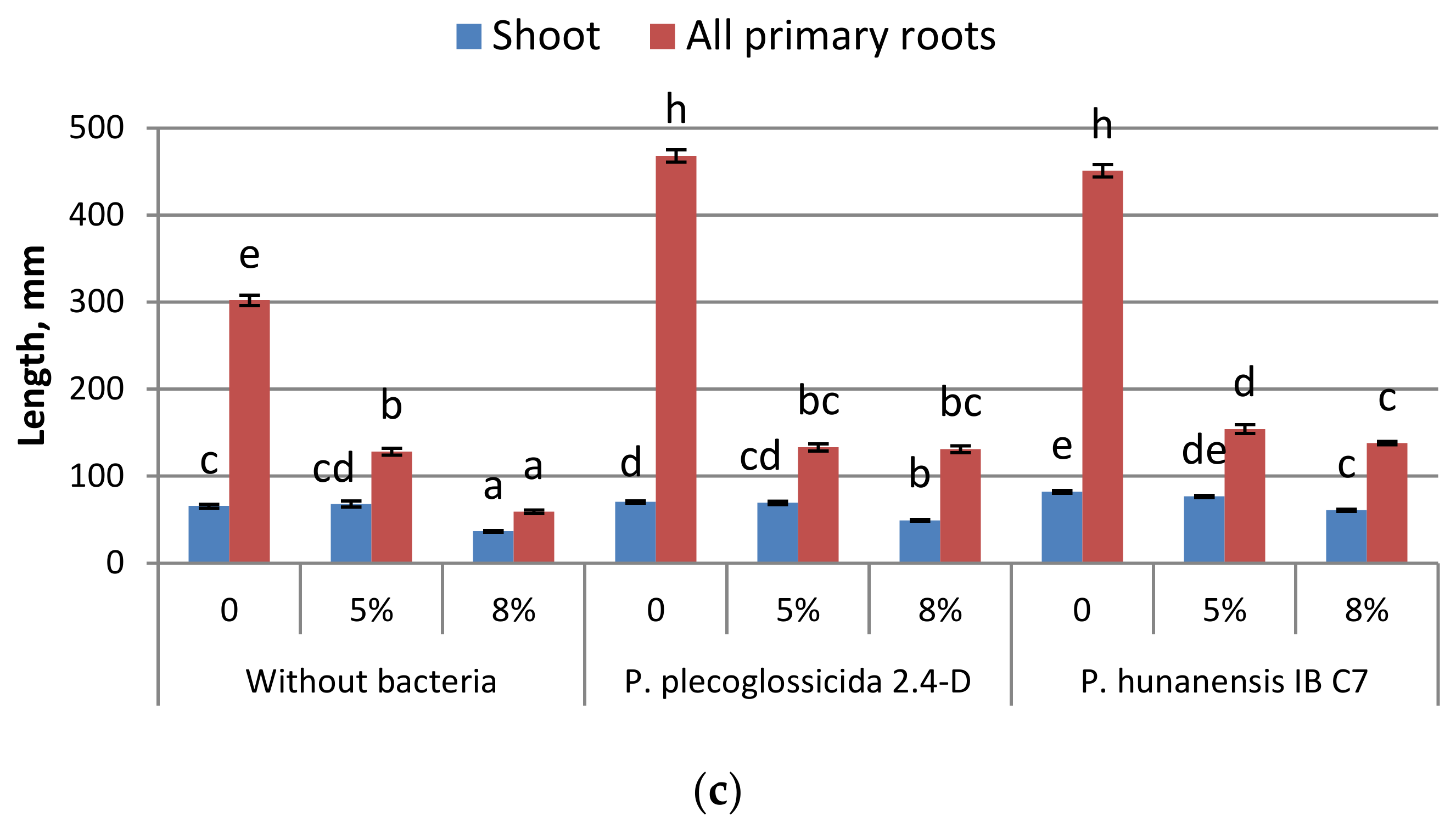
2.2. Effect of Microorganisms on Plants
3. Materials and Methods
Statistics
Author Contributions
Funding
Conflicts of Interest
References
- Afzal, M.; Khan, Q.M.; Sessitsch, A. Endophytic bacteria: Prospects and applications for the phytoremediation of organic pollutants. Chemosphere 2014, 117, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Fox, C.H.; O’Hara, P.D.; Bertazzon, S.; Morgan, K.; Underwood, F.E.; Paquet, P.C. A preliminary spatial assessment of risk: Marine birds and chronic oil pollution on Canada’s pacific coast. Sci. Total Environ. 2016, 573, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Varjani, S.; Gnansounou, E.; Gurunathan, B.; Pant, D.; Zakaria, Z. Polycyclic aromatic hydrocarbons from petroleum oil industry activities: Effect on human health and their biodegradation. In Waste Bioremediation, Energy, Environment and Sustainability; Springer: Singapore, 2018; pp. 185–199. [Google Scholar]
- Muratova, A.Y.; Dmitrieva, T.V.; Panchenko, L.V.; Turkovskaya, O.V. Phytoremediation of oil-sludge-contaminated soil. Int. J. Phytoremediat. 2008, 10, 486–502. [Google Scholar] [CrossRef] [PubMed]
- Fatima, K.; Imran, A.; Naveed, M.; Afzal, M. Plant-bacteria synergism: An innovative approach for the remediation of crude oil contaminated soils. Soil Environ. 2017, 6, 93–113. [Google Scholar] [CrossRef]
- Hatami, E.; Abbaspour, A.; Dorostkar, V. Phytoremediation of a petroleum-polluted soil by native plant species in Lorestan Province, Iran. Environ. Sci. Pollut. Res. Int. 2018, 22. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Afzal, M.; Iqbal, S.; Khan, Q.M. Plant-bacteria partnerships for the remediation of hydrocarbon contaminated soils. Chemosphere 2013, 90, 1317–1332. [Google Scholar] [CrossRef]
- Fatima, K.; Afzal, M.; Imran, A.; Khan, Q.M. Bacterial rhizosphere and endosphere populations associated with grasses and trees to be used for phytoremediation of crude oil contaminated soil. Bull. Environ. Contam. Toxicol. 2015, 94, 314–320. [Google Scholar] [CrossRef]
- Benson, A.; Ram, G.; John, A.; Joe, M.M. Inoculation of 1-aminocyclopropane-1-carboxylate deaminase–producing bacteria along with biosurfactant application enhances the phytoremediation efficiency of Medicago sativa in hydrocarbon-contaminated soils. Bioremediat. J. 2017, 21, 20–29. [Google Scholar] [CrossRef]
