

Intratumoral Microbiota: Metabolic Influences and Biomarker Potential in Gastrointestinal Cancer
Abstract
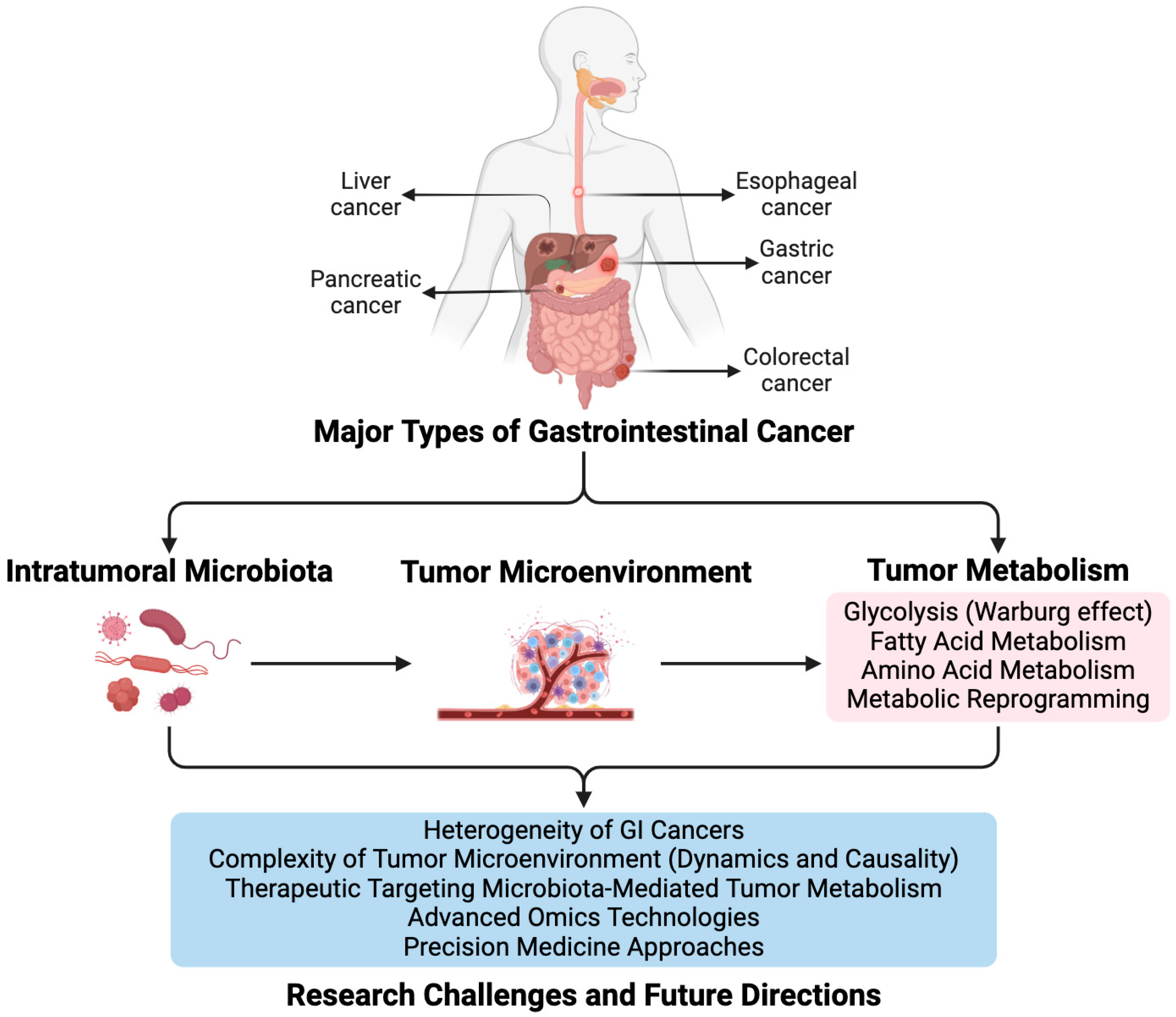
1. Introduction
2. Intratumoral Microbiota and GI Cancer
2.1. Origin of Intratumoral Microbiota in GI Cancer
2.2. Recent Studies on the Potential Mechanisms and Biomarker Significance of Intratumoral Microbiota in GI Cancer
2.2.1. Esophageal Cancer
2.2.2. Gastric Cancer
2.2.3. Liver Cancer
2.2.4. Pancreatic Cancer
2.2.5. Colorectal Cancer

{kind=link}
{kind=link}
| Year of Study | Nature of Study | Participants | Aim and Main Findings | Refs. |
|---|---|---|---|---|
| 2024 | Observational study | CRC patients | This study aimed to elucidate the genetic factors of F. nucleatum facilitating tumor colonization in CRC by analyzing closed genomes of 135 F. nucleatum strains. It identifies a distinct clade, Fna C2, as predominant in CRC tumors, showing increased metabolic potential and colonization of the GI tract, and providing insights into the pathoadaptation of Fna C2 to the CRC tumor niche. | [76] |
| 2023 | Observational study | CRC patients | The study proposes a method to detect bacterial signals in human RNA sequencing data and associates them with clinical and molecular properties of tumors. The analysis reveals correlations between intratumoral microbiome composition and survival, anatomic location, microsatellite instability, consensus molecular subtype, and immune cell infiltration in colon tumors. | [84] |
| 2024 | Prospective-Observational study | CRC patients | This study investigates the tumor microbial profile of young-onset CRC (yoCRC) compared to average-onset CRC (aoCRC), revealing higher microbial diversity and distinct microbial compositions in yoCRC tumors. Akkermansia and Bacteroides are enriched in yoCRC tumors, while aoCRC tumors show more abundances of several other bacteria. | [85] |
| 2023 | Observational study | Patients with locally advanced rectal cancer | This study investigates the tumor-bearing microbiota in patients with locally advanced rectal cancer before neoadjuvant chemoradiation therapy (nCRT) and its association with treatment response. The findings reveal specific microbial biomarkers and functional pathways associated with resistance to nCRT, highlighting the potential role of intratumoral microbiota in modulating treatment outcomes in rectal cancer patients. | [86] |
| 2023 | Observational study | CRC patients (data from the TCGA) | The aim of this study was to unravel the potential remodeling mechanisms of immune cell infiltration and tumorigenesis in CRC by integrating genetic, epigenetic, and intratumor microbial factors. Results reveal the significant influence of intratumor microbes on immune cell infiltration patterns, prognosis, and response to immune checkpoint blockade therapy in CRC. | [87] |
| 2023 | Observational study | Locally advanced rectal cancer (LARC) patients (data from a published European cohort) | This study investigates the intratumoral microbiota in LARC patients and its association with the response to nCRT. It identifies microbial signatures associated with pathological complete response (pCR) and non-pCR groups, highlighting their potential as independent predictive markers for nCRT response and revealing interactions between intratumoral microbes and cancer-associated fibroblasts (CAFs) in mediating treatment response. | [88] |
| 2024 | Observational study | CRC patients | The study aimed to understand how Colibactin-producing E. coli influences tumor heterogeneity, chemoresistance, and patient survival in right-sided CRC tumors. The main findings show that Colibactin-producing E. coli-infected tumors had high glycerophospholipid environments, reduced CD8+ T lymphocyte infiltration, and increased chemoresistance through lipid droplet accumulation and phosphatidylcholine remodeling. | [89] |
| 2021 | Observational study | CRC patients | The study explored the association between the intratumor microbiome and host genetic alterations in CRC patients. Fusobacterium was associated with mutated genes and cell cycle-related pathways, while Campylobacter abundance was linked to mutational signature 3, suggesting a potential role of bacterial-induced DNA damage in CRC. | [90] |
