Abstract
The periderm is a corky tissue that replaces the epidermis when the latter is damaged, and is critical for preventing pathogen invasion and water loss. The periderm is formed through the meristematic activity of phellogen cells (cork cambium). The potato skin (phellem cells) composes the outer layers of the tuber periderm and is a model for studying cork development. Early in tuber development and following tuber expansion, the phellogen becomes active and produces the skin. New skin layers are continuously added by division of the phellogen cells until tuber maturation. Some physiological disorders of the potato tuber are related to abnormal development of the skin, including skinning injuries and russeting of smooth-skinned potatoes. Thus, characterizing the potato periderm contributes to modeling cork development in plants and helps to resolve critical agricultural problems. Here, we summarize the data available on potato periderm formation, highlighting tissue characteristics rather than the suberization processes.
1. The Plant Periderm
The periderm, or cork, is a protective tissue of secondary origin. Its defensive characteristics are mainly due to the deposition of suberin polyester on the walls of its outer cell layers. The suberin consists of aromatic and aliphatic domains crosslinked by glycerol, and is localized between the primary wall and the plasmalemma [1]. The suberized cells are filled with air and, therefore, provide thermal insulation, the suberized walls prevent invasion by microorganisms (mechanically and chemically), and wax deposits that are embedded in the suberin material prevent desiccation of internal tissues [1,2].
The periderm occurs in several plants and plant organs: in the skin of underground organs such as potato and sweet potato; the cork of woody species due to increased thickness via secondary growth; the lenticels; the wound-healing periderm [3]; the russeting of fruit peels of, e.g., apple, pear, and kiwifruit; the netted rind of reticulated fruit [4,5,6,7], etc. A review on periderm evolution and ontogenesis has been recently published [8].
The periderm is made up of three types of cells [9]. The outer layers are composed of suberized phellem cells. The inner layers, the phelloderm, are made up of parenchyma-like cells. Between the phellem and the phelloderm is a meristematic layer of secondary origin called the phellogen or cork cambium. Outward cell divisions of the phellogen form the phellem and inward divisions form the phelloderm.
2. Native Periderm of Potato
Following potato tuberization, the original epidermis is replaced by periderm tissue, with its outer suberized phellem cells comprising the tuber skin. The development of the potato periderm can be divided into three stages [10] (Figure 1): (1) periderm initiation—cambial phellogen is formed through dedifferentiation of subepidermal cells, and sequentially, skin formation is initiated; (2) immature periderm development—active phellogen adds more skin layers to the expanding tuber; the dividing phellogen is labile and prone to fracture, resulting in separation of the skin from the underlying tuber flesh, and resulting in the costly agricultural problem of skinning injuries [11,12]; (3) periderm maturation—when the tuber ceases to expand at the end of the growing period, no new skin cells are required and the phellogen becomes inactive. As a result, skin layers adhere strongly to the tuber flesh in a process known as skin set.
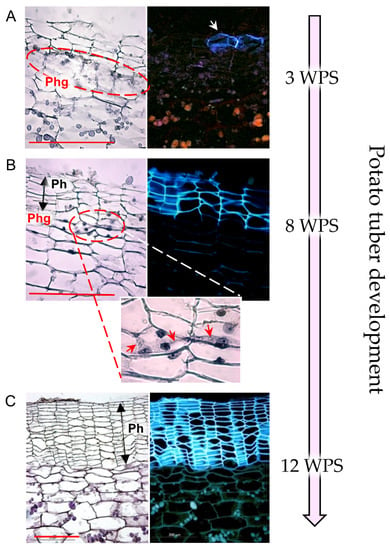
Figure 1.
Developmental stages of potato native periderm. Potato cv. Desirée plants were grown in pots (20 L) filled with perlite in a greenhouse under natural Israeli winter conditions (average temperature range of 10–18 °C). Tubers were collected at three time points during tuber development—3, 8, and 12 weeks post-sprouting (WPS) of seed tubers. Tissue samples were taken from their surface, fixed in FAA, dehydrated in an ethanol-xylene series, and embedded in paraplast. Cross-sections were stained with hematoxylin and viewed under a light microscope (left panel) and a UV microscope (right panel, black background), to examine tissue morphology and cell nuclei and the autofluorescence of suberized cell walls, respectively. (A) Periderm initiation—subepidermal cells undergo dedifferentiation to form phellogen (Phg) initials (encircled) that sequentially produce the phellem cells (white arrow). (B) Immature periderm development—the phellogen remains active and adds more phellem (Ph) cells to the expanding tuber. The close-up image (X2.5 magnification) displays the dividing phellogen cells, showing cell plates between daughter cells (red arrows). The cell plate is prone to fracture, resulting in separation of the immature skin from the tuber surface. (C) Periderm maturation—following foliage removal or plant senescence, tuber expansion ceases, the phellogen cells stop dividing, and the skin-set process is induced. The phellogen layer is undetectable at the maturation stage. Note the characteristic morphology of the phellem: columns of flattened cells that autofluoresce under UV light. Scale bars: 200 µm. Images were taken from [13].
In addition to the protective characteristic of the suberized phellem, the immature skin of potato is enriched with proteins involved in defense responses to biotic and abiotic stresses [13]. Similarly, the parenchyma-like phelloderm is enriched with defensive secondary metabolites, such as anthocyanins and the toxic steroidal glycoalkaloids [14].
