


Photosynthetic Traits of Páramo Plants Subjected to Short-Term Warming in OTC Chambers
 , , , , ,
, , , , ,  ,
,
Abstract
1. Introduction
2. Results
2.1. Climate
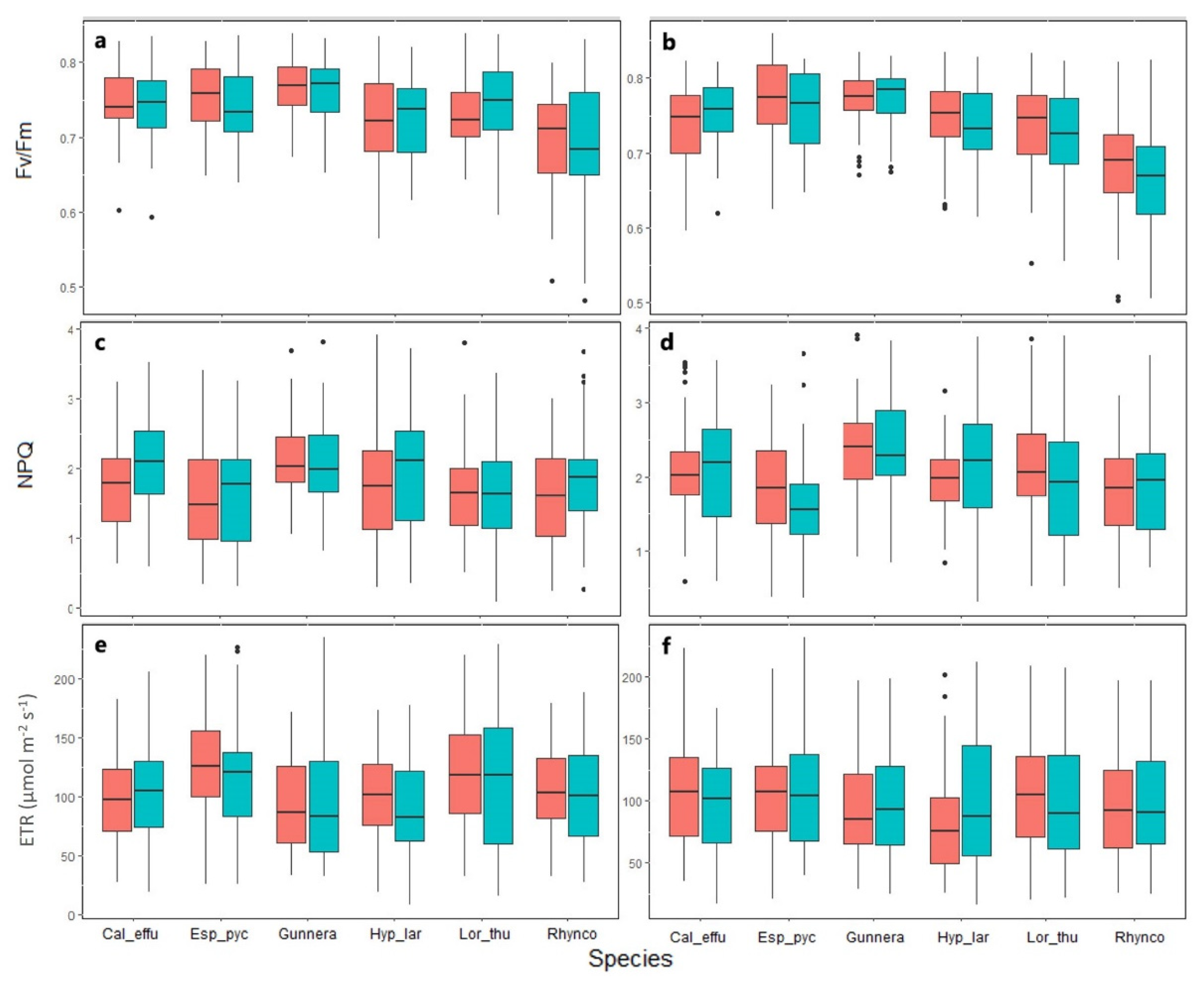
2.2. Chlorophyll a Fluorescence
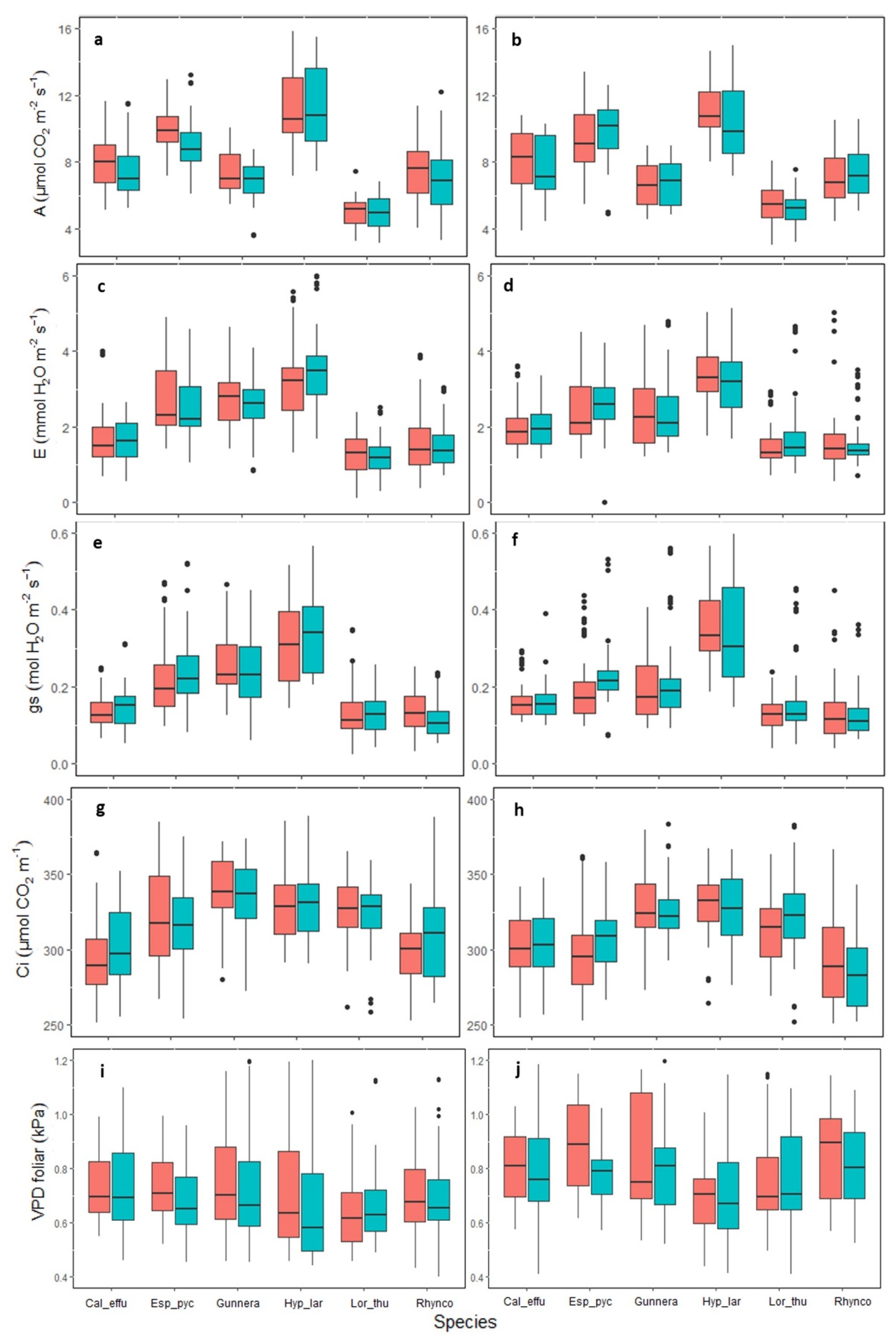
2.3. Gas Exchange Variables
2.4. Response of Photosynthesis to Light (An/PPFD) and to CO2 Concentration (An/Ci)
3. Discussion
4. Materials and Methods
4.1. Study Area
4.2. Plant Material and Experimental Design
4.3. Climatic Variables
4.4. Chlorophyll a Fluorescence
4.5. Gas exchange Variables
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Intergovernmental Panel on Climate Change (IPCC). Summary for Policymakers. In Climate Change 2021: The Physical Science Basis; Working Group I Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2021. [Google Scholar]
- Vuille, M.; Carey, M.; Huggel, C.; Buytaert, W.; Rabatel, A.; Jacobsen, D.; Soruco, A.; Villacis, M.; Yarleque, C.; Timm, O.E.; et al. Rapid decline of snow and ice in the tropical Andes–Impacts, uncertainties and challenges ahead. Earth-Sci. Rev. 2018, 176, 195–213. [Google Scholar] [CrossRef]
- Pepin, N.; Bradley, R.S.; Diaz, H.F.; Baraer, M.; Caceres, E.B.; Forsythe, N.; Fowler, H.; Greenwood, G.; Hashmi, M.Z.; Liu, X.D.; et al. Elevation-dependent warming in mountain regions of the world. Nat. Clim. Chang. 2015, 5, 424–430. [Google Scholar]
