
The Known Antimammalian and Insecticidal Alkaloids Are Not Responsible for the Antifungal Activity of Epichloë Endophytes

 and
and 
Abstract
1. Introduction
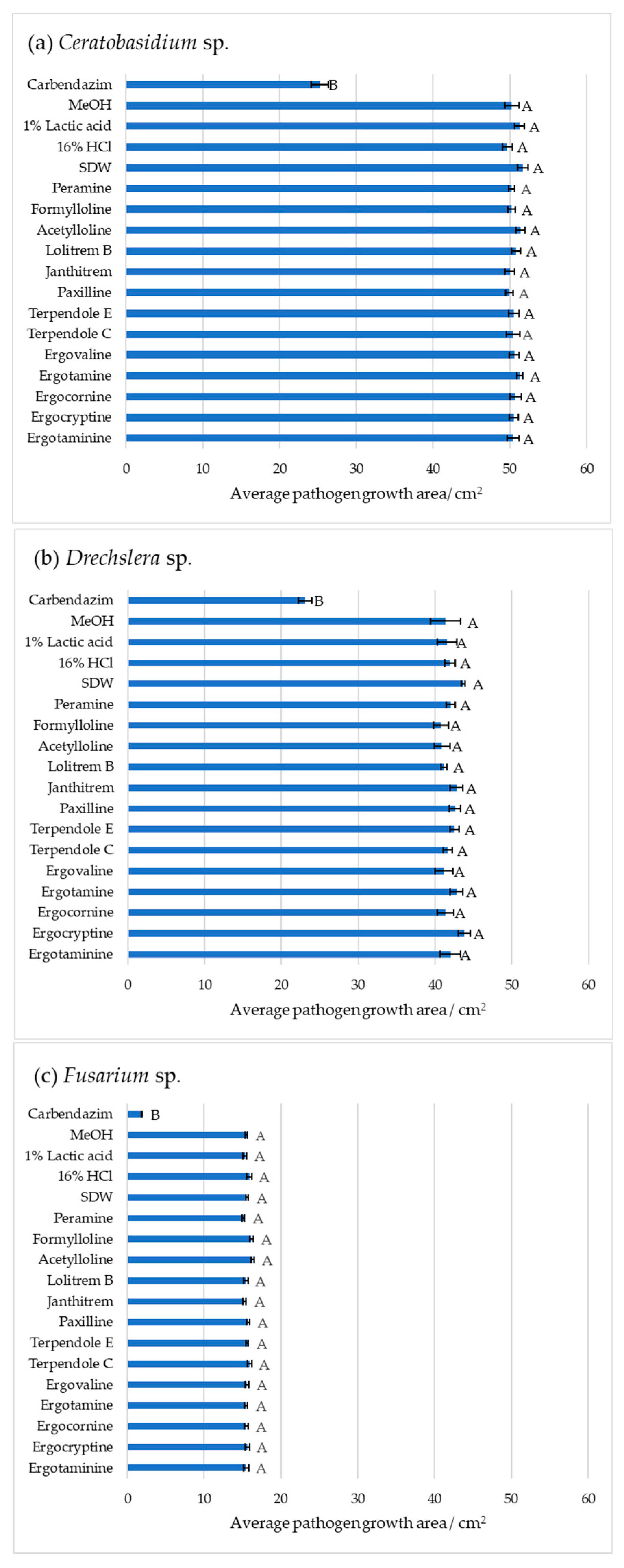
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material
5.2. Plant Pathogens
5.3. Chemicals
5.4. Agar Well Diffusion Assay
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Christensen, M. Antifungal activity in grasses infected with Acremonium and Epichloë endophytes. Australas. Plant Pathol. 1996, 25, 186–191. [Google Scholar] [CrossRef]
- Scott, B. Epichloë endophytes: Fungal symbionts of grasses. Curr. Opin. Microbiol. 2001, 4, 393–398. [Google Scholar] [CrossRef]
- Tian, P.; Nan, Z.; Li, C.; Spangenberg, G. Effect of the endophyte Neotyphodium lolii on susceptibility and host physiological response of perennial ryegrass to fungal pathogens. Eur. J. Plant Pathol. 2008, 122, 593–602. [Google Scholar] [CrossRef]
- Ruppert, K.G.; Matthew, C.; McKenzie, C.M.; Popay, A.J. Impact of Epichloë endophytes on adult Argentine stem weevil damage to perennial ryegrass seedlings. Entomol. Exp. Appl. 2017, 163, 328–337. [Google Scholar] [CrossRef]
- Reddy, P.; Guthridge, K.; Vassiliadis, S.; Hemsworth, J.; Hettiarachchige, I.; Spangenberg, G.; Rochfort, S. Tremorgenic mycotoxins: Structure diversity and biological activity. Toxins 2019, 11, 302. [Google Scholar] [CrossRef]
- Reddy, P.; Hemsworth, J.; Guthridge, K.M.; Vinh, A.; Vassiliadis, S.; Ezernieks, V.; Spangenberg, G.C.; Rochfort, S.J. Ergot alkaloid mycotoxins: Physiological effects, metabolism and distribution of the residual toxin in mice. Sci. Rep. 2020, 10, 9714. [Google Scholar] [CrossRef]
- Bacon, C.W. Procedure for isolating the endophyte from tall fescue and screening isolates for ergot alkaloids. Appl. Environ. Microbiol. 1988, 54, 2615–2618. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, R.T.; Hawkes, A.D.; Steyn, P.S.; Vleggaar, R. Tremorgenic neurotoxins from perennial ryegrass causing ryegrass staggers disorder of livestock: Structure elucidation of lolitrem B. J. Chem. Soc. Chem. Commun. 1984, 9, 614–616. [Google Scholar] [CrossRef]
- Xia, C.; Li, N.; Zhang, Y.; Li, C.; Zhang, X.; Nan, Z. Role of Epichloë endophytes in defense responses of cool-season grasses to pathogens: A review. Plant Dis. 2018, 102, 2061–2073. [Google Scholar] [CrossRef] [PubMed]
- Latch, G. Fungous diseases of ryegrasses in New Zealand: I. Foliage diseases. N. Z. J. Agric. Res. 1966, 9, 394–409. [Google Scholar] [CrossRef]
- Kolliker, R.; Kraehenbuehl, R.; Boller, B.; Widmer, F. Genetic diversity and pathogenicity of the grass pathogen Xanthomonas translucens pv. graminis. Syst. Appl. Microbiol. 2006, 29, 109–119. [Google Scholar] [CrossRef]