A study comparing potato skin and tuber-flesh transcriptomes, followed by functional analysis of genes that were highly and differentially expressed in the skin, demonstrated genes involved in developmental processes such as cell division, cell differentiation, morphogenesis, secondary cell wall formation (lignification and suberization), and stress-related activities [15,16]. Among these genes, several transcription regulators were identified that may play roles in skin initiation and maturation [15]. These included the homeobox-leucine zipper protein, StHAT3-like gene (Sotub01g029660), which is involved in controlling embryonic apical patterning and meristem function [17]; the NUCLEAR TRANSCRIPTION FACTOR Y SUBUNIT B-3, StHAP3 (Sotub06g007420), which regulates embryo identity and development [18]; the MYB family REGULATOR OF AXILLARY MERISTEMS 2, StRAX2-like gene (Sotub09g006290), which controls a very early step in axillary meristem initiation [19]; the NAC (NAM/ATAF1/2/CUC)-domain protein, StNAC83 (Sotub07g026930), which regulates shoot apical meristem formation [20]; the RELATED TO APETALA AP2-LIKE, StRAP2-7-like gene (Sotub11g029540), an ethylene-responsive transcription factor that is involved in the establishment of floral meristem identity [21]; NO APICAL MERISTEM 4-LIKE, NAM4 (Sotub05g028670), and the transcription factor TCP (Sotub09g007560), which regulate the expression of flavonoid/anthocyanin-biosynthesis genes in response to the stress of intense light [22,23] and may be related to the accumulation of pigment in the red skin of cv. Desirée tubers.
3. Wound Periderm of Potato
Potato tuber skin is often damaged during mechanical harvest or due to an incomplete skin set. These skinning injuries are healed by the development of a wound periderm. Native and wound periderms are similar in terms of tissue origin, structure, and morphology, but differ in their maturation process and in the composition of unesterified pectin [12] and anthocyanin (in colored cultivars [24]) (Figure 2A). In addition, the suberin of the wound periderm is enriched in wax alkyl ferulates and is more permeable to water [25]. Recently, regulators of the wound-suberization process in tubers were identified—StMYB74 and StMYB102, and polymorphisms in the latter were suggested to influence cultivar-specific wound-suberization capacity [26]. In potato, periderm development can be studied by inducing the formation of a wound periderm via removal of the tuber native skin or slicing the tuber flesh, and allowing the exposed tissue to heal in the dark. Within 1–3 days, wounding induces the formation of a closing layer, in which the walls of the exposed tuber parenchyma cells undergo lignification/suberization [10]. On day 3, phellogen initials become noticeable and columns of new phellem cells can be clearly seen below the closing layer [10,27,28,29]. From day 4 on, the newly formed phellem undergoes suberization from the outside layers inward, and on day 8, the suberized phellem layers become flattened and compacted, indicating maturation of the wound periderm [10].
A transient increase in auxin and lipid hydroxyperoxide levels at 20 to 30 min after wounding was suggested to initiate the cytological events that lead to wound periderm formation [30]. A mitotic activity of cells is followed to produce the periderm, starting 120 min post-wounding [30], parallel to the expression of cell-cycle genes [29], and an increase in polymerized actin and microfilament bundles in cells at the wounding site [31]. Further studies on hormonal regulation of wound periderm formation showed that abscisic acid, ethylene, and jasmonic acid levels increase transiently shortly after wounding and prior to periderm initiation [32,33,34,35].

Figure 2.
(A) Injuries of tuber surface healed by wound periderm that is devoid of the red pigmentation (red arrows). (B) Physiological russeting of smooth skin cultivar exhibiting dark brown patches of protruding rough skin tissue (right), compared to healthy-looking smooth skin tubers (left). Images were taken from [24] and [36], respectively.
Postharvest conditions of wounded tubers influence their capacity to heal. The effect of temperature, oxygen (O2) concentration, and relative humidity (RH) has been extensively studied in the context of the agricultural practice of curing skinning injuries of newly harvested potato tubers [27]. Wound-induced periderm formation occurred most rapidly at around 20–25 °C, was delayed at lower temperatures (10–15 °C), and was inhibited at temperatures as high as 35 °C [37,38]. The combination of O2 and temperature indicated inhibition at an O2 concentration of 1% or lower when stored at 15 °C or above [39]. In general, different combinations of temperature, O2, and RH should be adjusted according to the physiological state of the tubers [40] and local practices.
In this context, the timing of wound periderm initiation and development is critical for sealing the exposed internal tissues, to prevent pathogen invasion and water loss. Recently, brassinosteroid was shown to accelerate wound healing [41]. This steroidal hormone plays a positive role in the accumulation of lignin and polyphenolic suberin at the wound, reducing tuber weight loss and increasing disease resistance. The effect of brassinosteroid is achieved via upregulation of PHENYLALANINE AMMONIA LYASE (PAL), 4-COUMALATE:COA LIGASE (4-CL), and CINNAMYL ALCOHOL DEHYDROGENASE (CAD) expression and related enzyme activities in phenylpropanoid metabolism, promoting the synthesis of lignin precursors and phenolic acids, mainly caffeic acid, sinapic acid, and cinnamyl alcohol [41]. In addition, increased production of O2− and H2O2 following induction of NADPH oxidase promoted oxidative crosslinking of the phenolic acids and lignin precursors to form the polyphenolic domain of suberin [41].
4. Potato Skin Russeting
Impaired skin development that results in russeting of smooth-skinned varieties (Figure 2B) is often associated with suboptimal growth conditions [42]. This physiological skin disorder is not caused by pathogens [36]. Russeting may be a genetic trait, such as in the well-known US variety Russet Burbank where it is a desired character [43].
Tubers with russet skin have a thicker layer of phellem than smooth-skinned potatoes [36,42,44,45]. The buildup of phellem cell layers can result from increased activity of the phellogen as a result of, for example, high soil temperature [10,42] or strong adhesion between neighboring phellem cells so that they are not sloughed off during tuber development [36]. This may be due to increased suberization [45] or increased levels of pectin and hemicellulose [44]. Nevertheless, as the tuber expands during development, the thick skin cracks, resulting in netted or russet skin. We do not know if or how the undesirable phenomenon of physiological russeting is related to the russeting trait. That is, are these two different russeting phenomena determined by the same set of genes that interact with each other and with the environment to give different roughness outcomes, or are they determined by two different sets of genes? The roughness of Russet Burbank skin is dependent on growth temperature [45] and the quantity and source of fertilizer [46], suggesting that genetic and physiological russeting may involve the same set of genes.