- Korshunova, T.Y.; Chetverikov, S.P.; Bakaeva, M.D.; Kuzina, E.V.; Rafikova, G.F.; Chetverikova, D.V.; Loginov, O.N. Microorganisms in the elimination of oil pollution consequences (review). Appl. Biochem. Microbiol. 2019, 55, 344–354. [Google Scholar] [CrossRef]
- Gkorezis, P.; Daghio, M.; Franzetti, A.; Van Hamme, J.D.; Sillen, W.; Vangronsveld, J. The Interaction between plants and bacteria in the remediation of petroleum hydrocarbons: An environmental perspective. Front Microbiol. 2016; 7, 1836. [Google Scholar] [CrossRef]
- Glick, B.R. Modifying a plant’s response to stress by decreasing ethylene production. In Phytoremediation and Rhizoremediation. Theoretical Background. Focus on Biotechnology; Mackova, M., Dowling, D., Macek, T., Eds.; Springer: Dordrecht, The Netherlands, 2006; Volume 9A, pp. 227–236. [Google Scholar]
- Hou, J.; Liu, W.; Wang, B.; Wang, Q.; Luo, Y.; Franks, A.E. PGPR enhanced phytoremediation of petroleum contaminated soil and rhizosphere microbial community response. Chemosphere 2015, 138, 592–598. [Google Scholar] [CrossRef]
- Agnello, A.C.; Bagard, M.; van Hullebusch, E.D.; Espositob, G.; Huguenot, D. Comparative bioremediation of heavy metals and petroleum hydrocarbons co-contaminated soil by natural attenuation, phytoremediation, bioaugmentation and bioaugmentation-assisted phytoremediation. Sci. Total Environ. 2016, 563–564, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, K.E.; Huang, X.-D.; Glick, B.R.; Greenberg, B.M. Phytoremediation and rhizoremediation of organic soil contaminants: Potential and challenges. Plant Sci. 2009, 176, 20–30. [Google Scholar] [CrossRef]
- Rubin, R.L.; van Groenigen, K.J.; Hungate, B.A. Plant growth promoting rhizobacteria are more effective under drought: A meta-analysis. Plant Soil 2017, 416, 309–323. [Google Scholar] [CrossRef]
- Ullah, S.; Ashraf, M.; Asgar, H.N.; Iqbal, Z.; Ali, R. Plant growth promoting rhizobacteria mediated amelioration of drought in crop plants. Soil Environ. 2019, 38, 1–20. [Google Scholar] [CrossRef]
- Kudoyarova, G.R.; Vysotskaya, L.B.; Arkhipova, T.N.; Kuzmina, L.Y.; Galimsyanova, N.F.; Sidorova, L.V.; Gabbasova, I.M.; Melentiev, A.I.; Veselov, S.Y. Effect of auxin producing and phosphate solubilizing bacteria on mobility of soil phosphorus, growth rate, and P acquisition by wheat plants. Acta Physiol. Plant. 2017, 39, 253. [Google Scholar] [CrossRef]
