
Identification and Characterization of Colletotrichum fructicola and Colletotrichum siamense Causing Anthracnose on Luffa Sponge Gourd in China

Abstract
:1. Introduction
2. Results
2.1. Fungal Isolation
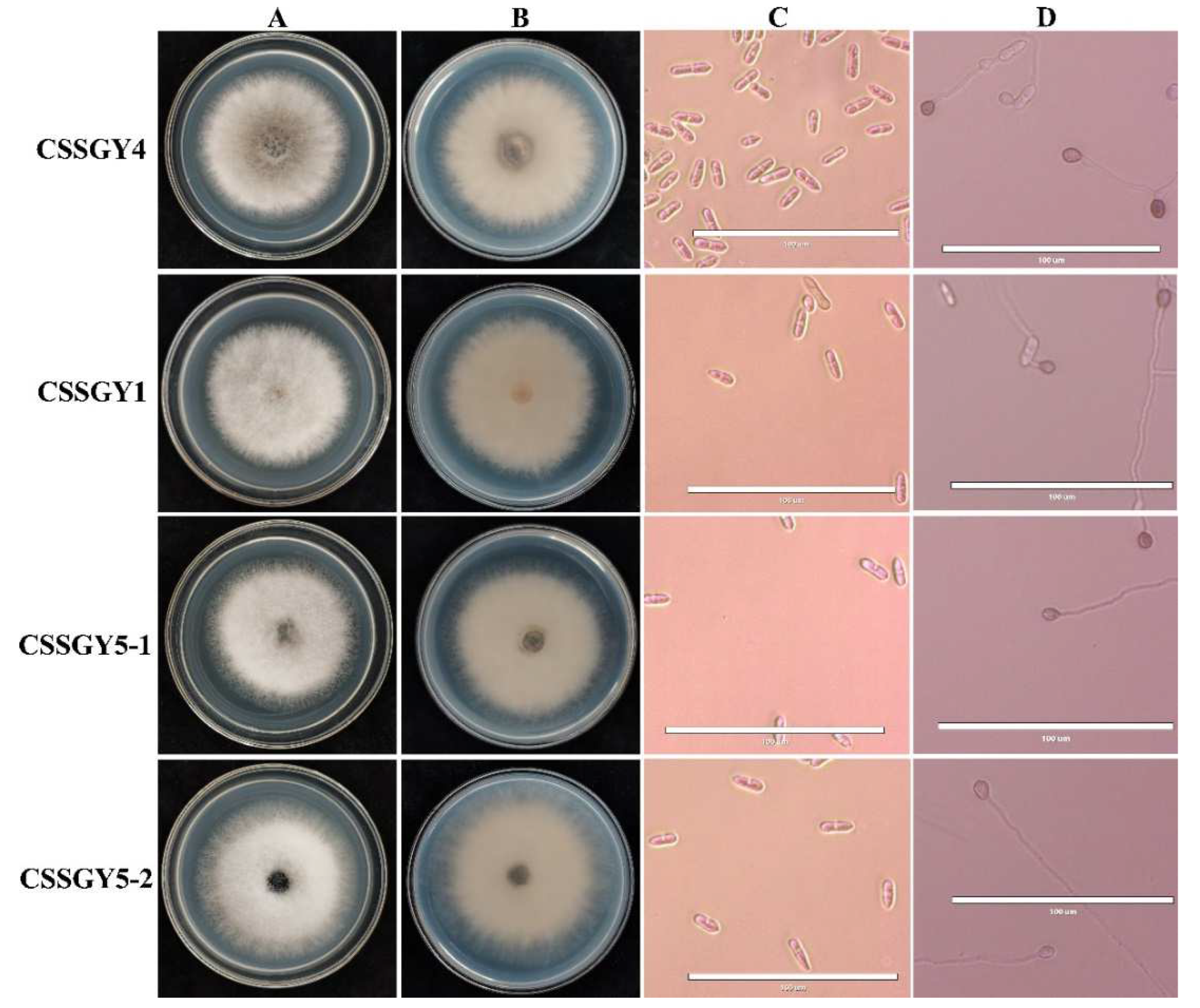
2.2. Morphological Characterization
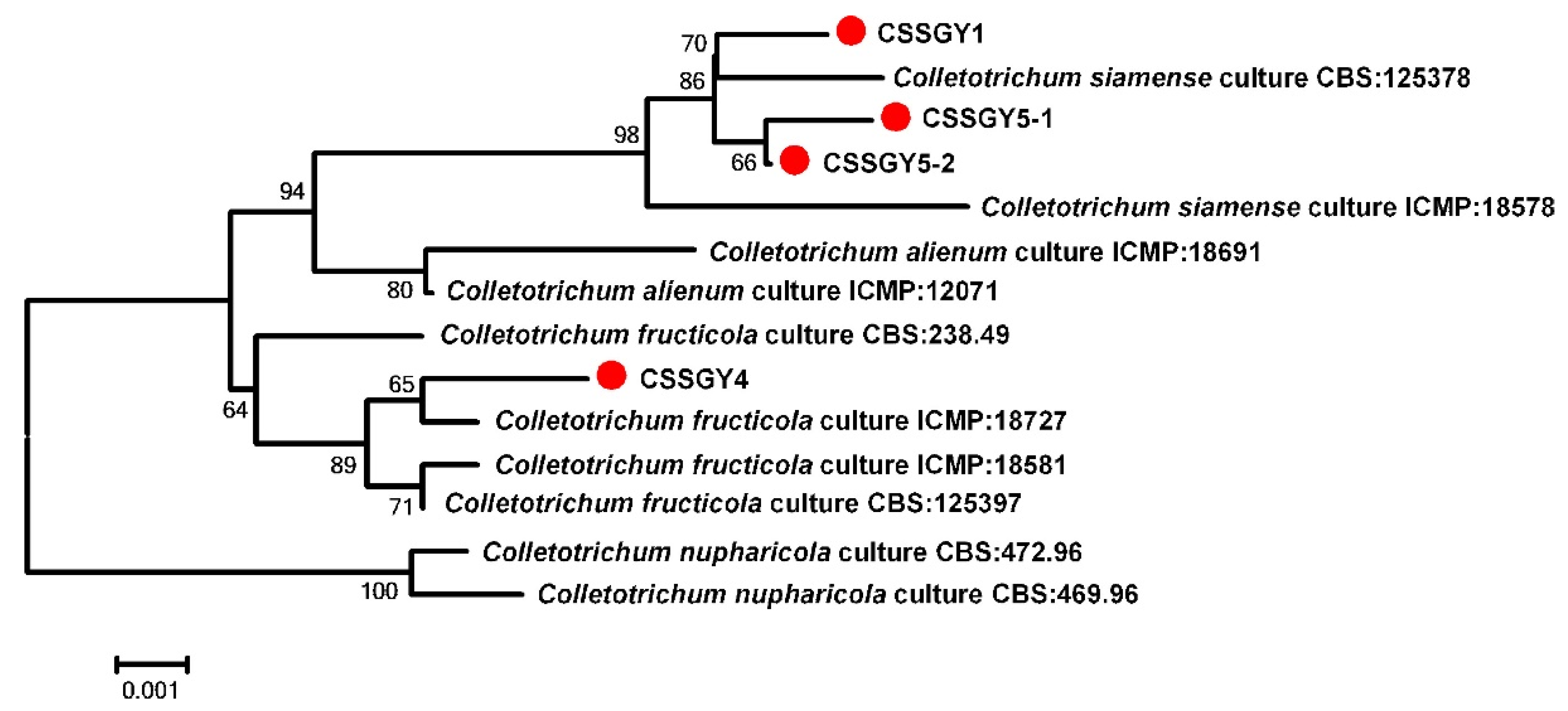
2.3. Molecular Identification
2.4. Pathogenicity Test
3. Discussion and Conclusions
4. Materials and Methods
4.1. Sample Collection and Fungal Isolation
4.2. Morphological and Cultural Characterization
4.3. Molecular Identification
4.4. Pathogenicity Assay
4.5. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, H.B.; Gong, H.; Liu, P.; He, X.L.; Luo, S.B.; Zheng, X.M.; Zhang, C.Y.; He, X.Y.; Luo, J. Large-scale development of EST-SSR markers in sponge gourd via transcriptome sequencing. Mol. Breed. 2014, 34, 1903–1915. [Google Scholar] [CrossRef]
- Wu, H.; Zhao, G.; Gong, H.; Li, J.; Luo, C.; He, X.; Luo, S.B.; Zheng, X.M.; Liu, X.X.; Guo, J.; et al. A high-quality sponge gourd (Luffa cylindrica) genome. Hortic. Res. 2020, 7, 128. [Google Scholar] [CrossRef]
- Zhang, T.; Ren, X.; Zhang, Z.; Ming, Y.; Yang, Z.; Hu, J.; Li, S.L.; Wang, Y.; Sun, S.R.; Sun, K.L.; et al. Long-read sequencing and de novo assembly of the Luffa cylindrica (L.) Roem. genome. Mol. Ecol. Res. 2020, 20, 511–519. [Google Scholar] [CrossRef]
- Cohen, Y.; Rubin, A.E.; Liu, X.L.; Wang, W.Q.; Zhang, Y.J.; Hermann, D. First report on the occurrence of A2 mating type of the cucurbit downy mildew agent Pseudoperonospora cubensis in China. Plant Dis. 2013, 97, 559. [Google Scholar] [CrossRef] [PubMed]
- Nayak, A.K.; Bandamaravuri, K.B. First report of downy mildew caused by Pseudoperonospora cubensis on Luffa cylindrica in India. J. Plant Pathol. 2019, 101, 447. [Google Scholar] [CrossRef] [Green Version]
- Ko, Y.; Yao, K.S.; Chen, C.Y.; Lin, C.H. First report of gray mold disease of sponge gourd (Luffa cylindrica) caused by Botrytis cinerea in Taiwan. Plant Dis. 2007, 91, 1199. [Google Scholar] [CrossRef]
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; Van Kan, J.A. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.-Y.; Zhang, J.-Z. Taxonomic studies of Alternaria from China VI. New species and new records to China on Cruciferae, Cucurbitaceae, and Euphorbiaceae. Mycotaxon 1999, 72, 423–432. [Google Scholar]
