


Genome-Wide Identification and Expression Analysis of PkNRT Gene Family in Korean Pine (Pinus koraiensis)

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Characterization of the PkNRT Gene Family Members in Korean Pine
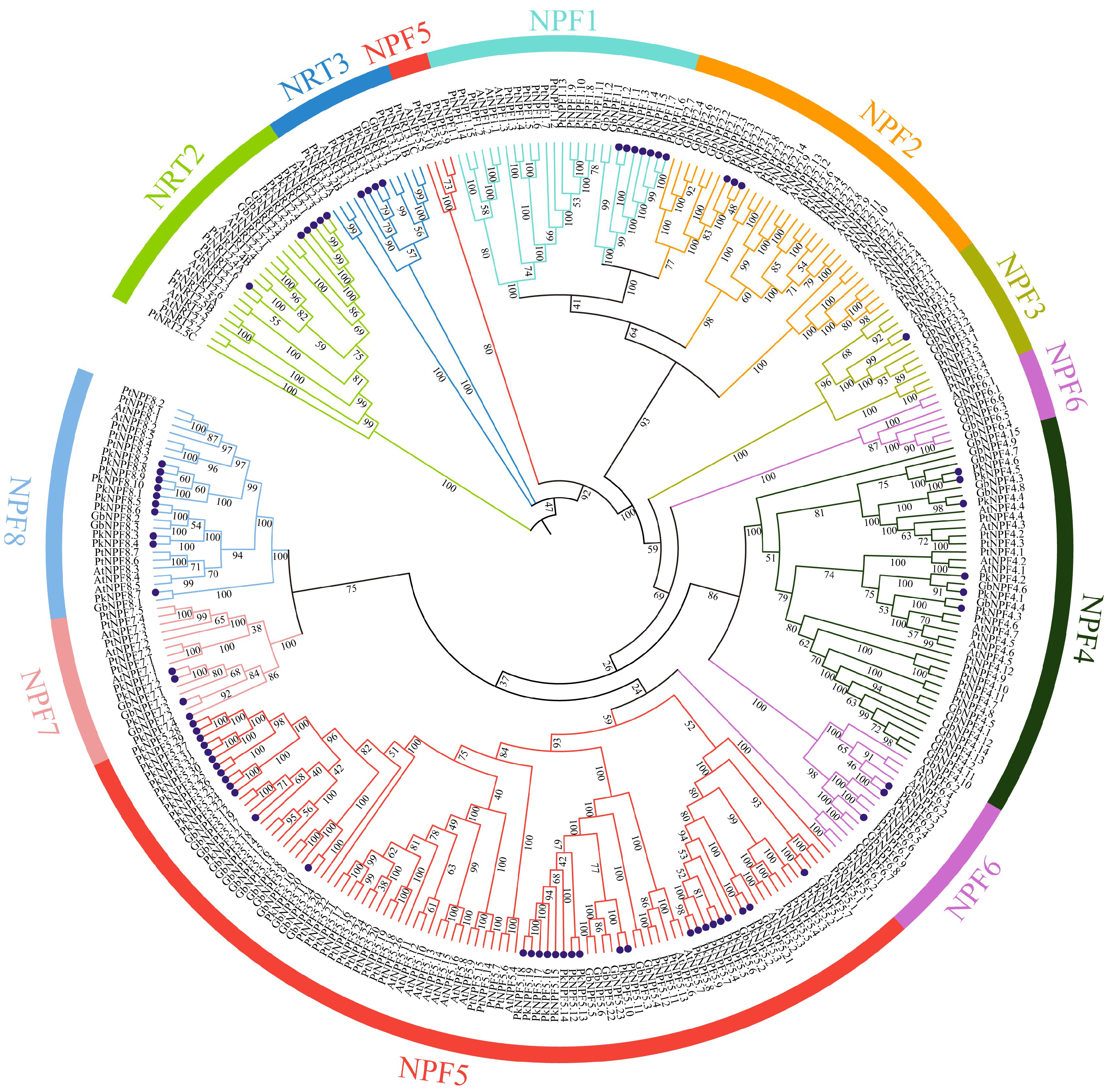
2.2. Phylogenetic Analysis of PkNRTs
2.3. Motifs, Conserved Domain, and Gene Structure Analysis of PkNRTs
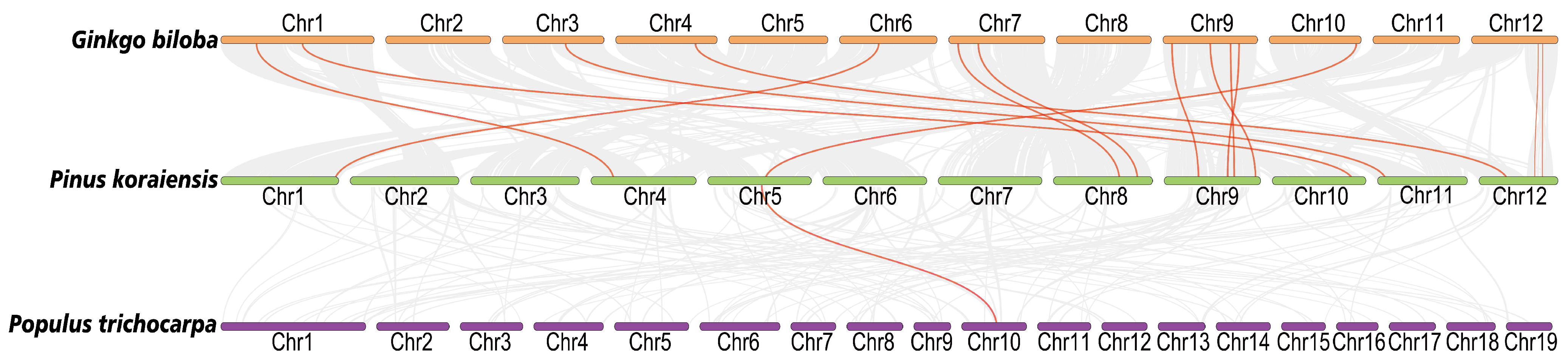
2.4. Analysis of Chromosomal Distribution and Collinearity of PkNRTs
2.5. Characterization of CREs in the Promoter Regions of PkNRTs
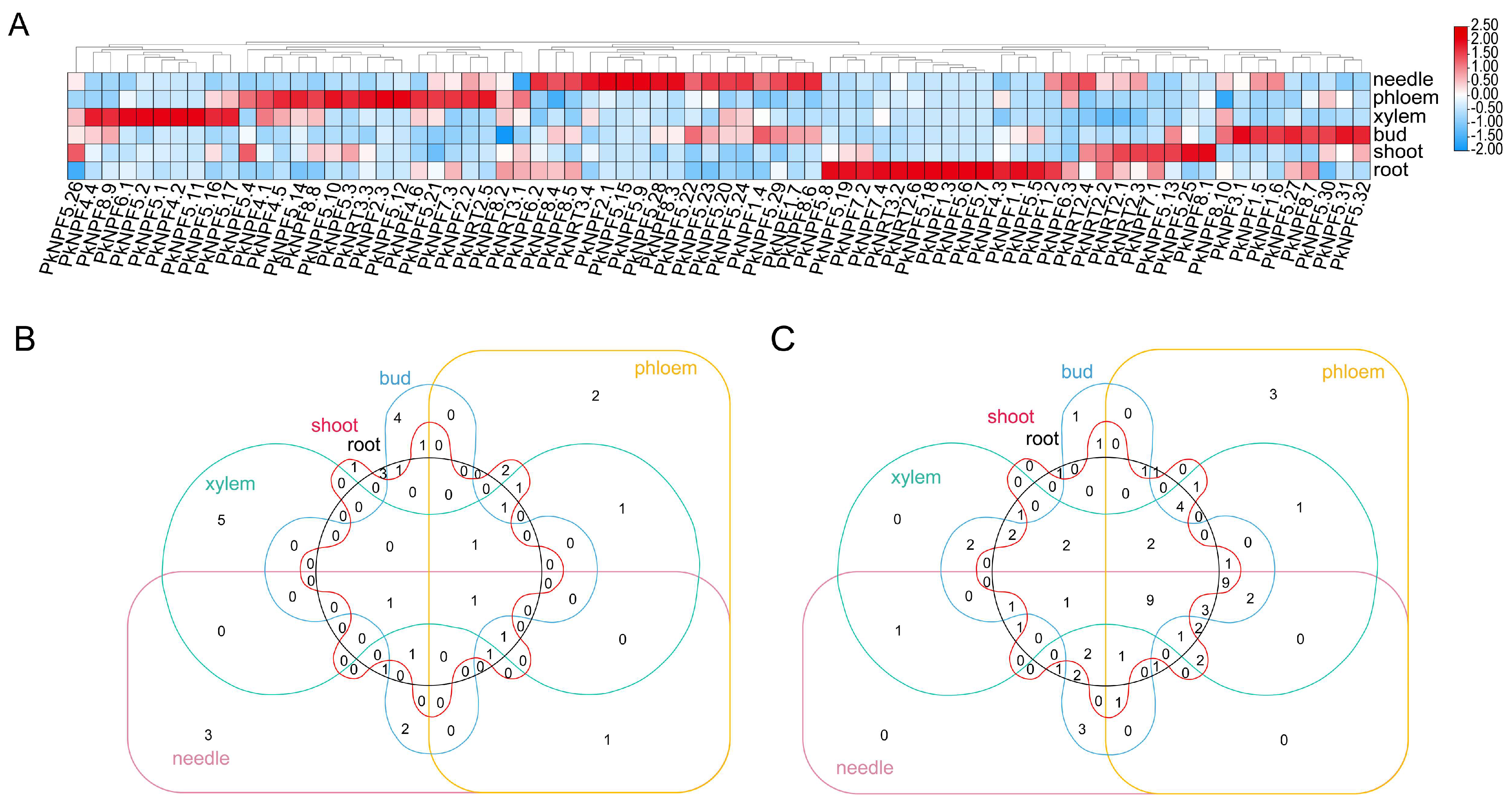
2.6. Transcript Abundance Analysis for Different Tissues in PkNRTs
2.7. Effects of N Treatment on Growth of Korean Pine Seedlings
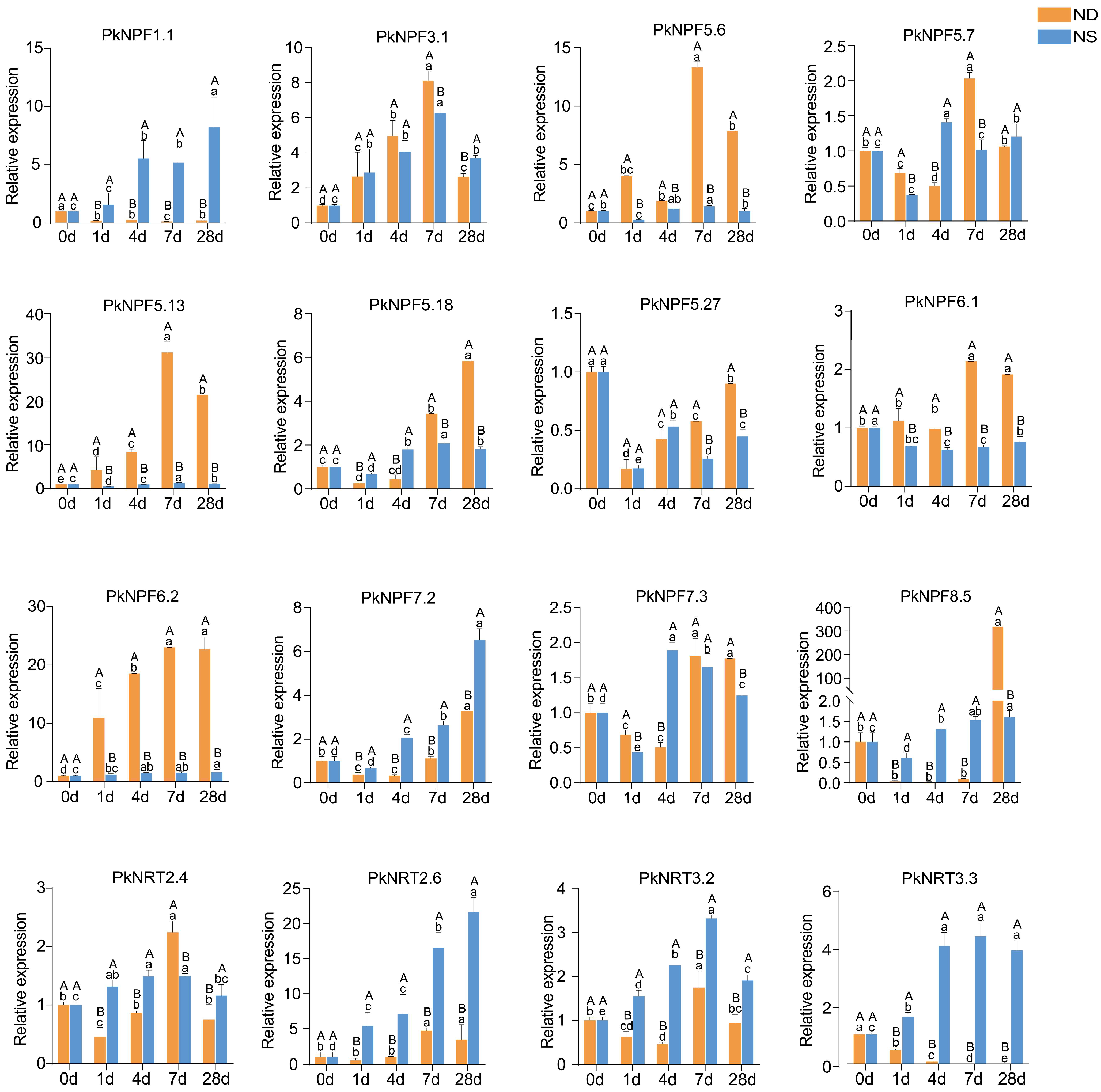
2.8. Expression Profiles of PkNRTs Under Different N Conditions
3. Discussion
4. Materials and Methods
4.1. Identification of NRT Gene Family in Korean Pine
4.2. Phylogenetic Analysis and Structural Characterization of PkNRTs
4.3. Colinearity Analysis and Calculation of Ka/Ks Values of PkNRTs
4.4. Analysis of CREs in the Promoters of PkNRTs
4.5. Transcriptome Sequencing for Tissue-Specific Abundance Analysis
4.6. Plant Materials and Treatments
4.7. Seedling Biomass and Root Morphology
4.8. Quantitative Real-Time PCR Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gaudinier, A.; Rodriguez-Medina, J.; Zhang, L.; Olson, A.; Liseron-Monfils, C.; Bågman, A.-M.; Foret, J.; Abbitt, S.; Tang, M.; Li, B.; et al. Transcriptional regulation of nitrogen-associated metabolism and growth. Nature 2018, 563, 259–264. [Google Scholar] [CrossRef]
- Luo, L.; Zhang, Y.; Xu, G. How does nitrogen shape plant architecture? J. Exp. Bot. 2020, 71, 4415–4427. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.-Q.; Lin, H.-X. Higher yield with less nitrogen fertilizer. Nat. Plants 2020, 6, 1078–1079. [Google Scholar] [CrossRef]
- You, L.; Ros, G.H.; Chen, Y.; Shao, Q.; Young, M.D.; Zhang, F.; de Vries, W. Global mean nitrogen recovery efficiency in croplands can be enhanced by optimal nutrient, crop and soil management practices. Nat. Commun. 2023, 14, 5747. [Google Scholar] [CrossRef]
- Krouk, G.; Lacombe, B.; Bielach, A.; Perrine-Walker, F.; Malinska, K.; Mounier, E.; Hoyerova, K.; Tillard, P.; Leon, S.; Ljung, K.; et al. Nitrate-Regulated Auxin Transport by NRT1.1 Defines a Mechanism for Nutrient Sensing in Plants. Dev. Cell 2010, 18, 927–937. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Y.; Hsu, P.-K.; Tsay, Y.-F. Uptake, allocation and signaling of nitrate. Trends Plant Sci. 2012, 17, 458–467. [Google Scholar] [CrossRef]