5. Chemical Composition of the Periderm
5.1. Suberin
Our knowledge of the biosynthesis pathway and assembly of suberin macromolecules is not complete. However, recent studies have provided new information on genes involved in suberin regulation. These are detailed in two reviews included in the present Special Issue [47,48], and are briefly discussed here in the context of potato.
The aromatic domain of potato suberin is composed of monolignols and hydroxycinnamic (ferulic) acids, and is covalently bound to the primary cell wall [49,50]. The aliphatic domain is a polyester that, upon transesterification, releases mainly C16–C28 α,ω-diacids and ω-hydroxyacids, with minor amounts of alkan-1-ols, alkanoic acids, and glycerol. The latter may be involved in crosslinking between the aromatic and aliphatic domains [51,52].
Several enzymatic activities and associated gene families have been identified in suberin biosynthesis. β-Ketoacyl CoA synthases (KCSs) are involved in fatty-acid elongation, and in potato, StKCS6 catalyzes suberin and wax compounds with chain lengths of C28 and longer [53]. Cytochrome P450 (CYP) oxygenases play a crucial role in the end- and mid-chain oxidation steps, starting from monofunctional fatty acids and leading to the ω-hydroxyacids and α,ω-diacids [54]. In potato, CYP86A33 was shown to promote the ω-hydroxylation step, and its silencing led to a periderm with a 60% decrease in its aliphatic suberin load, mainly due to reduced levels of C18:1 ω-hydroxyacid and α,ω-diacid monomers (by about 70% and 90%, respectively) compared to the wild type, and increased periderm permeability [55]. Acyltransferases promote the esterification of suberin acids to glycerol and the esterification of ω-hydroxyacids to ferulic acid [56], and the potato glycerol phosphate acyltransferase-encoding GPAT3 was found to be highly and differentially expressed at the phellogen-initiation stage [10].
The incorporation of ferulate into potato suberin is catalyzed by feruloyl transferase (FHT; [57]). It has been suggested that the product ω-hydroxyacid ferulate may be the preferred structure for linking the suberin aliphatic polyester to the neighboring aromatics [51]. Phenylpropanoid precursors and the activity of anionic peroxidase, which favored feruloyl substrates, may be involved in the synthesis of the aromatic domain [58,59].
Suberin also contains fatty-acid derivatives such as C18–C22 primary fatty alcohols that are catalyzed by fatty acyl-CoA reductases (FARs) [60]. In potato skin, FAR3 was highly expressed in the developmental stages of primary phellem formation and was observed in the inner layers of the skin phellem; it was downregulated upon skin maturation [15].
Finally, ATP-binding cassette G-transporters (ABCGs) were shown to be required for the synthesis of an effective suberin barrier in roots [61], and ABCG6 and ABCG19 were found to be expressed in the potato phellogen transcriptome [10].
In addition, two orthologs of a protein family that mediates Casparian strip formation—CASP—in the suberized endodermis of Arabidopsis roots, StCASP1B2 and StCASP1, were identified in potato skin, along with StCYP86A22 and StPOD72, whose sequences suggest that they may be closely related to known suberin-related genes [15].
5.2. Other Chemical Components
In addition to the insoluble protective suberin polymer that is embedded in the phellem cell walls, the potato tuber periderm can be a source of protective chemicals, such as antioxidant, antibacterial, and insecticidal compounds [62,63]. These molecules can be biosynthetic intermediates of the suberin polyester, or independent defense metabolites.
Nonpolar metabolites include nonpolar waxes, saturated and unsaturated fatty acids, saturated dicarboxylic acids, monoacylglycerols, 1-alkanols, n-alkanes, sterols, and polyphenolics [64]. Polar metabolites include quinic acid, phenolic amines, phenolic acids, flavonoid glycoalkaloids (solanine, chaconine, leptin, solanidine, solatriose, etc.), saponins, polyamines (putrescine, spermine, and spermidine derivatives), and methylprotodioscin and protodioscin [64].
Wound periderms may contain similar metabolites. Polar extracts include polyphenolic amines (kukoamine isomers, feruloyl putrescine, and its isomer), spermidine derivatives, feruloyl tyramine, ferulic acid, and caffeoyl putrescine [63]. Additional metabolites are flavonoid glycosides (kaempferol glycosides), phenolic acids, the steroidal glycoalkaloids, hydroxycinnamoyl putrescines, chlorogenic acid, ferulic acid, iso-chlorogenic acid, caffeic acid, and coniferyl alcohol [65].
Wound healing is also associated with oxidative stress, and free radical-scavenging activities have been demonstrated in periderm extract [63]. In both native and wound periderms, metabolite composition and level may be cultivar-dependent.
6. The Phellogen—A Key Player in Periderm Formation
Despite the importance of the corky periderm in plants, there is a lack of knowledge on the mechanism of phellogen cells activity with respect to their initiation, proliferative activity, and inactivation following the completion of periderm development. The potato periderm and the cork of Quercus suber are the accepted models for studying cork development. Periderm development from the pericycle of Arabidopsis root and hypocotyl has also been described [66].
To characterize phellogen activity, potato phellogen cells were isolated by laser-capture microdissection and their transcriptome was analyzed [10]. The phellogen is a lateral meristem. Accordingly, the potato phellogen transcriptome shared more genes with the vascular cambium than with stem or root apical meristems or the flower meristem, and had 68% shared genes with the transcriptome of oak bark (phellem and phellogen) [10,67]. These included genes that determine specific aspects of the stem cell niche and radial patterning. Furthermore, several genes that were shown to be expressed in the inner layers of the potato skin, where phellem cells are newly formed, were also found to be expressed in the vascular cambium [15].