- Bellé, C.; Moccellin, R.; Meneses, P.R.; Neves, C.G.; Groth, M.Z.; Kaspary, T.E.; de Barros, D.R.; de Farias, C.R.J. First report of Sclerotium rolfsii causing stem rot of Luffa cylindrica in Brazil. Plant Dis. 2018, 102, 250. [Google Scholar] [CrossRef]
- Kousik, C.S.; Parada, C.; Quesada-Ocampo, L. First report of Phytophthora fruit rot on bitter gourd (Mormodica charantia) and sponge gourd (Luffa cylindrica) caused by Phytophthora capsici. Plant Health Progress 2015, 16, 93–94. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.; Freitas, N.M.; Mendonça, H.L.; Barreto, R.W. First report of Stagonosporiopsis cucurbitacearum causing fruit rot of Luffa cylindrica in Brazil. Plant Dis. 2013, 97, 1120. [Google Scholar] [CrossRef]
- Damm, U.; Woudenberg, J.H.C.; Cannon, P.F.; Crous, P.W. Colletotrichum species with curved conidia from herbaceous hosts. Fungal Divers. 2009, 39, 45. [Google Scholar]
- Zhuang, W.-Y. Higher Fungi of Tropical China; Mycotaxon, Ltd.: Ithaca, NY, USA, 2001; p. 485. [Google Scholar]
- Cai, L.; Hyde, K.D.; Taylor, P.W.J.; Weir, B.; Waller, J.; Abang, M.M.; Zhang, J.Z.; Yang, Y.L.; Phoulivong, S.; Liu, Z.Y.; et al. A polyphasic approach for studying Colletotrichum. Fungal Divers. 2009, 39, 183–204. [Google Scholar]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.R.; Yu, S.Y.; Chang, T.D.; Lin, Y.J.; Wen, C.J.; Lin, Y.H. First report of anthracnose caused by Colletotrichum fructicola on tea in Taiwan. Plant Dis. 2021, 105, 710. [Google Scholar] [CrossRef]
- Sharma, G.; Shenoy, B.D. Colletotrichum fructicola and C. siamense are involved in chilli anthracnose in India. Arch. Phytopathol. Plant Prot. 2014, 47, 1179–1194. [Google Scholar] [CrossRef]
- Wang, C.X.; Zhang, Z.F.; Li, B.H.; Wang, H.Y.; Dong, X.L. First report of Glomerella leaf spot of apple caused by Glomerella cingulata in China. Plant Dis. 2012, 96, 912. [Google Scholar] [CrossRef]
- Rockenbach, M.F.; Velho, A.C.; Gonçalves, A.E.; Mondino, P.E.; Alaniz, S.M.; Stadnik, M.J. Genetic structure of Colletotrichum fructicola associated to apple bitter rot and Glomerella leaf spot in southern Brazil and Uruguay. Phytopathology 2016, 106, 774–781. [Google Scholar] [CrossRef] [Green Version]