- Zhao, L.; Chen, P.; Liu, P.; Song, Y.; Zhang, D. Genetic Effects and Expression Patterns of the Nitrate Transporter (NRT) Gene Family in Populus tomentosa. Front. Plant Sci. 2021, 12, 661635. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Li, M.; Xu, W.; Liu, J.-H.; Li, C. Genome-Wide Identification of NRT Gene Family and Expression Analysis of Nitrate Transporters in Response to Salt Stress in Poncirus trifoliata. Genes 2022, 13, 1115. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yan, M.; Hu, B.; Priyadarshani, S.V.G.N.; Hou, Z.; Ojolo, S.P.; Xiong, J.; Zhao, H.; Qin, Y. Characterization and the Expression Analysis of Nitrate Transporter (NRT) Gene Family in Pineapple. Trop. Plant Biol. 2018, 11, 177–191. [Google Scholar] [CrossRef]
- Tahir, M.M.; Wang, H.; Ahmad, B.; Liu, Y.; Fan, S.; Li, K.; Lei, C.; Shah, K.; Li, S.; Zhang, D. Identification and characterization of NRT gene family reveals their critical response to nitrate regulation during adventitious root formation and development in apple rootstock. Sci. Hortic. 2021, 275, 109642. [Google Scholar] [CrossRef]
- Li, W.; Wang, Y.; Okamoto, M.; Crawford, N.M.; Siddiqi, M.Y.; Glass, A.D.M. Dissection of the AtNRT2.1:AtNRT2.2 Inducible High-Affinity Nitrate Transporter Gene Cluster. Plant Physiol. 2007, 143, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Kotur, Z.; Mackenzie, N.; Ramesh, S.; Tyerman, S.D.; Kaiser, B.N.; Glass, A.D.M. Nitrate transport capacity of the Arabidopsis thaliana NRT2 family members and their interactions with AtNAR2.1. New Phytol. 2012, 194, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Walch-Liu, P.; Forde, B.G. Nitrate signalling mediated by the NRT1.1 nitrate transporter antagonises l-glutamate-induced changes in root architecture. Plant J. 2008, 54, 820–828. [Google Scholar] [CrossRef]
- Tal, I.; Zhang, Y.; Jørgensen, M.E.; Pisanty, O.; Barbosa, I.C.R.; Zourelidou, M.; Regnault, T.; Crocoll, C.; Erik Olsen, C.; Weinstain, R.; et al. The Arabidopsis NPF3 protein is a GA transporter. Nat. Commun. 2016, 7, 11486. [Google Scholar] [CrossRef]
- Akbudak, M.A.; Filiz, E.; Çetin, D. Genome-wide identification and characterization of high-affinity nitrate transporter 2 (NRT2) gene family in tomato (Solanum lycopersicum) and their transcriptional responses to drought and salinity stresses. J. Plant Physiol. 2022, 272, 153684. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; He, M.; Zhang, W.; Han, Y.; Zhang, X.; Zhang, X.; Zhu, Y.; Wang, Y.; Liu, L.; Xu, L. Genome-wide identification and expression analysis of RsNRT gene family reveals their potential roles in response to low-nitrogen condition in radish (Raphanus sativus L.). Sci. Hortic. 2023, 321, 112273. [Google Scholar] [CrossRef]
