
Pollen Morphological Characteristics of 46 Germplasm Resources of Polygonatum and Its Taxonomic Implications

 , ,
, ,
Abstract
1. Introduction
2. Results
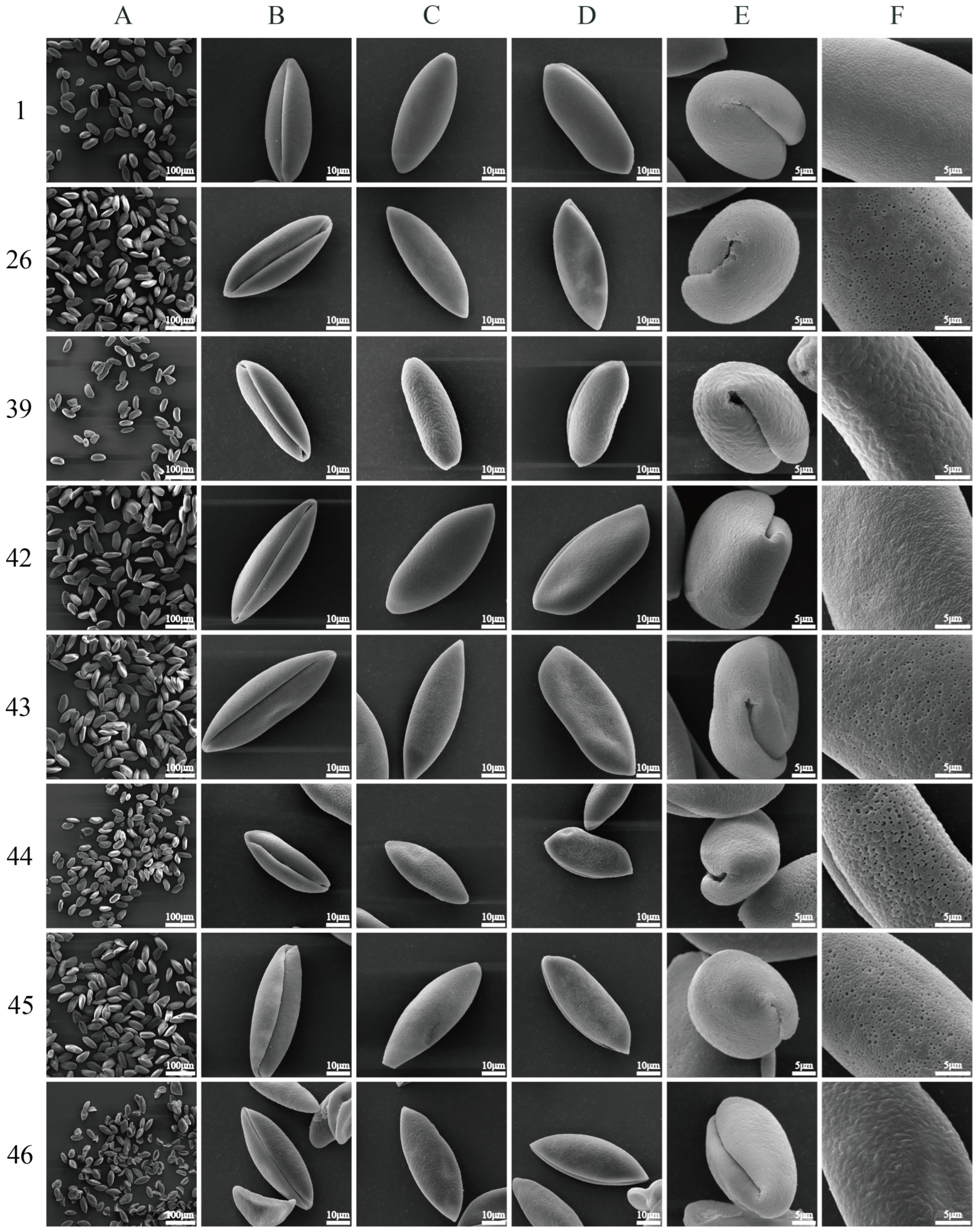
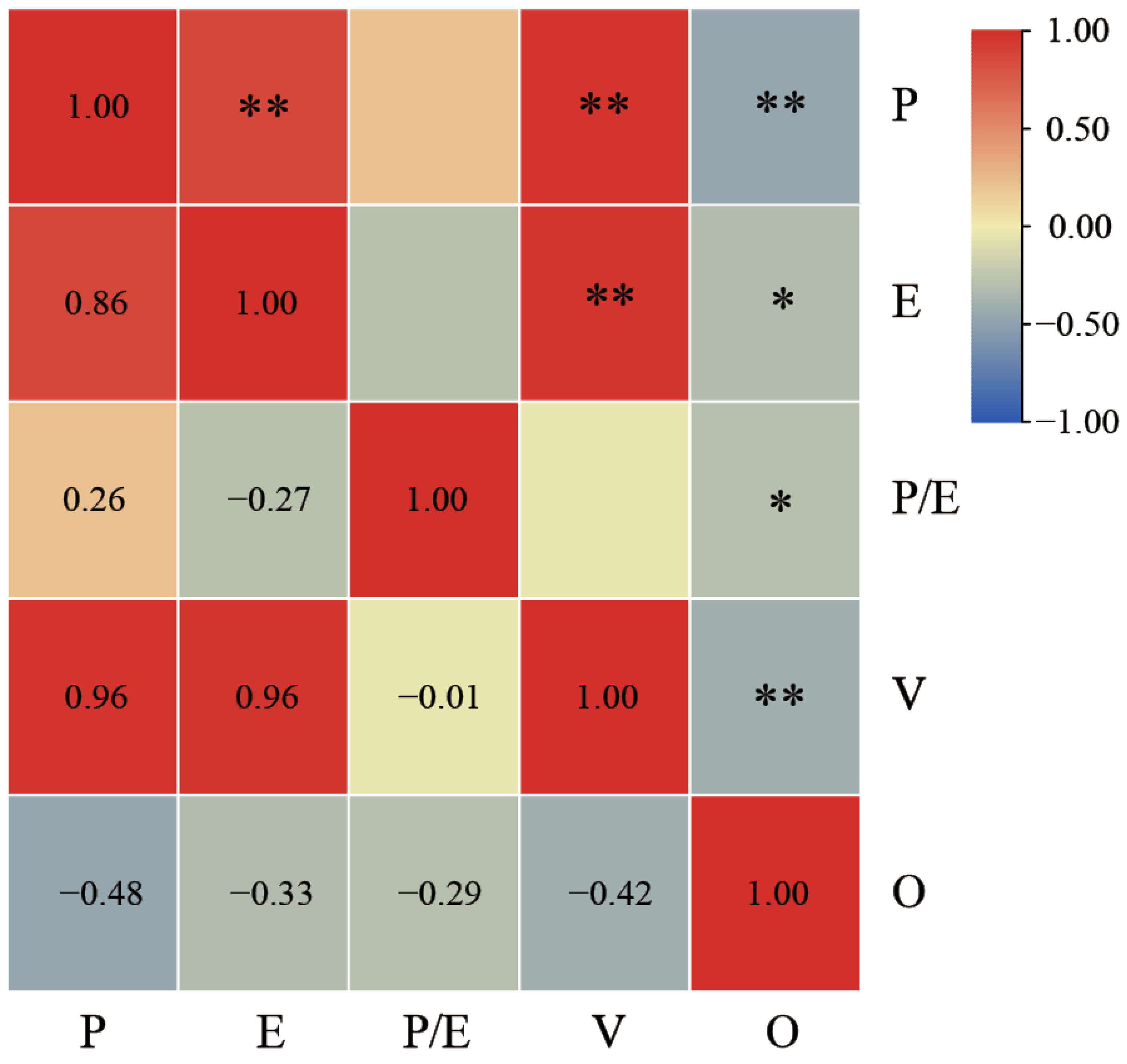
2.1. Pollen Shape and Size
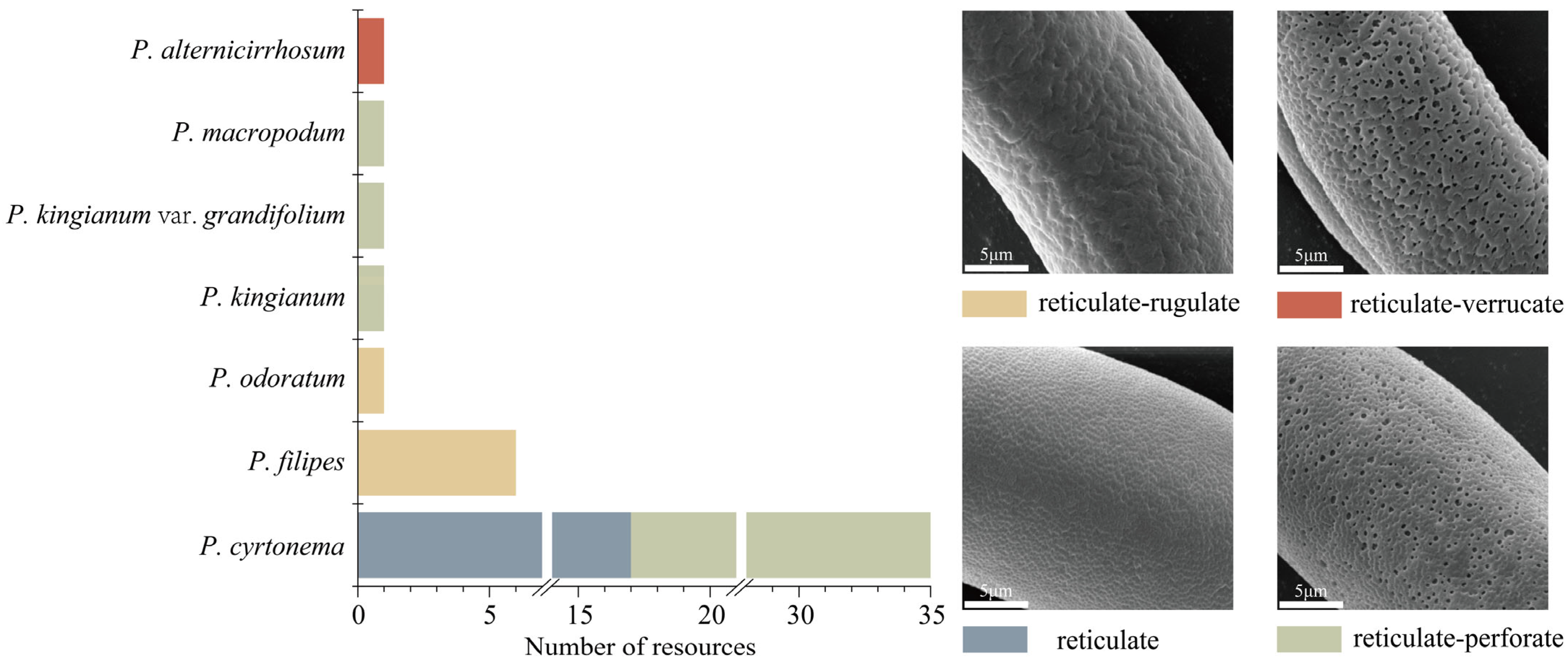
2.2. Pollen Ornamentation
2.3. Cluster Analysis of Pollen Morphology
3. Discussion
3.1. Pollen Morphological Characteristics
3.2. Evolutionary Trends in Pollen Morphological Characteristicss
3.3. Genetic Relationship Analysis Based on Pollen Morphological Characteristicss
4. Materials and Methods
4.1. Plant Material
4.2. Experimental Method
4.2.1. Pollen Collection, Preservation, and Treatment
4.2.2. Scanning Electron Microscopy Observation
4.3. Pollen Morphology Description and Data Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Editorial Committee of Flora of China; Chinese Academy of Sciences. Flora of China; Science Press: Beijing, China, 2016; Volume 15, pp. 58–80. (In Chinese) [Google Scholar]
- Xu, Q.H.; Huang, X.Z.; Wang, T. Study of the Pollen Morphology of Common Plant Species in the Mountains of Northern China—Exploration of Precise Identification Under an Optical Microscope; Science Press: Beijing, China, 2022; p. 51. (In Chinese) [Google Scholar]
- Chen, X.Q.; Tamura, M.N. Flora of China; Science Press and Missouri Botanical Garden Press: Beijing, China; St. Louis, MO, USA, 2000; Volume 24, pp. 223–235. [Google Scholar]
- Wei, Y.F.; Jiang, X.H.; Song, R.; She, C.W. Molecular cytogenetic characterization of 9 populations of four species in the genus Polygonatum (Asparagaceae). Comp. Cytogenet. 2024, 18, 73–95. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Zhao, C.C.; Li, X.; Gao, Q.Z.; Huang, L.Q.; Xiao, P.G.; Gao, W.Y. The genus Polygonatum: A review of ethnopharmacology, phytochemistry and pharmacology. J. Ethnopharmacol. 2018, 214, 274–291. [Google Scholar] [CrossRef] [PubMed]
