
Genome-Wide Identification and Functional Analysis of AP2/ERF Gene Family in Passiflora edulis Sims
 ,
,
Abstract
1. Introduction
2. Results
2.1. Identification of Ninety-One PeAP2/ERF Genes in the Passion Fruit Genome
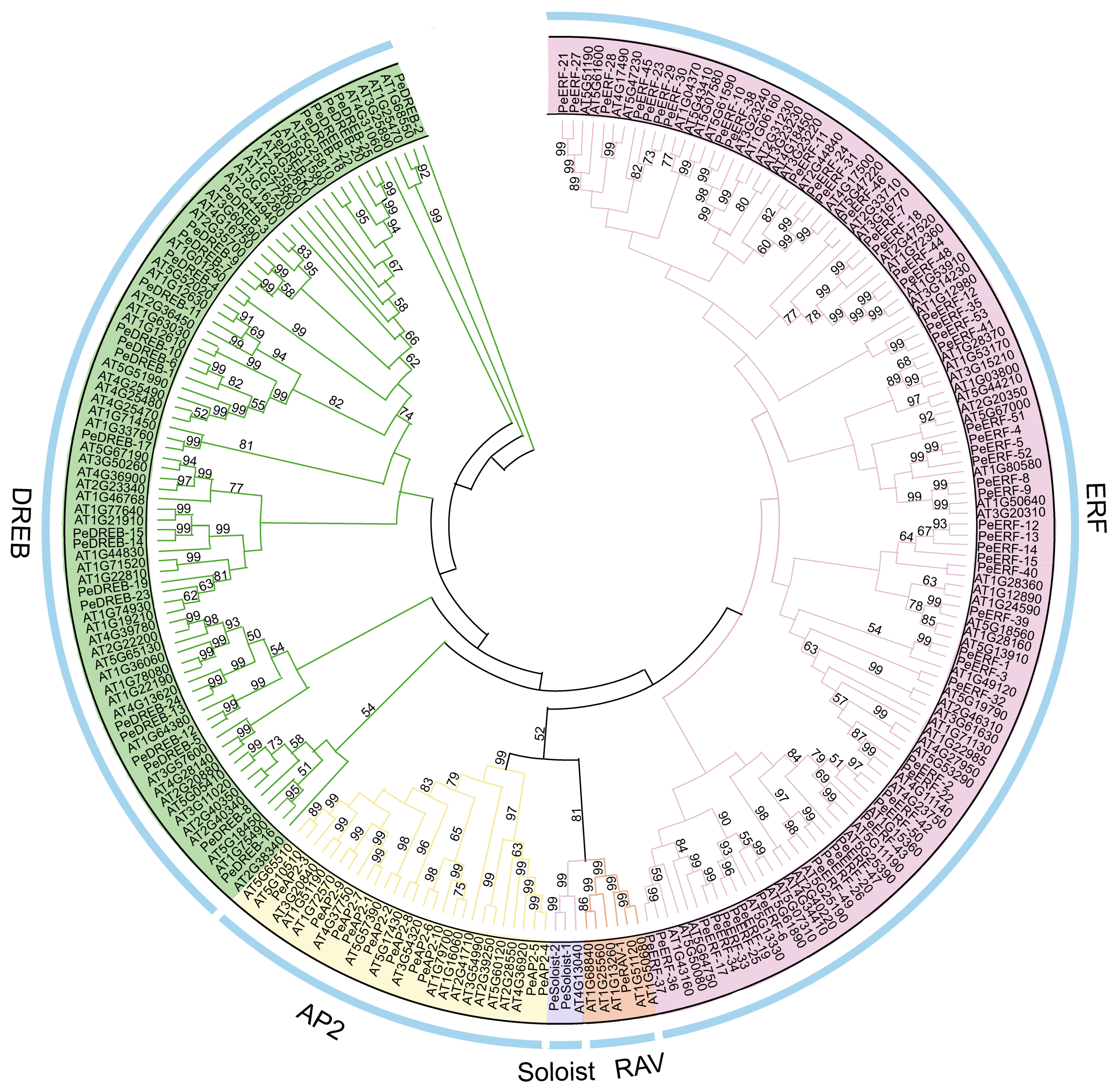
2.2. PeAP2/ERF Proteins Are Classified into Five Subfamilies
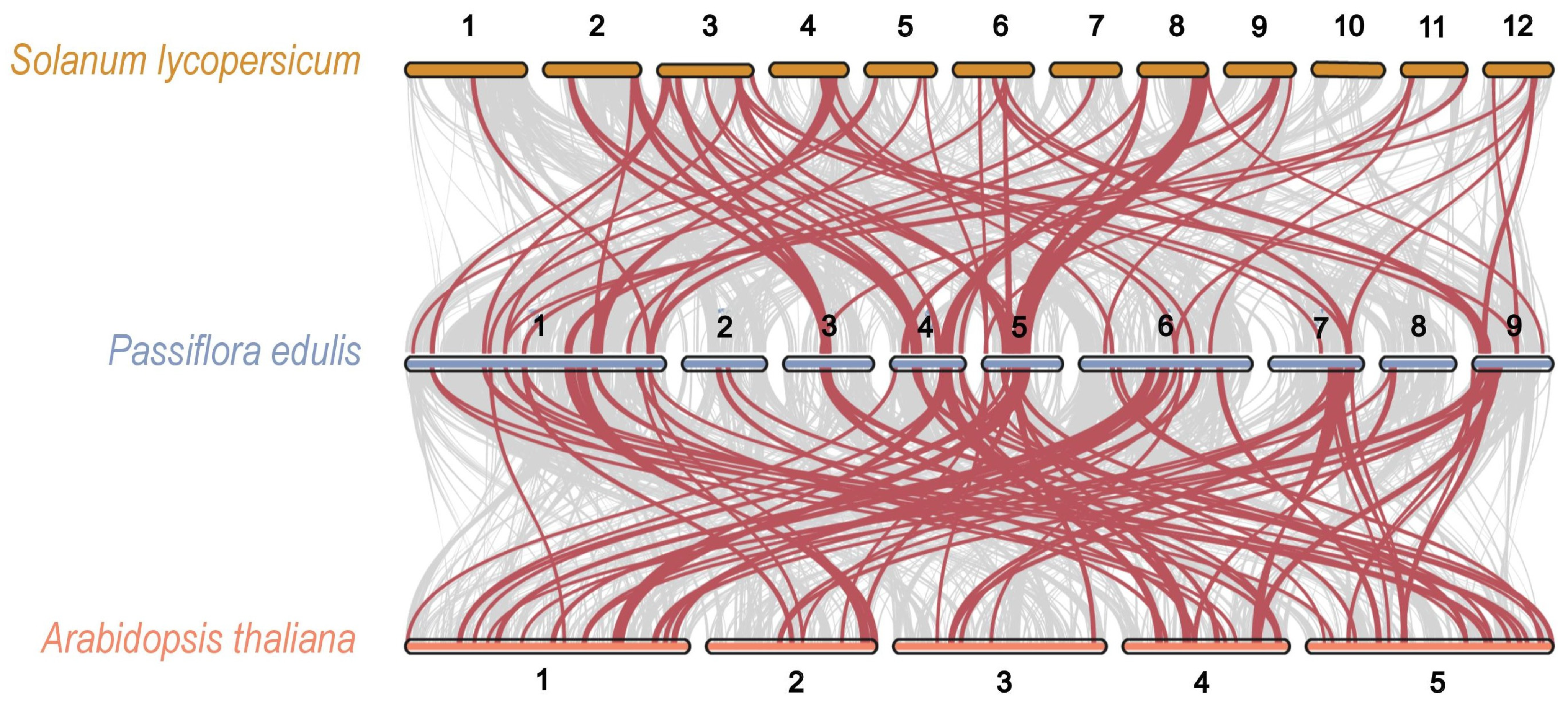
2.3. Chromosome Localization and Collinearity Analysis of the PeAP2/ERF Genes
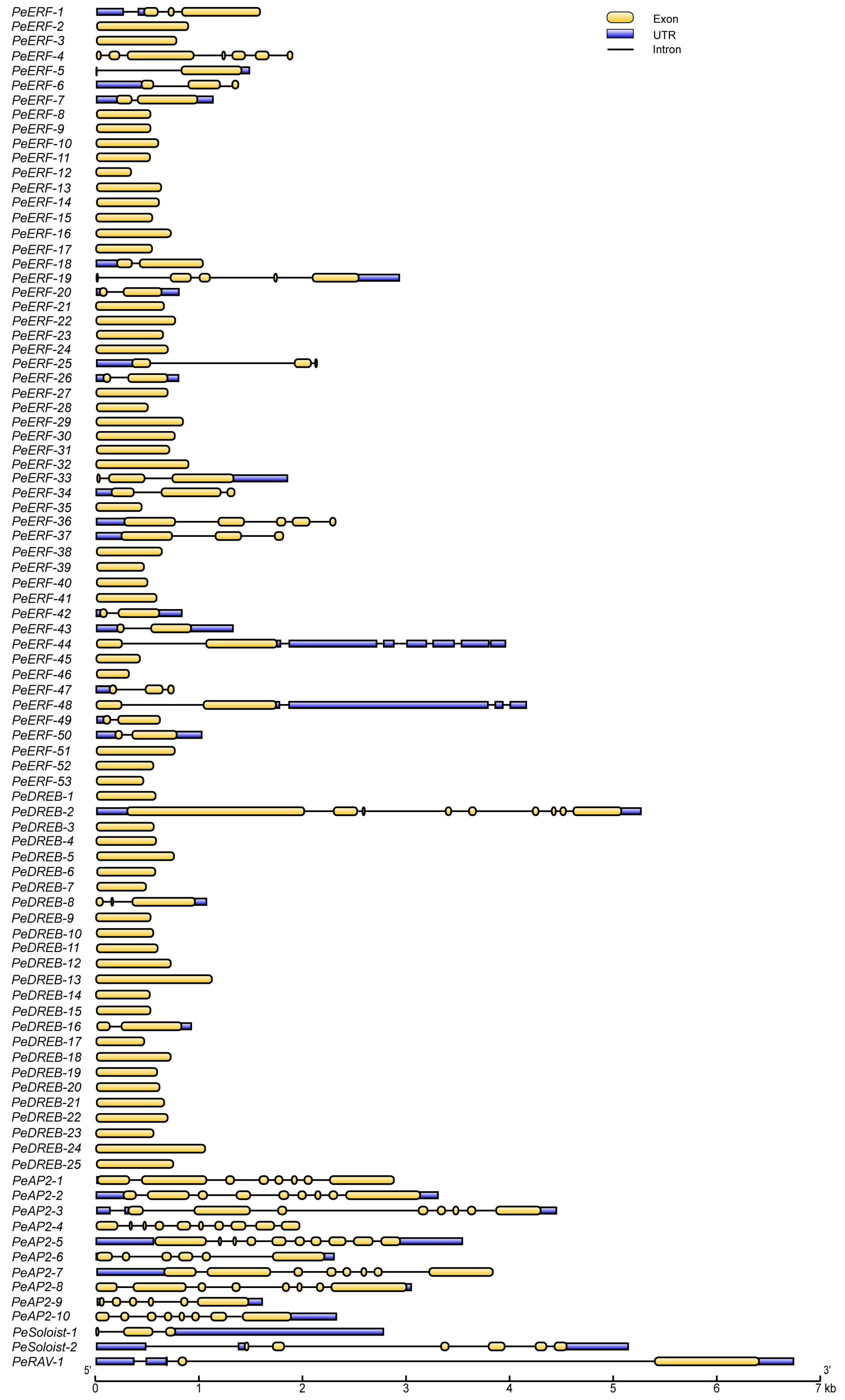
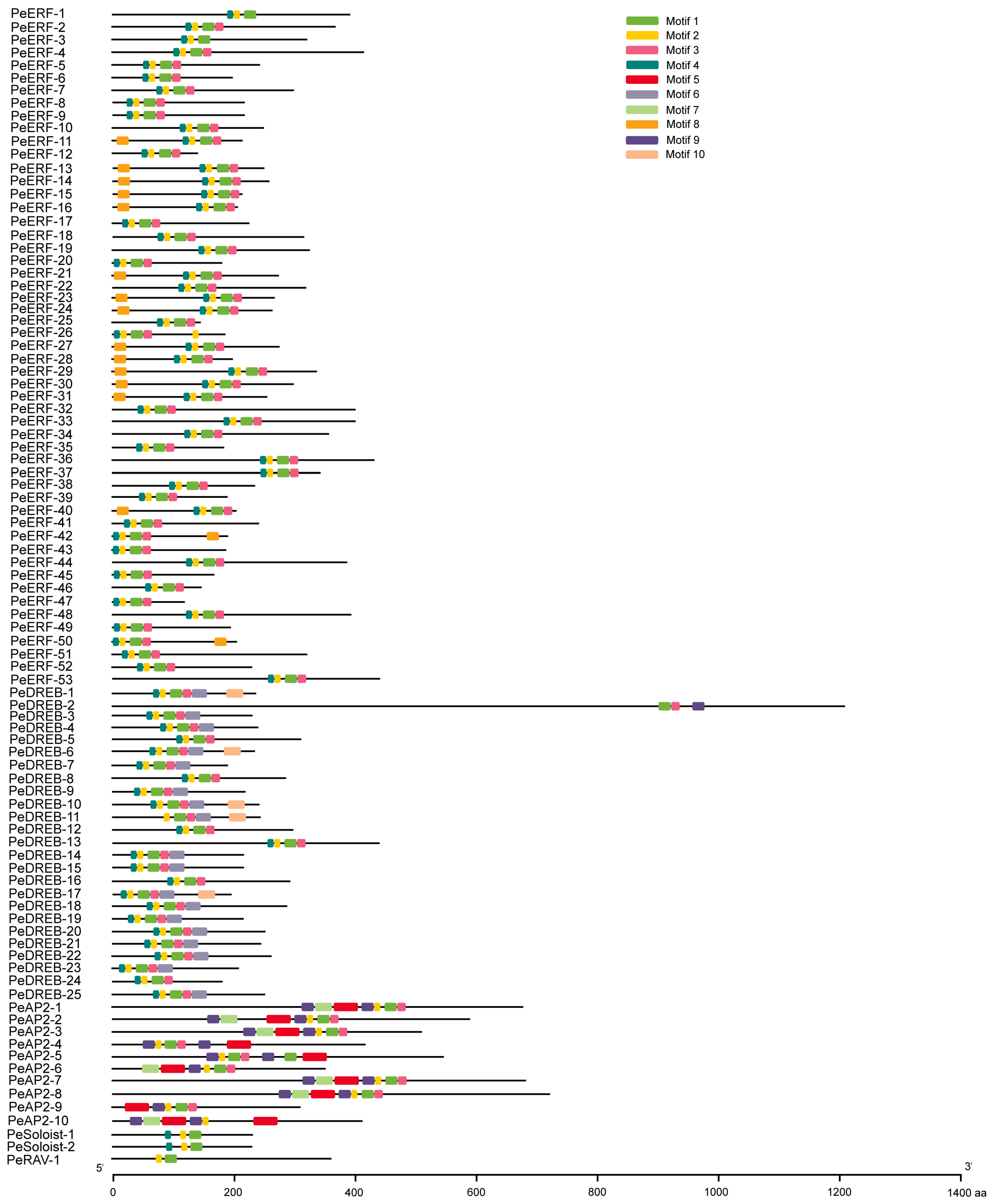
2.4. The Structure of PeAP2/ERFs Is Highly Conserved
2.5. Identification of Cis-Acting Elements in PeAP2/ERF Promoters
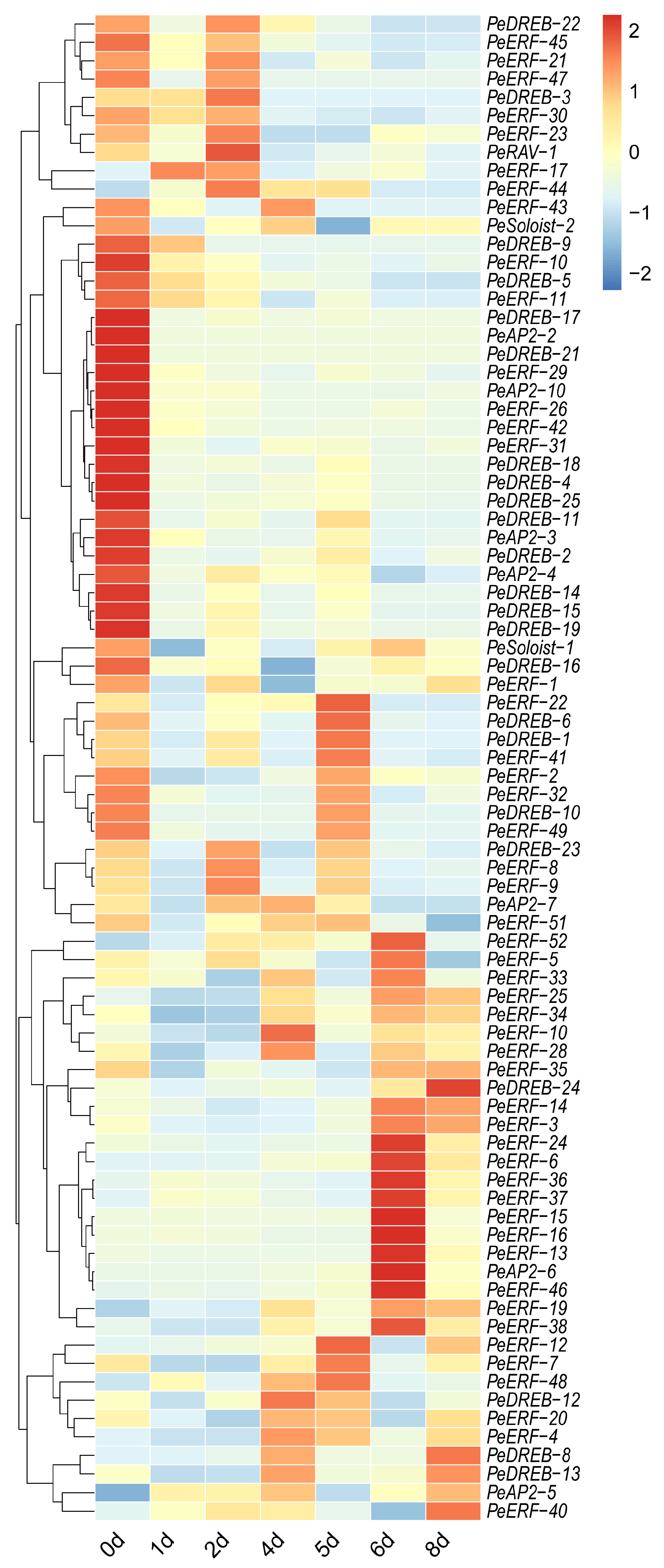
2.6. Analysis of PeAP2/ERFs Expression Pattern in Postharvest Passion Fruit
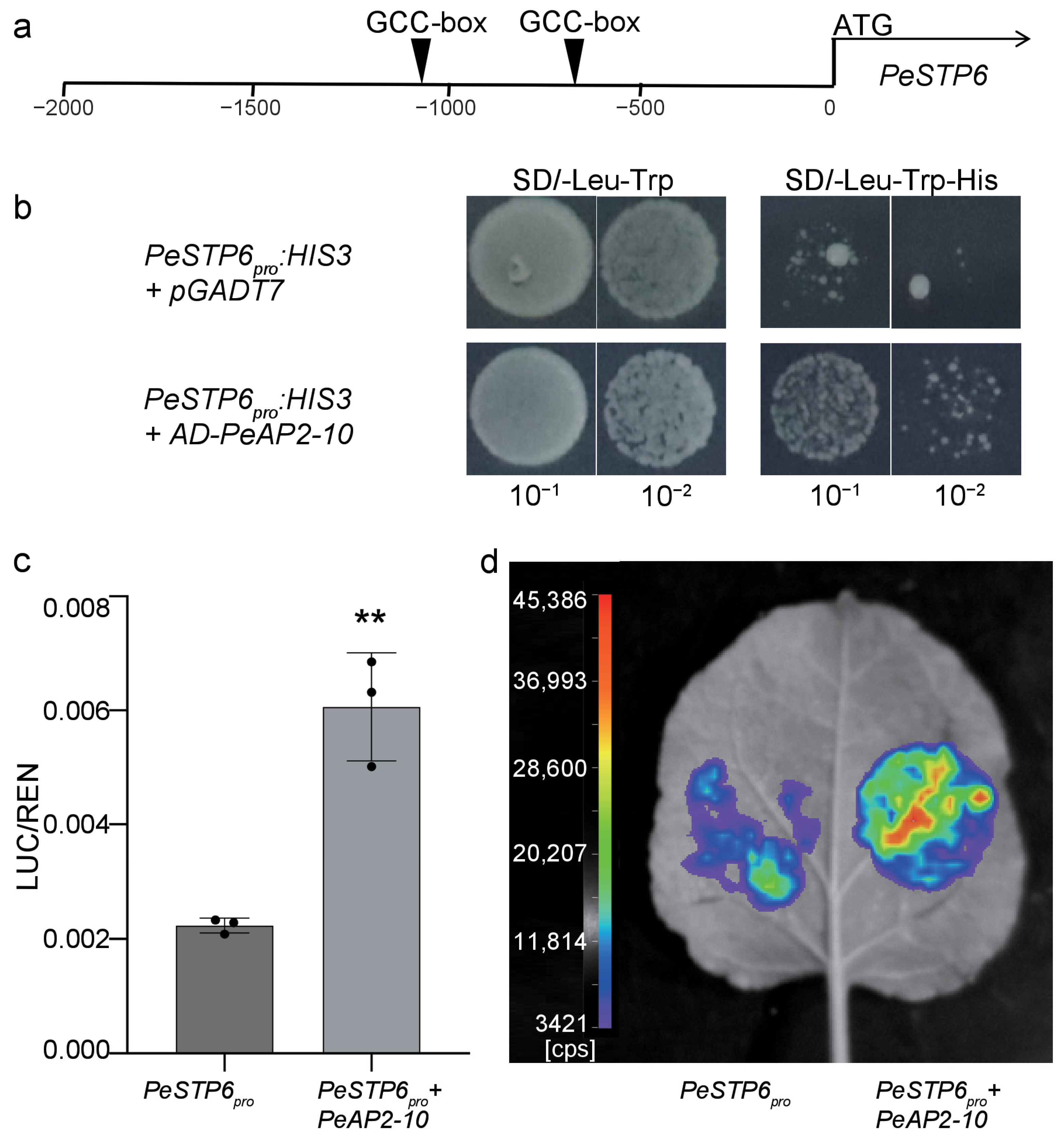
2.7. PeAP2-10 Directly Regulates PeSTP6
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Identification of the PeAP2/ERF Family Members in Passion Fruit
4.3. Phylogenetic Analysis
4.4. Chromosomal Distribution and Collinearity Analysis
4.5. Gene Structure and Conserved Motifs of PeAP2/ERF
4.6. Prediction of Cis-Elements in PeAP2/ERF Genes Promoter
4.7. Yeast One-Hybrid
4.8. Transient Dual Luciferase Reporter Assays
4.9. RNA Extraction and qRT-PCR
4.10. Co-Expression Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Licausi, F.; Ohme-Takagi, M.; Perata, P. APETALA/Ethylene Responsive Factor (AP2/ERF) Transcription Factors: Mediators of Stress Responses and Developmental Programs. New Phytol. 2013, 199, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Dossa, K.; Wei, X.; Li, D.; Fonceka, D.; Zhang, Y.; Wang, L.; Yu, J.; Boshou, L.; Diouf, D.; Cisse, N.; et al. Insight into the AP2/ERF Transcription Factor Superfamily in Sesame and Expression Profiling of Dreb Subfamily under Drought Stress. BMC Plant Biol. 2016, 16, 171. [Google Scholar] [CrossRef] [PubMed]
- Dipp-Alvarez, M.; Cruz-Ramirez, A. A Phylogenetic Study of the Ant Family Points to a Preant Gene as the Ancestor of Basal and Euant Transcription Factors in Land Plants. Front. Plant Sci. 2019, 10, 17. [Google Scholar] [CrossRef]
- Feng, K.; Hou, X.-L.; Xing, G.-M.; Liu, J.-X.; Duan, A.-Q.; Xu, Z.-S.; Li, M.-Y.; Zhuang, J.; Xiong, A.-S. Advances in AP2/ERF Super-Family Transcription Factors in Plant. Crit. Rev. Biotechnol. 2020, 40, 750–776. [Google Scholar] [CrossRef] [PubMed]
- Lv, B.; Wei, K.; Hu, K.; Tian, T.; Zhang, F.; Yu, Z.; Zhang, D.; Su, Y.; Sang, Y.; Zhang, X.; et al. MPK14-Mediated Auxin Signaling Controls Lateral Root Development Via ERF13-Regulated Very-Long-Chain Fatty Acid Biosynthesis. Mol. Plant 2021, 14, 285–297. [Google Scholar] [CrossRef]