| 2017 | Observational study | Microsatellite instability-high (MSI-H) CRC patients | The study aimed to investigate the clinicopathologic and molecular associations of F. nucleatum in MSI-H CRC patients. High intratumoral F. nucleatum were associated with increased macrophage infiltration and CDKN2A promoter methylation in MSI-H CRC. | [91] |
| 2021 | Observational study | MSI-H CRC patients | In MSI-H CRC, high levels of intratumoral F. nucleatum are associated with larger tumor size and advanced invasion depth. Additionally, F. nucleatum-enriched tumors exhibit decreased density of FoxP3+ T cells and an increased proportion of M2-polarized macrophages in the tumor center. | [92] |
| 2018 | Observational study | CRC patients | The aim of this study was to investigate the association between the amount of Bifidobacteria in CRC tissue and tumor differentiation, specifically the extent of signet ring cells, as well as the immune response to CRC. The main findings reveal that intratumor bifidobacteria were detected in 30% of cases and were associated with the extent of signet ring cells, suggesting a possible role of bifidobacteria in determining distinct tumor characteristics or as an indicator of dysfunctional mucosal barrier in CRC. | [93] |
| 2019 | Observational study | CRC patients | The aim of this study was to investigate the prognostic impact of intratumoral F. nucleatum in CRC patients treated with adjuvant chemotherapy. Intratumoral F. nucleatum load was found to be a potential prognostic factor in stage II/III CRC patients treated with oxaliplatin-based adjuvant chemotherapy, particularly in non-MSI-H/non-sigmoid/non-rectal cancer subsets. | [94] |
3. Role of Intratumoral Microbiota in Modulating GI Tumor Metabolism
3.1. Influence of Microbial Dysbiosis on Tumor Metabolic Reprogramming
3.2. Specific Metabolic Pathways Affected by Intratumoral Microbiota in GI Cancer
3.2.1. Glycolysis
3.2.2. Fatty Acid Metabolism
3.2.3. Amino Acid Metabolism
4. Clinical Implications of Intratumoral Microbiota-Mediated Tumor Metabolism in GI Cancer
4.1. Prognostic Significance of Intratumoral Microbiota Composition
4.2. Therapeutic Opportunities Targeting Microbiota-Mediated Tumor Metabolism in GI Cancer
5. Challenges and Future Directions
5.1. Current Limitations in Understanding the Complex Interplay between Intratumoral Microbiota and Tumor Metabolism
5.2. Future Research Directions for Unraveling the Mechanisms and Clinical Applications in GI Cancer
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Arnold, M.; Abnet, C.C.; Neale, R.E.; Vignat, J.; Giovannucci, E.L.; McGlynn, K.A.; Bray, F. Global Burden of 5 Major Types of Gastrointestinal Cancer. Gastroenterology 2020, 159, 335–349.e15. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.H.; Guan, C.A.; Bujang, M.A.; Lai, W.H.; Voon, P.J.; Sim, E.U.H. Identification of Phenomic Data in the Pathogenesis of Cancers of the Gastrointestinal (GI) Tract in the UK Biobank. Sci. Rep. 2024, 14, 1997. [Google Scholar] [CrossRef] [PubMed]
- Jardim, S.R.; de Souza, L.M.P.; de Souza, H.S.P. The Rise of Gastrointestinal Cancers as a Global Phenomenon: Unhealthy Behavior or Progress? Int. J. Environ. Res. Public Health 2023, 20, 3640. [Google Scholar] [CrossRef] [PubMed]
- Marabotto, E.; Kayali, S.; Buccilli, S.; Levo, F.; Bodini, G.; Giannini, E.G.; Savarino, V.; Savarino, E.V. Colorectal Cancer in Inflammatory Bowel Diseases: Epidemiology and Prevention: A Review. Cancers 2022, 14, 4254. [Google Scholar] [CrossRef] [PubMed]
- McGuigan, A.; Kelly, P.; Turkington, R.C.; Jones, C.; Coleman, H.G.; McCain, R.S. Pancreatic Cancer: A Review of Clinical Diagnosis, Epidemiology, Treatment and Outcomes. World J. Gastroenterol. 2018, 24, 4846–4861. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, P.; Mei, W.; Zeng, C. Intratumoral Microbiota: Implications for Cancer Onset, Progression, and Therapy. Front. Immunol. 2023, 14, 1301506. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, A.; Wang, Y.; Zhang, Y. Intratumoral Microbiota: Roles in Cancer Initiation, Development and Therapeutic Efficacy. Signal Transduct. Target. Ther. 2023, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Li, J.; Cai, W.; Huang, Y.; Liu, X.; Ma, Z.; Tang, Z.; Bian, X.; Zheng, J.; Jiang, J.; et al. The Emerging Tumor Microbe Microenvironment: From Delineation to Multidisciplinary Approach-Based Interventions. Acta Pharm. Sin. B 2024, 14, 1560–1591. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, K.; Taylor, C.T. The Impact of Hypoxia on Bacterial Infection. FEBS J. 2015, 282, 2260–2266. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Yu, B.; Rao, B.; Sun, Y.; Yu, J.; Wang, D.; Cui, G.; Ren, Z. The Effect of the Intratumoral Microbiome on Tumor Occurrence, Progression, Prognosis and Treatment. Front. Immunol. 2022, 13, 1051987. [Google Scholar] [CrossRef]