- Guo, F.-Q.; Wang, R.; Chen, M.; Crawford, N.M. The Arabidopsis Dual-Affinity Nitrate Transporter Gene AtNRT1.1 (CHL1) Is Activated and Functions in Nascent Organ Development during Vegetative and Reproductive Growth. Plant Cell 2001, 13, 1761–1777. [Google Scholar] [CrossRef]
- Guo, F.Q.; Wang, R.; Crawford, N.M. The Arabidopsis dual-affinity nitrate transporter gene AtNRT1.1 (CHL1) is regulated by auxin in both shoots and roots. J. Exp. Bot. 2002, 53, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Segonzac, C.C.; Boyer, J.-C.; Ipotesi, E.; Szponarski, W.; Tillard, P.; Touraine, B.; Sommerer, N.; Rossignol, M.; Gibrat, R.M. Nitrate Efflux at the Root Plasma Membrane: Identification of an Arabidopsis Excretion Transporter. Plant Cell 2007, 19, 3760–3777. [Google Scholar] [CrossRef]
- von Wittgenstein, N.J.J.B.; Le, C.H.; Hawkins, B.J.; Ehlting, J. Evolutionary classification of ammonium, nitrate, and peptide transporters in land plants. BMC Evol. Biol. 2014, 14, 11. [Google Scholar] [CrossRef] [PubMed]
- Kant, S. Understanding nitrate uptake, signaling and remobilisation for improving plant nitrogen use efficiency. Semin. Cell Dev. Biol. 2018, 74, 89–96. [Google Scholar] [CrossRef]
- Xia, X.; Fan, X.; Wei, J.; Feng, H.; Qu, H.; Xie, D.; Miller, A.J.; Xu, G. Rice nitrate transporter OsNPF2.4 functions in low-affinity acquisition and long-distance transport. J. Exp. Bot. 2015, 66, 317–331. [Google Scholar] [CrossRef]
- Wang, Y.-Y.; Tsay, Y.-F. Arabidopsis Nitrate Transporter NRT1.9 Is Important in Phloem Nitrate Transport. Plant Cell 2011, 23, 1945–1957. [Google Scholar] [CrossRef]
- Kiba, T.; Feria-Bourrellier, A.-B.; Lafouge, F.; Lezhneva, L.; Boutet-Mercey, S.; Orsel, M.; Bréhaut, V.; Miller, A.; Daniel-Vedele, F.; Sakakibara, H.; et al. The Arabidopsis Nitrate Transporter NRT2.4 Plays a Double Role in Roots and Shoots of Nitrogen-Starved Plants. Plant Cell 2012, 24, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Bi, L.; Song, G.; Wang, Q.; Jin, G. Species–habitat associations in an old-growth temperate forest in northeastern China. BMC Ecol. 2018, 18, 20. [Google Scholar] [CrossRef]
- Li, X.; Liu, X.-T.; Wei, J.-T.; Li, Y.; Tigabu, M.; Zhao, X.-Y. Genetic Improvement of Pinus koraiensis in China: Current Situation and Future Prospects. Forests 2020, 11, 148. [Google Scholar] [CrossRef]