- Prihastuti, H.; Cai, L.; Chen, H.; McKenzie, E.H.C.; Hyde, K.D. Characterization of Colletotrichum species associated with coffee berries in northern Thailand. Fungal Divers. 2009, 39, 89–109. [Google Scholar]
- Udayanga, D.; Manamgoda, D.S.; Liu, X.; Chukeatirote, E.; Hyde, K.D. What are the common anthracnose pathogens of tropical fruits? Fungal Divers. 2013, 61, 165–179. [Google Scholar] [CrossRef]
- Liu, F.; Tang, G.; Zheng, X.; Li, Y.; Sun, X.; Qi, X.; Zhou, Y.; Xu, J.; Chen, H.; Chang, X.; et al. Molecular and phenotypic characterization of Colletotrichum species associated with anthracnose disease in peppers from Sichuan Province, China. Sci. Rep. 2016, 6, 32761. [Google Scholar] [CrossRef]
- Wang, Y.C.; Hao, X.Y.; Wang, L.; Xiao, B.; Wang, X.C.; Yang, Y.J. Diverse Colletotrichum species cause anthracnose of tea plants (Camellia sinensis (L.) O. Kuntze) in China. Sci. Rep. 2016, 6, 35287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, O.; Jeon, J.Y.; Chang, T.; Shin, J.S.; Oh, N.K.; Lee, Y.S. Molecular and morphological characterization of Colletotrichum species in the Colletotrichum gloeosporioides complex associated with persimmon anthracnose in South Korea. Plant Dis. 2018, 102, 1015–1024. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.; Gleason, M.L.; Zhang, R.; Sun, G. Genome sequence resource of the wide-host-range anthracnose pathogen Colletotrichum siamense. Mol. Plant-Microbe Interact. 2019, 32, 931–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Feng, W.; Yang, J.; Gao, Z.; Zhang, Z.; Zhang, W.; Wang, S.; Wang, W.; Gong, D.; Hu, M. First report of anthracnose caused by Colletotrichum siamense on avocado fruits in China. Crop Prot. 2022, 155, 105922. [Google Scholar] [CrossRef]
- Innis, M.A.; Gelfand, D.H.; Sninsky, J.J.; White, T.J. PCR Protocols: A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- De Hoog, G.; Van den Ended, A.H.G.G. Molecular diagnostics of clinical strains of filamentous Basidiomycetes. Mycoses 1998, 41, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Templeton, M.D.; Rikkerink, E.H.; Solon, S.L.; Crowhurst, R.N. Cloning and molecular characterization of the glyceraldehyde-3-phosphate dehydrogenase-encoding gene and cDNA from the plant pathogenic fungus Glomerella cingulata. Gene 1992, 122, 225–230. [Google Scholar] [CrossRef]