Having the potato phellogen transcriptome enabled the identification of cellular functions that play a role in its cambial activity—cell division and differentiation, histone modification, chromatin remodeling, a high level of ribosomal proteins to support the extensive translation activity, and stress-associated genes—including heat-shock proteins that have been reported to play a role in cell-cycle regulation and meristem dedifferentiation [10]. The regulation of phellogen cell-division activity in the potato tuber was suggested to involve modulation of cytokinin homeostasis [10], similar to the regulation of phellogen in Populus [68]. High levels of vesicle trafficking activity required for the formation of a cell plate between two daughter cells (Figure 1B) were also seen in the transcriptome of the phellogen. The deposition of a new cell wall is controlled by a cytoskeletal array known as the phragmoplast [69]. The potato phellogen transcriptome includes 145 genes that are putatively related to cell-plate assembly, including genes that code for actin and myosin, as well as different aspects of vesicle trafficking, the cytoskeleton, and microtubule organization.
7. Phellogen Putative Role in Physiological Tuber Skin Blemishes
The potato phellogen is agriculturally important. Several physiological disorders of potato skin including incomplete skin set, skinning injuries, skin russeting, and loss of red pigmentation in wounded periderms may be related to impaired phellogen activity [11,24,42]. Phellogen activity can be divided into three stages: (i) initiation, when skin phellem initials are formed; (ii) proliferation, in which the skin is formed continuously and is at its immature stage, (iii) inactivation, inducing the skin maturation/skin-set process (Figure 1). Characterization of the phellogen-associated genes involved in these stages may contribute to the improved management of physiological potato skin blemishes.
Preparation of the potato phellogen transcriptome enabled the identification of genes with differential and high expression levels (1) during phellogen initiation and the early stages of skin formation and (2) following phellogen inactivation and the induction of skin maturation and skin-set processes. Moreover, the unique differential expression patterns were confirmed in both native and wound periderms [10].
The initiation of potato phellogen requires the transition of somatic cells (hypodermis or tuber parenchyma cells) to pluripotent stem cells (phellogen) that can produce different types of progeny (i.e., phellem and phelloderm). During the somatic-to-meristematic transition, cells have to dedifferentiate, activate their cell-division cycle, and reorganize their physiology, metabolism, and gene-expression patterns [70]. Accordingly, specific potato phellogen-activation genes included the histones H2B (Sotub03g016600), H3 (Sotub10g009520), and H4 (Sotub11g029670); the inducer of procambial activity VASCULAR TISSUE SIZE (VAS, Sotub01g040060); the lignin/suberin-related CAFFEOYL COENZYME A O-METHYLTRANSFERASE 1 (CCoAOMT1, Sotub02g031720), GDSL LIPASE- like (CFT, Sotub01g036860), and GLYCEROL-3-PHOSPHATE SN-2-ACYLTRANSFERASE 3 (GPAT3, Sotub01g032090) [10]—with main functions involving the chromatin remodeling and cell-wall synthesis processes that are required for the establishment of a dedifferentiation state. CFT and GPAT3 may also provide precursors for the synthesis of suberin at a later stage.
The phellogen inactivation/skin maturation (skin set)-related genes included PHD-FINGER FAMILY HOMEODOMAIN PROTEIN/HAT3.1 (PHDZnP/HAT3, Sotub01g044570), ACTIN 7 (ACT7, Sotub03g020330), NON-RECOGNITION-OF-BTH 4/MEDIATOR 15 (BTH4/MED15, Sotub04g009440), PEROXIDASE 49-LIKE (POD, Sotub02g027930), PEROXISOMAL DEFECTIVE 3/COMATOSE (PED3/CTS, Sotub04g020700), PROTEIN KINASE (APK1/AtATH8, Sotub04g014120), and ENHANCER OF AG-4 PROTEIN 2 (HGF3/HUA2, Sotub02g005440), with putative functions in the aging meristem [10].
8. Future Perspectives—Epigenetic Regulation of Potato Skin Development and Quality
Epigenetic factors such as DNA methylation and histone modifications have been shown to play a role in phellogen and phellem formation of cork oak and to have altered patterns in high- vs. low-quality phellem/cork [71,72,73,74]. Cork of low quality was characterized by the reduced meristematic activity of the phellogen, having a thinner cork layer, and the upregulation of genes belonging to the flavonoid pathway instead of promoting lignification and suberization [74]. These distinctions may not be applied to potato; however, they suggest that skin quality may be defined by phellogen characteristics. The russet skin disorder that develops on smooth-skinned potato cultivars (Section 4, above) may be viewed as a low-quality cork [36].
Potato displays strong genotype-by-environment interaction effects, and epigenetic regulation is known to govern plant responses to unfavorable growth conditions. Generally, the periderm/cork is a protective tissue that responds to stress conditions [13,45]. For example, the potato periderm was shown to react to heat stress by enhancing the production and accumulation of skin layers to create a thick and russeted protective cover [42]. Although the stress response in potato skin has not yet been shown to be epigenetically regulated, this demonstrates the potential for epigenetic modulation of skin development under unfavorable growth conditions. Agronomical practices may also induce chromatin modifications that affect skin quality [75,76]. A preliminary study determining global methylation level in potato skin indicated a significant reduction in the percentage of 5-methylcytosine (5-mC) in maturing skin compared to early immature skin, and in russet compared to smooth skin (personal communication). Similarly, a reduced concentration of 5-mC was determined in cork of low compared to high quality [73].
The potato phellogen transcriptome included orthologs of genes involved in epigenetic modifications in cork oak, including chromatin-remodeling genes, DNA methyltransferases, histone methyltransferases, and histone deacetylases [10]. Epigenetic factors were shown to be involved in lignification/suberization pathways of phellem cell walls [77] and may be regulated by the coordinated activity of histone variant H1.3 and the transcription factor MYB1 [78], which are also found in the potato phellogen transcriptome. Gene-expression studies indicated differential expression of some chromatin modifiers, higher in russeted vs. smooth skin (personal communication).
Taken together, the data support the hypothesis that epigenetic modifications are involved in regulating potato skin development and quality.