- Travaglia, C.; Masciarelli, O.; Fortuna, J.; Marchetti, G.; Cardozo, P.; Lucero, M.; Zorza, E.; Luna, V.; Reinoso, H. Towards sustainable maize production: Glyphosate detoxification by Azospirillum sp. and Pseudomonas sp. Crop Prot. 2015, 77, 102–109. [Google Scholar] [CrossRef]
- Han, Y.-Y.; Han, G.; Li, K.-R.; Zhang, X.-X. Effects of brassinolide on photosynthetic parameters of Robinia pseudoacacia seedlings in petroleum polluted soil. Nat. Environ. Pollut. Technol. 2017, 16, 199–204. [Google Scholar]
- Spaepen, S.; Vanderleyden, J. Auxin and plant-microbe interactions. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef]
- Shi, T.-Q.; Peng, H.; Zeng, S.-Y.; Ji, R.-Y.; Shi, K.; Huang, H.; Ji, X. Microbial production of plant hormones: Opportunities and challenges. Bioengineered 2017, 8, 124–128. [Google Scholar] [CrossRef]
- Kudoyarova, G.; Arkhipova, T.; Korshunova, T.; Bakaeva, M.; Loginov, O.; Dodd, I.C. Phytohormone mediation of interactions between plants and non-symbiotic growth promoting bacteria under edaphic stresses. Front. Plant Sci. 2019, 10, 1368. [Google Scholar] [CrossRef]
- Golubev, S.N.; Muratova, A.Y.; Wittenmayer, L.; Bondarenkova, A.D.; Hirche, F.; Matora, L.Y.; Merbach, W.; Turkovskaya, O.V. Rhizosphere indole-3-acetic acid as a mediator in the Sorghum bicolor–phenanthrene–Sinorhizobium melilotiinteractions. Plant Physiol. Biochem. 2011, 49, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Korshunova, T.Y.; Chetverikov, S.P.; Loginov, O.; Ramírez-Bahena, M.-H.; Igual, J.M.; Peix, Á. Pseudomonas turukhanskensis sp. nov., isolated from oil-contaminated soils. Int. J. Syst. Evol. Microbiol. 2016, 6, 4657–4664. [Google Scholar] [CrossRef] [PubMed]
- Vignesh, R.; Arularasan, A.; Gandhiraj, V.; Deepika, R.C. Isolation identification and characterization of potential oil degrading bacteria from oil contaminated sites. Int. Res. J. Eng. Technol. 2016, 3, 2503–2508. [Google Scholar]
- Koshlaf, E.; Ball, A.S. Soil bioremediation approaches for petroleum hydrocarbon polluted environments. AIMS Microbiol. 2017, 3, 25–49. [Google Scholar] [CrossRef]
- Xu, J.G.; Johnson, R.L. Nitrogen dynamics in soils with different hydrocarbon contents planted to barley and field pea. Can. J. Soil Sci. 1997, 77, 453–458. [Google Scholar] [CrossRef]
