
Interpopulational Variation in Cyclotide Production in Heavy-Metal-Treated Pseudometallophyte (Viola tricolor L.)
 , , , ,
, , , , 
Abstract
1. Introduction
2. Results
2.1. Similar Accumulation of Zn and Pb in M and NM Specimens of Viola tricolor
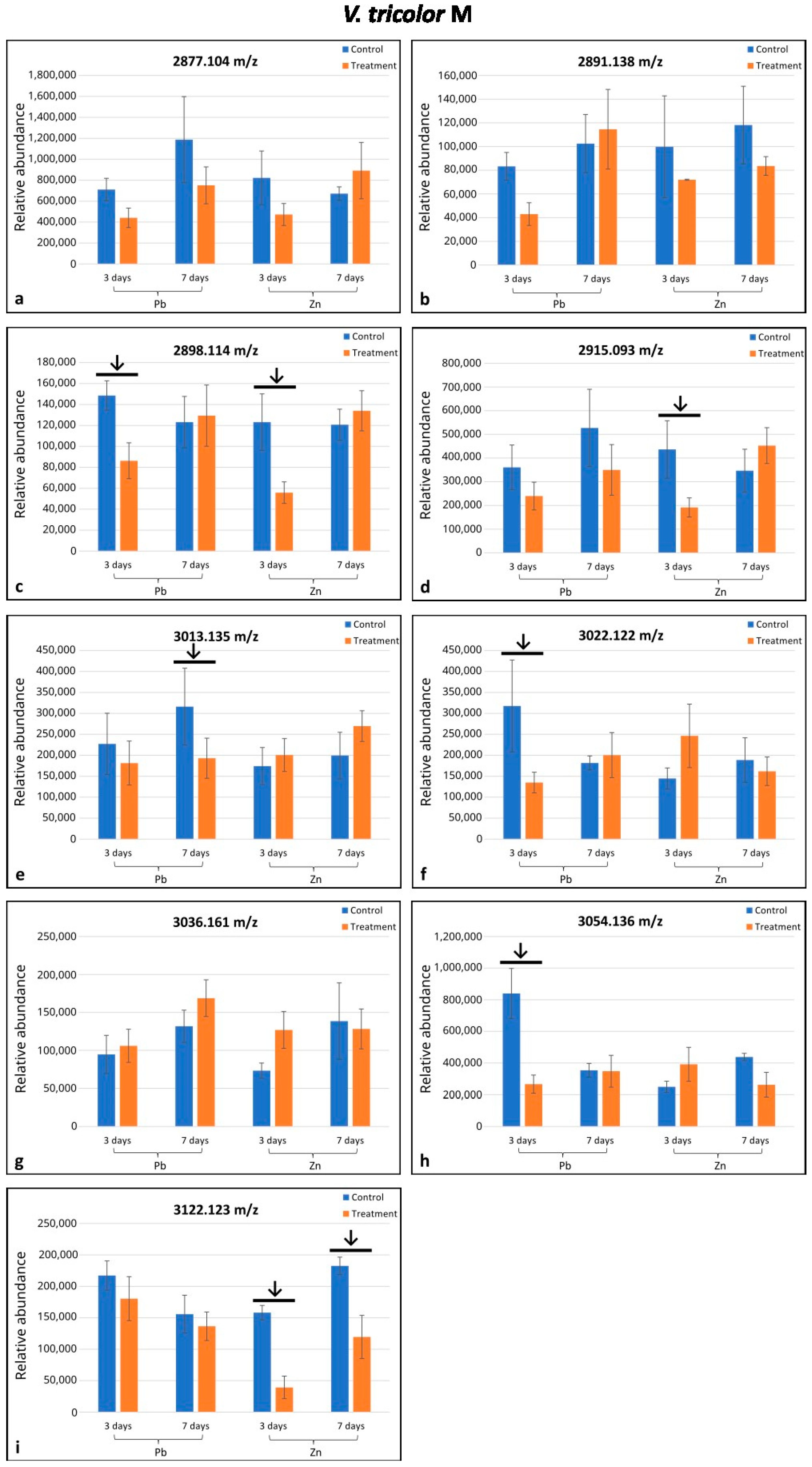
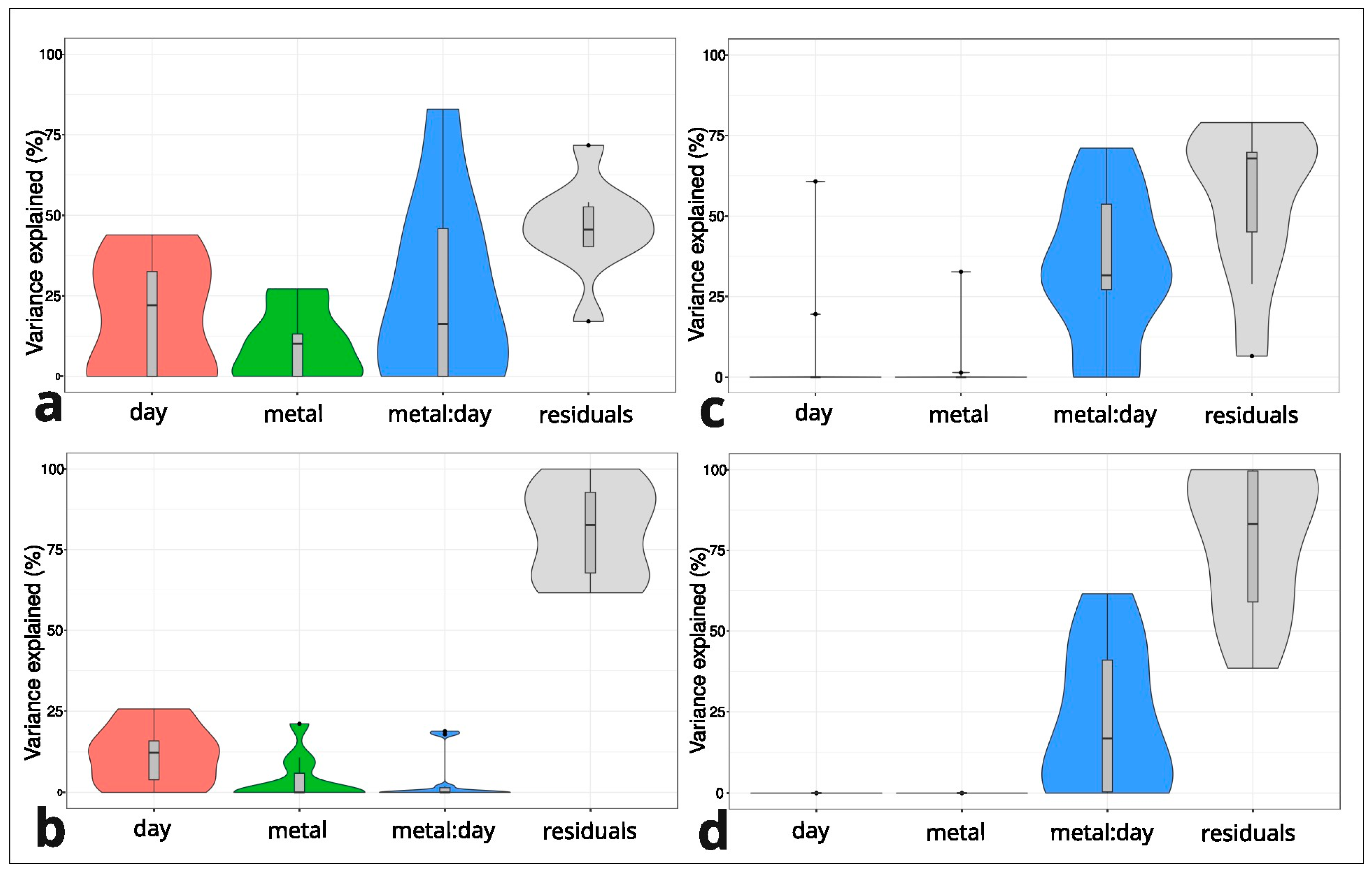
2.2. Variation in Cyclotide Production Between M and NM Individuals of Viola tricolor Under Treatment with Heavy Metals
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Quantification of Content of Heavy Metals in Plants by Atomic Absorption Spectroscopy (AAS)
4.3. Cyclotide Detection by Matrix Assisted Laser Desorption and Ionization (MALDI-MS)
4.4. Statistical Analyses
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bothe, H.; Vogel-Mikuš, K.; Pongrac, P.; Matevž, L.; Stepic, N.; Pelicon, P.; Vavpetič, P.; Jeromel, L.; Regvar, M. Metallophyte status of violets of the section Melanium. Chemosphere 2013, 93, 1844–1855. [Google Scholar]
- Bothe, H.; Słomka, A. Divergent biology of facultative heavy metal plants. J. Plant Physiol. 2017, 29, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Sychta, K.; Słomka, A.; Suski, S.; Fiedor, E.; Gregoraszczuk, E.; Kuta, E. Suspended cells of metallicolous and nonmetallicolous Viola species tolerate, accumulate and detoxify zinc and lead. Plant Physiol. Biochem. 2018, 132, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Sychta, K.; Słomka, A.; Kuta, E. Garden pansy (Viola × wittrockiana Gams.)—A good candidate for the revitalisation of polluted areas. Plant. Soil Environ. 2020, 66, 272–280. [Google Scholar] [CrossRef]
- Sychta, K.; Słomka, A.; Sliwinska, E.; Migdałek, G.; Kuta, E. From cells highly tolerant to Zn and Pb to fully fertile plants–selection of tolerant lines with in vitro culture. Plant Physiol. Biochem. 2020, 146, 231–237. [Google Scholar] [CrossRef]
- Miszczak, S.; Sychta, K.; Dresler, S.; Kurdziel, A.; Hanaka, A.; Słomka, A. Innate, high tolerance to zinc and lead in violets confirmed at the suspended cell level. Cells 2022, 11, 2355. [Google Scholar] [CrossRef]
