

Convergent Adaptation of Multiple Herbicide Resistance to Auxin Mimics and ALS- and EPSPS-Inhibitors in Brassica rapa from North and South America

 ,
,  ,
,  ,
, 
 , and
, and
Abstract
1. Introduction
2. Results
2.1. Herbicide-Sensitivity: Cross- and Multiple-Herbicide Resistance
2.2. Dose–Response to TM, 2,4-D, and Glyphosate
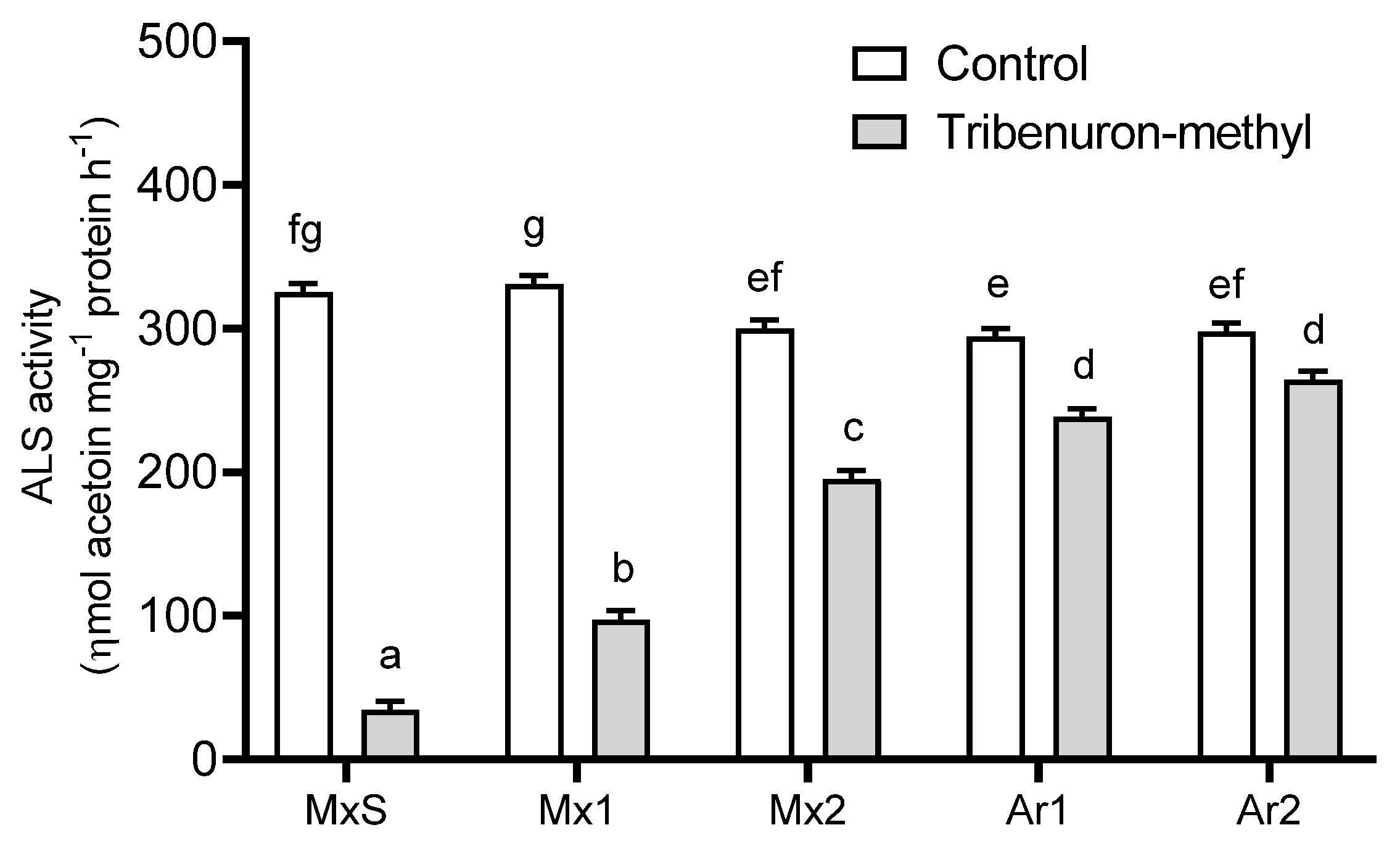
2.3. ALS Enzyme Activity in Response to TM
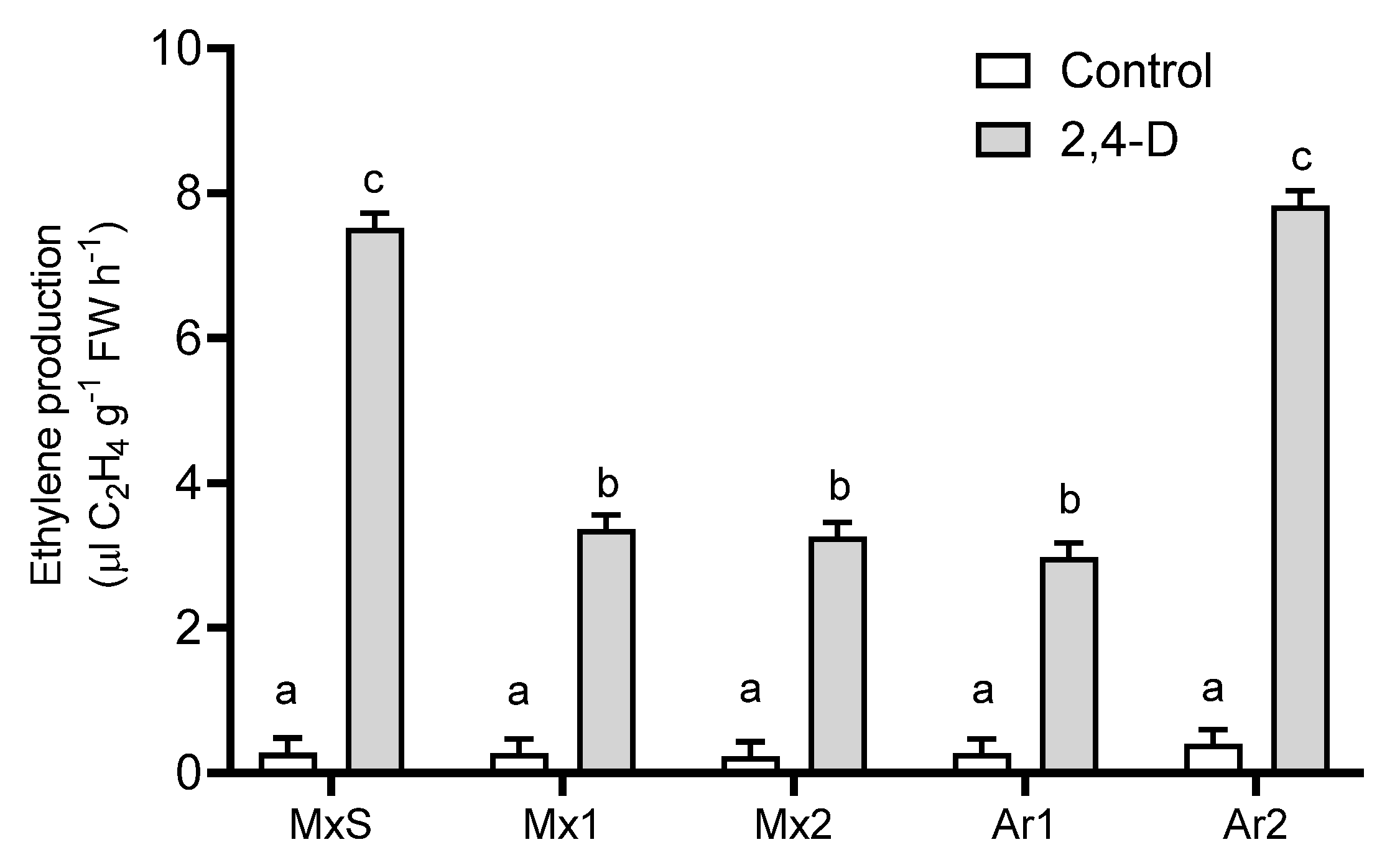
2.4. Ethylene Production in Response to 2,4-D
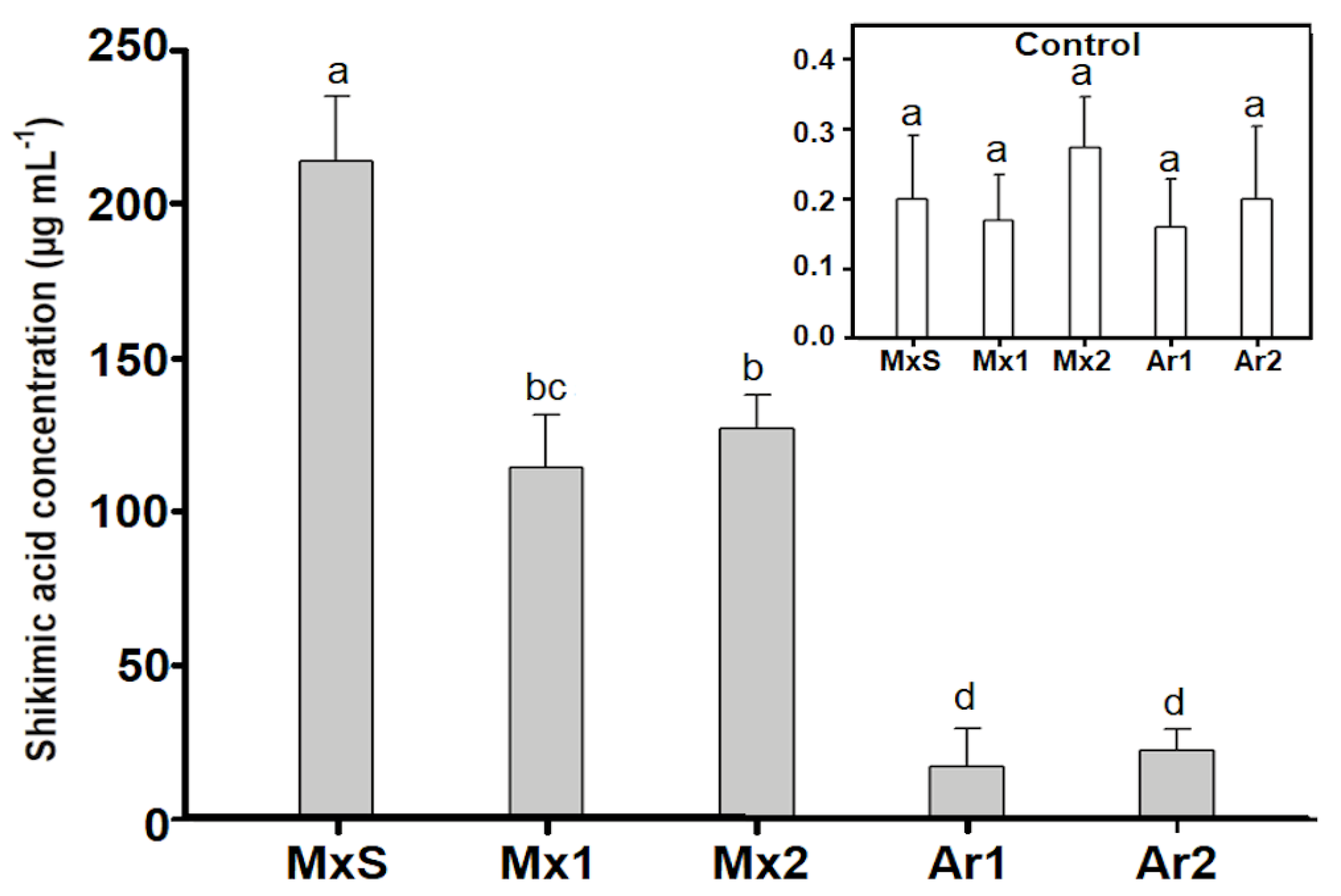
2.5. Shikimic Acid Accumulation Assay
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Materials
5.2. Herbicide-Sensitivity Experiment
5.3. Dose–Response Experiments
5.4. ALS Enzyme Activity in Response to TM
5.5. Ethylene Production in Response to 2,4-D
5.6. Shikimic Acid Accumulation in Response to Glyphosate
5.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dekker, J. Weed diversity and weed management. Weed Sci. 1997, 45, 357–363. [Google Scholar] [CrossRef]
- Clements, D.R.; Vanessa, L.J. Rapid evolution of invasive weeds under climate change: Present evidence and future research needs. Front. Agron. 2021, 3, 664034. [Google Scholar] [CrossRef]