9. Conclusions
Periderm development is an interesting topic involving some notable biological processes, such as dedifferentiation of the hypodermis or parenchyma cells into “stem cells” and meristematic phellogen activity. The latter gives rise to two different cell types, each with its own differentiation pathway—the phellem cells accumulate suberin and then die, whereas the parenchyma-like phelloderm accumulates secondary defense proteins and metabolites. This makes studying periderm development a challenge, especially when every element in the formation of the tissue has implications for tuber skin/cork quality and the respective agricultural product. The formation of potato native and wound periderms has been studied for many years, focusing mainly on the suberization of the phellem cell wall, a process that gives the periderm its main protective feature. We still lack information on the regulation of phellogen cell activation to form the tuber skin, and inactivation to initiate skin maturation (skin set), the latter corresponding to skinning injuries, or russeting disorders in smooth-skinned potato cultivars, among other physiological problems. We summarized the most up-to-date knowledge on the potato periderm. Parallel studies on periderm development in trees, Arabidopsis, and russeted fruit skin will provide additional information that may be relevant for potato tuber skin.
Author Contributions
P.K. and I.G. wrote the manuscript jointly. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The research is a contribution of the Agricultural Research Organization, Volcani Institute.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kolattukudy, P.E. Biochemistry and function of cutin and suberin. Can. J. Bot 1984, 62, 2918–2933. [Google Scholar] [CrossRef]
- Kolattukudy, P.E. Lipid polymers and associated phenols, their chemistry, biosynthesis and role in pathogenesis. Recent Adv. Phytochem. 1977, 77, 185–246. [Google Scholar] [CrossRef]
- Singh, B.; Bhardwaj, V.; Kaur, K.; Kukreja, S.; Goutam, U. Potato periderm is the first layer of defence against biotic and abiotic stresses: A review. Potato Res. 2021, 64, 131–146. [Google Scholar] [CrossRef]
- Arya, G.C.; Dong, Y.; Heinig, U.; Shahaf, N.; Kazachkova, Y.; Aviv-Sharon, E.; Nomberg, G.; Marinov, O.; Manasherova, E.; Aharoni, A.; et al. The metabolic and proteomic repertoires of periderm tissue in skin of the reticulated Sikkim cucumber fruit. In Horticulture Research; Oxford University Press: Oxford, UK, 2022; Volume 9. [Google Scholar] [CrossRef]
- Fogelman, E.; Redel, G.; Doron, I.; Naor, A.; Ben-Yashar, E.; Ginzberg, I. Control of apple russeting in a warm and dry climate. J. Hortic. Sci. Biotechnol. 2009, 84, 279–284. [Google Scholar] [CrossRef]
- Macnee, N.; Hilario, E.; Tahir, J.; Currie, A.; Warren, B.; Rebstock, R.; Hallett, I.C.; Chagné, D.; Schaffer, R.J.; Bulley, S.M. Peridermal fruit skin formation in Actinidia sp. (kiwifruit) is associated with genetic loci controlling russeting and cuticle formation. BMC Plant Biol. 2021, 21, 334. [Google Scholar] [CrossRef]
- Cohen, H.; Dong, Y.; Szymanski, J.; Lashbrooke, J.; Meir, S.; Almekias-Siegl, E.; Zeisler-Diehl, V.V.; Schreiber, L.; Aharoni, A. A multilevel study of melon fruit reticulation provides insight into skin ligno-suberization hallmarks. Plant Physiol. 2019, 179, 1486–1501. [Google Scholar] [CrossRef]
- Serra, O.; Mähönen, A.P.; Hetherington, A.J.; Ragni, L. The making of plant armor: The periderm. Annu. Rev. Plant Biol. 2021, 73, 405–432. [Google Scholar] [CrossRef]
- Reeve, R.M.; Hautala, E.; Weaver, M.L. Anatomy and compositional variation within potatoes. Am. Potato J. 1969, 46, 361–373. [Google Scholar] [CrossRef]
- Vulavala, V.K.R.; Fogelman, E.; Faigenboim, A.; Shoseyov, O.; Ginzberg, I. The transcriptome of potato tuber phellogen reveals cellular functions of cork cambium and genes involved in periderm formation and maturation. Sci. Rep. 2019, 9, 10216. [Google Scholar] [CrossRef]
- Lulai, E.C.; Freeman, T.P. The importance of phellogen cells and their structural characteristics in susceptibility and resistance to excoriation in immature and mature potato tuber (Solanum tuberosum L.) periderm. Ann. Bot. 2001, 88, 555–561. [Google Scholar] [CrossRef]
- Sabba, R.P.; Lulai, E.C. Histological analysis of the maturation of native and wound periderm in potato (Solanum tuberosum L.) tuber. In Annals of Botany; Oxford University Press: Oxford, UK, 2002; Volume 90, pp. 1–10. [Google Scholar] [CrossRef]
- Barel, G.; Ginzberg, I. Potato skin proteome is enriched with plant defence components. J. Exp. Bot. 2008, 59, 3347–3357. [Google Scholar] [CrossRef]
- Fogelman, E.; Oren-Shamir, M.; Hirschberg, J.; Mandolino, G.; Parisi, B.; Ovadia, R.; Tanami, Z.; Faigenboim, A.; Ginzberg, I. Nutritional value of potato (Solanum tuberosum) in hot climates: Anthocyanins, carotenoids, and steroidal glycoalkaloids. Planta 2019, 249, 1143–1155. [Google Scholar] [CrossRef]