- Shu, H.; Zhang, J.; Liu, F.; Bian, C.; Liang, J.; Liang, J.; Lin, Z.; Shu, W.; Li, J.; Shi, Q.; et al. Comparative transcriptomic studies on a cadmium hyperaccumulator Viola baoshanensis and its non-tolerant counterpart V. inconspicua. Int. J. Mol. Sci. 2019, 20, 1906. [Google Scholar] [CrossRef]
- Wojciechowski, K. Morphological and anatomical differences and reaction to zinc of the pseudometallophyte Viola tricolor L. grown hydroponically. Master’s Thesis, Jagiellonian University, Cracow, Poland, 2018. [Google Scholar]
- Kwiatkowska, M.; Żabicka, J.; Migdałek, G.; Żabicki, P.; Cubała, M.; Bohdanowicz, J.; Słomka, A.; Jędrzejczyk-Korycińska, M.; Sliwinska, E.; Sychta, K.; et al. Comprehensive characteristics and genetic diversity of the endemic Australian Viola banksii (section Erpetion, Violaceae). Aust. J. Bot. 2019, 67, 81–98. [Google Scholar] [CrossRef]
- Słomka, A.; Gubernat, M.; Pliszko, A.; Bothe, H. The unusual property of the sand violet, Viola rupestris, to cope with heavy metal toxicity. Flora 2020, 271, 151663. [Google Scholar] [CrossRef]
- Hildebrandt, U.; Hoef-Emden, K.; Backhausen, S.; Bothe, H.; Bożek, M.; Siuta, A.; Kuta, E. The rare, endemic zinc violets of Central Europe originate from Viola lutea Huds. Plant Syst. Evol. 2006, 257, 205–222. [Google Scholar] [CrossRef]
- Słomka, A.; Kuta, E.; Szarek-Łukaszewska, G.; Godzik, B.; Kapusta, P.; Tylko, G.; Bothe, H. Violets of the section Melanium, their colonization by arbuscular mycorrhizal fungi and their occurrence on heavy metal heaps. J. Plant Physiol. 2011, 168, 1191–1199. [Google Scholar] [CrossRef]
- Słomka, A.; Godzik, B.; Szarek-Łukaszewska, G.; Shuka, L.; Hoef-Emden, K.; Bothe, H. Albanian violets of the section Melanium, their morphological variability, genetic similarity and their adaptations to serpentine or chalk soils. J. Plant Physiol. 2015, 174, 110–123. [Google Scholar] [CrossRef] [PubMed]
- Nawrot, R.; Barylski, J.; Nowicki, G.; Broniarczyk, J.; Buchwald, W.; Goździcka-Józefiak, A. Plant antimicrobial peptides. Folia Microbiol. 2024, 59, 181–196. [Google Scholar] [CrossRef] [PubMed]
- Van der Weerden, N.L.; Anderson, M.A. Plant defensins: Common fold, multiple functions. Fungal Biol. Rev. 2013, 26, 121–131. [Google Scholar] [CrossRef]