- Wujisguleng, W.; Liu, Y.J.; Long, C.L. Ethnobotanical review of food uses of Polygonatum (Convallariaceae) in China. Acta Soc. Bot. Pol. 2012, 81, 239–244. [Google Scholar] [CrossRef]
- Tamura, M.N. Biosystematic studies on the genus Polygonatum (Liliaceae). I, Karyotype analysis of species indigenous to Japan and its adjacent regions. Cytologia 1990, 55, 443–466. [Google Scholar] [CrossRef]
- Baker, J.G. Revision of the Genera and Species of Asparagaceæ. Bot. J. Linn. Soc. 1875, 14, 508–629. [Google Scholar] [CrossRef]
- Tamura, M.N. Biosystematic studies on the genus Polygonatum (Liliaceae): III. Morphology of staminal filaments and karyology of eleven Eurasian species. Bot. Jahrb. Syst. 1993, 115, 1–26. [Google Scholar]
- Zhang, D.J.; Ren, J.; Jiang, H.; Wanga, V.O.; Dong, X.; Hu, G.W. Comparative and phylogenetic analysis of the complete chloroplast genomes of six Polygonatum species (Asparagaceae). Sci. Rep. 2023, 13, 7237. [Google Scholar] [CrossRef]
- Xia, M.Q.; Liu, Y.; Liu, J.J.; Chen, D.H.; Shi, Y.; Chen, Z.X.; Chen, D.R.; Jin, R.F.; Chen, H.L.; Comes, H.P.; et al. Out of the Himalaya-Hengduan Mountains: Phylogenomics, biogeography and diversification of Polygonatum Mill. (Asparagaceae) in the Northern Hemisphere. Mol. Phylogenet. Evol. 2022, 169, 107431. [Google Scholar] [CrossRef]
- Zhao, L.H.; Zhou, S.D.; He, X.J. A phylogenetic study of Chinese Polygonatum (Polygonateae, Asparagaceae). Nord. J. Bot. 2019, 37, e02019. [Google Scholar] [CrossRef]