- Yao, T.; Xue, Y.; Chen, D.; Chen, F.; Thompson, L.; Cui, P.; Koike, T.; Lau, W.K.-M.; Lettenmaier, D.; Mosbrugger, V.; et al. Recent third pole’s rapid warming accompanies cryospheric melt and water cycle intensification and interactions between monsoon and environment: Multidisciplinary approach with observations, modeling, and analysis. Bull. Am. Meteorol. Soc. 2019, 100, 423–444. [Google Scholar] [CrossRef]
- Verrall, B.; Pickering, C.M. Alpine vegetation in the context of climate change: A global review of past research and future directions. Sci. Total Environ. 2020, 748, 141344. [Google Scholar] [CrossRef]
- Dusenge, M.E.; Duarte, A.G.; Way, D.A. Plant carbon metabolism and climate change: Elevated CO2 and temperature impacts on photosynthesis, photorespiration and respiration. New Phytol. 2019, 221, 32–49. [Google Scholar] [CrossRef]
- Heikkinen, R.K.; Luoto, M.; Araujo, M.B.; Virkkala, R.; Thuiller, W.; Sykes, M.T. Methods and uncertainties in bioclimatic envelope modelling under climate change. Prog. Phys. Geogr. 2006, 30, 751–777. [Google Scholar] [CrossRef]
- Woodward, F.I.; Williams, B.G. Climate and plant distribution at global and local scales. Vegetatio 1987, 69, 189–197. [Google Scholar] [CrossRef]
- Luteyn, J.L. Páramos: A Checklist of Plant Diversity, Geographic Distribution, and Botanical Literature; Memoirs of the New York Botanical Garden: New York, NY, USA, 1999; p. 84. [Google Scholar]
- Young, B.; Josse, C.; Stern, M.; Vasconez, S.; Olander, J.; Sanchez, A.; Zador, M.; Smyth, R.; Comer, P.; Moull, K.; et al. Hots-pot de Biodiversidad de los Andes Tropicales. Nat. Y EcoDecisión, 2015. Available online: https://www.cepf.net/sites/default/files/tropicalandes_techsummary_sp.pdf (accessed on 15 October 2022).