- Xuan, M.; Gu, X.; Liu, Y.; Yang, L.; Li, Y.; Huang, D.; Li, J.; Xue, C. Intratumoral Microorganisms in Tumors of the Digestive System. Cell Commun. Signal 2024, 22, 69. [Google Scholar] [CrossRef]
- Hammoudi, N.; Ahmed, K.B.; Garcia-Prieto, C.; Huang, P. Metabolic Alterations in Cancer Cells and Therapeutic Implications. Chin. J. Cancer 2011, 30, 508–525. [Google Scholar] [CrossRef]
- Yang, J.; Shay, C.; Saba, N.F.; Teng, Y. Cancer Metabolism and Carcinogenesis. Exp. Hematol. Oncol. 2024, 13, 10. [Google Scholar] [CrossRef]
- Pavlova, N.N.; Zhu, J.; Thompson, C.B. The Hallmarks of Cancer Metabolism: Still Emerging. Cell Metab. 2022, 34, 355–377. [Google Scholar] [CrossRef]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does It Benefit Cancer Cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef]
- Zhou, D.; Duan, Z.; Li, Z.; Ge, F.; Wei, R.; Kong, L. The Significance of Glycolysis in Tumor Progression and Its Relationship with the Tumor Microenvironment. Front. Pharmacol. 2022, 13, 1091779. [Google Scholar] [CrossRef] [PubMed]
- Gatenby, R.A.; Gillies, R.J. Why Do Cancers Have High Aerobic Glycolysis? Nat. Rev. Cancer 2004, 4, 891–899. [Google Scholar] [CrossRef]
- Lunt, S.Y.; Vander Heiden, M.G. Aerobic Glycolysis: Meeting the Metabolic Requirements of Cell Proliferation. Annu. Rev. Cell Dev. Biol. 2011, 27, 441–464. [Google Scholar] [CrossRef]
- Schiliro, C.; Firestein, B.L. Mechanisms of Metabolic Reprogramming in Cancer Cells Supporting Enhanced Growth and Proliferation. Cells 2021, 10, 1056. [Google Scholar] [CrossRef]
- Elia, I.; Haigis, M.C. Metabolites and the Tumour Microenvironment: From Cellular Mechanisms to Systemic Metabolism. Nat. Metab. 2021, 3, 21–32. [Google Scholar] [CrossRef]
- Zhao, Y.; Shen, M.; Wu, L.; Yang, H.; Yao, Y.; Yang, Q.; Du, J.; Liu, L.; Li, Y.; Bai, Y. Stromal Cells in the Tumor Microenvironment: Accomplices of Tumor Progression? Cell Death Dis. 2023, 14, 587. [Google Scholar] [CrossRef]
- Lobel, G.P.; Jiang, Y.; Simon, M.C. Tumor Microenvironmental Nutrients, Cellular Responses, and Cancer. Cell Chem. Biol. 2023, 30, 1015–1032. [Google Scholar] [CrossRef]
- Kaymak, I.; Williams, K.S.; Cantor, J.R.; Jones, R.G. Immunometabolic Interplay in the Tumor Microenvironment. Cancer Cell 2021, 39, 28–37. [Google Scholar] [CrossRef]
- Thursby, E.; Juge, N. Introduction to the Human Gut Microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef]
- Singh, R.; Zogg, H.; Wei, L.; Bartlett, A.; Ghoshal, U.C.; Rajender, S.; Ro, S. Gut Microbial Dysbiosis in the Pathogenesis of Gastrointestinal Dysmotility and Metabolic Disorders. J. Neurogastroenterol. Motil. 2021, 27, 19–34. [Google Scholar] [CrossRef]
- Agagunduz, D.; Cocozza, E.; Cemali, O.; Bayazit, A.D.; Nani, M.F.; Cerqua, I.; Morgillo, F.; Saygili, S.K.; Berni Canani, R.; Amero, P.; et al. Understanding the Role of the Gut Microbiome in Gastrointestinal Cancer: A Review. Front. Pharmacol. 2023, 14, 1130562. [Google Scholar] [CrossRef]
- Wang, M.; Yu, F.; Li, P. Intratumor Microbiota in Cancer Pathogenesis and Immunity: From Mechanisms of Action to Therapeutic Opportunities. Front. Immunol. 2023, 14, 1269054. [Google Scholar] [CrossRef]
- Xie, Y.; Xie, F.; Zhou, X.; Zhang, L.; Yang, B.; Huang, J.; Wang, F.; Yan, H.; Zeng, L.; Zhang, L.; et al. Microbiota in Tumors: From Understanding to Application. Adv. Sci. 2022, 9, e2200470. [Google Scholar] [CrossRef]
- Tjalsma, H.; Boleij, A.; Marchesi, J.R.; Dutilh, B.E. A Bacterial Driver-Passenger Model for Colorectal Cancer: Beyond the Usual Suspects. Nat. Rev. Microbiol. 2012, 10, 575–582. [Google Scholar] [CrossRef]