- Vulavala, V.K.R.; Fogelman, E.; Rozental, L.; Faigenboim, A.; Tanami, Z.; Shoseyov, O.; Ginzberg, I. Identification of genes related to skin development in potato. Plant Mol. Biol. 2017, 94, 481–494. [Google Scholar] [CrossRef]
- Soler, M.; Serra, O.; Fluch, S.; Molinas, M.; Figueras, M. A potato skin SSH library yields new candidate genes for suberin biosynthesis and periderm formation. Planta 2011, 233, 933–978. [Google Scholar] [CrossRef]
- Turchi, L.; Carabelli, M.; Ruzza, V.; Possenti, M.; Sassi, M.; Peñalosa, A.; Sessa, G.; Salvi, S.; Forte, V.; Morelli, G.; et al. Arabidopsis HD-Zip II transcription factors control apical embryo development and meristem function. Development 2013, 140, 2118–2129. [Google Scholar] [CrossRef]
- Kwong, R.W.; Bui, A.Q.; Lee, H.; Kwong, L.W.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. LEAFY COTYLEDON1-LIKE defines a class of regulators essential for embryo development. Plant Cell 2003, 15, 5–18. [Google Scholar] [CrossRef]
- Müller, D.r.; Schmitz, G.; Theres, K. Blind homologous R2R3 myb genes control the pattern of lateral meristem initiation in Arabidopsis. Plant Cell 2006, 18, 586–597. [Google Scholar] [CrossRef]
- Takada, S.; Hibara, K.; Ishida, T.; Tasaka, M. The CUP-SHAPED COTYLEDON1 gene of Arabidopsis regulates shoot apical meristem formation. Development 2001, 128, 1127–1135. [Google Scholar] [CrossRef]
- Liu, X.; Dinh, T.T.; Li, D.; Shi, B.; Li, Y.; Cao, X.; Guo, L.; Pan, Y.; Jiao, Y.; Chen, X. AUXIN RESPONSE FACTOR 3 integrates the functions of AGAMOUS and APETALA2 in floral meristem determinacy. Plant J. 2014, 80, 629–641. [Google Scholar] [CrossRef]
- Morishita, T.; Kojima, Y.; Maruta, T.; Nishizawa-Yokoi, A.; Yabuta, Y.; Shigeoka, S. Arabidopsis NAC transcription factor, ANAC078, regulates flavonoid biosynthesis under high-light. Plant Cell Physiol. 2009, 50, 2210–2222. [Google Scholar] [CrossRef]
- Viola, I.L.; Camoirano, A.; Gonzalez, D.H. Redox-dependent modulation of anthocyanin biosynthesis by the TCP transcription factor TCP15 during exposure to high light intensity conditions in Arabidopsis. Plant Physiol. 2015, 170, 74–85. [Google Scholar] [CrossRef]
- Fogelman, E.; Tanami, S.; Ginzberg, I. Anthocyanin synthesis in native and wound periderms of potato. Physiol. Plant 2014, 153, 616–626. [Google Scholar] [CrossRef]
- Schreiber, L.; Franke, R.; Hartmann, K. Wax and suberin development of native and wound periderm of potato (Solanum tuberosum L.) and its relation to peridermal transpiration. Planta 2005, 220, 520–530. [Google Scholar] [CrossRef]
- Wahrenburg, Z.; Benesch, E.; Lowe, C.; Jimenez, J.; Vulavala, V.K.R.; Lü, S.; Hammerschmidt, R.; Douches, D.; Yim, W.C.; Santos, P.; et al. Transcriptional regulation of wound suberin deposition in potato cultivars with differential wound healing capacity. Plant J. 2021, 107, 77–99. [Google Scholar] [CrossRef]
- Ginzberg, I. Wound-periderm formation. In Induced Plant Resistabce to Herbivory; Schaller, A., Ed.; Springer Publishing: New York, NY, USA, 2008; pp. 131–146. [Google Scholar]
- Lulai, E.C.; Neubauer, J.D. Wound-induced suberization genes are differentially expressed, spatially and temporally, during closing layer and wound periderm formation. Postharvest Biol. Technol. 2014, 90, 24–33. [Google Scholar] [CrossRef]
- Neubauer, J.D.; Lulai, E.C.; Thompson, A.L.; Suttle, J.C.; Bolton, M.D. Wounding coordinately induces cell wall protein, cell cycle and pectin methyl esterase genes involved in tuber closing layer and wound periderm development. J. Plant Physiol. 2012, 169, 586–595. [Google Scholar] [CrossRef]
- Fabbri, A.A.; Fanelli, C.; Reverberi, M.; Ricelli, A.; Camera, E.; Urbanelli, S.; Rossini, A.; Picardo, M.; Altamura, M.M. Early physiological and cytological events induced by wounding in potato tuber. J. Exp. Bot. 2000, 51, 1267–1275. [Google Scholar] [CrossRef]
- Morelli, J.K.; Zhou, W.; Yu, J.; Lu, C.; Vayda, M.E. Actin depolymerization affects stress-induced translational activity of potato tuber tissue. Plant Physiol. 1998, 116, 1227–1237. [Google Scholar] [CrossRef][Green Version]
- Lulai, E.; Huckle, L.; Neubauer, J.; Suttle, J. Coordinate expression of AOS genes and JA accumulation: JA is not required for initiation of closing layer in wound healing tubers. J. Plant Physiol. 2011, 168, 976–982. [Google Scholar] [CrossRef]
- Lulai, E.C.; Suttle, J.C. The involvement of ethylene in wound-induced suberization of potato tuber (Solanum tuberosum L.): A critical assessment. Postharvest Biol. Technol. 2004, 34, 105–112. [Google Scholar] [CrossRef]
- Lulai, E.C.; Suttle, J.C.; Pederson, S.M. Regulatory involvement of abscisic acid in potato tuber wound-healing. J. Exp. Bot. 2008, 59, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Lulai, E.C.; Suttle, J.C. Signals involved in tuber wound-healing. Plant Signal. Behav. 2009, 4, 620–622. [Google Scholar] [CrossRef] [PubMed]
