
Establishment of a Virus-Induced Gene Silencing System in Abelmoschus manihot L.

 , ,
, ,
Abstract
1. Introduction
2. Results
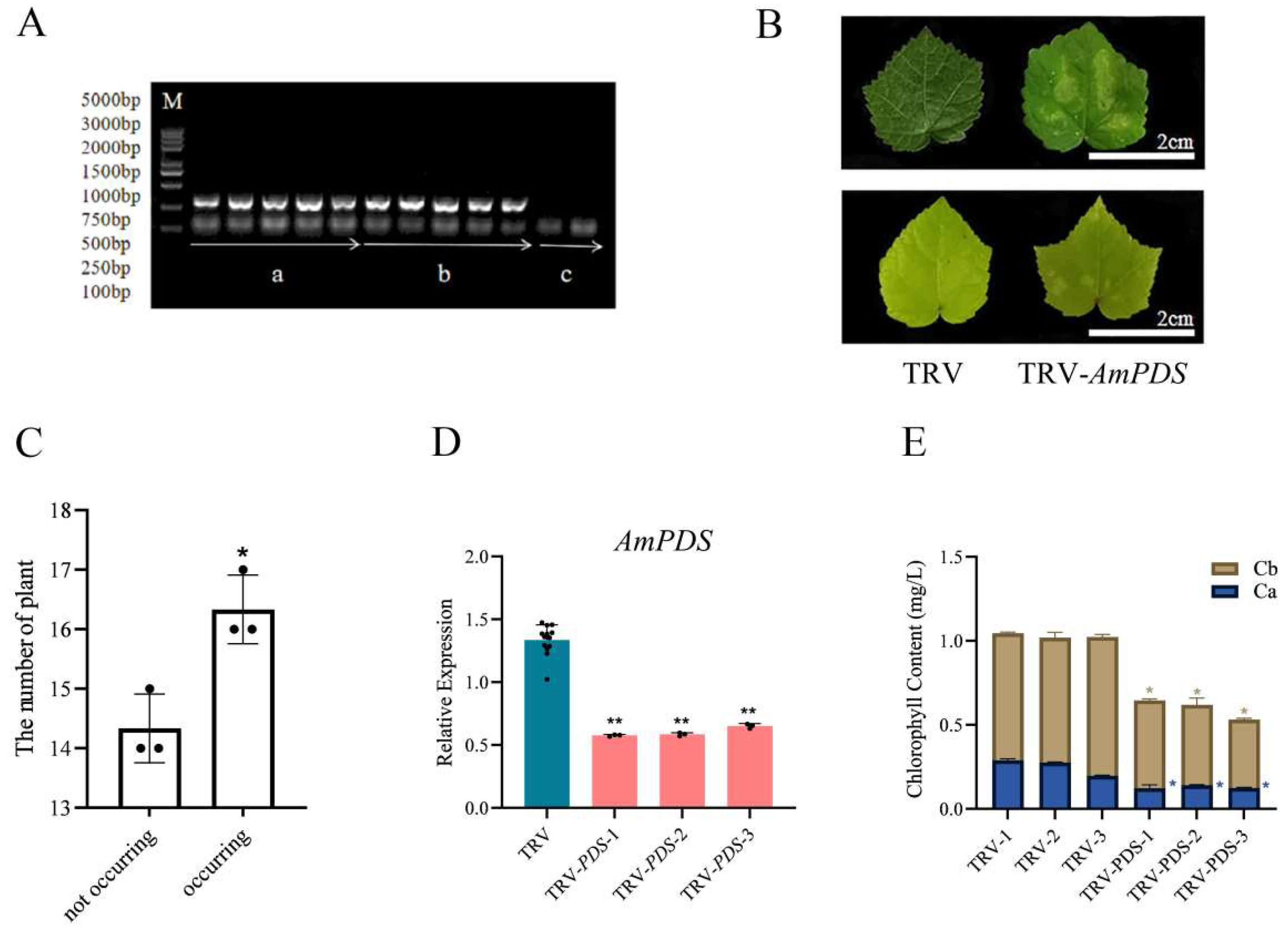
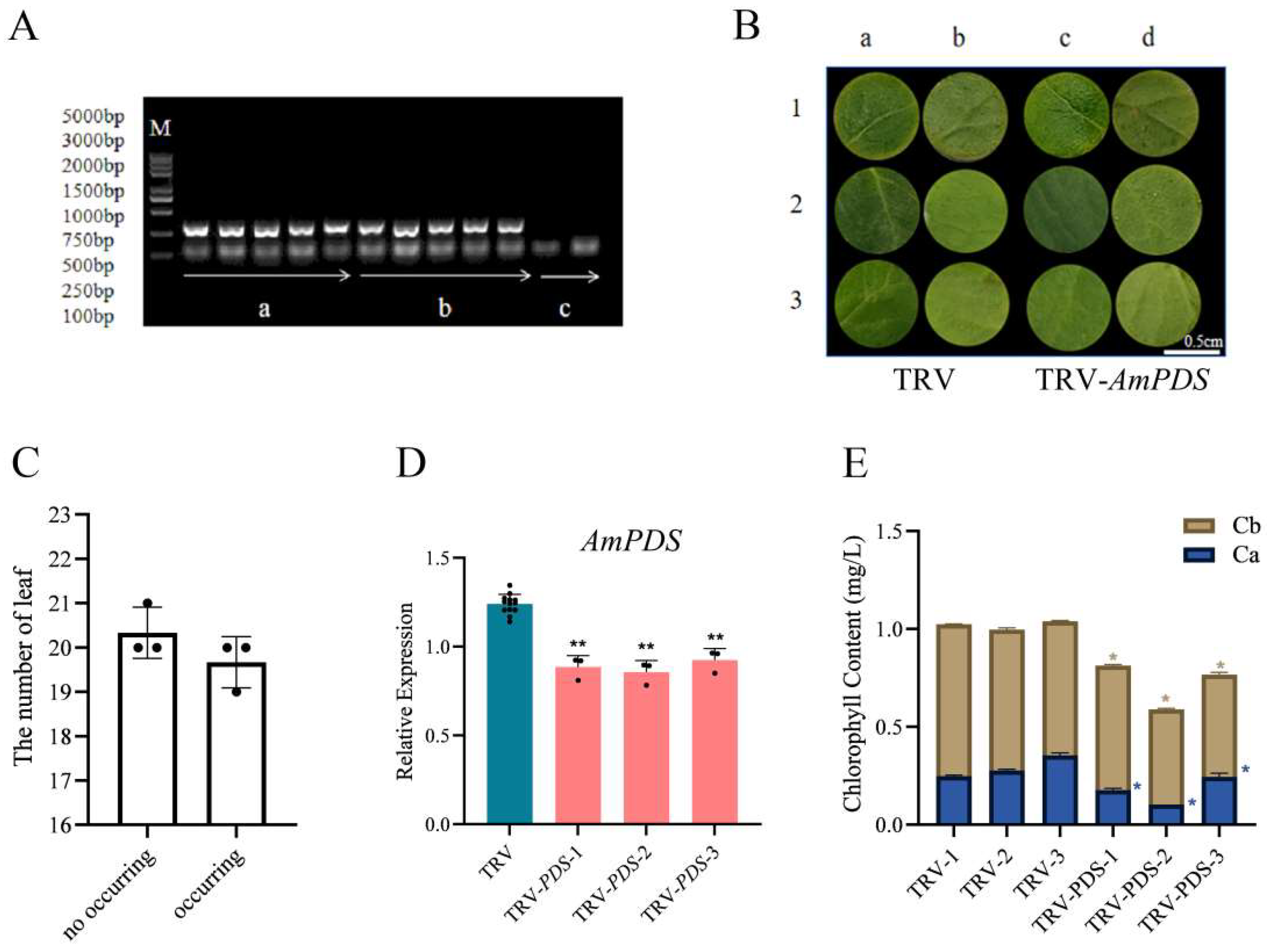
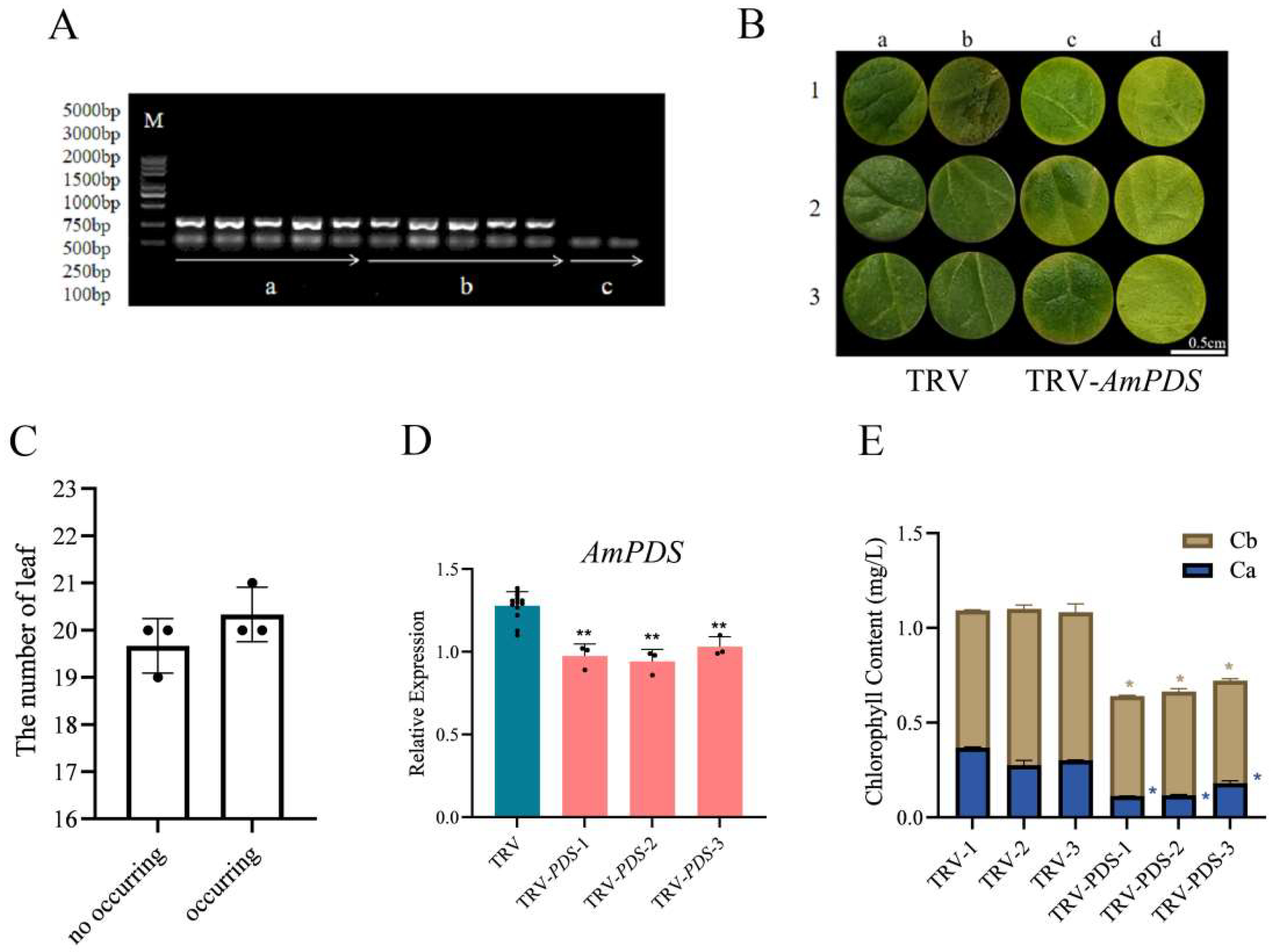
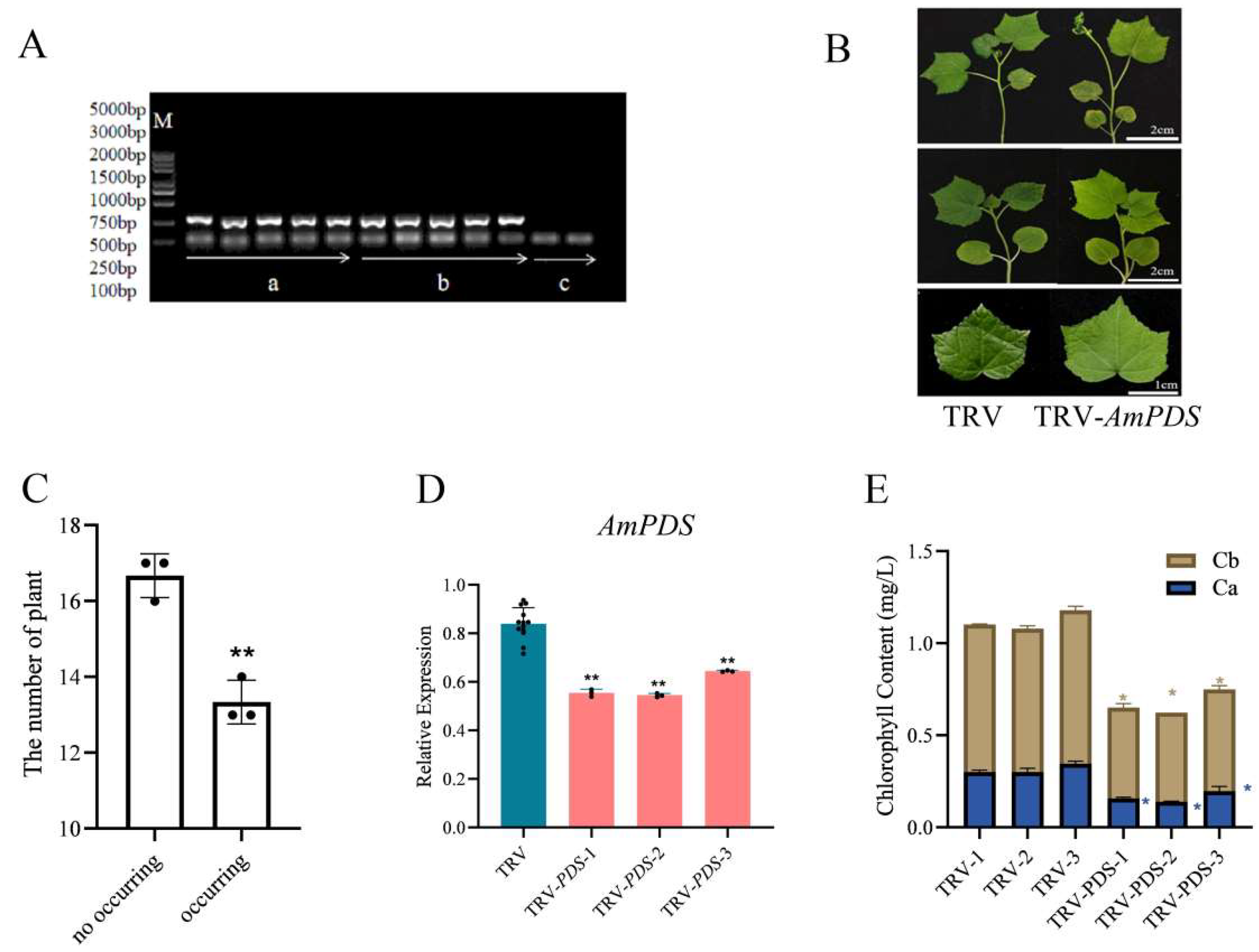
2.1. Cloning Results of the AmPDS Fragment
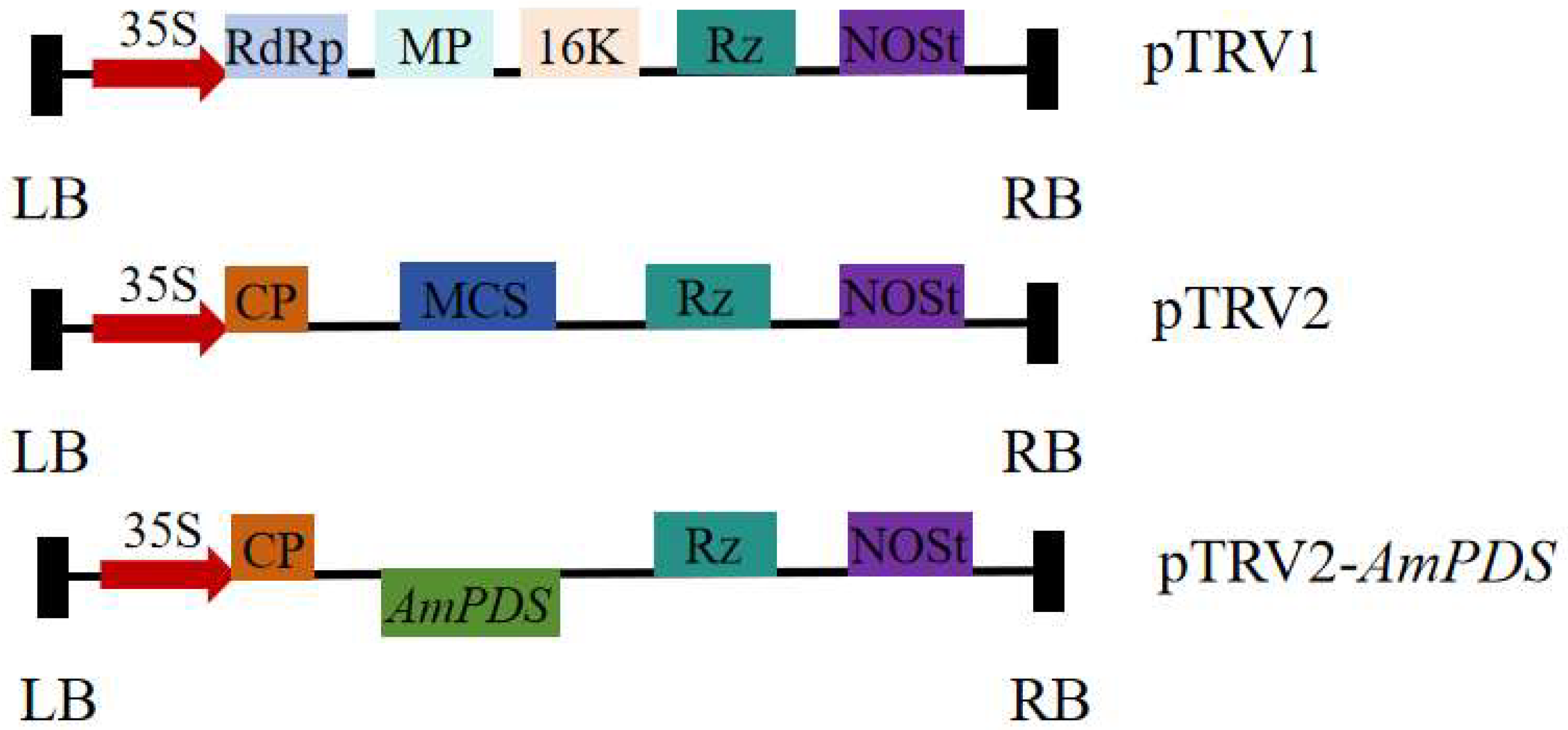
2.2. Construction of the AmPDS VIGS Recombinant Vector
2.3. Comparison of Silencing Efficiency of Different Inoculation Methods Mediated by Agrobacterium
2.3.1. Back-of-Blade Injection
2.3.2. Vacuum Infiltration Method
2.3.3. Direct Soaking Method
2.3.4. Root Drenching Method
2.4. Comparison of Different Agrobacterium Inoculation Methods
3. Discussion
4. Materials and Methods
4.1. Plant Materials, Strains, and Plasmids
4.2. Reagents
4.3. Cloning of the Core Fragment of AmPDS
4.4. Construction of the Recombinant Viral Vector pTRV–AmPDS
4.5. Preparation of the Inoculation Solution
4.6. Inoculation Methods
4.6.1. Back-of-Blade Injection Method
4.6.2. The Method of Vacuum Infiltration
4.6.3. The Method of Direct Soaking
4.6.4. The Method of Root Drenching
4.7. Gene Expression Analysis
4.8. TRV CP mRNA Detection
4.9. Determination of Chlorophyll Content
4.10. Statistical Analyses