- Dohlman, A.B.; Arguijo Mendoza, D.; Ding, S.; Gao, M.; Dressman, H.; Iliev, I.D.; Lipkin, S.M.; Shen, X. The Cancer Microbiome Atlas: A Pan-Cancer Comparative Analysis to Distinguish Tissue-Resident Microbiota from Contaminants. Cell Host Microbe 2021, 29, 281–298.e5. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y.; Li, Z.; Gao, X.; Huang, D. Global Analysis of Microbiota Signatures in Four Major Types of Gastrointestinal Cancer. Front. Oncol. 2021, 11, 685641. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, H.; Gao, X.; Wang, J. The Intratumor Microbiota Signatures Associate with Subtype, Tumor Stage, and Survival Status of Esophageal Carcinoma. Front. Oncol. 2021, 11, 754788. [Google Scholar] [CrossRef]
- Wu, H.; Leng, X.; Liu, Q.; Mao, T.; Jiang, T.; Liu, Y.; Li, F.; Cao, C.; Fan, J.; Chen, L.; et al. Intratumoral Microbiota Composition Regulates Chemoimmunotherapy Response in Esophageal Squamous Cell Carcinoma. Cancer Res. 2023, 83, 3131–3144. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, S.; Ma, X.; Zhan, J.; Pan, C.; Zhang, H.; Xie, X.; Wen, J.; Xie, X. Intratumoral Microbiome Impacts Immune Infiltrates in Tumor Microenvironment and Predicts Prognosis in Esophageal Squamous Cell Carcinoma Patients. Front. Cell Infect. Microbiol. 2023, 13, 1165790. [Google Scholar] [CrossRef]
- Yamamura, K.; Izumi, D.; Kandimalla, R.; Sonohara, F.; Baba, Y.; Yoshida, N.; Kodera, Y.; Baba, H.; Goel, A. Intratumoral Fusobacterium nucleatum Levels Predict Therapeutic Response to Neoadjuvant Chemotherapy in Esophageal Squamous Cell Carcinoma. Clin. Cancer Res. 2019, 25, 6170–6179. [Google Scholar] [CrossRef]
- Li, Y.; Wei, B.; Xue, X.; Li, H.; Li, J. Microbiome Changes in Esophageal Cancer: Implications for Pathogenesis and Prognosis. Cancer Biol. Med. 2023, 21, 163–174. [Google Scholar] [CrossRef]
- Gillespie, M.R.; Rai, V.; Agrawal, S.; Nandipati, K.C. The Role of Microbiota in the Pathogenesis of Esophageal Adenocarcinoma. Biology 2021, 10, 697. [Google Scholar] [CrossRef]
- Khatoon, J.; Rai, R.P.; Prasad, K.N. Role of Helicobacter pylori in Gastric Cancer: Updates. World J. Gastrointest. Oncol. 2016, 8, 147–158. [Google Scholar] [CrossRef]
- Maleki Kakelar, H.; Barzegari, A.; Dehghani, J.; Hanifian, S.; Saeedi, N.; Barar, J.; Omidi, Y. Pathogenicity of Helicobacter pylori in Cancer Development and Impacts of Vaccination. Gastric Cancer 2019, 22, 23–36. [Google Scholar] [CrossRef]
- Xi, Y.; Zhang, X.L.; Luo, Q.X.; Gan, H.N.; Liu, Y.S.; Shao, S.H.; Mao, X.H. Helicobacter pylori Regulates Stomach Diseases by Activating Cell Pathways and DNA Methylation of Host Cells. Front. Cell Dev. Biol. 2023, 11, 1187638. [Google Scholar] [CrossRef]
- Abate, M.; Vos, E.; Gonen, M.; Janjigian, Y.Y.; Schattner, M.; Laszkowska, M.; Tang, L.; Maron, S.B.; Coit, D.G.; Vardhana, S.; et al. A Novel Microbiome Signature in Gastric Cancer: A Two Independent Cohort Retrospective Analysis. Ann. Surg. 2022, 276, 605–615. [Google Scholar] [CrossRef]
- Peng, R.; Liu, S.; You, W.; Huang, Y.; Hu, C.; Gao, Y.; Jia, X.; Li, G.; Xu, Z.; Chen, Y. Gastric Microbiome Alterations Are Associated with Decreased CD8+ Tissue-Resident Memory T Cells in the Tumor Microenvironment of Gastric Cancer. Cancer Immunol. Res. 2022, 10, 1224–1240. [Google Scholar] [CrossRef]
- Yue, K.; Sheng, D.; Xue, X.; Zhao, L.; Zhao, G.; Jin, C.; Zhang, L. Bidirectional Mediation Effects between Intratumoral Microbiome and Host DNA Methylation Changes Contribute to Stomach Adenocarcinoma. Microbiol. Spectr. 2023, 11, e0090423. [Google Scholar] [CrossRef]
- Ji, J.; Ji, F.; Bayarsaikhan, E. Intratumoral Microbiota in HCC: A New Kid on the Block? Hepatology 2023, 78, 1012–1014. [Google Scholar] [CrossRef]
- Li, S.; Xia, H.; Wang, Z.; Zhang, X.; Song, T.; Li, J.; Xu, L.; Zhang, N.; Fan, S.; Li, Q.; et al. Intratumoral Microbial Heterogeneity Affected Tumor Immune Microenvironment and Determined Clinical Outcome of HBV-Related HCC. Hepatology 2023, 78, 1079–1091. [Google Scholar] [CrossRef]
- Liu, Y.; Kim, E.S.; Guo, H. Hepatitis B Virus-Related Hepatocellular Carcinoma Exhibits Distinct Intratumoral Microbiota and Immune Microenvironment Signatures. J. Med. Virol. 2024, 96, e29485. [Google Scholar] [CrossRef]
- Xue, C.; Gu, X.; Shi, Q.; Ma, X.; Jia, J.; Su, Y.; Bao, Z.; Lu, J.; Li, L. The Interaction between Intratumoral Bacteria and Metabolic Distortion in Hepatocellular Carcinoma. J. Transl. Med. 2024, 22, 237. [Google Scholar] [CrossRef]
- He, Y.; Zhang, Q.; Yu, X.; Zhang, S.; Guo, W. Overview of Microbial Profiles in Human Hepatocellular Carcinoma and Adjacent Nontumor Tissues. J. Transl. Med. 2023, 21, 68. [Google Scholar] [CrossRef]