- Kamath, R.; Rentz, J.A.; Schnoor, J.L.; Alvarez, P.J.J. Phytoremediation of hydrocarbon-contaminated soils: Principles and applications. In Petroleum Biotechnology: Developments and Perspectives Studies in Surface Science and Catalysis; Vazquez-Duhalt, R., Quintero-Ramirez, R., Eds.; Elsevier Science: Oxford, UK, 2004; Volume 51, pp. 447–478. [Google Scholar]
- Shtangeeva, I.; Perämäki, P.; Niemelä, M.; Kurashov, E.; Krylova, Y. Potential of wheat (Triticum aestivum L.) and pea (Pisum sativum) for remediation of soils contaminated with bromides and PAHs. Int. J. Phytoremediat. 2018, 20, 560–566. [Google Scholar] [CrossRef]
- Dickinson, S.J.; Rutherford, P.M. Utilization of biosolids during the phytoremediation of hydrocarbon-contaminated soil. J. Environ. Qual. 2006, 35, 982–991. [Google Scholar] [CrossRef]
- Soleimani, M.; Afyuni, M.; Hajabbasi, M.A.; Nourbakhsh, F.; Sabzalian, M.R.; Christensen, J.H. Phytoremediation of an aged petroleum contaminated soil using endophyte infected and non-infected grasses. Chemosphere 2010, 81, 1084–1090. [Google Scholar] [CrossRef]
- Ertekin, O.; Erol, C.; Unlu, S.; Yildizhan, Y.; Pelitli, V.; Yuksel, B.; Memon, A. Aliphatic hydrocarbon fingerprints in Trifolium spp. Fresenius Environ. Bull. 2011, 20, 367–371. [Google Scholar]
- Chetverikov, S.P.; Sharipov, D.A.; Korshunova, T.Y.; Loginov, O.N. Degradation of perfluorooctanyl sulfonate by strain Pseudomonas plecoglossicida 2.4-D. Appl. Biochem. Microbiol. 2017, 53, 533–538. [Google Scholar] [CrossRef]
- Nie, M.; Wang, Y.; Yu, J.; Xiao, M.; Jiang, L.; Yang, J.; Fang, C.; Chen, J.; Li, B. Understanding plant–microbe interactions for phytoremediation of petroleumpolluted soil. PLoS ONE 2011, 6, e17961. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: Chichester, UK, 1991; pp. 115–177. [Google Scholar]
- Raymond, R.L. Microbial oxidation of n-paraffinic hydrocarbons. Develop. Ind. Microbiol. 1961, 2, 23–32. [Google Scholar]
- King, E.O.; Ward, M.K.; Raney, D.E. Two simple media for the demonstration of pyocyanin and fluorescein. J. Lab. Clin. Med. 1954, 44, 301–307. [Google Scholar] [PubMed]
- Veselov, S.Y.; Kudoyarova, G.R.; Egutkin, N.L.; Guili-Zade, V.Z.; Mustafina, A.R.; Kof, E.M. Modified solvent partitioning scheme providing increased specificity and rapidity of immunoassay for indole 3- acetic acid. Physiol. Plant. 1992, 86, 93–96. [Google Scholar] [CrossRef]