- Buytaert, W.; Cuesta-Camacho, F.; Tobón, C. Potential impacts of climate change on the environmental services of humid tropical alpine regions. Glob. Ecol. Biogeogr. 2011, 20, 19–33. [Google Scholar] [CrossRef]
- Díaz-Granados Ortiz, M.; Navarrete González, J.; Suárez López, T. Páramos: Hidro Sistemas Sensibles. 2005. Available online: Http://www.Scielo.Org.Co/Scielo.Php?Script=Sci_Arttext&Pid=S0121-49932005000200008 (accessed on 15 October 2022).
- Daza Torres, M.; Hernández Florez, F.; Triana Flor, A. Efecto del Uso del Suelo en la Capacidad de Almacenamiento Hídrico en el Páramo de Sumapaz–Colombia. Rev. Fac. Nac. Agric. Medellín 2014, 67, 7189–7200. [Google Scholar] [CrossRef]
- Sarmiento, F.O. Anthropogenic change in the landscapes of highland Ecuador. Geogr. Rev. 2020, 92, 213–234. [Google Scholar] [CrossRef]
- Sklenář, P.; Romoleroux, K.; Muriel, P.; Jaramillo, R.; Bernardi, A.; Diazgranados, M.; Moret, P. Distribution changes in páramo plants from the equatorial high Andes in response to increasing temperature and humidity variation since 1880. Alp. Bot. 2021, 131, 201–212. [Google Scholar] [CrossRef]
- Madriñan, S.; Cortés, A.; Richardson, E. Páramo is the world’s fastest evolving and coolest biodiversity hotspot. Front. Genet. 2013, 4, 192. [Google Scholar] [CrossRef]
- Körner, C.; Hiltbrunner, E. Why Is the Alpine Flora Comparatively Robust against Climatic Warming? Diversity 2021, 13, 383. [Google Scholar] [CrossRef]
- Cuesta, F.; Peralvo, M.; Valarezo, N. Los Bosques Montanos de Los Andes Tropicales. Una Evaluación de su Estado de Conservación y de su Vulnerabilidad a Los Efectos del Cambio Climático; Serie Investigación y Sistematización #5; Programa Regional Ecobona Intercooperation: Quito, Ecuador, 2009. [Google Scholar]
- Bendix, J.; Behling, H.; Peters, T.; Richter, M.; Beck, E. Functional Biodiversity and Climate Change along an Altitudinal Gradient in a Tropical Mountain Rainforest; Tscharntke, T., Veldkamp, C.E., Faust, H., Guhardja, E., Bidin, A., Eds.; Tropical Rainforests and Agroforests under Global Change; Springer: Berlin, Germany, 2010; pp. 239–268. [Google Scholar]
- Morueta-Holme, N.; Engemann, K.; Sandoval-Acuña, P.; Jonas, J.D.; Segnitz, R.M.; Svenning, J.-C. Strong upslope shifts in Chimborazo’s vegetation over two centuries since Humboldt. Proc. Natl. Acad. Sci. USA 2015, 112, 12741–12745. [Google Scholar] [CrossRef]
- Hernández-Fuentes, C.; Bravo, L.A.; Cavieres, L.A. Photosynthetic responses and photoprotection strategies of Phacelia secunda plants exposed to experimental warming at different elevations in the central Chilean Andes. Alp. Bot. 2015, 125, 87–99. [Google Scholar] [CrossRef]
- Lasso, E.; Matheus-Arbeláez, P.; Gallery, R.E.; Garzón-López, C.; Cruz, M.; Leon-Garcia, I.V.; Curiel Yuste, J. Homeostatic Response to Three Years of Experimental Warming Suggests High Intrinsic Natural Resistance in the Páramos to Warming in the Short Term. Front. Ecol. Evol. 2021, 9, 615006. [Google Scholar] [CrossRef]
- Welker, J.M.; Fahnestock, J.T.; Henry, G.H.R.; O’dea, K.W.; Chimner, R.A. CO2 exchange in three Canadian High Arctic ecosystems: Response to long-term experimental warming. Glob. Chang. Biol. 2004, 10, 1981–1995. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Reum Han, A.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Zhou, Y.; Deng, J.; Tai, Z.; Jiang, L.; Han, J.; Meng, G.; Li, M.-H. Leaf Anatomy, Morphology and Photosynthesis of Three Tundra Shrubs after 7-Year Experimental Warming on Changbai Mountain. Plants 2019, 8, 271. [Google Scholar] [CrossRef]