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, M.J.; Hung, K.Y.; Li, L. Research progress on chemical constituents and biological activity of Hibiscus manihot L. J. Anhui Agric. Sci. 2019, 47, 5–7. [Google Scholar]
- Peng, Z.B.; Wu, Z.B. Analysis of hyperosidein flower of Hibiscus manihot L. by HPLC-mass spectrometry. J. Anhui Agric. Sci. 2008, 36, 10028–10029. [Google Scholar]
- Lan, R.; Li, B.; Liu, H.; Chen, L. Extraction and determination of total flavone in Abelmoschus manihot L. J. Jiangsu Agric. Sci. 2012, 40, 280–282. [Google Scholar]
- Lu, D.; Jia, R.B. Research progress on Chinese medicinal material Aurea Helianthus. Chin. J. Drug Eval. 2015, 32, 90–92. [Google Scholar]
- Li, B.; Hu, D.Y.; Li, S.S.; Xin, X.L.; Chen, L.; Liu, Q. Determination of the unsaturated fatty acids of Hibiscus manihot seed oil by GC-MS. Food Res. Dev. 2012, 33, 121–123. [Google Scholar]
- Chen, L.M.; Dong, S.; Liang, W.M.; Yang, L.L.; Chen, Y. Researches on extraction and antioxidant activity of soluble carbohydrate from stalk of Aurea Helianthus. Food Res. Dev. 2016, 37, 59–63. [Google Scholar]
- Chen, L.; Xin, X.L.; Su, D.H.; Wang, X.J.; Hu, X.M.; Li, B.; Liu, Q. Study on refining technology of total flavonoids from Hibiscus manihot L. J. Jiangsu Agric. Sci. 2016, 44, 339–343. [Google Scholar]