- Kudoyarova, G.R.; Melentiev, A.I.; Martynenko, E.V.; Timergalina, L.N.; Arkhipova, T.N.; Shendel, G.V.; Kuz’mina, L.Y.; Dodd, I.C.; Veselov, S.Y. Cytokinin producing bacteria stimulate amino acid deposition by wheat roots. Plant Physiol. Biochem. 2014, 83, 285–291. [Google Scholar] [CrossRef]
- Pikovskaya, R.I. Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Mikrobiologiya 1948, 17, 362–370. [Google Scholar]
- Hardy, R.W.F.; Burns, R.C.; Holsten, R.D. Application of the acetylene-ethylene assay for measurement of nitrogen fixation. Soil Biol. Biochem. 1973, 5, 47–81. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Number in the Collection of UIB UFRC RAS | Species/Strain | 16S rRNA Nucleotide Sequence Similarity, % | Strain Origin |
|---|---|---|---|
| IB K2 | Pseudomonas extremaustralis 14-3T | 99.93 | Soil contaminated with oil (Republic of Bashkortostan, Russia) |
| IB C7 (B-3229D) | P. hunanensis LVT | 99.56 | Steppe soil (Sol-Iletsky District, Orenburg region, Russia) |
| IB ND 1.1 | P. nitroreducens DSM 14399 | 99.90 | Soil from the site of oil spill (Khanty-Mansi Autonomous Area, Russia) |
| IB P1 | P. sihuiensis KCTC 32246T | 99.06 | Bilge water from a transport vessel (port of Novorossiysk, Russia) |
| Strains | Extent of Degradation of Hydrocarbon Substrate, % | ||||
|---|---|---|---|---|---|
| Oil | Diesel Fuel | Marine Fuel Oil | Lubricating Oil | Heavy Fuel Oil | |
| Pseudomonas extremaustralis IB K2 | 51.3 ± 0.7a | 65.5 ± 3.4 a | 47.6 ± 1.3 a | 17.1 ± 0.3 c | 22.2 ± 0.3 d |
| P. hunanensis IB C7 | 73.0 ± 1.6 b | 87.9 ± 2.2 c | 59.0 ± 1.2 c | 34.7 ± 0.4 d | 24.8 ± 0.2 e |
| P. nitroreducens IB ND 1.1 | 83.3 ± 1.6 c | 90.2 ± 1.8 c | 45.5 ± 0.9 a | 12.0 ± 0.3 a | 15.2 ± 0.4 b |
| P. plecoglossicida 2.4-D | 69.0 ± 1.5 b | 80.5 ± 2.0 b | Nm | Nm | 19.7 ± 0.3 c |
| P. sihuiensis IB P1 | 54.0 ± 1.9 a | 60.7 ± 3.2 a | 47.9 ± 1.1 a | 13.0 ± 0.2 b | 13.8 ± 0.4 a |
| P. turukhanskensis IB 1.1 | 84.5 ± 2.1 c | 89.0 ± 3.0 c | 51.6 ± 1.1 b | 18.0 ± 0.5 c | 20.5 ± 0.5 c |
| Strains | Without Additives | Oil | Hexadecane | Toluene |
|---|---|---|---|---|
| Pseudomonas extremaustralis IB K2 | 318 ± 37 c | 120 ± 13 ab | 84 ± 3 a | 140 ± 21 ab |
| P. hunanensis IB C7 | 661 ± 88 d | 702 ± 25 d | 654 ± 25 d | 80 ± 5 a |
| P. nitroreducens IB ND 1.1 | 1413 ± 64 e | 289 ± 40 c | 692 ± 94 d | 102 ± 10 a |