- Silva, D.N.; Talhinhas, P.; Várzea, V.; Cai, L.; Paulo, O.S.; Batista, D. Application of the Apn2/MAT locus to improve the systematics of the Colletotrichum gloeosporioides complex: An example from coffee (Coffea spp.) hosts. Mycologia 2012, 104, 396–409. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence align-ment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specimen Number | Host | Varieties | Geographic Origin (City, Province) |
|---|---|---|---|
| SGY1 | Luffa cylindrica | Zhenli | Changsha, Hunan |
| SGY2 | Luffa cylindrica | Zhenli | Changsha, Hunan |
| SGY3 | Luffa cylindrica | Zhenli | Changsha, Hunan |
| SGY4 | Luffa cylindrica | Jingpinbaili | Changsha, Hunan |
| SGY5-1 | Luffa cylindrica | Jingpinbaili | Changsha, Hunan |
| SGY5-2 | Luffa cylindrica | Jingpinbaili | Changsha, Hunan |
| SGY6 | Luffa cylindrica | Jingpinbaili | Changsha, Hunan |
| SGY7-1 | Luffa cylindrica | Changsharousigua | Changsha, Hunan |
| SGY7-2 | Luffa cylindrica | Changsharousigua | Changsha, Hunan |
| SGY8 | Luffa cylindrica | Changsharousigua | Changsha, Hunan |
| Strain Name | Conidia Size (μm) | Appressoria Size (μm) | ||
|---|---|---|---|---|
| Length Average ± SD a | Width Average ± SD a | Length Average ± SD a | Width Average ± SD a | |
| CSSGY1 | 23.57 ± 4.84 a | 6.22 ± 1.23 a | 8.47 ± 1.78 a | 6.12 ± 1.41 a |
| CSSGY4 | 16.33 ± 2.63 a | 6.12 ± 1.17 b | 8.16 ± 1.31 b | 6.12 ± 0.80 a |
| CSSGY5-1 | 15.31 ± 2.61 a | 5.71 ± 1.42 c | 8.88 ± 1.20 c | 6.12 ± 0.91 a |
| CSSGY5-2 | 15.31 ± 2.43 a | 5.71 ± 2.25 d | 9.11 ± 1.06 d | 7.06 ± 1.06 b |
| Species | Isolate/Strain | GenBank Acession Number | |||||
|---|---|---|---|---|---|---|---|
| ITS | ACT | CHS | GAPDH | TUB2 | ApMAT | ||
| Colletotrichum siamense | CBS 125378 | MH863512 | JX009441 | JX009875 | JX010019 | JX010410 | KP703513 |
| Colletotrichum siamense | ICMP 18578 | JX010171 | FJ907423 | JX009865 | JX009924 | JX010404 | JQ899289 |