- Huang, J.H.; Wang, J.; Chai, X.Q.; Li, Z.C.; Jiang, Y.H.; Li, J.; Liu, X.; Fan, J.; Cai, J.B.; Liu, F. The Intratumoral Bacterial Metataxonomic Signature of Hepatocellular Carcinoma. Microbiol. Spectr. 2022, 10, e0098322. [Google Scholar] [CrossRef]
- Sun, L.; Ke, X.; Guan, A.; Jin, B.; Qu, J.; Wang, Y.; Xu, X.; Li, C.; Sun, H.; Xu, H.; et al. Intratumoural Microbiome Can Predict the Prognosis of Hepatocellular Carcinoma after Surgery. Clin. Transl. Med. 2023, 13, e1331. [Google Scholar] [CrossRef]
- Song, Y.; Xiang, Z.; Lu, Z.; Su, R.; Shu, W.; Sui, M.; Wei, X.; Xu, X. Identification of a Brand Intratumor Microbiome Signature for Predicting Prognosis of Hepatocellular Carcinoma. J. Cancer Res. Clin. Oncol. 2023, 149, 11319–11332. [Google Scholar] [CrossRef]
- Chai, X.; Wang, J.; Li, H.; Gao, C.; Li, S.; Wei, C.; Huang, J.; Tian, Y.; Yuan, J.; Lu, J.; et al. Intratumor Microbiome Features Reveal Antitumor Potentials of Intrahepatic Cholangiocarcinoma. Gut Microbes 2023, 15, 2156255. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhang, W.; Zhang, Z.; Sha, G.; Wang, D.; Tang, D. Intratumoral Microbiota: A New Force in Diagnosing and Treating Pancreatic Cancer. Cancer Lett. 2023, 554, 216031. [Google Scholar] [CrossRef]
- Amara, S.; Yang, L.V.; Tiriveedhi, V.; Muzaffar, M. Complex Role of Microbiome in Pancreatic Tumorigenesis: Potential Therapeutic Implications. Cells 2022, 11, 1900. [Google Scholar] [CrossRef]
- Panebianco, C.; Ciardiello, D.; Villani, A.; Maiorano, B.A.; Latiano, T.P.; Maiello, E.; Perri, F.; Pazienza, V. Insights into the Role of Gut and Intratumor Microbiota in Pancreatic Ductal Adenocarcinoma as New Key Players in Preventive, Diagnostic and Therapeutic Perspective. Semin. Cancer Biol. 2022, 86, 997–1007. [Google Scholar] [CrossRef]
- Abe, S.; Masuda, A.; Matsumoto, T.; Inoue, J.; Toyama, H.; Sakai, A.; Kobayashi, T.; Tanaka, T.; Tsujimae, M.; Yamakawa, K.; et al. Impact of Intratumoral Microbiome on Tumor Immunity and Prognosis in Human Pancreatic Ductal Adenocarcinoma. J. Gastroenterol. 2024, 59, 250–262. [Google Scholar] [CrossRef]
- Riquelme, E.; Zhang, Y.; Zhang, L.; Montiel, M.; Zoltan, M.; Dong, W.; Quesada, P.; Sahin, I.; Chandra, V.; San Lucas, A.; et al. Tumor Microbiome Diversity and Composition Influence Pancreatic Cancer Outcomes. Cell 2019, 178, 795–806.e12. [Google Scholar] [CrossRef]
- Pushalkar, S.; Hundeyin, M.; Daley, D.; Zambirinis, C.P.; Kurz, E.; Mishra, A.; Mohan, N.; Aykut, B.; Usyk, M.; Torres, L.E.; et al. The Pancreatic Cancer Microbiome Promotes Oncogenesis by Induction of Innate and Adaptive Immune Suppression. Cancer Discov. 2018, 8, 403–416. [Google Scholar] [CrossRef]
- Alam, A.; Levanduski, E.; Denz, P.; Villavicencio, H.S.; Bhatta, M.; Alhorebi, L.; Zhang, Y.; Gomez, E.C.; Morreale, B.; Senchanthisai, S.; et al. Fungal Mycobiome Drives IL-33 Secretion and Type 2 Immunity in Pancreatic Cancer. Cancer Cell 2022, 40, 153–167.e11. [Google Scholar] [CrossRef]
- Tan, Q.; Ma, X.; Yang, B.; Liu, Y.; Xie, Y.; Wang, X.; Yuan, W.; Ma, J. Periodontitis Pathogen Porphyromonas gingivalis Promotes Pancreatic Tumorigenesis via Neutrophil Elastase from Tumor-Associated Neutrophils. Gut Microbes 2022, 14, 2073785. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Z.; Shen, X.; Xu, J.; Weng, Y.; Wang, W.; Xue, J. Clostridium butyricum and Its Metabolite Butyrate Promote Ferroptosis Susceptibility in Pancreatic Ductal Adenocarcinoma. Cell Oncol. 2023, 46, 1645–1658. [Google Scholar] [CrossRef] [PubMed]
- Pfisterer, N.; Ammer-Herrmenau, C.; Antweiler, K.; Kuffer, S.; Ellenrieder, V.; Neesse, A. Dynamics of Intestinal and Intratumoral Microbiome Signatures in Genetically Engineered Mice and Human Pancreatic Ductal Adenocarcinoma. Pancreatology 2023, 23, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Chakladar, J.; Kuo, S.Z.; Castaneda, G.; Li, W.T.; Gnanasekar, A.; Yu, M.A.; Chang, E.Y.; Wang, X.Q.; Ongkeko, W.M. The Pancreatic Microbiome Is Associated with Carcinogenesis and Worse Prognosis in Males and Smokers. Cancers 2020, 12, 2672. [Google Scholar] [CrossRef] [PubMed]
- Nalluri, H.; Jensen, E.; Staley, C. Role of Biliary Stent and Neoadjuvant Chemotherapy in the Pancreatic Tumor Microbiome. BMC Microbiol. 2021, 21, 280. [Google Scholar] [CrossRef] [PubMed]