- Neve, P.; Vila-Aiub, M.; Roux, F. Evolutionary-thinking in agricultural weed management. New Phytol. 2009, 184, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Heap, I. Global perspective of herbicide-resistant weeds. Pest Manag. Sci. 2014, 70, 1306–1315. [Google Scholar] [CrossRef]
- Peterson, M.A.; Collavo, A.; Ovejero, R.; Shivrain, V.; Walsh, M.J. The challenge of herbicide resistance around the world: A current summary. Pest Manag. Sci. 2018, 74, 2246–2259. [Google Scholar] [CrossRef]
- Benbrook, C.M. Trends in glyphosate herbicide use in the United States and globally. Environ. Sci. Eur. 2016, 28, 3. [Google Scholar] [CrossRef]
- Alcántara-de la Cruz, R.; Cruz-Hipólito, H.E.; Domínguez-Valenzuela, J.A.; De Prado, R. Glyphosate ban in Mexico: Potential impacts on agriculture and weed management. Pest Manag. Sci. 2021, 77, 3820–3831. [Google Scholar] [CrossRef]
- Duke, S.O.; Powles, S.B.; Sammons, R.D. Glyphosate—How it became a once in a hundred year herbicide and its future. Outlooks Pest Manag. 2018, 29, 247–251. [Google Scholar] [CrossRef]
- Beckie, H.J. Herbicide-resistant weeds: Management tactics and practices. Weed Technol. 2006, 20, 793–814. [Google Scholar] [CrossRef]