- Zhou, Y.H.; Xu, X.D.; Shi, Q.Y.; Gao, J.; Cao, L. Optimization of extraction of total flavonoids from Abelmoschus manihot L. and study on its components. Agric. Sci. J. Yanbian Univ. 2023, 21, 2024–2031. [Google Scholar]
- Peng, Z.B.; Wu, Z.B.; Yu, H.J. The GC-MS analysis of fatty acid composition in Hibiscus manihot seed oil. J. Anhui Agric. Sci. 2009, 37, 16753–16756. [Google Scholar]
- Wei, Q.; Lan, R.; Xin, X.L.; Chen, L. Determination of total flavonoids content in Golden Kwai seed by ultraviolet spectrophotometry. J. Anhui Agric. Sci. 2012, 40, 7050–7060. [Google Scholar]
- Song, L.L.; Huang, B.B.; Zhang, C.H. Ultrasonic extraction technology of total flavone from flower of Hibiscus manihot. Guizhou Agric. Sci. 2016, 44, 70–72. [Google Scholar]
- Wang, M.Z. The Extraction and Separation of Flavone in Hibiscus manihot Flower and the Investigation of Its Activities. Master’s Thesis, Hebei University of Science and Technology, Shijiazhuang, China, 2017. [Google Scholar]
- Rössner, C.; Lotz, D.; Becker, A. VIGS Goes Viral: How VIGS transforms our understanding of plant science. Annu. Rev. Plant Biol. 2022, 73, 703–728. [Google Scholar] [CrossRef] [PubMed]
- Schachtsiek, J.; Hussain, T.; Azzouhri, K.; Kayser, O.; Stehle, F. Virus-induced gene silencing (VIGS) in Cannabis sativa L. Plant Methods 2019, 15, 157. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.-H.; Slotkin, R.K. The initiation of RNA interference (RNAi) in plants. Curr. Opin. Plant Biol. 2021, 61, 102014. [Google Scholar] [CrossRef]