- Merali, N.; Chouari, T.; Terroire, J.; Jessel, M.D.; Liu, D.S.K.; Smith, J.H.; Wooldridge, T.; Dhillon, T.; Jimenez, J.I.; Krell, J.; et al. Bile Microbiome Signatures Associated with Pancreatic Ductal Adenocarcinoma Compared to Benign Disease: A UK Pilot Study. Int. J. Mol. Sci. 2023, 24, 16888. [Google Scholar] [CrossRef] [PubMed]
- Kohi, S.; Macgregor-Das, A.; Dbouk, M.; Yoshida, T.; Chuidian, M.; Abe, T.; Borges, M.; Lennon, A.M.; Shin, E.J.; Canto, M.I.; et al. Alterations in the Duodenal Fluid Microbiome of Patients With Pancreatic Cancer. Clin. Gastroenterol. Hepatol. 2022, 20, e196–e227. [Google Scholar] [CrossRef] [PubMed]
- Hozaka, Y.; Oi, H.; Satake, S.; Uchino, Y.; Goto, Y.; Idichi, T.; Tanoue, K.; Yamasaki, Y.; Kawasaki, Y.; Mataki, Y.; et al. Are Intratumoral Microbiota Involved in the Progression of Intraductal Papillary Mucinous Neoplasms of the Pancreas? Surgery 2023, 173, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.S.; Yang, C.Y.; Yeh, C.C.; Lin, R.T.; Chen, C.C.; Bai, L.Y.; Hung, M.C.; Lin, C.C.; Wu, C.Y.; Lin, J.T. Endoscopic Ultrasound-Guided Fine-Needle Biopsy as a Tool for Studying the Intra-Tumoral Microbiome in Pancreatic Ductal Adenocarcinoma: A Pilot Study. Sci. Rep. 2022, 12, 107. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhu, N.; Zheng, X.; Liu, Y.; Lu, H.; Yin, X.; Hao, H.; Tan, Y.; Wang, D.; Hu, H.; et al. Intratumor Microbiome Analysis Identifies Positive Association Between Megasphaera and Survival of Chinese Patients with Pancreatic Ductal Adenocarcinomas. Front. Immunol. 2022, 13, 785422. [Google Scholar] [CrossRef]
- Udayasuryan, B.; Ahmad, R.N.; Nguyen, T.T.D.; Umana, A.; Monet Roberts, L.; Sobol, P.; Jones, S.D.; Munson, J.M.; Slade, D.J.; Verbridge, S.S. Fusobacterium nucleatum Induces Proliferation and Migration in Pancreatic Cancer Cells through Host Autocrine and Paracrine Signaling. Sci. Signal 2022, 15, eabn4948. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, J.; Li, H.; Huang, B.; Zhang, B.; Song, B.; Bao, C.; Liu, Y.; Wang, Z. Integrated Multi-Omics Identified the Novel Intratumor Microbiome-Derived Subtypes and Signature to Predict the Outcome, Tumor Microenvironment Heterogeneity, and Immunotherapy Response for Pancreatic Cancer Patients. Front. Pharmacol. 2023, 14, 1244752. [Google Scholar] [CrossRef] [PubMed]
- Kinskey, J.C.; Huda, T.I.; Gozlan, E.C.; Quach, J.U.; Arturo, J.F.; Chobrutskiy, A.; Chobrutskiy, B.I.; Blanck, G. The Presence of Intratumoral Porphyromonas gingivalis Correlates with a Previously Defined Pancreatic Adenocarcinoma, Immune Cell Expression Phenotype and with Tumor Resident, Adaptive Immune Receptor Features. Carcinogenesis 2023, 44, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Wan, M.L.; Wang, Y.; Zeng, Z.; Deng, B.; Zhu, B.S.; Cao, T.; Li, Y.K.; Xiao, J.; Han, Q.; Wu, Q. Colorectal Cancer (CRC) as a Multifactorial Disease and Its Causal Correlations with Multiple Signaling Pathways. Biosci. Rep. 2020, 40, BSR20200265. [Google Scholar] [CrossRef]
- Heavey, P.M.; McKenna, D.; Rowland, I.R. Colorectal Cancer and the Relationship between Genes and the Environment. Nutr. Cancer 2004, 48, 124–141. [Google Scholar] [CrossRef]
- Li, J.; Zhu, Y.; Yang, L.; Wang, Z. Effect of Gut Microbiota in the Colorectal Cancer and Potential Target Therapy. Discov. Oncol. 2022, 13, 51. [Google Scholar] [CrossRef]
- Zepeda-Rivera, M.; Minot, S.S.; Bouzek, H.; Wu, H.; Blanco-Miguez, A.; Manghi, P.; Jones, D.S.; LaCourse, K.D.; Wu, Y.; McMahon, E.F.; et al. A Distinct Fusobacterium nucleatum Clade Dominates the Colorectal Cancer Niche. Nature 2024, 628, 424–432. [Google Scholar] [CrossRef]
- Leake, I. Genotoxins from Gut Bacteria. Nat. Biotechnol. 2022, 40, 1765. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.T.; Kantilal, H.K.; Davamani, F. The Mechanism of Bacteroides fragilis Toxin Contributes to Colon Cancer Formation. Malays. J. Med. Sci. 2020, 27, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Shang, F.M.; Liu, H.L. Fusobacterium nucleatum and Colorectal Cancer: A Review. World J. Gastrointest. Oncol. 2018, 10, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Liu, X.; Liu, Z.; Pan, C.; Zhang, X.; Zhao, Z.; Sun, H. Fusobacterium nucleatum in Tumors: From Tumorigenesis to Tumor Metastasis and Tumor Resistance. Cancer Biol. Ther. 2024, 25, 2306676. [Google Scholar] [CrossRef]