- Smith, B.; Towers, N. Mycotoxicoses of grazing animals in New Zealand. N. Z. Vet. J. 2002, 50, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Di Menna, M.E.; Smith, B.L.; Miles, C.O. A history of facial eczema (pithomycotoxicosis) research. N. Z. J. Agric. Res. 2009, 52, 345–376. [Google Scholar] [CrossRef]
- Tangni, E.K.; Pussemier, L.; Van Hove, F. Mycotoxin contaminating maize and grass silages for dairy cattle feeding: Current state and challenges. J. Anim. Sci. Adv. 2013, 3, 492–511. [Google Scholar]
- Bacon, C.; Porter, J.; Robbins, J.; Luttrell, E. Epichloë typhina from toxic tall fescue grasses. Appl. Environ. Microbiol. 1977, 34, 576–581. [Google Scholar] [CrossRef]
- Zhou, L.; Zhang, X.; Li, C.; Christensen, M.J.; Nan, Z. Antifungal activity and phytochemical investigation of the asexual endophyte of Epichloë sp. from Festuca sinensis. Sci. Chi. Life Sci. 2015, 58, 821–826. [Google Scholar] [CrossRef]
- Niones, J.T.; Takemoto, D. An isolate of Epichloë festucae, an endophytic fungus of temperate grasses, has growth inhibitory activity against selected grass pathogens. J. Gen. Plant Pathol. 2014, 80, 337–347. [Google Scholar] [CrossRef]
- Siegel, M.R.; Latch, G.C. Expression of antifungal activity in agar culture by isolates of grass endophytes. Mycologia 1991, 83, 529–537. [Google Scholar] [CrossRef]
- Christensen, M.J.; Latch, G.C.M. Variation among isolates of Acremonium endophytes (A. coenophialum and possibly A. typhinum) from tall fescue (Festuca arundinacea). Mycol. Res. 1991, 95, 1123–1126. [Google Scholar] [CrossRef]
- Fernando, K.; Reddy, P.; Hettiarachchige, I.K.; Spangenberg, G.C.; Rochfort, S.J.; Guthridge, K.M. Novel antifungal activity of Lolium-associated Epichloë endophytes. Microorganisms 2020, 8, 955. [Google Scholar] [CrossRef]
- Yue, Q.; Miller, C.J.; White, J.F.; Richardson, M.D. Isolation and characterization of fungal inhibitors from Epichloë festucae. J. Agric. Food Chem. 2000, 48, 4687–4692. [Google Scholar] [CrossRef]
- Pańka, D.; Jeske, M.; Troczyński, M. Effect of Neotyphodium uncinatum endophyte on meadow fescue yielding, health status and ergovaline production in host-plants. J. Plant Prot. Res. 2011, 51, 362–370. [Google Scholar] [CrossRef]
- White, J.F.; Cole, G.T. Endophyte-host associations in forage grasses. IV. The endophyte of Festuca versuta. Mycologia 1986, 78, 102–107. [Google Scholar] [CrossRef]
- Hiroyuki, K.; Satoshi, T.; Shun-ichi, T.; Yoshihara, T.; Sakamura, S.; Shimanuki, T.; Sato, T.; Tajimi, A. New fungitoxic sesquiterpenoids, chokols AG, from stromata of Epichloe typhina and the absolute configuration of chokol E. Agric. Biol. Chem. 1989, 53, 789–796. [Google Scholar] [CrossRef]