- Yamori, W.; Hikosaka, K.; Way, D.A. Temperature response of photosynthesis in C3, C4, and CAM plants: Temperature acclimation and temperature adaptation. Photosynth. Res. 2014, 119, 101–117. [Google Scholar] [CrossRef]
- Way, D.A.; Yamori, W. Thermal acclimation of photosynthesis: On the importance of adjusting our definitions and accounting for thermal acclimation of respiration. Photosyn Res. 2014, 119, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Salvucci, M.E.; Crafts-Brandner, S.J. Inhibition of photosynthesis by heat stress: The activation state of Rubisco as a limiting factor in photosynthesis. Physiol. Plant. 2004, 120, 179–186. [Google Scholar] [CrossRef]
- Sharkey, T.D. Effects of moderate heat stress on photosynthesis: Importance of thylakoid reactions, rubisco deactivation, reactive oxygen species, and thermotolerance provided by isoprene. Plant Cell Environ. 2005, 28, 269–277. [Google Scholar] [CrossRef]
- Wang, H.; Atkin, O.K.; Keenan, T.F.; Smith, N.G.; Wright, I.J.; Bloomfield, K.J.; Kattge, J.; Reich, P.B.; Prentice, I.C. Acclimation of leaf respiration consistent with optimal photosynthetic capacity. Glob. Chang. Biol. 2020, 26, 2573–2583. [Google Scholar] [CrossRef]
- Sáez, P.L.; Galmés, J.; Ramírez, C.F.; Poblete, L.; Rivera, B.K.; Cavieres, L.A.; Clemente-Moreno, M.J.; Flexas, J.; Bravo, L. Mesophyll conductance to CO2 is the most significant limitation to photosynthesis at different temperatures and water availabilities in Antarctic vascular species. Environ. Exp. Bot. 2018, 156, 279–287. [Google Scholar] [CrossRef]
- Cavieres, L.A.; Sáez, P.; Sanhueza, C.; Sierra-Almeida, A.; Rabert, C.; Corcuera, L.J.; Alberdi, M.; Bravo, L.A. 2016. Ecophysiological traits of Antarctic vascular plants: Their importance in the responses to climate change. Plant Ecol. 2016, 217, 343–358. [Google Scholar] [CrossRef]
- Allison, S.D.; Romero-Olivares, A.L.; Lu, Y.W.; Treseder, K.K. Temperature sensitivities of extracellular enzyme Vmax and Km across thermal environments. Glob. Chang. Biol. 2018, 24, 2884–2897. [Google Scholar] [CrossRef]
- Samaniego, L.; Thober, S.; Kumar, R.; Wanders, N.; Rakovec, O.; Pan, M.; Zink, M.; Sheffield, J.; Wood, E.F.; Marx, A. Anthropogenic warming exacerbates European soil moisture droughts. Nat. Clim. 2018, 8, 421. [Google Scholar] [CrossRef]
- Bhusal, N.; Adhikari, A.; Lee, M.; Han, A.; Han, A.R.; Kim, H.S. Evaluation of growth responses of six gymnosperm species under long-term excessive irrigation and traits determining species resistance to waterlogging. Agric. For. Meteorol. 2022, 323, 109071. [Google Scholar] [CrossRef]
- Reich, P.B.; Sendall, K.M.; Stefanski, A.; Rich, R.L.; Hobbie, S.E.; Montgomery, R.A. Effects of climate warming on photosynthesis in boreal tree species depend on soil moisture. Nature 2018, 562, 263–267. [Google Scholar] [CrossRef]
- Shen, H.; Dong, S.; Li, S. Effects of Warming and N deposition on the physiological performances of Leymus secalinus in alpine meadow of Qinghai-Tibetan plateau. Front. Plant Sci. 2020, 10, 1804. [Google Scholar] [CrossRef] [PubMed]
- Elmendorf, S.C.; Henry, G.H.; Hollister, R.D.; Björk, R.G.; Bjorkman, A.D.; Callaghan, T.V.; Collier, L.S.; Cooper, E.J.; Cornelissen, J.H.; Day, T.A.; et al. Global assessment of experimental climate warming on tundra vegetation: Heterogeneity over space and time. Ecol. Lett. 2012, 15, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.Y.; Xia, J.Y.; Liu, L.L.; Wan, S.Q. Global patterns of the responses of leaf-level photosynthesis and respiration in terrestrial plants to experimental warming. J. Plant Ecol. 2013, 6, 437–447. [Google Scholar] [CrossRef]