- Xiang, D.; Yang, J.; Liu, L.; Yu, H.; Gong, X.; Liu, D. The Regulation of Tissue-Specific Farnesoid X Receptor on Genes and Diseases Involved in Bile Acid Homeostasis. Biomed. Pharmacother. 2023, 168, 115606. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Qian, L. Research on Gut Microbiota-Derived Secondary Bile Acids in Cancer Progression. Integr. Cancer Ther. 2022, 21, 15347354221114100. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, Y.; Mima, K.; Ishimoto, T.; Ogata, Y.; Imai, K.; Miyamoto, Y.; Akiyama, T.; Daitoku, N.; Hiyoshi, Y.; Iwatsuki, M.; et al. Relationship between Fusobacterium nucleatum and Antitumor Immunity in Colorectal Cancer Liver Metastasis. Cancer Sci. 2021, 112, 4470–4477. [Google Scholar] [CrossRef] [PubMed]
- Sambruni, G.; Macandog, A.D.; Wirbel, J.; Cagnina, D.; Catozzi, C.; Dallavilla, T.; Borgo, F.; Fazio, N.; Fumagalli-Romario, U.; Petz, W.L.; et al. Location and Condition Based Reconstruction of Colon Cancer Microbiome from Human RNA Sequencing Data. Genome Med. 2023, 15, 32. [Google Scholar] [CrossRef] [PubMed]
- Barot, S.V.; Sangwan, N.; Nair, K.G.; Schmit, S.L.; Xiang, S.; Kamath, S.; Liska, D.; Khorana, A.A. Distinct Intratumoral Microbiome of Young-Onset and Average-Onset Colorectal Cancer. eBioMedicine 2024, 100, 104980. [Google Scholar] [CrossRef]
- Huang, X.; Chen, C.; Xie, W.; Zhou, C.; Tian, X.; Zhang, Z.; Wang, Q.; Chang, H.; Xiao, W.; Zhang, R.; et al. Metagenomic Analysis of Intratumoral Microbiome Linking to Response to Neoadjuvant Chemoradiotherapy in Rectal Cancer. Int. J. Radiat. Oncol. Biol. Phys. 2023, 117, 1255–1269. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, X.; Zhang, H.; Zhang, H.; Yi, Z.; Zhang, Q.; Liu, Q.; Liu, X. Multi-Omics Analysis Reveals Intratumor Microbes as Immunomodulators in Colorectal Cancer. Microbiol. Spectr. 2023, 11, e0503822. [Google Scholar] [CrossRef]
- Sun, L.; Qu, J.; Ke, X.; Zhang, Y.; Xu, H.; Lv, N.; Leng, J.; Zhang, Y.; Guan, A.; Feng, Y.; et al. Interaction between Intratumoral Microbiota and Tumor Mediates the Response of Neoadjuvant Therapy for Rectal Cancer. Front. Microbiol. 2023, 14, 1229888. [Google Scholar] [CrossRef]
- de Oliveira Alves, N.; Dalmasso, G.; Nikitina, D.; Vaysse, A.; Ruez, R.; Ledoux, L.; Pedron, T.; Bergsten, E.; Boulard, O.; Autier, L.; et al. The Colibactin-Producing Escherichia coli Alters the Tumor Microenvironment to Immunosuppressive Lipid Overload Facilitating Colorectal Cancer Progression and Chemoresistance. Gut Microbes 2024, 16, 2320291. [Google Scholar] [CrossRef]
- Okuda, S.; Shimada, Y.; Tajima, Y.; Yuza, K.; Hirose, Y.; Ichikawa, H.; Nagahashi, M.; Sakata, J.; Ling, Y.; Miura, N.; et al. Profiling of Host Genetic Alterations and Intra-Tumor Microbiomes in Colorectal Cancer. Comput. Struct. Biotechnol. J. 2021, 19, 3330–3338. [Google Scholar] [CrossRef]
- Park, H.E.; Kim, J.H.; Cho, N.Y.; Lee, H.S.; Kang, G.H. Intratumoral Fusobacterium nucleatum Abundance Correlates with Macrophage Infiltration and CDKN2A Methylation in Microsatellite-Unstable Colorectal Carcinoma. Virchows Arch. 2017, 471, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.A.; Yoo, S.Y.; Oh, H.J.; Jeong, S.; Cho, N.Y.; Kang, G.H.; Kim, J.H. Differential Immune Microenvironmental Features of Microsatellite-Unstable Colorectal Cancers According to Fusobacterium nucleatum Status. Cancer Immunol. Immunother. 2021, 70, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Kosumi, K.; Hamada, T.; Koh, H.; Borowsky, J.; Bullman, S.; Twombly, T.S.; Nevo, D.; Masugi, Y.; Liu, L.; da Silva, A.; et al. The Amount of Bifidobacterium Genus in Colorectal Carcinoma Tissue in Relation to Tumor Characteristics and Clinical Outcome. Am. J. Pathol. 2018, 188, 2839–2852. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.J.; Kim, J.H.; Bae, J.M.; Kim, H.J.; Cho, N.Y.; Kang, G.H. Prognostic Impact of Fusobacterium nucleatum Depends on Combined Tumor Location and Microsatellite Instability Status in Stage II/III Colorectal Cancers Treated with Adjuvant Chemotherapy. J. Pathol. Transl. Med. 2019, 53, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Proffitt, C.; Bidkhori, G.; Moyes, D.; Shoaie, S. Disease, Drugs and Dysbiosis: Understanding Microbial Signatures in Metabolic Disease and Medical Interventions. Microorganisms 2020, 8, 1381. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Li, R.; Chen, Z.; Li, G.; Liu, B.; Guo, S.; Yue, Q.; Yang, S.; Xie, L.; Zhang, Y.; et al. The Role of the Symbiotic Microecosystem in Cancer: Gut Microbiota, Metabolome, and Host Immunome. Front. Immunol. 2023, 14, 1235827. [Google Scholar] [CrossRef]