- Meng, Y.; Nie, Z.L.; Deng, T.; Wen, J.; Yang, Y.P. Phylogenetics and evolution of phyllotaxy in the Solomon’s seal genus Polygonatum (Asparagaceae: Polygonateae). Bot. J. Linn. Soc. 2014, 176, 435–451. [Google Scholar] [CrossRef]
- Tsymbalyuk, Z.M.; Nitsenko, L.M.; Mosyakin, S.L. Taxonomic significance of pollen morphology of selected taxa of Bassia, Sedobassia, Spirobassia and Neokochia (Chenopodiaceae/Amaranthaceae sensu APG IV). Palynology 2023, 47, 2233586. [Google Scholar] [CrossRef]
- Li, M.; Tian, C.F.; Idrees, M.; Pathak, M.; Xiong, X.H.; Gao, X.F.; Wang, X.R. Pollen Morphology in Sorbus L. (Rosaceae) and Its Taxonomic Implications. Plants 2023, 12, 3318. [Google Scholar] [CrossRef] [PubMed]
- Halbritter, H.; Ulrich, S.; Grímsson, F.; Weber, M.; Zetter, R.; Hesse, M.; Buchner, R.; Svojtka, M.; Frosch-Radivo, A. Illustrated Pollen Terminology; Springer International Publishing AG Part of Springer Nature: Cham, Switzerland, 2018; p. 483. [Google Scholar]
- Wodehouse, R.P. The Phylogenetic Value of Pollen-grain Characters. Ann. Bot. 1928, 42, 891–934. [Google Scholar] [CrossRef]
- Ali, M.; Liu, Y.-J.; Xia, Q.-P.; Bahadur, S.; Hussain, A.; Shao, J.-W.; Shuaib, M. Pollen micromorphology of eastern Chinese Polygonatum and its role in taxonomy by using scanning electron microscopy. Microsc. Res. Tech. 2021, 84, 1451–1461. [Google Scholar] [CrossRef]
- Wang, L.; Gu, L.; Zhao, C.; Liu, J. Pollen morphology of Polygonatae and its systematic significance. Palynology 2018, 42, 255–272. [Google Scholar] [CrossRef]
- Deng, X.Y.; Zhou, S.D.; He, X.J. Pollen morphology of 13 Chinese species of Polygonatum and its systematic significations. Plant Sci. J. 2007, 25, 11–18. (In Chinese) [Google Scholar]