- Benedito, V.A.; Visser, P.B.; Angenent, G.C.; Krens, F.A. The potential of virus-induced gene silencing for speeding up functional characterization of plant genes. Genet. Mol. Res. 2004, 3, 323–341. [Google Scholar]
- Burch-Smith, T.M.; Anderson, J.C.; Martin, G.B.; Dinesh-Kumar, S.P. Applications and advantages of virus-induced gene silencing for gene function studies in plants. Plant J. 2004, 39, 734–746. [Google Scholar] [CrossRef]
- Chen, X.; Rechavi, O. Plant and animal small RNA communications between cells and organisms. Nat. Rev. Mol. Cell Biol. 2022, 23, 185–203. [Google Scholar] [CrossRef]
- Chen, Y.G.; Hur, S. Cellular origins of dsRNA, their recognition and consequences. Nat. Rev. Mol. Cell Biol. 2022, 23, 286–301. [Google Scholar] [CrossRef]
- Liu, Y.; Schiff, M.; Dinesh-Kumar, S.P. Virus-induced gene silencing in tomato. Plant J. 2002, 31, 777–786. [Google Scholar] [CrossRef]
- Constantin, G.D.; Krath, B.N.; MacFarlane, S.A.; Nicolaisen, M.; Johansen, I.E.; Lund, O.S. Virus-induced gene silencing as a tool for functional genomics in a legume species. Plant J. 2004, 40, 622–631. [Google Scholar] [CrossRef]