- Moss, S.; Ulber, L.; den Hoed, I. A herbicide resistance risk matrix. Crop Prot. 2019, 115, 13–19. [Google Scholar] [CrossRef]
- Heap, I.; Duke, S.O. Overview of glyphosate-resistant weeds worldwide. Pest Manag. Sci. 2018, 74, 1040–1049. [Google Scholar] [CrossRef] [PubMed]
- Braz, G.B.P.; Takano, H.K. Chemical control of multiple herbicide-resistant Amaranthus: A review. Adv. Weed Sci. 2022, 40, e0202200062. [Google Scholar] [CrossRef] [PubMed]
- Jasieniuk, M.; Brûlé-Babel, A.L.; Morrison, I.N. The evolution and genetics of herbicide resistance in weeds. Weed Sci. 1996, 44, 176–193. [Google Scholar] [CrossRef]
- Powles, S.B.; Yu, Q. Evolution in action: Plants resistant to herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef] [PubMed]
- Vencill, W.K.; Nichols, R.L.; Webster, T.M.; Soteres, J.K.; Mallory-Smith, C.; Burgos, N.R.; Johnson, W.G.; McClelland, M.R. Herbicide resistance: Toward an understanding of resistance development and the impact of herbicide-resistant crops. Weed Sci. 2012, 60, 2–30. [Google Scholar] [CrossRef]