- Mirouze, M.; Sels, J.; Richard, O.; Czernic, P.; Loubet, S.; Jacquier, A.; Francois, I.E.; Cammue, B.P.; Lebrun, M.; Berthomieu, P.; et al. A putative novel role for plant defensins: A defensin from the zinc hyper-accumulating plant, Arabidopsis halleri, confers zinc tolerance. Plant J. 2006, 47, 329–342. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, Z.; Ranwez, V.; Fizames, C.; Marquès, L.; Le Martret, B.; Alasimone, J.; Godé, C.; Lacombe, E.; Castillo, T.; Saumitou-Laprade, P.; et al. Plant defensin type 1 (PDF1): Protein promiscuity and expression variation within the Arabidopsis genus shed light on zinc tolerance acquisition in Arabidopsis halleri. New Phytol. 2013, 200, 820–833. [Google Scholar] [CrossRef]
- Marquès, L.; Oomen, R.J. On the way to unravel zinc hyperaccumulation in plants: A mini review. Metallomics 2011, 3, 1265–1270. [Google Scholar] [CrossRef]
- Luo, J.S.; Gu, T.; Yang, Y.; Zhang, Z. A non-secreted plant defensin AtPDF2.6 conferred cadmium tolerance via its chelation in Arabidopsis. Plant Mol. Biol. 2019, 100, 561–569. [Google Scholar] [CrossRef]
- Burman, R.; Yeshak, M.Y.; Larsson, S.; Craik, D.J.; Rosengren, K.J.; Göransson, U. Distribution of circular proteins in plants: Large-scale mapping of cyclotides in the Violaceae. Front. Plant Sci. 2015, 6, 855. [Google Scholar] [CrossRef]
- Jennings, C.; West, J.; Waine, C.; Craik, D.; Anderson, M.A. Biosynthesis and insecticidal properties of plant cyclotides: The cyclic knotted proteins from Oldenlandia affinis. Proc. Natl. Acad. Sci. USA 2001, 98, 10614–10619. [Google Scholar] [CrossRef]
- Slazak, B.; Kapusta, M.; Strömstedt, A.A.; Słomka, A.; Krychowiak, M.; Shariatgorji, M.; Andrén, P.E.; Bohdanowicz, J.; Kuta, E.; Göransson, U. How does the sweet violet (Viola odorata L.) fight pathogens and pests–cyclotides as a comprehensive plant host defense system? Front. Plant Sci. 2018, 9, 1296. [Google Scholar] [CrossRef] [PubMed]
- Cao, P.; Yang, Y.; Uche, F.I.; Hart, S.R.; Li, W.W.; Yuan, C. Coupling plant-derived cyclotides to metal surfaces: An antibacterial and antibiofilm study. Int. J. Mol. Sci. 2018, 19, 793. [Google Scholar] [CrossRef] [PubMed]