- Zhang, P.; Lü, X.-T.; Jin, G.; Liu, Z.; Li, M.-H. Leaf nitrogen resorption is more important than litter nitrogen mineralization in mediating the diversity–productivity relationship along a nitrogen-limited temperate forest succession chronosequence. For. Ecosyst. 2023, 10, 100102. [Google Scholar] [CrossRef]
- Cao, J.; Liu, H.; Zhao, B.; Li, Z.; Liang, B.; Shi, L.; Song, Z.; Wu, L.; Wang, Q.; Cressey, E.L.; et al. Nitrogen addition enhances tree radial growth but weakens its recovery from drought impact in a temperate forest in northern China. Sci. Total Environ. 2023, 903, 166884. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Zhu, J.; Lu, D.; Zhu, C.; Gao, P.; Yang, X. Effects of Korean Pine Basal Area in Mixed Broadleaved–Korean Pine Forest Stands on Its Natural Regeneration in Northeast China. For. Sci. 2021, 67, 179–191. [Google Scholar] [CrossRef]
- Xiang, G.; Wu, R.; Zhang, M.; Li, K.; He, X.; Song, Y.; Song, F. Simulated nitrogen deposition alters the ectomycorrhizal fungal community structure in northern Korean pine plantations. Catena 2023, 233, 107525. [Google Scholar] [CrossRef]
- Yang, G.; Zhang, M.; Jin, G. Effects of nitrogen addition on species composition and diversity of early spring herbs in a Korean pine plantation. Ecol. Evol. 2023, 13, e10498. [Google Scholar] [CrossRef]
- Wang, X.; Cai, X.; Xu, C.; Wang, Q. Identification and characterization of the NPF, NRT2 and NRT3 in spinach. Plant Physiol. Biochem. 2021, 158, 297–307. [Google Scholar] [CrossRef]
- Zhang, J.; Han, Z.; Lu, Y.; Zhao, Y.; Wang, Y.; Zhang, J.; Ma, H.; Han, Y.Z. Genome-wide identification, structural and gene expression analysis of the nitrate transporters (NRTs) family in potato (Solanum tuberosum L.). PLoS ONE 2021, 16, e0257383. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Yang, D.; Hu, Y.; Xu, J.; Lu, Z. Genome-Wide Identification and Expression Analysis of Nitrate Transporter (NRT) Gene Family in Eucalyptus grandis. Genes 2024, 15, 930. [Google Scholar] [CrossRef]
- Feng, J.; Zhu, C.; Cao, J.; Liu, C.; Zhang, J.; Cao, F.; Zhou, X. Genome-wide identification and expression analysis of the NRT genes in Ginkgo biloba under nitrate treatment reveal the potential roles during calluses browning. BMC Genom. 2023, 24, 633. [Google Scholar] [CrossRef]
- Chiba, Y.; Shimizu, T.; Miyakawa, S.; Kanno, Y.; Koshiba, T.; Kamiya, Y.; Seo, M. Identification of Arabidopsis thaliana NRT1/PTR FAMILY (NPF) proteins capable of transporting plant hormones. J. Plant Res. 2015, 128, 679–686. [Google Scholar] [CrossRef]