- Koshino, H.; Yoshihara, T.; Ichihara, A.; Tajimi, A.; Shimanuki, T. Two sphingoid derivatives from stromata of Epichloë typhina on Phleum pratense. Phytochemistry 1992, 31, 3757–3759. [Google Scholar] [CrossRef]
- Koshino, H.; Yoshihara, T.; Okuno, M.; Sakamura, S.; Tajimi, A.; Shimanuki, T. Gamahonolides A, B, and gamahorin, novel antifungal compounds from stromata of Epichloë typhina on Phleum pratense. Biosci. Biotechnol. Biochem. 1992, 56, 1096–1099. [Google Scholar] [CrossRef]
- Seto, Y.; Takahashi, K.; Matsuura, H.; Kogami, Y.; Yada, H.; Yoshihara, T.; Nabeta, K. Novel cyclic peptide, epichlicin, from the endophytic fungus, Epichloe typhina. Biosci. Biotechnol. Biochem. 2007, 71, 1470–1475. [Google Scholar] [CrossRef][Green Version]
- Purev, E.; Kondo, T.; Takemoto, D.; Niones, J.T.; Ojika, M. Identification of ε-poly-l-lysine as an antimicrobial product from an Epichloë endophyte and isolation of fungal ε-PL synthetase gene. Molecules 2020, 25, 1032. [Google Scholar] [CrossRef]
- Tian, Z.; Wang, R.; Ambrose, K.V.; Clarke, B.B.; Belanger, F.C. The Epichloë festucae antifungal protein has activity against the plant pathogen Sclerotinia homoeocarpa, the causal agent of dollar spot disease. Sci. Rep. 2017, 7, 5643. [Google Scholar] [CrossRef]
- Kaur, J.; Ekanayake, P.N.; Tian, P.; van Zijll de Jong, E.; Dobrowolski, M.P.; Rochfort, S.J.; Mann, R.C.; Smith, K.F.; Forster, J.W.; Guthridge, K.M.; et al. Discovery and characterisation of novel asexual Epichloë endophytes from perennial ryegrass (Lolium perenne L.). Crop. Past. Sci. 2015, 66, 1058. [Google Scholar] [CrossRef]
- Ekanayake, P.N.; Kaur, J.; Tian, P.; Rochfort, S.J.; Guthridge, K.M.; Sawbridge, T.I.; Spangenberg, G.C.; Forster, J.W. Genomic and metabolic characterisation of alkaloid biosynthesis by asexual Epichloë fungal endophytes of tall fescue pasture grasses. Genome 2017, 60, 496–509. [Google Scholar] [CrossRef]
- Babu, J.V.; Popay, A.J.; Miles, C.O.; Wilkins, A.L.; di Menna, M.E.; Finch, S.C. Identification and structure elucidation of janthitrems A and D from Penicillium janthinellum and determination of the tremorgenic and anti-insect activity of janthitrems A and B. J. Agric. Food Chem. 2018, 66, 13116–13125. [Google Scholar] [CrossRef] [PubMed]
- Tapper, B.A.; Cooper, B.M.; Easton, H.S.; Fletcher, L.R.; Hume, D.E.; Lane, G.A.; Latch, G.C.M.; Pennell, C.G.L.; Popay, A.J.; Christensen, M.J. Grass Endophytes; World Intellectual Property Organization: Geneva, Switzerland, 2004; Available online: https://patents.google.com/patent/WO2004106487A2/un (accessed on 12 November 2021).