- Skjeldal, L.; Gran, L.; Sletten, K.; Volkman, B.F. Refined structure and metal binding site of the Kalata B1 peptide. Arch. Biochem. Biophys. 2002, 399, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Shenkarev, Z.O.; Nadezhdin, K.D.; Lyukmanova, E.N.; Sobol, V.A.; Skjeldal, L.; Arseniev, A.S. Divalent cation coordination and mode of membrane interaction in cyclotides: NMR spatial structure of ternary complex Kalata B7/Mn2+/DPC micelle. J. Inorg. Biochem. 2008, 102, 1246–1256. [Google Scholar] [CrossRef]
- Sychta, K.; Słomka, A.; Shariatgorji, R.; Andrén, P.E.; Samardakiewicz, S.; Göransson, U.; Slazak, B. The involvement of cyclotides in the heavy metal tolerance of Viola spp. Sci. Rep. 2024, 14, 19306. [Google Scholar] [CrossRef]
- Hellinger, R.; Koehbach, J.; Soltis, D.E.; Carpenter, E.J.; Wong, G.K.S.; Gruber, C.W. Peptidomics of circular cysteine-rich plant peptides: Analysis of the diversity of cyclotides from Viola tricolor by transcriptome and proteome mining. J. Proteome Res. 2015, 14, 4851–4862. [Google Scholar] [CrossRef]
- Slazak, B.; Kaltenböck, K.; Steffen, K.; Rogala, M.; Rodriguez-Rodriguez, P.; Nilsson, A.; Shariatgorji, R.; Andrén, P.E.; Göransson, U. Cyclotide host-defense tailored for species and environments in violets from the Canary Islands. Sci. Rep. 2021, 11, 12452. [Google Scholar] [CrossRef]
- Kawalec, P. Badania Biosystematyczne nad Viola tricolor L. z Hałd Cynkowo-Ołowiowych w Okolicach Olkusza. Master’s Thesis, Jagiellonian University in Krakow, Cracow, Poland, 2008. [Google Scholar]
- Camarero, J.A. Cyclotides, a versatile ultrastable micro-protein scaffold for biotechnological applications. Bioorganic Med. Chem. Lett. 2017, 27, 5089–5099. [Google Scholar]
- Słomka, A.; Sutkowska, A.; Szczepaniak, M.; Malec, P.; Mitka, J.; Kuta, E. Increased genetic diversity of Viola tricolor L. (Violaceae) in metal-polluted environments. Chemosphere 2011, 83, 435–442. [Google Scholar] [CrossRef]
- Slazak, B.; Jędrzejska, A.; Badyra, B.; Shariatgorji, R.; Nilsson, A.; Andrén, P.E.; Göransson, U. The influence of plant stress hormones and biotic elicitors on cyclotide production in Viola uliginosa cell suspension cultures. Plants 2022, 11, 1876. [Google Scholar] [CrossRef]