| P. plecoglossicida 2.4-D | 1205 ± 72 e | 588 ± 32 d | 518 ± 21 d | 89 ± 4 a |
| P. sihuiensis IB P1 | 1324 ± 38 e | 189 ± 36 bc | 698 ± 7 d | 83 ± 4 a |
| P. turukhanskensis IB 1.1 | 11261 ± 897 g | 6048 ± 809 f | 7301 ± 406 f | 154 ± 12 b |
| Strains | Additives to the Medium | |||
|---|---|---|---|---|
| Without Additives | + Oil | + Hexadecane | + Toluene | |
| Pseudomonas extremaustralis IB K2 | (5.1 ± 0.2) × 109 | (1.8 ± 0.1) × 108 | (2.0 ± 0.1) × 108 | (2.0 ± 0.1) × 105 |
| P. hunanensis IB C7 | (7.9 ± 0.2) × 109 | (8.0 ± 0.2) × 108 | (1.0 ± 0.1) × 109 | (3.0 ± 0.1) × 105 |
| P. nitroreducens IB ND 1.1 | (5.7 ± 0.3) × 109 | (2.6 ± 0.1) × 108 | (2.3 ± 0.1) × 108 | <105 |
| P. plecoglossicida 2.4-D | (5.4 ± 0.3) × 109 | (2.0 ± 0.1) × 108 | (2.6 ± 0.1) × 108 | <105 |
| P. sihuiensis IB P1 | (1.0 ± 0.1) × 1010 | (9.0 ± 0.2) × 107 | (8.0 ± 0.2) × 107 | <105 |
| P. turukhanskensis IB 1.1 | (4.0 ± 0.2) × ·109 | (7.4 ± 0.2) × 108 | (3.3 ± 0.2) × 108 | <105 |
| Plant Species | Pseudomonas extremaustralis IB K2 | P. hunanensis IB C7 | P. nitroreducens IB ND 1.1 | P. plecoglossicida 2.4-D | P. sihuiensis IB P1 | P. turukhanskensis IB 1.1 | Control |
|---|---|---|---|---|---|---|---|
| Avena sativa | 34.40 ± 1.33 | 39.00 ± 1.15 | 56.10 ± 2.14 | 52.50 ± 1.82 | 36.60 ± 1.50 | 37.50 ± 1.26 | 40.00 ± 2.05 |
| Bromus inermis | 57.90 ± 2.30 | 73.33 ± 1.04 | 66.05 ± 2.17 | 69.38 ± 1.76 | 56.65 ± 1.22 | 59.38 ± 1.08 | 59.18 ± 1.40 |
| Festuca pratensis | 40.62 ± 1.85 | 44.38 ± 1.35 | 37.00 ± 1.28 | 39.38 ± 1.15 | 43.75 ± 2.18 | 40.63 ± 1.43 | 43.13 ± 1.07 |
| Hordeum vulgare | 96.65 ± 2.5 | 96.65 ± 2.42 | 96.65 ± 1.98 | 91.65 ± 2.13 | 97.50 ± 1.63 | 97.50 ± 1.54 | 91.65 ± 2.00 |
| Pisum sativum | 95.00 ± 2.73 | 96.65 ± 1.84 | 90.00 ± 2.87 | 98.35 ± 2.15 | 98.35 ± 2.35 | 95.00 ± 2.46 | 91.10 ± 1.55 |
| Sorghum × drummondii | 80.00 ± 3.12 | 83.35 ± 2.25 | 88.35 ± 2.08 | 82.50 ± 1.81 | 78.35 ± 3.07 | 83.35 ± 2.85 | 83.35 ± 2.54 |
| Trifolium pratense | 39.18 ± 1.50 | 60.00 ± 1.49 | 69.18 ± 1.88 | 60.83 ± 1.43 | 36.88 ± 1.40 | 60.00 ± 1.80 | 35.00 ± 1.39 |
| Plant Species | Pseudomonas extremaustralis IB K2 | P. hunanensis IB C7 | P. nitroreducens IB ND 1.1 | P. plecoglossicida 2.4-D | P. sihuiensis IB P1 | P. turukhanskensis IB 1.1 | Control |
|---|---|---|---|---|---|---|---|