- Wessels, B.; Seyfferth, C.; Escamez, S.; Vain, T.; Antos, K.; Vahala, J.; Delhomme, N.; Kangasjarvi, J.; Eder, M.; Felten, J.; et al. An AP2/ERF Transcription Factor ERF139 Coordinates Xylem Cell Expansion and Secondary Cell Wall Deposition. New Phytol. 2019, 224, 1585–1599. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.; Hao, Z.; Tu, Z.; Shen, Y.; Zhang, C.; Wen, S.; Yang, L.; Ma, J.; Li, H. Genome-Wide Survey and Identification of AP2/ERF Genes Involved in Shoot and Leaf Development in Liriodendron chinense. BMC Genom. 2021, 22, 807. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Du, H.; Huang, Z.; He, M.; Kong, L.; Fang, C.; Chen, L.; Yang, H.; Zhang, Y.; Liu, B.; et al. The AP2/ERF Transcription Factor Toe4b Regulates Photoperiodic Flowering and Grain Yield Per Plant in Soybean. Plant Biotechnol. J. 2023, 21, 1682–1694. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Liu, L.; Liu, C.; Song, L.; Dong, Y.; Chen, L.; Li, M. Sweet Cherry AP2/ERF Transcription Factor, PAVRAV2, Negatively Modulates Fruit Size by Directly Repressing PavKLUH Expression. Physiol. Plant. 2023, 175, e14065. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Zhang, K.; Li, C.-Y.; Xie, G.-W.; Lu, M.-T.; Qian, Y.; Shu, Y.-P.; Shen, Q. Genome-Wide Comprehensive Characterization and Transcriptomic Analysis of AP2/ERF Gene Family Revealed Its Role in Seed Oil and Ala Formation in Perilla (Perilla frutescens). Gene 2023, 889, 147808. [Google Scholar] [CrossRef]
- Du, C.; Hu, K.; Xian, S.; Liu, C.; Fan, J.; Tu, J.; Fu, T. Dynamic Transcriptome Analysis Reveals AP2/ERF Transcription Factors Responsible for Cold Stress in Rapeseed (Brassica napus L.). Mol. Genet. Genom. 2016, 291, 1053–1067. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Nishida, S.; Shitan, N.; Sato, F. Genome-Wide Identification of AP2/ERF Transcription Factor-Encoding Genes in California Poppy (Eschscholzia californica) and Their Expression Profiles in Response to Methyl Jasmonate. Sci. Rep. 2020, 10, 18066. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Zhai, Y.; He, J.; Song, M.; Flaishman, M.A.; Ma, H. AP2/ERF Genes Associated with Superfast Fig (Ficus carica L.) Fruit Ripening. Front. Plant Sci. 2022, 13, 1040796. [Google Scholar] [CrossRef]
- Wan, R.; Song, J.; Lv, Z.; Qi, X.; Han, X.; Guo, Q.; Wang, S.; Shi, J.; Jian, Z.; Hu, Q.; et al. Genome-Wide Identification and Comprehensive Analysis of the AP2/ERF Gene Family in Pomegranate Fruit Development and Postharvest Preservation. Genes 2022, 13, 895. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.-C.; Han, Y.-C.; Qi, X.-Y.; Shan, W.; Chen, J.-Y.; Lu, W.-J.; Kuang, J.-F. Papaya CPERF9 Acts as a Transcriptional Repressor of Cell-Wall-Modifying Genes CPPME1/2 and CPPG5 Involved in Fruit Ripening. Plant Cell Rep. 2016, 35, 2341–2352. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.-K.; Li, X.; Xu, Q.; Chen, J.-Y.; Yin, X.-R.; Ferguson, I.B.; Chen, K.