- Bai, H.; Euring, D.; Volmer, K.; Janz, D.; Polle, A. The Nitrate Transporter (NRT) Gene Family in Poplar. PLoS ONE 2013, 8, e72126. [Google Scholar] [CrossRef]
- Drechsler, N.; Courty, P.-E.; Brulé, D.; Kunze, R. Identification of arbuscular mycorrhiza-inducible Nitrate Transporter 1/Peptide Transporter Family (NPF) genes in rice. Mycorrhiza 2018, 28, 93–100. [Google Scholar] [CrossRef] [PubMed]
- You, H.; Liu, Y.; Minh, T.N.; Lu, H.; Zhang, P.; Li, W.; Xiao, J.; Ding, X.; Li, Q. Genome-wide identification and expression analyses of nitrate transporter family genes in wild soybean (Glycine soja). J. Appl. Genet. 2020, 61, 489–501. [Google Scholar] [CrossRef]
- Fan, X.; Tang, Z.; Tan, Y.; Zhang, Y.; Luo, B.; Yang, M.; Lian, X.; Shen, Q.; Miller, A.J.; Xu, G. Overexpression of a pH-sensitive nitrate transporter in rice increases crop yields. Proc. Natl. Acad. Sci. USA 2016, 113, 7118–7123. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Qiao, D.; Zhang, Q.; Li, Y.; Xu, H.; Wei, L.; Gu, Y.; Cao, Y. Cloning and expression study of a putative high-affinity nitrate transporter gene from Dunaliella salina. J. Appl. Phycol. 2004, 16, 395–400. [Google Scholar] [CrossRef]
- Yin, X.-M.; Luo, W.; Wang, S.-W.; Shen, Q.-R.; Long, X.-H. Effect of Nitrogen Starvation on the Responses of Two Rice Cultivars to Nitrate Uptake and Utilization. Pedosphere 2014, 24, 690–698. [Google Scholar] [CrossRef]
- Gao, Y.; Qi, S.; Wang, Y. Nitrate signaling and use efficiency in crops. Plant Commun. 2022, 3, 100353. [Google Scholar] [CrossRef] [PubMed]
- Lima, J.E.; Serezino, L.H.D.; Alves, M.K.; Tagliaferro, A.L.; Vitti, M.; Creste, S.; Riaño-Pachón, D.M.; dos Santos, R.V.; Figueira, A. Root nitrate uptake in sugarcane (Saccharum spp.) is modulated by transcriptional and presumably posttranscriptional regulation of the NRT2.1/NRT3.1 transport system. Mol. Genet. Genom. 2022, 297, 1403–1421. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Wu, J.; Yang, S.; Schiefelbein, J.; Gan, Y. Nitrate regulation of lateral root and root hair development in plants. J. Exp. Bot. 2020, 71, 4405–4414. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Jennings, A.; Barlow, P.W.; Forde, B.G. Dual pathways for regulation of root branching by nitrate. Proc. Natl. Acad. Sci. USA 1999, 96, 6529–6534. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Yordanov, Y.S.; Georgieva, T.; Li, X.; Busov, V. Nitrogen deprivation promotes Populus root growth through global transcriptome reprogramming and activation of hierarchical genetic networks. New Phytol. 2013, 200, 483–497. [Google Scholar] [CrossRef]