- Oliver, A.M.; Leathwick, D.M.; Waghorn, T.S.; Miller, C.M.; Atkinson, D.S. Carbendazim, at concentrations used on pasture for facial eczema control, reduces development of Trichostrongylus colubriformis when sprayed onto infected sheep faeces. N. Z. Vet. J. 2009, 57, 383–387. [Google Scholar] [CrossRef]
- Al-Rahmah, A.; Mostafa, A.; Abdel-Megeed, A.; Yakout, S.; Hussein, S. Fungicidal activities of certain methanolic plant extracts against tomato phytopathogenic fungi. Afr. J. Microbiol. Res. 2013, 7, 517–524. [Google Scholar]
- Reddy, P.; Rochfort, S.; Read, E.; Deseo, M.; Jaehne, E.; Van Den Buuse, M.; Guthridge, K.; Combs, M.; Spangenberg, G.; Quinn, J. Tremorgenic effects and functional metabolomics analysis of lolitrem B and its biosynthetic intermediates. Sci. Rep. 2019, 9, 9364. [Google Scholar] [CrossRef]
- Reddy, P.; Vincent, D.; Hemsworth, J.; Ezernieks, V.; Guthridge, K.; Spangenberg, G.C.; Rochfort, S.J. Effects of ergotamine on the central nervous system using untargeted metabolomics analysis in a mouse model. Sci. Rep. 2021, 11, 19542. [Google Scholar] [CrossRef]
- Finch, S.C.; Prinsep, M.R.; Popay, A.J.; Wilkins, A.L.; Webb, N.G.; Bhattarai, S.; Jensen, J.G.; Hawkes, A.D.; Babu, J.V.; Tapper, B.A.; et al. Identification and structure elucidation of epoxyjanthitrems from lolium perenne infected with the endophytic fungus Epichloë festucae var. lolii and determination of the tremorgenic and anti-insect activity of epoxyjanthitrem I. Toxins 2020, 12, 526. [Google Scholar] [CrossRef]
- Cherewyk, J.E.; Parker, S.E.; Blakley, B.R.; Al-Dissi, A.N. Assessment of the vasoactive effects of the (S)-epimers of ergot alkaloids in vitro. J. Anim. Sci. 2020, 98, skaa203. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Sharma, V.K.; Manikyam, H.K.; Krishna, A.B. Ergot alkaloids: A review on therapeutic applications. Eur. J. Med. Plants 2016, 14, 1–17. [Google Scholar] [CrossRef]
- Rowan, D.D.; Dymock, J.J.; Brimble, M.A. Effect of fungal metabolite peramine and analogs on feeding and development of argentine stem weevil (Listronotus bonariensis). J. Chem. Ecol. 1990, 16, 1683–1695. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, H.H.; Siegel, M.R.; Blankenship, J.D.; Mallory, A.C.; Bush, L.P.; Schardl, C.L. Contribution of fungal loline alkaloids to protection from aphids in a grass-endophyte mutualism. Mol. Plant-Microbe Interact. 2000, 13, 1027–1033. [Google Scholar] [CrossRef]
- Schardl, C.L.; Grossman, R.B.; Nagabhyru, P.; Faulkner, J.R.; Mallik, U.P. Loline alkaloids: Currencies of mutualism. Phytochemistry 2007, 68, 980–996. [Google Scholar] [CrossRef] [PubMed]
- Vassiliadis, S.; Elkins, A.C.; Reddy, P.; Guthridge, K.M.; Spangenberg, G.C.; Rochfort, S.J. A simple LC-MS method for the quantitation of alkaloids in endophyte-infected perennial ryegrass. Toxins 2019, 11, 649. [Google Scholar] [CrossRef] [PubMed]
- Reed, K.; Nie, Z.; Walker, L.; Kearney, G. Fluctuations in the concentration of ergovaline and lolitrem B produced by the wild-type endophyte (Neotyphodium lolii) in perennial ryegrass (Lolium perenne) pasture. Anim. Prod. Sci. 2011, 51, 1098–1108. [Google Scholar] [CrossRef]