- Chung, E.; Seong, E.; Kim, Y.-C.; Chung, E.J.; Oh, S.-K.; Lee, S.; Park, J.M.; Joung, Y.H.; Choi, D. A method of high frequency virus-induced gene silencing in chili pepper (Capsicum annuum L. cv. Bukang). Mol. Cells 2004, 17, 377–380. [Google Scholar] [CrossRef] [PubMed]
- Holzberg, S.; Brosio, P.; Gross, C.; Pogue, G.P. Barley stripe mosaic virus-induced gene silencing in a monocot plant. Plant J. 2002, 30, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Peele, C.; Jordan, C.V.; Muangsan, N.; Turnage, M.; Egelkrout, E.; Eagle, P.; Hanley-Bowdoin, L.; Robertson, D. Silencing of a meristematic gene using geminivirus-derived vectors. Plant J. 2001, 27, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Ratcliff, F.G.; MacFarlane, S.A.; Baulcombe, D.C. Gene silencing without DNA: RNA-mediated cross-protection between viruses. Plant Cell 1999, 11, 1207. [Google Scholar] [CrossRef]
- Liu, Y.; Schiff, M.; Marathe, R.; Dinesh-Kumar, S.P. Tobacco Rar1, EDS1 and NPR1/NIM1 like genes are required for N-mediated resistance to tobacco mosaic virus. Plant J. 2002, 30, 415–429. [Google Scholar] [CrossRef]
- Zhao, J.; Jiang, H.; Wang, G.; Wang, Z.; Dong, J.; Song, J. Virus-induced gene silencing in diploid and tetraploid potato species. Methods Mol. Biol. 2020, 2172, 39–50. [Google Scholar]