- Atkin, O.K.; Tjoelker, M.G. Thermal acclimation and the dynamic response of plant respiration to temperature. Trends Plant Sci. 2003, 8, 343–351. [Google Scholar] [CrossRef]
- Way, D.A.; Sage, R.F. Thermal acclimation of photosynthesis in black spruce Picea mariana (Mill.) BSP. Plant Cell Environ. 2008, 31, 1250–1262. [Google Scholar] [CrossRef]
- Way, D.A.; Oren, R. Differential responses to changes in growth temperature between trees from different functional groups and biomes: A review and synthesis of data. Tree Physiol. 2010, 30, 669–688. [Google Scholar] [CrossRef]
- Crous, K.Y.; Quentin, Y.S. Photosynthesis of temperate Eucalyptus globulus trees outside their native range has limited adjustment to elevated CO2 and climate warming. Glob. Chang. Biol. 2013, 19, 3790–3807. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). Climate Change: Impacts, Adaptation and Vulnerability. In Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007; pp. 1–6. [Google Scholar]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence-a practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C.; Knapp, N. How to correctly determine the different chlorophyll fluorescence parameters and the chlorophyll fluorescence decrease ratio Rfd of leaves with the PAM fluorometer. Photosynthetica 2005, 43, 379–393. [Google Scholar] [CrossRef]
- Shi, F.S.; Wu, Y.; Wu, N.; Luo, P. Different growth and physiological responses to experimental warming of two dominant plant species Elymus nutans and Potentilla anserina in an alpine meadow of the eastern Tibetan Plateau. Photosynthetica 2010, 48, 437–445. [Google Scholar] [CrossRef]
- Lawlor, D.; Tezara, W. Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells: A critical evaluation of mechanisms and integration of processes. Ann. Bot. 2009, 103, 561–579. [Google Scholar] [CrossRef]
- Tezara, W.; Mitchell, V.; Driscoll, S.P.; Lawlor, D.W. Effects of water deficit and its interaction with CO2 supply on the biochemistry and physiology of photosynthesis in sunflower. J. Exp. Bot. 2002, 53, 1781–1791. [Google Scholar] [CrossRef]
- Wilhelm, C.; Selmar, D. Energy dissipation is an essential mechanism to sustain the viability of plants: The physiological limits of improved photosynthesis. J. Plant Physiol. 2011, 168, 79–87. [Google Scholar] [CrossRef]
- Albert, K.R.; Mikkelsen, T.N.; Michelsen, A.; Ro-Poulsen, H.; Van der Linden, L. Interactive effects of drought, elevated CO2 and warming on photosynthetic capacity and photosystem performance in temperate heath plants. J. Plant Physiol. 2011, 168, 1550–1561. [Google Scholar] [CrossRef]
- Buchner, O.; Stoll, M.; Karadar, M.; Kranner, I.; Neuner, G. Application of heat stress in situ demonstrates a protective role of irradiation on photosynthetic performance in alpine plants. Plant Cell Environ. 2015, 38, 812–826. [Google Scholar] [CrossRef]
- Magaña Ugarte, R.; Escudero, A.; Gavilán, R.G. Metabolic and physiological responses of Mediterranean high-mountain and Alpine plants to combined abiotic stresses. Physiol. Plant. 2018, 165, 403–412. [Google Scholar] [CrossRef]
- Goetz, S.J.; Bunn, A.G.; Fiske, G.J.; Houghton, R.A. Satellite-observed photosynthetic trends across boreal North America associated with climate and fire disturbance. Proc. Natl. Acad. Sci. USA 2005, 102, 13521–13525. [Google Scholar] [CrossRef]