- Ureta, M.S.; Torres-Carbonell, F.; Pandolfo, C.; Presotto, A.; Cantamutto, M.A.; Poverene, M. IMI resistance associated to crop-weed hybridization in a natural Brassica rapa population: Characterization and fate. Environ. Monit. Assess. 2017, 189, 101. [Google Scholar] [CrossRef]
- Pandolfo, C.E.; Presotto, A.; Carbonell, F.T.; Ureta, S.; Poverene, M.; Cantamutto, M. Transgene escape and persistence in an agroecosystem: The case of glyphosate-resistant Brassica rapa L. in central Argentina. Environ. Sci. Pollut. Res. 2018, 25, 6251–6264. [Google Scholar] [CrossRef]
- Heap, I. The International Herbicide-Resistant Weed Database. Available online: www.weedscience.org (accessed on 10 March 2023).
- Kreiner, J.M.; Latorre, S.M.; Burbano, H.A.; Stinchcombe, J.R.; Otto, S.P.; Weigel, D.; Wright, S.I. Rapid weed adaptation and range expansion in response to agriculture over the past two centuries. Science 2022, 378, 1079–1085. [Google Scholar] [CrossRef]
- Villaseñor, J.L.; Espinosa-Garcia, F.J. The alien flowering plants of Mexico. Divers. Distrib. 2004, 10, 113–123. [Google Scholar] [CrossRef]
- CONASIPRO—Comité Nacional del Sistema Producto Oleaginosas. Oleaginosas en Cadena. 2006. Available online: http://www.oleaginosas.org/cargas/4o__bolet_n_.pdf (accessed on 10 March 2023).