- Ekengren, S.K.; Liu, Y.; Schiff, M.; Dinesh-Kumar, S.P.; Martin, G.B. Two MAPK cascades, NPR1, and TGA transcription factors play a role in Pto-mediated disease resistance in tomato. Plant J. 2003, 36, 905–917. [Google Scholar] [CrossRef]
- Choi, H.W.; Kim, Y.J.; Lee, S.C.; Hong, J.K.; Hwang, B.K. Hydrogen peroxide generation by the pepper extracellular peroxidase CaPO2 activates local and systemic cell death and defense response to bacterial pathogens. Plant Physiol. 2007, 145, 890–904. [Google Scholar] [CrossRef]
- Gao, X.; Britt, R.C., Jr.; Shan, L.; He, P. Agrobacterium-mediated virus-induced gene silencing assay in cotton. J. Vis. Exp. 2011, 54, 2938. [Google Scholar] [CrossRef]
- Ryu, C.; Anand, A.; Kang, L.; Mysore, K.S. Agrodrench: A novel and effective agroinoculation method for virus-induced gene silencing in roots and diverse Solanaceous species. Plant J. 2004, 40, 322–331. [Google Scholar] [CrossRef]
- Hileman, L.C.; Drea, S.; de Martino, G.; Litt, A.; Irish, V.F. Virus-induced gene silencing is an effective tool for assaying gene function in the basal eudicot species Papaver somniferum (opium poppy). Plant J. 2005, 44, 334–341. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, D.; Zhang, Y.; Liu, K.; Xu, K.; Zhang, F.; Wang, J.; Tan, G.; Nie, X.; Ji, Q.; et al. Vacuum and co-cultivation agroinfiltration of (germinated) seeds results in tobacco rattle Virus (TRV) mediated whole-plant virus-induced gene silencing (VIGS) in wheat and maize. Front. Plant Sci. 2017, 8, 393. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liang, M.; Chen, J.; Wang, H.; Ma, L. Rapid generation of fragrant thermo-sensitive genic male sterile rice with enhanced disease resistance via CRISPR/Cas9. Planta 2024, 259, 112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Gai, M.; Xue, B.; Jia, N.; Wang, C.; Wang, J.; Sun, H. The use of miRNAs as reference genes for miRNA expression normalization during Lilium somatic embryogenesis by real-time reverse transcription PCR analysis. Plant Cell Tissue Organ Cult. (PCTOC) 2017, 129, 105–118. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′ to 3′) | |
|---|---|---|
| AmPDS | Forward | ATGCTGACTTGGCCTGAGAAAG |
| Reverse | TCAGTGTCTGCTTGCTCCAGTC | |
| AmPDS silencing fragment | Forward | gtgagtaaggttaccgaattcATGCTGACTTGGCCTGAGAAAG |
| Reverse | cgtgagctcggtaccggatccCCAAGAATGCCATCTTTGATCC | |
| Number | Gene Name | Sequence (5′ to 3′) | |
|---|---|---|---|
| 1 | AmPDS | Forward | CCTGATCGTGTGACTGAGGA |
| Reverse | ATTGGCATGCAAAGCCTCTC | ||
| 2 | Actin | Forward | TCTTTCATCGGGATGGAAGC |
| Reverse | ACTGAGCACAATGTTACCGTAGAG | ||
| Reagent | Volume |
|---|---|
| 5× M-MLV buffer | 2 μL |