- Ginzberg, I.; Minz, D.; Faingold, I.; Soriano, S.; Mints, M.; Fogelman, E.; Warshavsky, S.; Zig, U.; Yermiyahu, U. Calcium mitigated potato skin physiological disorder. Am. J. Potato Res. 2012, 89, 351–362. [Google Scholar] [CrossRef]
- Morris, S.C.; Forbes-Smith, M.R.; Scriven, F.M. Determination of optimum conditions for suberization, wound periderm formation, cellular desiccation and pathogen resistance in wounded Solanum tuberosum tubers. Physiol. Mol. Plant Pathol. 1989, 35, 177–190. [Google Scholar] [CrossRef]
- Thomas, P. Wound-induced suberization and periderm development in potato tubers as affected by temperature and gamma irradiation. Potato Res. 1982, 25, 155–164. [Google Scholar] [CrossRef]
- Lipton, W.J. Some effects of low-oxygen atmospheres on potato tubers. Am. Potato J. 1967, 44, 292–299. [Google Scholar] [CrossRef]
- Wigginton, M. Effects of temperature, oxygen tension and relative humidity on the wound-healing process in the potato tuber. Potato Res. 1974, 17, 200–214. [Google Scholar] [CrossRef]
- Han, Y.; Yang, R.; Zhang, X.; Wang, Q.; Wang, B.; Zheng, X.; Li, Y.; Prusky, D.; Bi, Y. Brassinosteroid accelerates wound healing of potato tubers by activation of reactive oxygen metabolism and phenylpropanoid metabolism. Foods 2022, 11, 906. [Google Scholar] [CrossRef]
- Ginzberg, I.; Barel, G.; Ophir, R.; Tzin, E.; Tanami, Z.; Muddarangappa, T.; De Jong, W.; Fogelman, E. Transcriptomic profiling of heat-stress response in potato periderm. J. Exp. Bot. 2009, 60, 4411–4421. [Google Scholar] [CrossRef]
- De Jong, H. Inheritance of russeting in cultivated diploid potatoes. Potato Res. 1981, 24, 309–313. [Google Scholar] [CrossRef]
- Okazawa, Y.; Iriuda, N. On occurrence of defected potato tubers with rough russeted skin. Jpn. J. Crop Sci. 1980, 49, 58–65. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Timm, H.; Spurr, A.R. Effects of soil temperature on growth and nutrition of potato plants and tuberization, composition, and periderm structure of tubers. Proc. Am. Soc. Hortic. Sci. 1964, 84, 412–423. [Google Scholar]
- Murphy, H.; Goven, M. Influence of source of potash on yield, specific gravity, and surface russeting of the Russet Burbank variety in Maine. Am. Potato J. 1965, 42, 192–194. [Google Scholar] [CrossRef]
- Woolfson, K.N.; Esfandiari, M.; Bernards, M.A. Suberin biosynthesis, assembly, and regulation. Plants 2022, 11, 555. [Google Scholar] [CrossRef]
- Nomberg, G.; Marinov, O.; Arya, G.C.; Manasherova, E.; Cohen, H. The key enzymes in the suberin biosynthetic pathway in plants: An update. Plants 2022, 11, 392. [Google Scholar] [CrossRef]
- Bernards, M.A.; Lewis, N.G. The macromolecular aromatic domain in suberized tissue: A changing paradigm. Phytochemistry 1998, 47, 915–933. [Google Scholar] [CrossRef]
- Yan, B.; Stark, R.E. Biosynthesis, molecular structure, and domain architecture of potato suberin: A C-13 NMR study using isotopically labeled precursors. J. Agric. Food Chem. 2000, 48, 3298–3304. [Google Scholar] [CrossRef]
- Graça, J. Suberin: The biopolyester at the frontier of plants. Front. Chem. 2015, 3, 62. [Google Scholar] [CrossRef]
- Graça, J.; Pereira, H. Suberin structure in potato periderm: Glycerol, long-chain monomers, and glyceryl and feruloyl dimers. J. Agric. Food Chem. 2000, 48, 5476–5483. [Google Scholar] [CrossRef]
- Serra, O.; Soler, M.; Hohn, C.; Franke, R.; Schreiber, L.; Prat, S.; Molinas, M.; Figueras, M. Silencing of StKCS6 in potato periderm leads to reduced chain lengths of suberin and wax compounds and increased peridermal transpiration. J. Exp. Bot. 2009, 60, 697–707. [Google Scholar] [CrossRef]
- Kolattukudy, P.E. Polyestrs in higher plants. Adv. Biochem. Eng./Biotechnol. 2001, 71, 2–49. [Google Scholar]
- Serra, O.; Soler, M.; Hohn, C.; Sauveplane, V.; Pinot, F.; Franke, R.; Schreiber, L.; Prat, S.; Molinas, M.; Figueras, M. CYP86A33—targeted gene silencing in potato tuber alters suberin composition, distorts suberin lamellae, and impairs the periderm’s water barrier function. Plant Physiol. 2009, 149, 1050–1060. [Google Scholar] [CrossRef]
- Beisson, F.; Li, Y.; Bonaventure, G.; Pollard, M.; Ohlrogge, J.B. The acyltransferase GPAT5 is required for the synthesis of suberin in seed coat and root of Arabidopsis. Plant Cell 2007, 19, 351–368. [Google Scholar] [CrossRef]
- Serra, O.; Hohn, C.; Franke, R.; Prat, S.; Molinas, M.; Figueras, M. A feruloyl transferase involved in the biosynthesis of suberin and suberin-associated wax is required for maturation and sealing properties of potato periderm. Plant J. 2010, 62, 277–290. [Google Scholar] [CrossRef]
- Bernards, M.A. Demystifying suberin. Can. J. Bot.-Rev. Can. De Bot. 2002, 80, 227–240. [Google Scholar] [CrossRef]
- Bernards, M.A.; Fleming, W.D.; Llewellyn, D.B.; Priefer, R.; Yang, X.L.; Sabatino, A.; Plourde, G.L. Biochemical characterization of the suberization-associated anionic peroxidase of potato. Plant Physiol. 1999, 121, 135–145. [Google Scholar] [CrossRef]