- Bhullar, M.S.; Kaur, S.; Kaur, T.; Singh, T.; Singh, M.; Jhala, A.J. Control of broadleaf weeds with post-emergence herbicides in four barley (Hordeum spp.) cultivars. Crop Prot. 2013, 43, 216–222. [Google Scholar] [CrossRef]
- Scursoni, J.A.; Vera, A.C.D.; Oreja, F.H.; Kruk, B.C.; de la Fuente, E.B. Weed management practices in Argentina crops. Weed Technol. 2019, 33, 459–463. [Google Scholar] [CrossRef]
- Shahzad, M.; Jabran, K.; Hussain, M.; Raza, M.A.S.; Wijaya, L.; El-Sheikh, M.A.; Alyemeni, M.N. The impact of different weed management strategies on weed flora of wheat-based cropping systems. PLoS ONE 2021, 16, e0247137. [Google Scholar] [CrossRef] [PubMed]
- Tranel, P.J.; Wright, T.R. Resistance of weeds to ALS-inhibiting herbicides: What have we learned? Weed Sci. 2002, 50, 700–712. [Google Scholar] [CrossRef]
- Brown, H.M.; Cotterman, J.C. Recent advances in sulfonylurea herbicides. In Herbicides Inhibiting Branched-Chain Amino Acid Biosynthesis, 1st ed.; Stetter, J., Ed.; Springer: Berlin/Heidelberg, Germany, 1994; Volume 10, pp. 47–81. [Google Scholar]
- Merotto, A., Jr.; Gazziero, D.L.P.; Oliveira, M.C.; Scursoni, J.; Garcia, M.A.; Figueroa, R.; Turra, G.M. Herbicide use history and perspective in South America. Adv. Weed Sci. 2022, 40, e020220050. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Wei, L.; Li, X.; Wang, Y.; Wu, J.; Liu, M.; Zhao, C.; Chen, Z.; Xiao, Z.; Jian, H.; et al. Whole-genome resequencing reveals Brassica napus origin and genetic loci involved in its improvement. Nat. Commun. 2019, 10, 1154. [Google Scholar] [CrossRef]
- Wilkinson, M.J.; Elliott, L.J.; Allainguillaume, J.; Shaw, M.W.; Norris, C.; Welters, R.; Alexander, M.; Sweet, J.; Mason, D.C. Hybridization between Brassica napus and B. rapa on a national scale in the United Kingdom. Science 2003, 302, 457–459. [Google Scholar] [CrossRef]
- Sohn, S.-I.; Thamilarasan, S.K.; Pandian, S.; Oh, Y.-J.; Ryu, T.-H.; Lee, G.-S.; Shin, E.-K. Interspecific Hybridization of Transgenic Brassica napus and Brassica rapa—An Overview. Genes 2022, 13, 1442. [Google Scholar] [CrossRef]
- ASCDMA—Agencia de Servicios a la Comercialización y Desarrollo de Mercados Agropecuarios. La Canola. 2018. Available online: https://www.gob.mx/aserca/articulos/la-canola?idiom=es (accessed on 11 March 2023).
- Busi, R.; Goggin, D.E.; Heap, I.M.; Horak, M.J.; Jugulam, M.; Masters, R.A.; Napier, R.M.; Riar, D.S.; Satchivi, N.M.; Torra, J.; et al. Weed resistance to synthetic auxin herbicides. Pest Manag. Sci. 2018, 74, 2265–2276. [Google Scholar] [CrossRef]
- Juan, V.F.; Fré, F.R.N.; Saint-André, H.M.; Fernández, R.R. Responses of 2,4-D resistant Brassica rapa L. biotype to various 2,4-D formulations and other auxinic herbicides. Crop Prot. 2021, 145, 105621. [Google Scholar] [CrossRef]
- Tafoya-Razo, J.A. Control de maleza resistente a herbicidas en el cultivo de cebada en la región del altiplano. In Memoria del XLIII Congreso Nacional de la Ciencia de la Maleza; Delgado-Castillo, J.C., Rosales-Robles, E., Delgado-Tinoco, J.C., Eds.; 2022; Available online: https://somecima.com/memorias/ (accessed on 11 March 2023).