| Avena sativa | 86.0 ± 1.2 | 112.4 ± 2.3 | 80.5 ± 0.9 | 99.3 ± 1.6 | 75.3 ± 1.1 | 74.6 ± 1.8 | 69.0 ± 1.5 |
| Bromus inermis | 21.5 ± 0.41 | 20.6 ± 0.18 | 17.7 ± 0.58 | 20.2 ± 0.43 | 17.3 ± 0.53 | 23.7 ± 0.61 | 19.5 ± 0.29 |
| Festuca pratensis | 3.90 ± 0.18 | 4.88 ± 0.07 | 4.23 ± 0.09 | 3.88 ± 0.24 | 4.72 ± 0.08 | 4.84 ± 0.09 | 4.06 ± 0.10 |
| Hordeum vulgare | 150 ± 3 | 172 ± 5 | 160 ± 3 | 156 ± 4 | 179 ± 5 | 179 ± 5 | 105 ± 2 |
| Pisum sativum | 626 ± 9 | 643 ± 4 | 702 ± 10 | 591 ± 14 | 669 ± 4 | 602 ± 5 | 598 ± 4 |
| Sorghum × drummondii | 28.0 ± 0.4 | 40.2 ± 0.9 | 37.7 ± 0.6 | 44.1 ± 0.6 | 39 ± 0.7 | 41.3 ± 1.0 | 34 ± 0.5 |
| Trifolium pratense | 2.67 ± 0.27 | 6.05 ± 0.25 | 4.53 ± 0.23 | 2.54 ± 0.04 | 2.84 ± 0.14 | 2.76 ± 0.17 | 2.60 ± 0.25 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakaeva, M.; Kuzina, E.; Vysotskaya, L.; Kudoyarova, G.; Arkhipova, T.; Rafikova, G.; Chetverikov, S.; Korshunova, T.; Chetverikova, D.; Loginov, O. Capacity of Pseudomonas Strains to Degrade Hydrocarbons, Produce Auxins and Maintain Plant Growth under Normal Conditions and in the Presence of Petroleum Contaminants. Plants 2020, 9, 379. https://doi.org/10.3390/plants9030379
Bakaeva M, Kuzina E, Vysotskaya L, Kudoyarova G, Arkhipova T, Rafikova G, Chetverikov S, Korshunova T, Chetverikova D, Loginov O. Capacity of Pseudomonas Strains to Degrade Hydrocarbons, Produce Auxins and Maintain Plant Growth under Normal Conditions and in the Presence of Petroleum Contaminants. Plants. 2020; 9(3):379. https://doi.org/10.3390/plants9030379
Chicago/Turabian StyleBakaeva, Margarita, Elena Kuzina, Lidiya Vysotskaya, Guzel Kudoyarova, Tat’yana Arkhipova, Gulnaz Rafikova, Sergey Chetverikov, Tat’yana Korshunova, Dar’ya Chetverikova, and Oleg Loginov. 2020. "Capacity of Pseudomonas Strains to Degrade Hydrocarbons, Produce Auxins and Maintain Plant Growth under Normal Conditions and in the Presence of Petroleum Contaminants" Plants 9, no. 3: 379. https://doi.org/10.3390/plants9030379
APA StyleBakaeva, M., Kuzina, E., Vysotskaya, L., Kudoyarova, G., Arkhipova, T., Rafikova, G., Chetverikov, S., Korshunova, T., Chetverikova, D., & Loginov, O. (2020). Capacity of Pseudomonas Strains to Degrade Hydrocarbons, Produce Auxins and Maintain Plant Growth under Normal Conditions and in the Presence of Petroleum Contaminants. Plants, 9(3), 379. https://doi.org/10.3390/plants9030379