- Domergue, F.; Vishwanath, S.J.; Joubes, J.; Ono, J.; Lee, J.A.; Bourdon, M.; Alhattab, R.; Lowe, C.; Pascal, S.; Lessire, R.; et al. Three Arabidopsis fatty acyl-coenzyme A reductases, FAR1, FAR4, and FAR5, generate primary fatty alcohols associated with suberin deposition. Plant Physiol. 2010, 153, 1539–1554. [Google Scholar] [CrossRef]
- Landgraf, R.; Smolka, U.; Altmann, S.; Eschen-Lippold, L.; Senning, M.; Sonnewald, S.; Weigel, B.; Frolova, N.; Strehmel, N.; Hause, G.; et al. The ABC transporter ABCG1 is required for suberin formation in potato tuber periderm. Plant Cell 2014, 26, 3403–3415. [Google Scholar] [CrossRef]
- Dastmalchi, K.; Perez Rodriguez, M.; Lin, J.; Yoo, B.; Stark, R.E. Temporal resistance of potato tubers: Antibacterial assays and metabolite profiling of wound-healing tissue extracts from contrasting cultivars. Phytochemistry 2019, 159, 75–89. [Google Scholar] [CrossRef]
- Dastmalchi, K.; Cai, Q.; Zhou, K.; Huang, W.; Serra, O.; Stark, R.E. Solving the jigsaw puzzle of wound-healing potato cultivars: Metabolite profiling and antioxidant activity of polar extracts. J. Agric. Food Chem. 2014, 62, 7963–7975. [Google Scholar] [CrossRef]
- Huang, W.; Serra, O.; Dastmalchi, K.; Jin, L.; Yang, L.; Stark, R.E. Comprehensive MS and solid-state NMR metabolomic profiling reveals molecular variations in native periderms from four Solanum tuberosum potato cultivars. J. Agric. Food Chem. 2017, 65, 2258–2274. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.-L.; Bernards, M. Metabolite profiling of potato (Solanum tuberosum L.) tubers during wound-induced suberization. Metabolomics 2007, 3, 147–159. [Google Scholar] [CrossRef]
- Wunderling, A.; Ripper, D.; Barra-Jimenez, A.; Mahn, S.; Sajak, K.; Targem, M.B.; Ragni, L. A molecular framework to study periderm formation in Arabidopsis. New Phytol. 2018, 219, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Boher, P.; Soler, M.; Sánchez, A.; Hoede, C.; Noirot, C.; Paiva, J.A.P.; Serra, O.; Figueras, M. A comparative transcriptomic approach to understanding the formation of cork. Plant Mol. Biol. 2018, 96, 103–118. [Google Scholar] [CrossRef]
- Miguel, A.; Milhinhos, A.; Novák, O.; Jones, B.; Miguel, C.M. The SHORT-ROOT-like gene PtSHR2B is involved in Populus phellogen activity. J. Exp. Bot. 2016, 67, 1545–1555. [Google Scholar] [CrossRef]
- Smertenko, A.; Assaad, F.; Baluška, F.; Bezanilla, M.; Buschmann, H.; Drakakaki, G.; Hauser, M.-T.; Janson, M.; Mineyuki, Y.; Moore, I.; et al. Plant cytokinesis: Terminology for structures and processes. Trends Cell Biol. 2017, 27, 885–894. [Google Scholar] [CrossRef]
- Fehér, A.; Pasternak, T.P.; Dudits, D. Transition of somatic plant cells to an embryogenic state. Plant Cell Tissue Organ Cult. 2003, 74, 201–228. [Google Scholar] [CrossRef]
- Inácio, V.; Barros, P.M.; Costa, A.; Roussado, C.; Gonçalves, E.; Costa, R.; Graça, J.; Oliveira, M.M.; Morais-Cecílio, L. Differential DNA methylation patterns are related to phellogen origin and quality of Quercus suber cork. PLoS ONE 2017, 12, e0169018. [Google Scholar] [CrossRef]
- Inácio, V.; Martins, M.T.; Graça, J.; Morais-Cecílio, L. Cork oak young and traumatic periderms show PCD typical chromatin patterns but different chromatin-modifying genes expression. Front. Plant Sci. 2018, 9, 1194. [Google Scholar] [CrossRef]
- Ramos, M.; Rocheta, M.; Carvalho, L.; Inácio, V.; Graça, J.; Morais-Cecilio, L. Expression of DNA methyltransferases is involved in Quercus suber cork quality. Tree Genet. Genomes 2013, 9, 1481–1492. [Google Scholar] [CrossRef][Green Version]
- Teixeira, R.T.; Fortes, A.M.; Bai, H.; Pinheiro, C.; Pereira, H. Transcriptional profiling of cork oak phellogenic cells isolated by laser microdissection. Planta 2018, 247, 317–338. [Google Scholar] [CrossRef]
- Kooke, R.; Johannes, F.; Wardenaar, R.; Becker, F.; Etcheverry, M.; Colot, V.; Vreugdenhil, D.; Keurentjes, J.J.B. Epigenetic basis of morphological variation and phenotypic plasticity in Arabidopsis thaliana. Plant Cell 2015, 27, 337–348. [Google Scholar] [CrossRef]
- Secco, D.; Whelan, J.; Rouached, H.; Lister, R. Nutrient stress-induced chromatin changes in plants. Curr. Opin. Plant Biol. 2017, 39, 1–7. [Google Scholar] [CrossRef]
- Yu, N.; Cao, L.; Yuan, L.; Zhi, X.; Chen, Y.; Gan, S.; Chen, L. Maintenance of grafting-induced epigenetic variations in the asexual progeny of Brassica oleracea and B. juncea chimera. Plant J. 2018, 96, 22–38. [Google Scholar] [CrossRef]
- Soler, M.; Plasencia, A.; Larbat, R.; Pouzet, C.; Jauneau, A.; Rivas, S.; Pesquet, E.; Lapierre, C.; Truchet, I.; Grima-Pettenati, J. The Eucalyptus linker histone variant EgH1.3 cooperates with the transcription factor EgMYB1 to control lignin biosynthesis during wood formation. New Phytol. 2017, 213, 287–299. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).