-S. EjAP2-1, an AP2/ERF Gene, Is a Novel Regulator of Fruit Lignification Induced by Chilling Injury, Via Interaction with EjMYB Transcription Factors. Plant Biotechnol. J. 2015, 13, 1325–1334. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yuan, J.; Zhang, D.; Deng, K.; Chai, G.; Huang, Y.; Ma, S.; Qin, Y.; Wang, L. Genome-Wide Identification and Characterization of the SBP Gene Family in Passion Fruit (Passiflora edulis Sims). Int. J. Mol. Sci. 2022, 23, 14153. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-S.; Xu, Y.; Xing, W.-T.; Wu, B.; Huang, D.-M.; Ma, F.-N.; Zhan, R.-L.; Sun, P.-G.; Xu, Y.-Y.; Song, S. Identification of the Passion Fruit (Passiflora edulis Sims) MYB Family in Fruit Development and Abiotic Stress, and Functional Analysis of PeMYB87 in Abiotic Stresses. Front. Plant Sci. 2023, 14, 1124351. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ma, F.; Wu, B.; Lv, W.; Xu, Y.; Xing, W.; Chen, D.; Xu, B.; Song, S. Genome-Wide Association and Expression Analysis of the Lipoxygenase Gene Family in Passiflora edulis Revealing PeLOX4 Might Be Involved in Fruit Ripeness and Ester Formation. Int. J. Mol. Sci. 2022, 23, 12496. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Wu, B.; Chen, G.; Xing, W.; Xu, Y.; Ma, F.; Li, H.; Hu, W.; Huang, H.; Yang, L.; et al. Genome-Wide Analysis of the Passion Fruit Invertase Gene Family Reveals Involvement of PeCWINV5 in Hexose Accumulation. BMC Plant Biol. 2024, 24, 836. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Huang, D.; Zhang, S.; Wang, W.; Ma, F.; Wu, B.; Xu, Y.; Xu, B.; Chen, D.; Zou, M.; et al. Chromosome-Scale Genome Assembly Provides Insights into the Evolution and Flavor Synthesis of Passion Fruit (Passiflora edulis Sims). Hortic. Res. 2021, 8, 14. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Li, K.; Cheng, Y.; Li, M.; Liu, L.; Mur, L.A.J.; Luo, J.; Fang, C. PeWRKY20 Represses PeMDH1 to Modulate Malic Acid Metabolism and Flavor Formation in Postharvest Passion Fruit. Postharvest Biol. Technol. 2024, 218, 113164. [Google Scholar] [CrossRef]
- Thirugnanasambantham, K.; Durairaj, S.; Saravanan, S.; Karikalan, K.; Muralidaran, S.; Islam, V.I.H. Role of Ethylene Response Transcription Factor (ERF) and Its Regulation in Response to Stress Encountered by Plants. Plant Mol. Biol. Report. 2015, 33, 347–357. [Google Scholar] [CrossRef]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-Wide Analysis of the ERF Gene Family in Arabidopsis and Rice. Plant Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef] [PubMed]
- Dan, Y.; Niu, Y.; Wang, C.; Yan, M.; Liao, W. Genome-Wide Identification and Expression Analysis of the Trehalose-6-Phosphate Synthase (TPS) Gene Family in Cucumber (Cucumis sativus L.). PeerJ 2021, 9, e11398. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Wei, Y.; Xu, R.; Lin, S.; Luan, H.; Lv, C.; Zhang, X.; Song, X.; Xu, R. Genome-Wide Analysis of Apetala2/Ethylene-Responsive Factor (AP2/ERF) Gene Family in Barley (Hordeum vulgare L.). PLoS ONE 2016, 11, e0161322. [Google Scholar] [CrossRef]