- Lv, X.; Zhang, Y.; Hu, L.; Zhang, Y.; Zhang, B.; Xia, H.; Du, W.; Fan, S.; Kong, L. Low-Nitrogen Stress Stimulates Lateral Root Initiation and Nitrogen Assimilation in Wheat: Roles of Phytohormone Signaling. J. Plant Growth Regul. 2021, 40, 436–450. [Google Scholar] [CrossRef]
- Ötvös, K.; Marconi, M.; Vega, A.; O’Brien, J.; Johnson, A.; Abualia, R.; Antonielli, L.; Montesinos, J.C.; Zhang, Y.; Tan, S.; et al. Modulation of plant root growth by nitrogen source-defined regulation of polar auxin transport. EMBO J. 2021, 40, e106862. [Google Scholar] [CrossRef]
- Huang, N.-C.; Liu, K.-H.; Lo, H.-J.; Tsay, Y.-F. Cloning and Functional Characterization of an Arabidopsis Nitrate Transporter Gene That Encodes a Constitutive Component of Low-Affinity Uptake. Plant Cell 1999, 11, 1381–1392. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-H.; Kuo, H.-F.; Canivenc, G.v.; Lin, C.-S.; Lepetit, M.; Hsu, P.-K.; Tillard, P.; Lin, H.-L.; Wang, Y.-Y.; Tsai, C.-B.; et al. Mutation of the Arabidopsis NRT1.5 Nitrate Transporter Causes Defective Root-to-Shoot Nitrate Transport. Plant Cell 2008, 20, 2514–2528. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Song, Y.; Zhu, J.; Yan, Q.; Wang, G. Korean pine seed: Linking changes in dormancy to germination in the 2 years following dispersal. For. Int. J. For. Res. 2018, 91, 98–109. [Google Scholar] [CrossRef]
- Ren, H.; Gao, G.; Ma, Y.; Li, Z.; Wang, S.; Gu, J. Shift of root nitrogen-acquisition strategy with tree age is mediated by root functional traits along the collaboration gradient of the root economics space. Tree Physiol. 2023, 43, 1341–1353. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | No. of AA | MW (kDa) | pI | GRAVY | No. of TMs |
|---|---|---|---|---|---|---|
| PkNPF1.1 | Pkor04G00574 | 633 | 69.73 | 8.69 | 0.142 | 10 |
| PkNPF1.2 | Pkor04G00573 | 630 | 69.28 | 8.92 | 0.141 | 9 |
| PkNPF1.3 | Pkor04G00568 | 635 | 70.17 | 9.34 | 0.17 | 10 |
| PkNPF1.4 | Pkor04G04403 | 639 | 70.13 | 9.25 | 0.148 | 10 |
| PkNPF1.5 | Pkor04G00571 | 639 | 70.45 | 9.35 | 0.096 | 8 |
| PkNPF1.6 | Pkor04G00570 | 639 | 70.33 | 9.21 | 0.088 | 8 |
| PkNPF1.7 | Pkor04G00569 | 639 | 70.33 | 9.21 | 0.088 | 8 |
| PkNPF2.1 | Pkor10G02120 | 661 | 73.07 | 9.21 | 0.201 | 10 |
| PkNPF2.2 | Pkor10G02119 | 638 | 70.83 | 8.53 | 0.125 | 8 |
| PkNPF2.3 | Pkor10G02121 | 658 | 72.67 | 7.12 | 0.159 | 10 |
| PkNPF3.1 | Pkor12G01674 | 648 | 71.75 | 9.17 | 0.154 | 11 |