- Yang, X.; Li, Y.; Chen, H.; Huang, J.; Zhang, Y.; Qi, M.; Liu, Y.; Li, T. Photosynthetic response mechanism of soil salinity-induced cross-tolerance to subsequent drought stress in tomato plants. Plants 2020, 9, 363. [Google Scholar] [CrossRef]
- Bravo, L.; Saavedra-Mella, F.; Vera, F.; Guerra, A.; Cavieres, L.; Ivanov, A.; Huner, N.; Corcuera, L. Effect of cold acclimation on the photosynthetic performance of two ecotypes of Colobanthus quitensis (Kunth) Bartl. J. Exp. Bot. 2007, 58, 3581–3590. [Google Scholar] [CrossRef]
- Athanasiou, K.; Dyson, B.C.; Webster, R.E.; Johnson, G.N. Dynamic acclimation of photosynthesis increases plant fitness in changing environments. Plant Physiol. 2010, 152, 366–373. [Google Scholar] [CrossRef]
- Flexas, J.; Zhang, Y.; Gulías, J.; Xiong, D.; Carriquí, M.; Baraza, E.; Xu, W. Leaf physiological traits of plants from the Qinghai-Tibet Plateau and other arid sites in China: Identifying susceptible species and well-adapted extremophiles. J. Plant Physiol. 2022, 272, 153689. [Google Scholar] [CrossRef] [PubMed]
- Crous, K.Y. Plant responses to climate warming: Physiological adjustments and implications for plant functioning in a future, warmer world. Am. J. Bot. 2019, 106, 1049. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Su, P.; Wu, X.; Shi, R.; Ding, X. Leaf and community photosynthetic carbon assimilation of alpine plants under in-situ warming. Front. Plant Sci. 2021, 12, 1151. [Google Scholar] [CrossRef] [PubMed]
- Fauset, S.; Oliveira, L.; Buckeridge, M.S.; Foyer, C.H.; Galbraith, D.; Tiwari, R. Contrasting responses of stomatal conductance and photosynthetic capacity to warming and elevated CO2 in the tropical tree species Alchornea glandulosa under heatwave conditions. Environ. Exp. Bot. 2019, 158, 28–39. [Google Scholar] [CrossRef]
- Instituto de Hidrología, Meteorología y Estudios Ambientales (IDEAM). Inventario de Emisiones de Gases Efecto Invernadero para la Región Bogotá–Cundinamarca, 1st ed.; Instituto de Hidrología, Meteorología y Estudios Ambientales (IDEAM): Cundinmarca, Colombia, 2012. [Google Scholar]
- Marion, G.M.; Pidgeon, D.E. Passive Techniques for Manipulating Field Soil Temperatures; Cold Regions Research and Engineering Lab: Hanover, NH, USA, 1992. [Google Scholar]
- Baker, N.R. Chlorophyll Fluorescence: A Probe of Photosynthesis In Vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef]
- Von Caemmerer, S. Biochemical Models of Leaf Photosynthesis, 2nd ed.; CSIRO Publishing: Clayton, Australia, 2000; pp. 1–65. [Google Scholar]
- Varela, S.; Caballé, G.; Curetti, M.; Portela, J. Sistemas Irga en la Medición de Intercambio Gaseoso. In En Técnicas en Medición en Ecofisiología Vegetal: Conceptos y Procedimientos, 1st ed.; Fernández, M.E., Gyenge, E., Eds.; Ediciones INTA: Buenos Aires, Argentina, 2010; pp. 89–108. [Google Scholar]
- Valladares, F.; Niinemets, Ü. Shade Tolerance, a Key Plant Feature of Complex Nature and Consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef]
- de Lobo, F.A.; de Barros, M.P.; Dalmagro, H.J.; Dalmolin, Â.C.; Pereira, W.E.; de Souza, É.C.; Vourlitis, G.L.; Ortíz, C.E. Fitting net photosynthetic light-response curves with Microsoft Excel-a critical look at the models. Photosynthetica 2013, 51, 445–456. [Google Scholar] [CrossRef]
- Kaipiainen, E. Parameters of photosynthesis light curve in Salix dasyclados and their changes during the growth season. Russ. J. Plant Physiol. 2009, 56, 445–453. [Google Scholar] [CrossRef]
- Long, S.P.; Bernacchi, C.J. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. J. Exp. Bot. 2003, 54, 2393–2401. [Google Scholar] [CrossRef]