- Yang, H.; Sun, Y.; Wang, H.; Zhao, T.; Xu, X.; Jiang, J.; Li, J. Genome-Wide Identification and Functional Analysis of the ERF2 Gene Family in Response to Disease Resistance against Stemphylium lycopersici in Tomato. BMC Plant Biol. 2021, 21, 72. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Haxim, Y.; Liang, Y.; Qiao, S.; Gao, B.; Zhang, D.; Li, X. Genome-Wide Investigation of AP2/ERF Gene Family in the Desert Legume Eremosparton songoricum: Identification, Classification, Evolution, and Expression Profiling under Drought Stress. Front. Plant Sci. 2022, 13, 885694. [Google Scholar] [CrossRef]
- Guo, Z.; He, L.; Sun, X.; Li, C.; Su, J.; Zhou, H.; Liu, X. Genome-Wide Analysis of the Rhododendron AP2/ERF Gene Family: Identification and Expression Profiles in Response to Cold, Salt and Drought Stress. Plants 2023, 12, 994. [Google Scholar] [CrossRef]
- Jarambasa, T.; Regon, P.; Jyoti, S.Y.; Gupta, D.; Panda, S.K.; Tanti, B. Genome-Wide Identification and Expression Analysis of the Pisum sativum (L.) Apetala2/Ethylene-Responsive Factor (AP2/ERF) Gene Family Reveals Functions in Drought and Cold Stresses. Genetica 2023, 151, 225–239. [Google Scholar] [CrossRef]
- Zhuang, J.; Zhu, B. Analysis of Brassica napus Ests: Gene Discovery and Expression Patterns of AP2/ERF-Family Transcription Factors. Mol. Biol. Rep. 2014, 41, 45–56. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Zhao, B.-Y.; Ye, X.; Du, J.; Song, J.-L.; Wang, W.-J.; Huang, X.-L.; Ouyang, K.-X.; Zhang, X.-Q.; Liao, F.-X.; et al. Genome-Wide Analysis of the AP2/ERF Gene Family in Pennisetum glaucum and the Negative Role of PgRAV_01 in Drought Tolerance. Plant Physiol. Biochem. 2024, 216, 109112. [Google Scholar] [CrossRef]
- Rashid, M.; He, G.; Yang, G.; Hussain, J.; Yan, X. AP2/ERF Transcription Factor in Rice: Genome-Wide Canvas and Syntenic Relationships between Monocots and Eudicots. Evol. Bioinform. 2012, 8, 321–355. [Google Scholar] [CrossRef]
- Riaz, M.W.; Lu, J.; Shah, L.; Yang, L.; Chen, C.; Mei, X.D.; Xue, L.; Manzoor, M.A.; Abdullah, M.; Rehman, S.; et al. Expansion and Molecular Characterization of AP2/ERF Gene Family in Wheat (Triticum aestivum L.). Front. Genet. 2021, 12, 632155. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.-Y.; Chen, L.-H.; Fan, B.-L.; Xu, Z.; Wang, Q.; Zhao, B.-Y.; Gao, M.; Yuan, M.-H.; ul Qamar, M.T.; Jiang, Y.; et al. Integrative Multiomics Profiling of Passion Fruit Reveals the Genetic Basis for Fruit Color and Aroma. Plant Physiol. 2024, 194, 2491–2510. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.P.; Crabtree, J.; Zhao, Q.; Lorenzi, H.; Orvis, J.; Puiu, D.; Melake-Berhan, A.; Jones, K.M.; Redman, J.; Chen, G.; et al. Draft Genome Sequence of the Oilseed Species Ricinus communis. Nat. Biotechnol. 