- Palma-Bautista, C.; Vázquez-García, J.G.; Osuna, M.D.; Garcia-Garcia, B.; Torra, J.; Portugal, J.; De Prado, R. An Asp376Glu substitution in ALS gene and enhanced metabolism confers high tribenuron-methyl resistance in Sinapis alba. Front. Plant Sci. 2022, 13, 1011596. [Google Scholar] [CrossRef]
- Mora, A.D.; Rosario, J.; Rojano-Delgado, A.M.; Palma-Bautista, C.; Torra, J.; Alcántara-de La Cruz, R.; De Prado, R. Multiple resistance to synthetic auxin herbicides and glyphosate in Parthenium hysterophorus occurring in citrus orchards. J. Agric. Food Chem. 2019, 67, 10010–10017. [Google Scholar] [CrossRef] [PubMed]
- Tahmasebi, B.K.; Alcántara-de la Cruz, R.; Alcántara, E.; Torra, J.; Valenzuela, J.A.D.; Cruz-Hipólito, H.E.; Rojano-Delgado, A.M.; De Prado, R. Multiple resistance evolution in bipyridylium-resistant Epilobium ciliatum after recurrent selection. Front. Plant Sci. 2018, 9, 695. [Google Scholar] [CrossRef] [PubMed]
- Yanniccari, M.; Palma-Bautista, C.; Vázquez-García, J.G.; Gigón, R.; Mallory-Smith, C.A.; De Prado, R. Constitutive overexpression of EPSPS by gene duplication is involved in glyphosate resistance in Salsola tragus. Pest Manag. Sci. 2023, 79, 1062–1068. [Google Scholar] [CrossRef] [PubMed]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-response analysis using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Population | MxS | Mx1 | Mx2 | Ar1 | Ar2 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Herbicide | Fw (g) | %Sur | Fw (g) | %Sur | Fw (g) | %Sur | Fw (g) | %Sur | Fw (g) | %Sur | |
| Control | 9.4 ± 1.6 | 100 | 10.1 ± 1.3 | 100 | 9.7 ± 2.6 | 100 | 9.4 ± 0.5 | 100 | 7.6 ± 1.1 | 100 | |
| Tribenuron-methyl | 0.0 | 0.0 | 10.0 ± 3.0 | 100 | 10.0 ± 2.4 | 100 | 7.1 ± 0.9 | 100 | 8.0 ± 1.6 | 100 | |
| Florasulam | 0.0 | 0.0 | 9.3 ± 0.7 | 100 | 10.7 ± 0.7 | 100 | 6.8 ± 0.9 | 100 | 7.1 ± 2.1 | 100 | |
| Imazamox | 0.0 | 0.0 | 10.4 ± 0.6 | 100 | 10.6 ± 0.6 | 100 | 5.4 ± 1.3 | 100 | 4.9 ± 0.5 | 100 | |
| Iodo. + Meso. 1 | 0.0 | 0.0 | 5.4 ± 1.1 | 100 | 6.1 ± 2.4 | 100 | 4.3 ± 0.6 | 100 | 3.8 ± 0.6 | 100 | |
| 2,4-D | 0.0 | 0.0 | 7.5 ± 2.6 | 100 | 6.7 ± 3.7 | 80 | 4.7 ± 0.5 | 100 | 0.0 | 0.0 | |
| Clopyralid | 7.8 ± 0.7 | 100 | 8.4 ± 1.7 | 100 | 5.9 ± 1.9 | 100 | 7.8 ± 1.8 | 100 | 7.2 ± 1.9 | 100 | |
| Dicamba | 6.1 ± 0.9 | 100 | 10.7 ± 1.4 | 100 | 8.1 ± 2.7 | 100 | 6.1 ± 1.8 | 100 | 6.4 ± 0.6 | 100 | |
| Fluroxypyr | 0.0 | 0.0 | 7.9 ± 1.4 | 100 | 7.1 ± 1.2 | 100 | 3.3 ± 0.2 | 100 | 0.0 | 0.0 | |
| MCPA | 0.0 | 0.0 | 6.2 ± 2.5 | 100 | 3.8 ± 2.7 | 90 | 4.0 ± 0.4 | 100 | 0.0 | 0.0 | |
| Glyphosate | 0.0 | 0.0 | 3.8 ± 0.4 | 70 | 4.2 ± 0.5 | 70 | 10.0 ± 1.2 | 100 | 8.1 ± 0.7 | 100 | |