- Chien-Ti, C.; Yen-Hsueh, T. Revision of Polygonatum (Asparagaceae, Nolinoideae, Polygonateae) of Taiwan. PhytoKeys 2019, 117, 99–118. [Google Scholar] [CrossRef]
- Zheng, Y.; Sun, Y.G.; Wang, Y.; Shao, J.Z.; Zhang, D.C. Study on the pollen morphology of polygonatum from Anhui, China. Bull. Bot. Res. 1998, 18, 414–417. (In Chinese) [Google Scholar]
- Patil, S.A.; Nimbalkar, M.S.; Pagariya, M.C.; Kulkarni, A.J.; Jadhav, P.R.; Mane, M.P.; Magdum, A.B.; Saha, T.N.; Shinde, K.V.; Prasad, K.V.; et al. Pollen morphology and variability among Indian cultivars of Chrysanthemum morifolium and comparative analysis with genera of the Asteraceae family. Genet. Resour. Crop Evol. 2024. [Google Scholar] [CrossRef]
- Tsymbalyuka, Z.M.; Ivanovab, D.; Nitsenko, L.M. Taxonomic significance of pollen morphology of African Oreobliton, and related genera Aphanisma, Beta, Hablitzia, and Patellifolia (Amaranthaceae). S. Afr. J. Bot. 2024, 169, 134–145. [Google Scholar] [CrossRef]
- Zhang, D.R. Origin study on six Polygonatum in Northeast China. Master’s Thesis, Shenyang Agricultural University, Shenyang, China, 2016. (In Chinese). [Google Scholar]
- Li, C.Q.; Li, Y.; Zhang, Y.M.; Chen, J.C.; Li, M.; Lu, Q. Pollen Specificity Analysis of Nitraria tangutorum in Different Geographic Families. Acta Agric. Boreali-Occident. Sin. 2023, 32, 1437–1444. (In Chinese) [Google Scholar]
- Li, H.C.; Wu, T.Y.; Luo, J. Pollen Morphological Characteristics of 30 Species of Gentianaceae in Shergyla Mountain Area, Tibet. Acta Hortic. Sin. 2021, 48, 2427–2442. (In Chinese) [Google Scholar]