2010, 28, 951–953. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, J.; Cai, B.; Peng, R.-H.; Zhu, B.; Jin, X.-F.; Xue, Y.; Gao, F.; Fu, X.-Y.; Tian, Y.-S.; Zhao, W.; et al. Genome-Wide Analysis of the AP2/ERF Gene Family in Populus trichocarpa. Biochem. Biophys. Res. Commun. 2008, 371, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Han, S.; Wang, H.; Yu, M.; Ma, R.; Yu, Z. The Regulation of 1-Methylcyclopropene Treatment on the Subfamily Genes Expression of Ethylene Response Factors in Peaches during Storage. Acta Sci. Pol. Technol. Aliment. 2021, 20, 313–323. [Google Scholar] [CrossRef]
- Zhang, J.; Yin, X.-R.; Li, H.; Xu, M.; Zhang, M.-X.; Li, S.-J.; Liu, X.-F.; Shi, Y.-N.; Grierson, D.; Chen, K.-S. Ethylene Response Factor39-Myb8 Complex Regulates Low-Temperature-Induced Lignification of Loquat Fruit. J. Exp. Bot. 2020, 71, 3172–3184. [Google Scholar] [CrossRef]
- Soares, C.G.; do Prado, S.B.R.; Andrade, S.C.S.; Fabi, J.P. Systems Biology Applied to the Study of Papaya Fruit Ripening: The Influence of Ethylene on Pulp Softening. Cells 2021, 10, 2339. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shi, C.; Ge, P.; Li, F.; Zhu, L.; Wang, Y.; Tao, J.; Zhang, X.; Dong, H.; Gai, W.; et al. A 21-bp Indel in the Promoter of STP1 Selected during Tomato Improvement Accounts for Soluble Solid Content in Fruits. Hortic. Res. 2023, 10, uhad009. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped Blast and Psi-Blast: A New Generation of Protein Database Search Programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) V3: An Online Tool for the Display and Annotation of Phylogenetic and Other Trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the Graphical Presentation of Linkage Maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H.; et al. MCSCANX: A Toolkit for Detection and Evolutionary Analysis of Gene Synteny and Collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An Information Aesthetic for Comparative Genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An Upgraded Gene Feature Visualization Server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for In Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Li, K.; Cheng, Y.; Fang, C. OsDWARF10, Transcriptionally Repressed by OsSPL3, Regulates the Nutritional Metabolism of Polished rice. Front. Plant Sci. 2023, 14, 1322463. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Guo, Z.; Wang, K.; Wang, R.; Fang, C. Comparative Metabolomic Analysis Reveals the Role of OsHPL1 in the Cold-Induced Metabolic Changes in Rice. Plants 2023, 12, 2032. [Google Scholar] [CrossRef] [PubMed]