- Fletcher, L.R.; Finch, S.C.; Sutherland, B.L.; deNicolo, G.; Mace, W.J.; van Koten, C.; Hume, D.E. The occurrence of ryegrass staggers and heat stress in sheep grazing ryegrass-endophyte associations with diverse alkaloid profiles. N. Z. Vet. J. 2017, 65, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.; Krischke, M.; Mueller, M.J.; Krauss, J. Plant age and seasonal timing determine endophyte growth and alkaloid biosynthesis. Fungal Ecol. 2017, 29, 52–58. [Google Scholar] [CrossRef]
- Vikuk, V.; Fuchs, B.; Krischke, M.; Mueller, M.J.; Rueb, S.; Krauss, J. Alkaloid concentrations of Lolium perenne infected with Epichloë festucae var. lolii with different detection methods—A re-evaluation of intoxication risk in Germany? J. Fungi 2020, 6, 177. [Google Scholar] [CrossRef] [PubMed]
- Repussard, C.; Zbib, N.; Tardieu, D.; Guerre, P. Ergovaline and lolitrem B concentrations in perennial ryegrass in field culture in southern France: Distribution in the plant and impact of climatic factors. J. Agric. Food Chem. 2014, 62, 12707–12712. [Google Scholar] [CrossRef]
- Ludlow, E.J.; Vassiliadis, S.; Ekanayake, P.N.; Hettiarachchige, I.K.; Reddy, P.; Sawbridge, T.I.; Rochfort, S.J.; Spangenberg, G.C.; Guthridge, K.M. Analysis of the indole diterpene gene cluster for biosynthesis of the epoxy-janthitrems in Epichloë endophytes. Microorganisms 2019, 7, 560. [Google Scholar] [CrossRef]
- Wachowska, U. Susceptibility of cereals and other crops to Rhizoctonia cerealis. Phytopathol. Pol. 2000, 20, 59–66. [Google Scholar]
- Burpee, L. Rhizoctonia cerealis causes yellow patch of turfgrasses. Plant Dis. 1980, 64, 1114–1116. [Google Scholar] [CrossRef]
- Sneh, B.; Jabaji-Hare, S.; Neate, S.; Dijst, G. Rhizoctonia Species: Taxonomy, Molecular Biology, Ecology, Pathology and Disease Control, 2nd ed.; Springer Science & Business Media, Kluwer: Dordrecht, The Netherlands, 1996. [Google Scholar]
- Labruyere, R. Contamination of ryegrass seed with Drechslera species and its effect on disease incidence in the ensuing crop. Neth. J. Pl. Path. 1977, 83, 205–215. [Google Scholar] [CrossRef]
- Hsiang, T. All you ever wanted to know about Fusarium patch/Microdochium patch/pink snow mold or whatever that disease is called. Green Master 2009, 44, 13–16. [Google Scholar]
- Reddy, P.; Deseo, M.A.; Ezernieks, V.; Guthridge, K.; Spangenberg, G.; Rochfort, S. Toxic indole diterpenes from endophyte-infected perennial ryegrass Lolium perenne L.: Isolation and stability. Toxins 2019, 11, 16. [Google Scholar] [CrossRef] [PubMed]
- Beloshapkina, O.O.; Katushova, M.S.; Kalashnikov, D.V. Improving methods for preventing diseases of pasture grasses and securing the quality of feedstuff. IOP Conf. Ser. Earth Environ. Sci. 2021, 624, 012150. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Yu, J.-J.; Zhang, Y.-N.; Zhang, X.; Cheng, C.-J.; Wang, J.-X.; Hollomon, D.W.; Fan, P.-S.; Zhou, M.-G. Effect of carbendazim resistance on trichothecene production and aggressiveness of Fusarium graminearum. Mol. Plant-Microbe Interact. 2009, 22, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Marei, G.I.K.; Abdel Rasoul, M.A.; Abdelgaleil, S.A.M. Comparative antifungal activities and biochemical effects of monoterpenes on plant pathogenic fungi. Pestic. Biochem. Physiol. 2012, 103, 56–61. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | Host/Source | Accession Number | Disease | Effect on Pasture/ Seed Production | References |