- Slazak, B.; Jędrzejska, A.; Badyra, B.; Sybilska, A.; Lewandowski, M.; Kozak, M.; Kapusta, M.; Shariatgorji, R.; Nilsson, A.; Andrén, P.E.; et al. The involvement of cyclotides in mutual interactions of violets and the two-spotted spider mite. Sci. Rep. 2022, 12, 1914. [Google Scholar] [CrossRef]
- Busoms, S.; Pérez-Martín, L.; Llimós, M.; Poschenrieder, C.; Martos, S. Genome-wide association study reveals key genes for differential lead accumulation and tolerance in natural Arabidopsis thaliana accessions. Front. Plant Sci. 2021, 12, 689316. [Google Scholar] [CrossRef] [PubMed]
- De Agostini, A.; Cogoni, A.; Cortis, P.; Vacca, A.; Becerril, J.M.; Hernández, A.; Esteban, R. Heavy metal tolerance strategies in metallicolous and non-metallicolous populations of mosses: Insights of γ+ β-tocopherol regulatory role. Environ. Exp. Bot. 2022, 194, 104738. [Google Scholar] [CrossRef]
- Boquete, M.T.; Schmid, M.W.; Wagemaker, N.C.; Carey, S.B.; McDaniel, S.F.; Richards, C.L.; Alonso, C. Molecular basis of intraspecific differentiation for heavy metal tolerance in the copper moss Scopelophila cataractae. Environ. Exp. Bot. 2022, 201, 104970. [Google Scholar] [CrossRef]
- Feng, D.; Wang, R.; Sun, X.; Liu, P.; Tang, J.; Zhang, C.; Liu, H. Heavy metal stress in plants: Ways to alleviate with exogenous substances. Sci. Total Environ. 2023, 897, 165397. [Google Scholar] [CrossRef] [PubMed]
- Babst-Kostecka, A.; Przybyłowicz, W.J.; Seget, B.; Mesjasz-Przybyłowicz, J. Zinc allocation to and within Arabidopsis halleri seeds: Different strategies of metal homeostasis in accessions under divergent selection pressure. Plant Environ. Interact. 2020, 1, 207–220. [Google Scholar] [CrossRef]
- Słomka, A. Microevolutionary processes in heavy metal contaminated sites based on Viola tricolor L. (Melanium Ging. section, Violaceae). Ph.D. Dissertation, Jagiellonian University in Cracow, Cracow, Poland, 2009. [Google Scholar]
- Krygier, E.; Molenda, D.; Saładziak, A. Katalog zabytków budownictwa przemysłowego w Polsce; Powiat Olkusz—Województwo krakowskie; Wydawnictwo PAN: Wrocław, Poland, 1971. [Google Scholar]
- Mackey, E.A.; Becker, D.A.; Oflaz, R.D.; Greenberg, R.R.; Lindstrom, R.M.; Lee, L.Y.; Wood, L.J.; Long, S.E.; Kelly, W.R.; Mann, J.L.; et al. Certification of NIST Standard Reference Material 1575a pine needles and results of an international laboratory comparison. J. Res. NIST 2004, 3, 130. [Google Scholar]