- Jessica, B.; Leif, V.; Christine, W.; Russel, P.B.; Craig, P.H.; Qi, L.; Lynn, E.; Sandra, E.S. HIP: A Method for High-Dimensional Multi-View Data Integration and Prediction Accounting for Subgroup Heterogeneity. Brief. Bioinform. 2024, 25, bbae470. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | ID | Chr No. | Protein Length (aa) | CDS (bp) | MW (Da) | pI | GRAVY |
|---|---|---|---|---|---|---|---|
| P_edulia010004144.g | PeAP2-1 | Chr1 | 678 | 2037 | 75,028 | 6.84 | −0.75 |
| P_edulia020006708.g | PeAP2-2 | Chr2 | 589 | 1770 | 63,971 | 7.8 | −0.39 |
| P_edulia030008406.g | PeAP2-3 | Chr3 | 510 | 1533 | 56,401 | 6.25 | −0.61 |
| P_edulia030008511.g | PeAP2-4 | Chr3 | 417 | 1254 | 46,541 | 9.72 | −0.82 |
| P_edulia040010127.g | PeAP2-5 | Chr4 | 546 | 1641 | 60,563 | 7.14 | −0.74 |
| P_edulia050011325.g | PeAP2-6 | Chr5 | 351 | 1056 | 39,533 | 5.16 | −0.66 |
| P_edulia060015442.g | PeAP2-7 | Chr6 | 682 | 2049 | 74,340 | 7.56 | −0.62 |
| P_edulia060015844.g | PeAP2-8 | Chr6 | 721 | 2166 | 78,270 | 6.54 | −0.71 |
| P_edulia070018496.g | PeAP2-9 | Chr7 | 310 | 933 | 34,092 | 6.18 | −0.66 |
| P_edulia080019291.g | PeAP2-10 | Chr8 | 411 | 1236 | 46,358 | 8.36 | −0.68 |
| P_edulia010000572.g | PeDREB-1 | Chr1 | 236 | 711 | 25,967 | 5.01 | −0.42 |
| P_edulia010001596.g | PeDREB-2 | Chr1 | 1209 | 3630 | 133,140 | 10.39 | −0.24 |
| P_edulia010003920.g | PeDREB-3 | Chr1 | 230 | 693 | 25,106 | 4.82 | −0.64 |
| P_edulia040009646.g | PeDREB-4 | Chr4 | 240 | 723 | 26,964 | 6.51 | −0.55 |
| P_edulia040010723.g | PeDREB-5 | Chr4 | 310 | 933 | 33,942 | 8.35 | −0.45 |
| P_edulia040010839.g | PeDREB-6 | Chr4 | 234 | 705 | 26,077 | 5.22 | −0.75 |
| P_edulia040011090.g | PeDREB-7 | Chr4 | 190 | 573 | 20,815 | 9.71 | −0.69 |
| P_edulia050011280.g | PeDREB-8 | Chr5 | 286 | 861 | 31,335 | 5.22 | −0.71 |
| P_edulia050011897.g | PeDREB-9 | Chr5 | 218 | 657 | 24,004 | 6.73 | −0.77 |
| P_edulia050012013.g | PeDREB-10 | Chr5 | 241 | 726 | 27,470 | 4.87 | −0.87 |
| P_edulia050012014.g | PeDREB-11 | Chr5 | 243 | 732 | 26,281 | 4.67 | −0.54 |
| P_edulia050012164.g | PeDREB-12 | Chr5 | 297 | 894 | 33,242 | 8.44 | −0.57 |
| P_edulia060013532.g | PeDREB-13 | Chr6 | 439 | 1320 | 48,352 | 8.76 | −0.60 |
| P_edulia060014565.g | PeDREB-14 | Chr6 | 215 | 648 | 23,342 | 4.66 | −0.43 |
| P_edulia060014877.g | PeDREB-15 | Chr6 | 215 | 648 | 23,376 | 4.56 | −0.48 |
| P_edulia060015588.g | PeDREB-16 | Chr6 | 292 | 879 | 32,099 | 5.74 | −0.41 |
| P_edulia070016972.g | PeDREB-17 | Chr7 | 194 | 585 | 20,703 | 4.84 | −0.39 |
| P_edulia070017934.g | PeDREB-18 | Chr7 | 287 | 864 | 31,127 | 4.48 | −0.71 |
| P_edulia070018079.g | PeDREB-19 | Chr7 | 215 | 648 | 23,856 | 4.26 | −0.70 |
| P_edulia070018189.g | PeDREB-20 | Chr7 | 251 | 756 | 27,266 | 4.74 | −0.55 |
| P_edulia070018581.g | PeDREB-21 | Chr7 | 244 | 735 | 26,542 | 4.99 | −0.47 |
| P_edulia090020954.g | PeDREB-22 | Chr9 | 262 | 789 | 28,430 | 5.14 | −0.55 |