|---|---|---|---|---|---|
| Ceratobasidium sp. | Triticum aestivum (wheat) | VPRI 22537 | yellow patch/ sharp eyespot | Reduced yield | [51,52,53] |
| Drechslera sp. | Briza maxima (rattle grass) | VPRI 12962 | brown blight and net blotch | Dry matter and herbage yield reduction | [10,54] |
| Fusarium sp. | Lolium perenne (perennial ryegrass) | VPRI 43403 | Fusarium patch | Reduced yield and dry matter | [55] |
| Biosynthesis Pathway | Purified Compound (Purity, %) | Chemical Formula | Concentration Used in Bioassay (ppm) | Concentration in Planta 1,3 (ppm) | Source |
|---|---|---|---|---|---|
| Peramine | Peramine hemisulfate | C12H17N5O.H2SO4 | 100 | 52.24 ± 3.10 | Toronto Research Chemicals, North York, ON, Canada |
| Loline | n-Formylloline-d3 (>98%) | C9H11D3N2O2 | 500 | 427.92 ± 26.62 | Toronto Research Chemicals, North York, ON, Canada |
| n-Acetylloline. Hydrochloride (>98%) | C10H16N2O2.HCl | 250 | typically trace levels in perennial ryegrass 2 | Toronto Research Chemicals, North York, ON, Canada | |
| Indole diterpene | Lolitrem B (>98%) | C42H55NO7 | 10 | 4.88 ± 0.64 | (Reddy et al., 2019) [56] |
| Janthitrem A (>95%) | C37H47NO6 | 50 | 11.86 ± 0.85 | BioAustralis, Smithfield NSW, Australia | |
| Paxilline (>98%) | C27H33NO4 | 10 | 0.24 ± 0.05 | Sigma-Aldrich, St. Louis, MO, USA | |
| Terpendole E (>95%) | C28H39NO3 | 10 | 0.061 ± 0.01 | Sigma-Aldrich, St. Louis, MO, USA | |
| Terpendole C (>95%) | C32H41NO5 | 10 | 1.33 ± 0.23 | Sigma-Aldrich, St. Louis, MO, USA | |
| Ergot alkaloid | Ergovaline (≥97%) | C29H35N5O5 | 10 | 0.40 ± 0.08 | (Reddy et al., 2020) [6] |
| Ergotamine (≥97%) | C33H35N5O5 | 10 | NA | LGC Standards, Wesel, Germany | |
| Ergocornine (>98%) | C31H39N5O5 | 10 | NA | LGC Standards, Wesel, Germany | |
| Ergocryptine (>98%) | C32H41N5O5 | 10 | NA | LGC Standards, Wesel, Germany | |
| Ergotaminine (>98%) | C33H35N5O5 | 10 | NA | LGC Standards, Wesel, Germany |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernando, K.; Reddy, P.; Vassiliadis, S.; Spangenberg, G.C.; Rochfort, S.J.; Guthridge, K.M. The Known Antimammalian and Insecticidal Alkaloids Are Not Responsible for the Antifungal Activity of Epichloë Endophytes. Plants 2021, 10, 2486. https://doi.org/10.3390/plants10112486
Fernando K, Reddy P, Vassiliadis S, Spangenberg GC, Rochfort SJ, Guthridge KM. The Known Antimammalian and Insecticidal Alkaloids Are Not Responsible for the Antifungal Activity of Epichloë Endophytes. Plants. 2021; 10(11):2486. https://doi.org/10.3390/plants10112486
Chicago/Turabian StyleFernando, Krishni, Priyanka Reddy, Simone Vassiliadis, German C. Spangenberg, Simone J. Rochfort, and Kathryn M. Guthridge. 2021. "The Known Antimammalian and Insecticidal Alkaloids Are Not Responsible for the Antifungal Activity of Epichloë Endophytes" Plants 10, no. 11: 2486. https://doi.org/10.3390/plants10112486
APA StyleFernando, K., Reddy, P., Vassiliadis, S., Spangenberg, G. C., Rochfort, S. J., & Guthridge, K. M. (2021). The Known Antimammalian and Insecticidal Alkaloids Are Not Responsible for the Antifungal Activity of Epichloë Endophytes. Plants, 10(11), 2486. https://doi.org/10.3390/plants10112486