- Sun, Y.; Ding, S.Q.; Shi, Y.M.; Tang, D.Q. Pollen Morphology of 15 Cultivars of Freesia hybrida. Bull. Bot. Res. 2019, 39, 17–26. (In Chinese) [Google Scholar]
- He, L.Q.; Wang, L.H.; Zhuang, Q.G.; Zhang, Y.; Zhang, Q. Observation and analysis of pollen morphology of 41 germplasm of 9 species of kiwifruit. S. China Fruits. 2024, 53, 132–139. (In Chinese) [Google Scholar]
- Wu, Y.; Yang, N.Y.; Song, Y.; Li, X.P.; Guo, W.Z.; Zhang, Y.C.; Zhang, Y.L. Pollen morphological study on 8 Camellia species using different methods and samples. S. China For. Sci. 2023, 51, 1–4. (In Chinese) [Google Scholar]
- Fang, C. Variation of pollen morphology under different observing methods. Master’s Thesis, East China Normal University, Shanghai, China, 2020. (In Chinese). [Google Scholar]
- Bhattacharya, R.; Saha, S.; Kostina, O.; Muravnik, L.; Mitra, A. Replacing critical point drying with a low-cost chemical drying provides comparable surface image quality of glandular trichomes from leaves of Millingtonia hortensis L. f. in scanning electron micrograph. Appl. Microsc. 2020, 50, 15. [Google Scholar] [CrossRef]
- Chissoe, W.F.; Vezey, E.L.; Skvarla, J.J. Hexamethyldisilazane as a Drying Agent for Pollen Scanning Electron Microscopy. Biotech. Histochem. 1994, 69, 192–198. [Google Scholar] [CrossRef]
- Wang, K.F.; Wang, X.Z. Introduction to Pollenology, 2nd ed.; Peking University Press: Beijing, China, 1983; pp. 21–34. (In Chinese) [Google Scholar]
- Walker, J.W. Evolution of exine structure in the pollen of primitive angiosperms. Am. J. Bot. 1974, 61, 891–902. [Google Scholar] [CrossRef]
- Makhkamov, T.; Rahmatov, A.; Zafar, M.; Elshikh, M.S.; Farraj, D.A.A.; Yuldashev, A.; Islamov, S.; Sotiboldiyeva, D.; Makkamov, Z.; Khakimova, D.; et al. Pollen morphology of some Nepeta L. Taxa in Uzbekistan. Am. J. Bot. 2024. [Google Scholar] [CrossRef]