- Ethier, G.J.; Livingston, N.J. On the need to incorporate sensitivity to CO2 transfer conductance into the Farquhar-von Caemmerer-Berry leaf photosynthesis model. Plant Cell Environ. 2004, 27, 137–153. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Bernacchi, C.J.; Farquhar, G.D.; Singsaas, E.L. Fitting photosynthetic carbon dioxide response curves for C3 leaves. Plant Cell Environ. 2007, 30, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, T.D. What gas exchange data can tell us about photosynthesis. Plant Cell Environ. 2015, 39, 1161–1163. [Google Scholar] [CrossRef] [PubMed]
- Hedges, L.; Olkin, I. Statistical Methods for Meta-Analysis; Academic Press: New York, NY, USA, 1985. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conserved Area | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Dec-20 | Apr-21 | Aug-21 | Jan-22 | ||||||
| Amax (μmol m−2 s−1) | OTC | Control | OTC | Control | OTC | Control | OTC | Control | Treatment Significance |
| C. effusa | 10.9 ± 0.04 | 10.7 ± 0.1 | 7.9 ± 0.1 | 7.7 ± 0.04 | 8.3 ± 0.6 | 7.4 ± 0.4 | 10.4 ± 1.7 | 7.5 ± 0.1 | p < 0.01 |
| E. pycnophylla | 17.7 ± 1.7 | 15.1 ± 0.9 | 9.3 ± 0.2 | 9.0 ± 0.1 | 13.2 ± 0.1 | 11.7 ± 0.3 | 11.9 ± 2.1 | 8.5 ± 2.1 | |
| G. magellanica | 10.3 ± 0.2 | 9.8 ± 0.3 | 7.6 ± 0.1 | 7.4 ± 0.1 | 7.1 ± 1.1 | 7.4 ± 0.1 | 8.4 ± 0.5 | 7.2 ± 0.02 | |
| H. laricifolium | 14.0 ± 0.1 | 13.7 ± 0.1 | 8.8 ± 0.01 | 8.7 ± 0.04 | 12.2 ± 1.9 | 9.9 ± 0.1 | 11.5 ± 0.3 | 12.5 ± 0.4 | |
| L. thuyoides | 13.1 ± 0.2 | 12.2 ± 0.03 | 8.5 ± 0.01 | 8.4 ± 0.04 | 6.0 ± 0.9 | 5.6 ± 0.7 | 6.0 ± 0.3 | 4.9 ± 0.2 | |
| R. macrochaeta | 11.7 ± 0.4 | 11.1 ± 0.1 | 8.2 ± 0.1 | 8.1 ± 0.03 | 9.2 ± 0.7 | 10.1 ± 0.3 | 7.8 ± 1.8 | 5.2 ± 0.5 | |
| Rd (μmol CO2 m−2 s−1) | |||||||||
| C. effusa | 0.4 ± 0.01 | 0.4 ± 0.1 | 0.7 ± 0.1 | 0.8 ± 0.1 | 0.4 ± 0.1 | 0.4 ± 0.1 | 1 ± 0.2 | 0.8 ± 0.04 | p > 0.05 |
| E. pycnophylla | 0.6 ± 0.1 | 0.7 ± 0.2 | 0.8 ± 0.2 | 1.0 ± 0.3 | 0.7 ± 0 | 0.7 ± 0.02 | 0.7 ± 0.2 | 0.9 ± 0.3 | |
| G. magellanica | 0.9 ± 1.1 | 0.9 ± 0.6 | 0.9 ± 0.6 | 0.5 ± 0.04 | 0.4 ± 0.2 | 0.6 ± 0.1 | 1.1 ± 0.4 | 0.5 ± 0.02 | |
| H. laricifolium | 2.0 ± 0.2 | 2.0 ± 0.01 | 1.3 ± 0.3 | 0.8 ± 0.2 | 2.2 ± 0.1 | 1.1 ± 0.01 | 2.1 ± 0.03 | 1.5 ± 0.2 | |
| L. thuyoides | 0.6 ± 0.3 | 1.3 ± 0.3 | 1.5 ± 0.2 | 0.7 ± 0.03 | 1.8 ± 0.4 | 0.9 ± 0.3 | 1.2 ± 0.4 | 0.5 ± 0.1 | |
| R. macrochaeta | 0.2 ± 0.1 | 0.2 ± 0.04 | 0.5 ± 0.6 | 0.6 ± 0.6 | 0.6 ± 0.2 | 0.3 ± 0.1 | 0.4 ± 0.02 | 0.5 ± 0.2 | |
| Vcmax_app (μmol m−2 s−1) | |||||||||
| C. effusa | 25.4 ± 2.52 | 17.2 ± 1.14 | 69.6 ± 5.92 | 65.5 ± 22 | 24.9 ± 4.73 | 24.6 ± 0.93 | 49.4 ± 1.28 | 45.4 ± 0.37 | p < 0.01 |
| E. pycnophylla | 20.7 ± 1.08 | 33.3 ± 6.37 | 65.6 ± 5.43 | 52.8 ± 0.87 | 42.2 ± 3.88 | 39.7 ± 8.3 | 47.3 ± 2.6 | 43.2 ± 2.85 | |
| G. magellanica | 31.0 ± 1.25 | 24.3 ± 0.35 | 84.9 ± 1.94 | 74.5 ± 0.49 | 41.6 ± 7.48 | 30.8 ± 8 | 40.1 ± 1.59 | 36.9 ± 1.67 | |
| H. laricifolium | 22.6 ± 7.23 | 44.4 ± 7.02 | 65.0 ± 1.97 | 43.8 ± 0.69 | 39.7 ± 6.81 | 35.3± 5.53 | 40.1 ± 6.9 | 31.6 ± 0.39 | |
| L. thuyoides | 28.0 ± 0.06 | 23.2 ± 2.62 | 48.6 ± 0.61 | 34.4 ± 0.75 | 42.8 ± 1.49 | 24.8 ± 1.41 | 34.3 ± 0.42 | 31.7 ± 1.75 | |
| R. macrochaeta | 26.7 ± 1.37 | 28.6 ± 1.46 | 63.1 ± 2.43 | 77.3 ± 3.5 | 47.6 ± 6.08 | 30.1 ± 7.2 | 51.4 ± 5.78 | 34.3 ± 2.9 | |
| Intervened Area | |||||||||
| Dec-20 | Apr-21 | Aug-21 | Jan-22 | ||||||
| Amax (μmol m−2 s−1) | OTC | Control | OTC | Control | OTC | Control | OTC | Control | Treatment significance |