| Atrazine | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Bromoxynil | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Glufosinate | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Mesotrione | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Herbicide | Population | Dry Weight Reduction | % Plant Survival | ||||||
|---|---|---|---|---|---|---|---|---|---|
| d | b | GR50 (95% CI) | RF | d | b | LD50 (95% CI) | RF | ||
| Tribenuron-methyl | MxS | 100.2 | 2.15 | 0.28 (0.25–0.31) | -- | 100.1 | 7.7 | 0.54 (0.43–0.66) | -- |
| Mx1 | 100.2 | 1.3 | 74.4 (56.9–92.9) | 266 | 100.7 | 3.4 | 511 (430–593) | 947 | |
| Mx2 | 104.2 | 1.3 | 259 (204–314) | 925 | 100.1 | 3.4 | 1535 (1294–1775) | 2842 | |
| Ar1 | 100.6 | 1.1 | 405 (296–513) | 1445 | 100.9 | 2.7 | 1434 (1182–1687) | 2656 | |
| Ar2 | 99.9 | 1.4 | 664 (530–797) | 2371 | 100.1 | 4.0 | 2197 (1875–2519) | 4069 | |
| 2,4-D | MxS | 99.6 | 2.7 | 43.3 (39.4–47.2) | -- | 100.5 | 3.7 | 117 (106–128) | -- |
| Mx1 | 96.2 | 0.96 | 398 (335–461) | 9.2 | 99.7 | 3.3 | 584 (553–614) | 5.0 | |
| Mx2 | 94.5 | 0.96 | 385 (314–456) | 8.9 | 100.1 | 94 | 675 (608–743) | 5.8 | |
| Ar1 | 99.0 | 1.2 | 337 (263–411) | 7.8 | 100.1 | 4.0 | 1099 (962–1235) | 9.4 | |
| Ar2 | 99.7 | 1.5 | 48.7 (38.8–58.5) | 1.1 | 101.2 | 3.4 | 182 (158–207) | 1.5 | |
| Glyphsate 2 | MxS | 100.1 | 0.7 | 97 (76–118) | -- | 100.7 | 2.9 | 225 (214–236) | -- |
| Mx1 | 98.5 | 1.7 | 355 (314–395 | 3.7 | 100.0 | 14.6 | 793 (643–943) | 3.5 | |
| Mx2 | 98.3 | 2.3 | 329 (301–357) | 3.4 | 100.0 | 10.8 | 600 (457–743) | 2.7 | |
| Ar1 | 99.6 | 1.4 | 5153 (4270–6036) | 53.0 | 100.3 | 2.9 | 9505 (8471–10,540) | 42.3 | |
| Ar2 | 99.2 | 1.9 | 2351 (2016–2687) | 24.2 | 100.5 | 2.2 | 6849 (5995–7703) | 30.5 | |
| Population | Location, Country | Crops | Herbicide History | Coordinates |
|---|---|---|---|---|
| Ar1 | Tandil, Arg. | Fallow and wheat | Glyphosate, metsulfuron-methyl, 2,4-D | 37°13′22.8″ S 59°17′42.0″ W |
| Ar2 | San Cayetano, Arg. | Fallow and wheat | Glyphosate, imazamox, 2,4-D | 38°17′42.0″ S 59°23′09.6″ W |
| MxS | Texcoco, Mex. | No crops | Non treated | 19°29′34.8″ N 98°52′30.0″ W |
| Mx1 | Tlaxcala, Mex. | Barley | Glyphosate, iodosulfuron-methyl sodium + mesosulfuron-methyl, 2,4-D amine | 19°36′14.4″ N 98°09′57.6″ W |
| Mx2 | Tlaxcala, Mex. | Barley | Glyphosate, iodosulfuron-methyl sodium + mesosulfuron-methyl, 2,4-D amine | 19°32′16.8″ N 98°09′57.6″ W |
| MoA/HRAC Group | Herbicide | Manufacturer | Commercial Product | Rate |
|---|---|---|---|---|
| ALS inhibitor/2 | Tribenuron-methyl | Nufarm | 75% w/v, Primma® SL | 20 |
| Florasulam | Nufarm | 5% w/v, Fragma® SL | 5 | |
| Imazamox | BASF | 4% w/v, Pulsar® 40 SC | 40 | |