- Marco, G.D.; D’Agostino, A.; Braglia, R.; Redi, E.L.; Iacobelli, S.; Gismondi, A.; Canini, A. Pollen variability in Quercus L. species and relative systematic implications. Plant Physiol. Biochem. 2023, 204, 108079. [Google Scholar] [CrossRef]
- Li, Q.L.; Luo, M.; Qin, M.J.; Lin, M.X.; Zhang, W.W.; Yang, Y. ISSR Analysis of Genetic Diversity Among Eight Polygonatum Species. J. Chin. Med. Mater. 2017, 40, 2042–2045. (In Chinese) [Google Scholar]
- Zhu, Q.; Deng, X.; Zhang, S.B.; Mei, S.Y.; Chen, X.J.; Zhang, J.F.; Xiao, Q.M.; Li, Y. Genetic diversity of 6 species in Polygonatum by SSR marker. China J. Chin. Mater. Med. 2018, 43, 2935–2943. (In Chinese) [Google Scholar]
- Dong, R.J.; Huang, L.; Zhang, Y.Q.; Yin, Y.P.; Liu, P.; Wang, M.J.; Wang, L.Z.; Liang, Q. Electron Microscopic Scanning Analysis of Morphological Characteristics of Pollens from Four Genotypes with Different Flower Color of Polygonatum kingianum Coll. et Hemsl. Southwest China J. Agric. Sci. 2019, 32, 1236–1240. (In Chinese) [Google Scholar]
- Li, T.Q.; Cao, H.J.; Kang, M.S.; Zhang, Z.X.; Zhao, N.; Zhang, H. Pollen Flora of China Woody Plants by SEM; Science Press: Beijing, China, 2011; p. 10. (In Chinese) [Google Scholar]
- Erdtman, G. Handbook of Palynology—An Introduction to the Study of Pollen Grains and Spores; Munksgaard: Copenhagen, Denmark, 1969; p. 496. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | SS | df | MS | F | p |
|---|---|---|---|---|---|
| P/μm | 2163.229 | 45 | 48.072 | 26.248 | 0.000 |
| E/μm | 14,410.124 | 45 | 320.225 | 45.955 | 0.000 |
| P/E | 0.193 | 45 | 0.004 | 7.519 | 0.000 |