| PkNPF4.1 | Pkor01G02841 | 564 | 63.08 | 8.82 | 0.36 | 11 |
| PkNPF4.2 | Pkor01G03784 | 574 | 64.51 | 8.29 | 0.35 | 12 |
| PkNPF4.3 | Pkor11G00219 | 580 | 64.53 | 8.46 | 0.3 | 11 |
| PkNPF4.4 | Pkor08G03209 | 623 | 69.08 | 7.61 | 0.234 | 12 |
| PkNPF4.5 | Pkor08G02109 | 607 | 66.91 | 9.02 | 0.171 | 12 |
| PkNPF4.6 | Pkor08G03210 | 606 | 67.09 | 9.01 | 0.144 | 11 |
| PkNPF5.1 | Pkor10G00974 | 618 | 69.15 | 9.1 | 0.255 | 10 |
| PkNPF5.2 | Pkor08G01587 | 608 | 67.49 | 9.27 | 0.179 | 9 |
| PkNPF5.3 | Pkor08G01589 | 598 | 66.37 | 9.14 | 0.228 | 9 |
| PkNPF5.4 | Pkor02G03103 | 609 | 68.36 | 9.15 | 0.214 | 11 |
| PkNPF5.5 | Pkor02G03104 | 537 | 59.25 | 9.47 | 0.277 | 10 |
| PkNPF5.6 | Pkor02G01331 | 629 | 70.39 | 9.27 | 0.163 | 12 |
| PkNPF5.7 | Pkor02G01336 | 623 | 69.98 | 9.09 | 0.173 | 10 |
| PkNPF5.8 | Pkor02G03105 | 623 | 69.81 | 9.33 | 0.232 | 10 |
| PkNPF5.9 | Pkor02G01335 | 622 | 69.88 | 9.35 | 0.2 | 10 |
| PkNPF5.10 | Pkor05G02326 | 614 | 68.27 | 9.05 | 0.269 | 10 |
| PkNPF5.11 | Pkor06G02154 | 614 | 68.49 | 9.02 | 0.264 | 9 |
| PkNPF5.12 | Pkor06G00882 | 605 | 67.07 | 9.14 | 0.301 | 12 |
| PkNPF5.13 | Pkor06G00883 | 607 | 67.27 | 8.54 | 0.382 | 12 |
| PkNPF5.14 | Pkor09G01222 | 612 | 68.18 | 8.95 | 0.29 | 11 |
| PkNPF5.15 | Pkor06G00884 | 606 | 67.37 | 8.89 | 0.323 | 12 |
| PkNPF5.16 | Pkor06G03064 | 611 | 68.28 | 7.54 | 0.296 | 12 |
| PkNPF5.17 | Pkor06G00881 | 607 | 68.42 | 9.05 | 0.254 | 12 |
| PkNPF5.18 | Pkor06G03063 | 608 | 67.87 | 8.88 | 0.305 | 12 |
| PkNPF5.19 | Pkor06G03062 | 608 | 67.78 | 8.59 | 0.318 | 12 |
| PkNPF5.20 | Pkor12G00815 | 559 | 61.88 | 5.83 | 0.393 | 10 |
| PkNPF5.21 | Pkor12G01382 | 605 | 67.84 | 5.92 | 0.248 | 12 |
| PkNPF5.22 | Pkor07G00455 | 578 | 64.67 | 5.86 | 0.274 | 9 |
| PkNPF5.23 | Pkor07G00456 | 579 | 64.83 | 6.32 | 0.364 | 10 |
| PkNPF5.24 | Pkor07G02566 | 547 | 61.45 | 5.71 | 0.277 | 8 |
| PkNPF5.25 | Pkor07G02567 | 606 | 68.19 | 8.79 | 0.227 | 9 |
| PkNPF5.26 | Pkor07G00457 | 590 | 66.15 | 8.82 | 0.253 | 9 |
| PkNPF5.27 | Pkor07G00460 | 592 | 66.1 | 8.12 | 0.338 | 10 |
| PkNPF5.28 | Pkor07G00458 | 595 | 66.58 | 7.53 | 0.33 | 10 |
| PkNPF5.29 | Pkor07G00459 | 595 | 66.58 | 6.86 | 0.335 | 10 |
| PkNPF5.30 | Pkor07G00461 | 578 | 64.18 | 7.91 | 0.304 | 10 |
| PkNPF5.31 | Pkor07G02568 | 576 | 63.74 | 8.44 | 0.325 | 10 |
| PkNPF5.32 | Pkor07G00462 | 576 | 63.83 | 8.27 | 0.335 | 10 |
| PkNPF6.1 | Pkor12G01587 | 610 | 66.98 | 8.43 | 0.243 | 11 |
| PkNPF6.2 | Pkor11G00350 | 601 | 66.65 | 8.96 | 0.242 | 11 |