| C. effusa | 6.2 ± 0.1 | 6 ± 0.1 | 4 ± 0.1 | 3.8 ± 0.1 | 9.5 ± 0.8 | 10 ± 0.1 | 8.5 ± 1.8 | 8.6 ± 0.01 | p > 0.05 |
| E. pycnophylla | 7.3 ± 0.01 | 7.2 ± 0.04 | 5.7 ± 0.1 | 5.6 ± 0.1 | 8.9 ± 1.1 | 5.4 ± 1.8 | 11.1 ± 1 | 12.5 ± 0.6 | |
| G. magellanica | 5.9 ± 0.01 | 5.8 ± 0.03 | 3.5 ± 0.1 | 3.1 ± 0.2 | 6.7 ± 0.3 | 4 ± 0.2 | 6.5 ± 0.03 | 6.9 ± 0.3 | |
| H. laricifolium | 6.9 ± 0.01 | 6.6 ± 0.02 | 5.4 ± 0.1 | 5.1 ± 0.1 | 9.4 ± 0.02 | 8.3 ± 0.5 | 12.9 ± 0.1 | 12.8 ± 0.2 | |
| L. thuyoides | 6.5 ± 0.01 | 6.5 ± 0.02 | 5 ± 0.1 | 4.7 ± 0.02 | 5.7 ± 0.02 | 5.4 ± 0.2 | 6.3 ± 0.2 | 6.2 ± 0.4 | |
| R. macrochaeta | 6.4 ± 0.01 | 6.3 ± 0.04 | 4.6 ± 0.1 | 4.4 ± 0.1 | 4.5 ± 0.4 | 7.7 ± 2.6 | 7.7 ± 0.5 | 9.3 ± 1.5 | |
| Rd (μmol CO2 m−2 s−1) | |||||||||
| C. effusa | 0.8 ± 0.1 | 0.4 ± 0.2 | 0.5 ± 0.1 | 0.8 ± 0.1 | 0.8 ± 0.1 | 0.4 ± 0.1 | 0.4 ± 0.2 | 0.7 ± 0.1 | p > 0.05 |
| E. pycnophylla | 0.7 ± 0.03 | 0.9 ± 0.1 | 0.6 ± 0.2 | 0.6 ± 0.2 | 0.8 ± 0.1 | 0.8 ± 0.1 | 0.6 ± 0.1 | 0.7 ± 0 | |
| G. magellanica | 0.8 ± 0.1 | 1 ± 0.1 | 0.9 ± 0.1 | 0.9 ± 0.1 | 0.9 ± 0.2 | 0.8 ± 0.1 | 0.7 ± 0.1 | 0.9 ± 0.02 | |
| H. laricifolium | 1.4 ± 0.1 | 1.7 ± 0.04 | 1.5 ± 0.3 | 1.4 ± 0.01 | 1.2 ± 0.2 | 1.3 ± 0.1 | 1.5 ± 0.02 | 1.5 ± 0.1 | |
| L. thuyoides | 1 ± 0.1 | 1 ± 0.1 | 0.4 ± 0 | 1.3 ± 0.1 | 1.1 ± 0.1 | 1.1 ± 0.2 | 0.8 ± 0.2 | 1.2 ± 0.1 | |
| R. macrochaeta | 0.4 ± 0.02 | 0.6 ± 0.02 | 0.7 ± 0.2 | 0.6 ± 0.1 | 0.6 ± 0.1 | 0.7 ± 0.1 | 0.6 ± 0.04 | 0.5 ± 0.03 | |
| Vcmax_app(μmol m−2 s−1) | |||||||||
| C. effusa | 43.65 ± 1.75 | 43.9 ± 0.04 | 38.05 ± 6.45 | 38.9 ± 8.55 | 26.83 ± 1.79 | 25.34 ± 0.8 | 22.64 ± 1.74 | 22.93 ± 0.01 | p > 0.05 |
| E. pycnophylla | 12.05 ± 0.65 | 40.3 ± 6 | 41.25 ± 5.95 | 30.5 ± 0.2 | 25.4 ± 1.16 | 22.03 ± 1.5 | 27.69 ± 2.89 | 28.71 ± 0.39 | |
| G. magellanica | 25.17 ± 9.2 | 30.95 ± 0.55 | 55.25 ± 4.15 | 35.8 ± 3.7 | 21.41 ± 0.42 | 12.01 ± 0.4 | 27.12 ± 1.83 | 24.07 ± 0.5 | |
| H. laricifolium | 23.7 ± 0.7 | 71.35 ± 0.15 | 48.65 ± 6.95 | 48.9 ± 0.9 | 28.4 ± 4.18 | 21.86 ± 0.65 | 33.01 ± 1.16 | 31.63 ± 3.17 | |
| L. thuyoides | 15.35 ± 1.25 | 16.9 ± 2.1 | 19.65 ± 4.25 | 36.6 ± 19.93 | 16.64 ± 0.86 | 16.11 ± 1.76 | 25.98 ± 1.75 | 26.74 ± 2.05 | |
| R. macrochaeta | 37.4 ± 6 | 66.6 ± 9 | 20.7 ± 4.8 | 20.25 ± 2.65 | 17.64 ± 1.98 | 21.11 ± 3.94 | 21.07 ± 1.76 | 21.84 ± 1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solarte, M.E.; Solarte Erazo, Y.; Ramírez Cupacán, E.; Enríquez Paz, C.; Melgarejo, L.M.; Lasso, E.; Flexas, J.; Gulias, J. Photosynthetic Traits of Páramo Plants Subjected to Short-Term Warming in OTC Chambers. Plants 2022, 11, 3110. https://doi.org/10.3390/plants11223110
Solarte ME, Solarte Erazo Y, Ramírez Cupacán E, Enríquez Paz C, Melgarejo LM, Lasso E, Flexas J, Gulias J. Photosynthetic Traits of Páramo Plants Subjected to Short-Term Warming in OTC Chambers. Plants. 2022; 11(22):3110. https://doi.org/10.3390/plants11223110
Chicago/Turabian StyleSolarte, María Elena, Yisela Solarte Erazo, Elizabeth Ramírez Cupacán, Camila Enríquez Paz, Luz Marina Melgarejo, Eloisa Lasso, Jaume Flexas, and Javier Gulias. 2022. "Photosynthetic Traits of Páramo Plants Subjected to Short-Term Warming in OTC Chambers" Plants 11, no. 22: 3110. https://doi.org/10.3390/plants11223110
APA StyleSolarte, M. E., Solarte Erazo, Y., Ramírez Cupacán, E., Enríquez Paz, C., Melgarejo, L. M., Lasso, E., Flexas, J., & Gulias, J. (2022). Photosynthetic Traits of Páramo Plants Subjected to Short-Term Warming in OTC Chambers. Plants, 11(22), 3110. https://doi.org/10.3390/plants11223110