| P_edulia090021123.g | PeDREB-23 | Chr9 | 208 | 627 | 22,834 | 4.27 | −0.46 |
| P_eduliaContig200022650.g | PeDREB-24 | Contig | 440 | 693 | 48,313 | 6.84 | −0.59 |
| P_eduliaContig30022789.g | PeDREB-25 | Contig | 256 | 1123 | 33,592 | 9.83 | −0.55 |
| P_edulia010000150.g | PeERF-1 | Chr1 | 251 | 1176 | 27,267 | 4.65 | −0.44 |
| P_edulia010001731.g | PeERF-2 | Chr1 | 391 | 1107 | 43,201 | 5.54 | −0.89 |
| P_edulia010001854.g | PeERF-3 | Chr1 | 368 | 966 | 41,985 | 4.73 | −0.59 |
| P_edulia010001961.g | PeERF-4 | Chr1 | 321 | 1248 | 35,875 | 4.99 | −0.45 |
| P_edulia010002055.g | PeERF-5 | Chr1 | 415 | 732 | 46,607 | 6.54 | −0.64 |
| P_edulia010002171.g | PeERF-6 | Chr1 | 243 | 597 | 26,686 | 6.25 | −0.93 |
| P_edulia010003619.g | PeERF-7 | Chr1 | 127 | 900 | 14,151 | 9.72 | −0.85 |
| P_edulia010004885.g | PeERF-8 | Chr1 | 299 | 651 | 33,729 | 7.14 | −0.75 |
| P_edulia010005010.g | PeERF-9 | Chr1 | 216 | 651 | 23,667 | 7.33 | −0.75 |
| P_edulia010005188.g | PeERF-10 | Chr1 | 249 | 750 | 28,127 | 4.79 | −0.76 |
| P_edulia010005329.g | PeERF-11 | Chr1 | 216 | 645 | 23,653 | 8.07 | −0.84 |
| P_edulia020006460.g | PeERF-12 | Chr2 | 382 | 423 | 42,764 | 5.06 | −0.58 |
| P_edulia030007663.g | PeERF-13 | Chr3 | 140 | 747 | 15,549 | 10.28 | −0.72 |
| P_edulia030007664.g | PeERF-14 | Chr3 | 257 | 774 | 28,824 | 6.51 | −0.78 |
| P_edulia030007668.g | PeERF-15 | Chr3 | 248 | 639 | 27,691 | 9.12 | −0.71 |
| P_edulia030007669.g | PeERF-16 | Chr3 | 212 | 618 | 23,722 | 4.68 | −0.68 |
| P_edulia030009441.g | PeERF-17 | Chr3 | 205 | 678 | 23,055 | 5.95 | −0.59 |
| P_edulia040009825.g | PeERF-18 | Chr4 | 225 | 945 | 24,644 | 7.09 | −0.82 |
| P_edulia040010539.g | PeERF-19 | Chr4 | 314 | 978 | 35,157 | 5.89 | −0.72 |
| P_edulia040010561.g | PeERF-20 | Chr4 | 325 | 543 | 36,419 | 6.68 | −0.86 |
| P_edulia040010637.g | PeERF-21 | Chr4 | 274 | 825 | 30,768 | 6.79 | −0.56 |
| P_edulia040010735.g | PeERF-22 | Chr4 | 180 | 957 | 20,655 | 7.25 | −0.53 |
| P_edulia040011072.g | PeERF-23 | Chr4 | 318 | 792 | 35,009 | 5 | −0.71 |
| P_edulia040011079.g | PeERF-24 | Chr4 | 267 | 804 | 29,686 | 7.95 | −0.31 |
| P_edulia050011662.g | PeERF-25 | Chr4 | 263 | 438 | 28,058 | 8.62 | −0.96 |
| P_edulia050011732.g | PeERF-26 | Chr5 | 145 | 558 | 16,267 | 7.51 | −0.77 |
| P_edulia050011872.g | PeERF-27 | Chr5 | 275 | 597 | 30,447 | 8.6 | −0.50 |
| P_edulia050011874.g | PeERF-28 | Chr5 | 185 | 828 | 20,952 | 8.03 | −0.63 |
| P_edulia050012238.g | PeERF-29 | Chr5 | 198 | 897 | 21,956 | 9.5 | −0.54 |
| P_edulia050012239.g | PeERF-30 | Chr5 | 337 | 1014 | 37,775 | 5.96 | −0.60 |
| P_edulia050012240.g | PeERF-31 | Chr5 | 298 | 765 | 32,899 | 8.61 | −0.43 |
| P_edulia060013160.g | PeERF-32 | Chr5 | 254 | 1203 | 27,510 | 9.13 | −0.82 |
| P_edulia060013663.g | PeERF-33 | Chr6 | 400 | 1203 | 44,660 | 4.45 | −0.64 |