| V | 5189.680 | 45 | 115.326 | 45.347 | 0.000 |
| Species | P/μm | E/μm | P/E | V |
|---|---|---|---|---|
| P. cyrtonema | 22.52 ± 0.79 | 58.11 ± 2.63 | 0.39 ± 0.01 | 36.15 ± 1.31 |
| P. filipes | 20.18 ± 1.25 | 53.75 ± 3.35 | 0.38 ± 0.02 | 32.90 ± 1.91 |
| P. kingianum var. grandifolium | 27.15 ± 1.18 | 66.92 ± 2.84 | 0.41 ± 0.02 | 42.61 ± 1.46 |
| P. kingianum | 25.32 ± 1.49 | 67.07 ± 2.44 | 0.38 ± 0.02 | 41.20 ± 1.72 |
| P. alternicirrhosum | 18.17 ± 1.36 | 44.11 ± 2.33 | 0.41 ± 0.03 | 28.30 ± 1.55 |
| P. macropodum | 22.23 ± 1.42 | 57.14 ± 2.81 | 0.39 ± 0.03 | 35.62 ± 1.66 |
| P. odoratum | 20.42 ± 1.16 | 54.55 ± 2.52 | 0.37 ± 0.02 | 33.36 ± 1.43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, M.; Xie, Q.; Jiang, L.; Liu, L.; Luo, W.; Su, H.; Chen, Q. Pollen Morphological Characteristics of 46 Germplasm Resources of Polygonatum and Its Taxonomic Implications. Plants 2024, 13, 3509. https://doi.org/10.3390/plants13243509
Ding M, Xie Q, Jiang L, Liu L, Luo W, Su H, Chen Q. Pollen Morphological Characteristics of 46 Germplasm Resources of Polygonatum and Its Taxonomic Implications. Plants. 2024; 13(24):3509. https://doi.org/10.3390/plants13243509
Chicago/Turabian StyleDing, Mingyue, Qian Xie, Lai Jiang, Lingling Liu, Wenbao Luo, Hailan Su, and Qingxi Chen. 2024. "Pollen Morphological Characteristics of 46 Germplasm Resources of Polygonatum and Its Taxonomic Implications" Plants 13, no. 24: 3509. https://doi.org/10.3390/plants13243509
APA StyleDing, M., Xie, Q., Jiang, L., Liu, L., Luo, W., Su, H., & Chen, Q. (2024). Pollen Morphological Characteristics of 46 Germplasm Resources of Polygonatum and Its Taxonomic Implications. Plants, 13(24), 3509. https://doi.org/10.3390/plants13243509