| Colletotrichum siamense | CSSGY1 | ON428107 | ON454464 | ON454476 | ON454480 | ON454468 | ON454472 |
| Colletotrichum siamense | CSSGY5-1 | ON428108 | ON454466 | ON454478 | ON454482 | ON454470 | ON454474 |
| Colletotrichum siamense | CSSGY5-2 | ON428109 | ON454467 | ON454479 | ON454483 | ON454471 | ON454475 |
| Colletotrichum alienum | ICMP 18691 | JX010217 | JX009580 | JX009754 | JX010018 | JX010385 | |
| Colletotrichum alienum | ICMP 12071 | JX010251 | JX009572 | JX009882 | GU174553 | JX010411 | KC888927 |
| Colletotrichum fructicola | ICMP 18727 | JX010179 | JX009565 | JX009812 | JX010035 | JX010394 | |
| Colletotrichum fructicola | ICMP 18581 | JX010165 | FJ907426 | JX009866 | JX010033 | JX010405 | JQ807838 |
| Colletotrichum fructicola | CBS 125397 | MH863502 | JX009581 | JX009874 | JX010032 | JX010409 | |
| Colletotrichum fructicola | CSSGY4 | ON428106 | ON454465 | ON454477 | ON454481 | ON454469 | ON454473 |
| Colletotrichum nupharicola | CBS:472.96 | JX010188 | JX009582 | JX009836 | JX010031 | JX145226 | JX145320 |
| Colletotrichum nupharicola | CBS 469.96 | JX010189 | JX009486 | JX009834 | JX009836 | JX010399 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Zhu, J.-Z.; Li, X.-G.; Zhong, J. Identification and Characterization of Colletotrichum fructicola and Colletotrichum siamense Causing Anthracnose on Luffa Sponge Gourd in China. Plants 2022, 11, 1537. https://doi.org/10.3390/plants11121537
Li P, Zhu J-Z, Li X-G, Zhong J. Identification and Characterization of Colletotrichum fructicola and Colletotrichum siamense Causing Anthracnose on Luffa Sponge Gourd in China. Plants. 2022; 11(12):1537. https://doi.org/10.3390/plants11121537
Chicago/Turabian StyleLi, Ping, Jun-Zi Zhu, Xiao-Gang Li, and Jie Zhong. 2022. "Identification and Characterization of Colletotrichum fructicola and Colletotrichum siamense Causing Anthracnose on Luffa Sponge Gourd in China" Plants 11, no. 12: 1537. https://doi.org/10.3390/plants11121537
APA StyleLi, P., Zhu, J.-Z., Li, X.-G., & Zhong, J. (2022). Identification and Characterization of Colletotrichum fructicola and Colletotrichum siamense Causing Anthracnose on Luffa Sponge Gourd in China. Plants, 11(12), 1537. https://doi.org/10.3390/plants11121537