| P_edulia060013843.g | PeERF-34 | Chr6 | 400 | 1071 | 43,717 | 8.58 | −0.63 |
| P_edulia060014975.g | PeERF-35 | Chr6 | 356 | 552 | 38,670 | 8.07 | −0.37 |
| P_edulia060015066.g | PeERF-36 | Chr6 | 183 | 1299 | 20,099 | 10.63 | −0.69 |
| P_edulia060015079.g | PeERF-37 | Chr6 | 432 | 1026 | 47,198 | 8.45 | −0.78 |
| P_edulia060015213.g | PeERF-38 | Chr6 | 234 | 705 | 26,272 | 5.24 | −0.65 |
| P_edulia060015996.g | PeERF-39 | Chr6 | 342 | 570 | 36,653 | 6.31 | −0.68 |
| P_edulia070017176.g | PeERF-40 | Chr7 | 189 | 615 | 21,274 | 10.54 | −0.73 |
| P_edulia070017525.g | PeERF-41 | Chr7 | 204 | 726 | 22,966 | 7.66 | −0.54 |
| P_edulia070017902.g | PeERF-42 | Chr7 | 190 | 573 | 21,462 | 10.11 | −0.77 |
| P_edulia070018289.g | PeERF-43 | Chr7 | 187 | 564 | 20,782 | 9.37 | −0.62 |
| P_edulia070018452.g | PeERF-44 | Chr7 | 241 | 1161 | 25,808 | 6.54 | −0.68 |
| P_edulia080019076.g | PeERF-45 | Chr8 | 386 | 504 | 42,800 | 4.76 | −0.70 |
| P_edulia080019171.g | PeERF-46 | Chr8 | 167 | 441 | 18,581 | 7.32 | −0.68 |
| P_edulia090020522.g | PeERF-47 | Chr9 | 118 | 357 | 13,273 | PI | −0.66 |
| P_edulia090020764.g | PeERF-48 | Chr9 | 393 | 1182 | 16,176 | 4.62 | −0.63 |
| P_edulia090020847.g | PeERF-49 | Chr9 | 194 | 585 | 43,325 | 7.56 | −0.63 |
| P_edulia090021477.g | PeERF-50 | Chr9 | 205 | 618 | 22,911 | 6.18 | −0.74 |
| P_edulia090022050.g | PeERF-51 | Chr9 | 321 | 966 | 22,228 | 8.36 | −0.42 |
| P_eduliaContig100022618.g | PeERF-52 | Contig | 230 | 543 | 34,318 | 7.38 | −0.78 |
| P_eduliaContig240022563.g | PeERF-53 | Contig | 180 | 1188 | 25,530 | 7.8 | −0.49 |
| P_edulia020006614.g | PeRAV-1 | Chr2 | 361 | 1086 | 41,452 | 5.16 | −0.64 |
| P_edulia010005093.g | PeSoloist-1 | Chr1 | 231 | 696 | 25,920 | 4.67 | −0.78 |
| P_edulia060014445.g | PeSoloist-2 | Chr6 | 229 | 690 | 25,703 | 9.13 | −0.88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, L.; Zhang, L.; Gu, R.; Ni, S.; Yu, J.; Gao, Y.; Fang, C. Genome-Wide Identification and Functional Analysis of AP2/ERF Gene Family in Passiflora edulis Sims. Plants 2025, 14, 645. https://doi.org/10.3390/plants14050645
Luo L, Zhang L, Gu R, Ni S, Yu J, Gao Y, Fang C. Genome-Wide Identification and Functional Analysis of AP2/ERF Gene Family in Passiflora edulis Sims. Plants. 2025; 14(5):645. https://doi.org/10.3390/plants14050645
Chicago/Turabian StyleLuo, Lanjun, Liping Zhang, Ronghao Gu, Shihao Ni, Jingyao Yu, Yachao Gao, and Chuanying Fang. 2025. "Genome-Wide Identification and Functional Analysis of AP2/ERF Gene Family in Passiflora edulis Sims" Plants 14, no. 5: 645. https://doi.org/10.3390/plants14050645
APA StyleLuo, L., Zhang, L., Gu, R., Ni, S., Yu, J., Gao, Y., & Fang, C. (2025). Genome-Wide Identification and Functional Analysis of AP2/ERF Gene Family in Passiflora edulis Sims. Plants, 14(5), 645. https://doi.org/10.3390/plants14050645