| Iodosulfuron + Mesosulfuron | Bayer CropScience | 5% + 0.75% w/w/v, Hussar® Plus | 150 1 | |
| Auxin mimics/4 | 2,4-D | Nufarm | 60% w/v, U 46 D Complet® SL | 600 |
| Clopyralid | Corteva | 72% w/v, Lontrel® WG | 300 | |
| Dicamba | Syngenta | 48% w/v, Banvel® D WG | 150 | |
| MCPA | Nufarm | 40% w/v, Procer M-40® | 800 | |
| Fluroxypyr | Nufarm | 20% w/v, Praxis® EC | 200 | |
| EPSPS inhibitor/9 | Glyphosate | Bayer CropScience | 36% w/v, Roundup® SL | 960 2 |
| PSII inhibitor/5 | Atrazine | Syngenta | 90% w/v, Gesaprim® SL | 1550 |
| PSII inhibitor/6 | Bromoxynil | Nufarm | 38.5% w/v, Emblem Flo® SC | 385 |
| GS inhibitor/10 | Glufosinate | BASF | 15% w/v, Finale® SL | 600 |
| HPPD inhibitor/27 | Mesotrione | Syngenta | 48% w/v, Callisto® SC | 150 |
| Herbicide | Dose (g ai ha−1) |
|---|---|
| 2,4-D | Susceptible: 0, 40, 80, 160, 320, 640, and 1280 Resistant: 0, 40, 80, 160, 320, 640, and 1280 |
| Glyphosate 1 | Susceptible: 0, 10, 20, 40, 80, 160, 320, and 640 Resistant: 0, 80, 160, 320, 640, 1280, 2560, 5120, and 10,240 |
| Tribenuron-methyl | Susceptible: 0, 0.125, 0.25, 0.5, 1, 2, 4, and 10 Resistant: 0, 20, 40, 80, 160, 320, 640, 1280, and 2560 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dominguez-Valenzuela, J.A.; Palma-Bautista, C.; Vazquez-Garcia, J.G.; Yanniccari, M.; Gigón, R.; Alcántara-de la Cruz, R.; De Prado, R.; Portugal, J. Convergent Adaptation of Multiple Herbicide Resistance to Auxin Mimics and ALS- and EPSPS-Inhibitors in Brassica rapa from North and South America. Plants 2023, 12, 2119. https://doi.org/10.3390/plants12112119
Dominguez-Valenzuela JA, Palma-Bautista C, Vazquez-Garcia JG, Yanniccari M, Gigón R, Alcántara-de la Cruz R, De Prado R, Portugal J. Convergent Adaptation of Multiple Herbicide Resistance to Auxin Mimics and ALS- and EPSPS-Inhibitors in Brassica rapa from North and South America. Plants. 2023; 12(11):2119. https://doi.org/10.3390/plants12112119
Chicago/Turabian StyleDominguez-Valenzuela, José Alfredo, Candelario Palma-Bautista, José G. Vazquez-Garcia, Marcos Yanniccari, Ramón Gigón, Ricardo Alcántara-de la Cruz, Rafael De Prado, and João Portugal. 2023. "Convergent Adaptation of Multiple Herbicide Resistance to Auxin Mimics and ALS- and EPSPS-Inhibitors in Brassica rapa from North and South America" Plants 12, no. 11: 2119. https://doi.org/10.3390/plants12112119
APA StyleDominguez-Valenzuela, J. A., Palma-Bautista, C., Vazquez-Garcia, J. G., Yanniccari, M., Gigón, R., Alcántara-de la Cruz, R., De Prado, R., & Portugal, J. (2023). Convergent Adaptation of Multiple Herbicide Resistance to Auxin Mimics and ALS- and EPSPS-Inhibitors in Brassica rapa from North and South America. Plants, 12(11), 2119. https://doi.org/10.3390/plants12112119