| PkNPF6.3 | Pkor11G02733 | 597 | 66.53 | 8.28 | 0.209 | 11 |
| PkNPF7.1 | Pkor11G02681 | 598 | 65.67 | 7.96 | 0.301 | 12 |
| PkNPF7.2 | Pkor11G00148 | 626 | 69.15 | 8.52 | 0.208 | 12 |
| PkNPF7.3 | Pkor09G02450 | 630 | 70.58 | 8.2 | 0.082 | 11 |
| PkNPF7.4 | Pkor09G02643 | 622 | 69.64 | 6.07 | 0.134 | 12 |
| PkNPF8.1 | Pkor09G00733 | 589 | 65.58 | 8.97 | 0.111 | 9 |
| PkNPF8.2 | Pkor09G00732 | 571 | 63.61 | 8.22 | 0.136 | 10 |
| PkNPF8.3 | Pkor05G01620 | 596 | 66.25 | 5.48 | 0.212 | 10 |
| PkNPF8.4 | Pkor05G01614 | 596 | 66.17 | 5.25 | 0.237 | 10 |
| PkNPF8.5 | Pkor05G01606 | 594 | 65.59 | 6.28 | 0.204 | 11 |
| PkNPF8.6 | Pkor05G01605 | 594 | 65.97 | 6.15 | 0.258 | 11 |
| PkNPF8.7 | Pkor01G04179 | 566 | 63.29 | 8.23 | 0.258 | 10 |
| PkNPF8.8 | Pkor09G00734 | 616 | 68.79 | 8.53 | 0.069 | 10 |
| PkNPF8.9 | Pkor09G00735 | 589 | 66.21 | 8.86 | 0.128 | 9 |
| PkNPF8.10 | Pkor09G00737 | 589 | 66.15 | 8.74 | 0.117 | 10 |
| PkNRT2.1 | Pkor08G00516 | 533 | 58 | 8.69 | 0.3 | 10 |
| PkNRT2.2 | Pkor12G01755 | 533 | 58 | 8.69 | 0.3 | 10 |
| PkNRT2.3 | Pkor12G02554 | 533 | 58 | 8.69 | 0.3 | 10 |
| PkNRT2.4 | Pkor12G02553 | 533 | 57.87 | 8.39 | 0.297 | 10 |
| PkNRT2.5 | Pkor12G01756 | 533 | 57.95 | 8.14 | 0.333 | 10 |
| PkNRT2.6 | Pkor09G01906 | 548 | 58.63 | 8.17 | 0.353 | 10 |
| PkNRT3.1 | Pkor09G03181 | 268 | 29.26 | 9.18 | 0.018 | 1 |
| PkNRT3.2 | Pkor09G01715 | 207 | 22.58 | 9.73 | 0.173 | 1 |
| PkNRT3.3 | Pkor09G01713 | 214 | 23.98 | 9.25 | 0.012 | 1 |
| PkNRT3.4 | Pkor09G01714 | 214 | 23.49 | 9.64 | −0.037 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Wu, H.; Li, B.; Wang, P.; Zhang, P.; Shen, H.; Yang, J. Genome-Wide Identification and Expression Analysis of PkNRT Gene Family in Korean Pine (Pinus koraiensis). Plants 2025, 14, 238. https://doi.org/10.3390/plants14020238
Zhao X, Wu H, Li B, Wang P, Zhang P, Shen H, Yang J. Genome-Wide Identification and Expression Analysis of PkNRT Gene Family in Korean Pine (Pinus koraiensis). Plants. 2025; 14(2):238. https://doi.org/10.3390/plants14020238
Chicago/Turabian StyleZhao, Xinyu, Haibo Wu, Boyang Li, Pengyang Wang, Peng Zhang, Hailong Shen, and Jianfei Yang. 2025. "Genome-Wide Identification and Expression Analysis of PkNRT Gene Family in Korean Pine (Pinus koraiensis)" Plants 14, no. 2: 238. https://doi.org/10.3390/plants14020238
APA StyleZhao, X., Wu, H., Li, B., Wang, P., Zhang, P., Shen, H., & Yang, J. (2025). Genome-Wide Identification and Expression Analysis of PkNRT Gene Family in Korean Pine (Pinus koraiensis). Plants, 14(2), 238. https://doi.org/10.3390/plants14020238