- Källback, P.; Nilsson, A.; Shariatgorji, M.; Andrén, P.E. msIQuant—Quantitation software for mass spectrometry imaging enabling fast access, visualization, and analysis of large data sets. Anal. Chem. 2016, 88, 4346–4353. [Google Scholar] [CrossRef]
- Mercier, C.; Truntzer, C.; Pecqueur, D.; Gimeno, J.P.; Belz, G.; Roy, P. Mixed-model of ANOVA for measurement reproducibility in proteomics. J. Proteom. 2009, 72, 974–981. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Heavy Metal Concentration (mg kg−1) | Bioaccumulation Factor | ||||
|---|---|---|---|---|---|
| Population Days of Treatment | Pb | Zn | Pb | Zn | |
| M | 3 | 2.835 ± [0.938] * | 284.206 ± [110.516] | 0.013 ± [0.004] * | 4.372 ± [1.700] |
| 7 | 277.360 ± [108.281] | 601.721 ± [197.362] | 1.339 ± [0.523] | 9.257 ± [3.036] | |
| NM | 3 | 155.818 ± [58.976] | 443.771 ± [139.720] | 0.752 ± [0.284] | 6.827 ± [2.149] |
| 7 | 199.207 ± [54.611] | 705.695 ± [358.215] | 0.962 ± [0.263] | 10.856 ± [5.511] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miszczak, R.; Slazak, B.; Sychta, K.; Göransson, U.; Nilsson, A.; Słomka, A. Interpopulational Variation in Cyclotide Production in Heavy-Metal-Treated Pseudometallophyte (Viola tricolor L.). Plants 2025, 14, 471. https://doi.org/10.3390/plants14030471
Miszczak R, Slazak B, Sychta K, Göransson U, Nilsson A, Słomka A. Interpopulational Variation in Cyclotide Production in Heavy-Metal-Treated Pseudometallophyte (Viola tricolor L.). Plants. 2025; 14(3):471. https://doi.org/10.3390/plants14030471
Chicago/Turabian StyleMiszczak, Rebecca, Blazej Slazak, Klaudia Sychta, Ulf Göransson, Anna Nilsson, and Aneta Słomka. 2025. "Interpopulational Variation in Cyclotide Production in Heavy-Metal-Treated Pseudometallophyte (Viola tricolor L.)" Plants 14, no. 3: 471. https://doi.org/10.3390/plants14030471
APA StyleMiszczak, R., Slazak, B., Sychta, K., Göransson, U., Nilsson, A., & Słomka, A. (2025). Interpopulational Variation in Cyclotide Production in Heavy-Metal-Treated Pseudometallophyte (Viola tricolor L.). Plants, 14(3), 471. https://doi.org/10.3390/plants14030471