| dNTP Mix (2.5 mM) | 2 μL |
| Random primer (10 μM) | 0.5 μL |
| Oligo(dT)18 (10 μM) | 0.5 μL |
| RNase inhibitor (40 U/μL) | 0.25 μL |
| M-MLV reverse transcriptase (200 U/μL) | 0.25 μL |
| RNA | 1 μL |
| RNase-free ddH2O | To a final volume of 10 µL |
| Primer | Sequence (5′ to 3′) | |
|---|---|---|
| TRV CP mRNA | Forward | CCTGCTGACTTGATGGACGA |
| Reverse | CCAGTGTTCGCCTTGGTAG | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Hou, J.; Chu, H.; Guo, P.; Sang, Q.; Liu, Z.; Cao, L. Establishment of a Virus-Induced Gene Silencing System in Abelmoschus manihot L. Plants 2025, 14, 150. https://doi.org/10.3390/plants14020150
Zhang T, Hou J, Chu H, Guo P, Sang Q, Liu Z, Cao L. Establishment of a Virus-Induced Gene Silencing System in Abelmoschus manihot L. Plants. 2025; 14(2):150. https://doi.org/10.3390/plants14020150
Chicago/Turabian StyleZhang, Ting, Jiaqi Hou, Hongtao Chu, Pengyu Guo, Qianzi Sang, Zhongxu Liu, and Li Cao. 2025. "Establishment of a Virus-Induced Gene Silencing System in Abelmoschus manihot L." Plants 14, no. 2: 150. https://doi.org/10.3390/plants14020150
APA StyleZhang, T., Hou, J., Chu, H., Guo, P., Sang, Q., Liu, Z., & Cao, L. (2025). Establishment of a Virus-Induced Gene Silencing System in Abelmoschus manihot L. Plants, 14(2), 150. https://doi.org/10.3390/plants14020150