- Liu, J.; Luo, F.; Wen, L.; Zhao, Z.; Sun, H. Current Understanding of Microbiomes in Cancer Metastasis. Cancers 2023, 15, 1893. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Mao, Y.; Wang, L. The Crosstalk of Intratumor Bacteria and the Tumor. Front. Cell Infect. Microbiol. 2023, 13, 1273254. [Google Scholar] [CrossRef]
- Ye, L.Y.; Zhang, Q.; Bai, X.L.; Pankaj, P.; Hu, Q.D.; Liang, T.B. Hypoxia-Inducible Factor 1alpha Expression and Its Clinical Significance in Pancreatic Cancer: A Meta-Analysis. Pancreatology 2014, 14, 391–397. [Google Scholar] [CrossRef]
- Tan, Z.; Xu, J.; Zhang, B.; Shi, S.; Yu, X.; Liang, C. Hypoxia: A Barricade to Conquer the Pancreatic Cancer. Cell Mol. Life Sci. 2020, 77, 3077–3083. [Google Scholar] [CrossRef]
- Gnanasekaran, J.; Binder Gallimidi, A.; Saba, E.; Pandi, K.; Eli Berchoer, L.; Hermano, E.; Angabo, S.; Makkawi, H.A.; Khashan, A.; Daoud, A.; et al. Intracellular Porphyromonas gingivalis Promotes the Tumorigenic Behavior of Pancreatic Carcinoma Cells. Cancers 2020, 12, 2331. [Google Scholar] [CrossRef] [PubMed]
- Harada, Y.; Sujino, T.; Miyamoto, K.; Nomura, E.; Yoshimatsu, Y.; Tanemoto, S.; Umeda, S.; Ono, K.; Mikami, Y.; Nakamoto, N.; et al. Intracellular Metabolic Adaptation of Intraepithelial CD4(+)CD8alphaalpha(+) T Lymphocytes. iScience 2022, 25, 104021. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Guo, F.; Lu, S.Y.; Shen, C.; Ma, D.; Zhang, X.; Xie, Y.; Yan, T.; Yu, T.; Sun, T.; et al. F. Nucleatum Targets LncRNA ENO1-IT1 to Promote Glycolysis and Oncogenesis in Colorectal Cancer. Gut 2021, 70, 2123–2137. [Google Scholar] [CrossRef] [PubMed]
- Ou, S.; Wang, H.; Tao, Y.; Luo, K.; Ye, J.; Ran, S.; Guan, Z.; Wang, Y.; Hu, H.; Huang, R. Fusobacterium nucleatum and Colorectal Cancer: From Phenomenon to Mechanism. Front. Cell Infect. Microbiol. 2022, 12, 1020583. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, Q.; Lu, W. Intratumoral Microbiome and Gastrointestinal Cancers. Front. Oncol. 2022, 12, 1047015. [Google Scholar] [CrossRef]
- Sheflin, A.M.; Whitney, A.K.; Weir, T.L. Cancer-Promoting Effects of Microbial Dysbiosis. Curr. Oncol. Rep. 2014, 16, 406. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Du, J.; Chao, S.; Li, S.; Cai, H.; Zhang, H.; Chen, G.; Liu, P.; Bu, P. Fusobacterium nucleatum Promotes Colorectal Cancer Cell to Acquire Stem Cell-Like Features by Manipulating Lipid Droplet-Mediated Numb Degradation. Adv. Sci. 2022, 9, e2105222. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Cai, J.; Hou, W.; Xu, K.; Wu, X.; Song, Y.; Bai, C.; Mo, Y.Y.; Zhang, Z. Microbiome and Spatially Resolved Metabolomics Analysis Reveal the Anticancer Role of Gut Akkermansia Muciniphila by Crosstalk with Intratumoral Microbiota and Reprogramming Tumoral Metabolism in Mice. Gut Microbes 2023, 15, 2166700. [Google Scholar] [CrossRef]
- Pacheco, J.H.L.; Elizondo, G. Interplay between Estrogen, Kynurenine, and AHR Pathways: An Immunosuppressive Axis with Therapeutic Potential for Breast Cancer Treatment. Biochem. Pharmacol. 2023, 217, 115804. [Google Scholar] [CrossRef]
- Dehhaghi, M.; Kazemi Shariat Panahi, H.; Heng, B.; Guillemin, G.J. The Gut Microbiota, Kynurenine Pathway, and Immune System Interaction in the Development of Brain Cancer. Front. Cell Dev. Biol. 2020, 8, 562812. [Google Scholar] [CrossRef]
- Canavese, M.; Wijesundara, D.; Maddern, G.J.; Grubor-Bauk, B.; Hauben, E. Hepatitis C Virus Drives the Pathogenesis of Hepatocellular Carcinoma: From Immune Evasion to Carcinogenesis. Clin. Transl. Immunol. 2016, 5, e101. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Chu, Q.; Zheng, Q.; Yuan, X.; Su, Y.; Bao, Z.; Lu, J.; Li, L. Current Understanding of the Intratumoral Microbiome in Various Tumors. Cell Rep. Med. 2023, 4, 100884. [Google Scholar] [CrossRef] [PubMed]
- Baghban, R.; Roshangar, L.; Jahanban-Esfahlan, R.; Seidi, K.; Ebrahimi-Kalan, A.; Jaymand, M.; Kolahian, S.; Javaheri, T.; Zare, P. Tumor Microenvironment Complexity and Therapeutic Implications at a Glance. Cell Commun. Signal 2020, 18, 59. [Google Scholar] [CrossRef] [PubMed]
- Feng, P.; Xue, X.; Bukhari, I.; Qiu, C.; Li, Y.; Zheng, P.; Mi, Y. Gut Microbiota and Its Therapeutic Implications in Tumor Microenvironment Interactions. Front. Microbiol. 2024, 15, 1287077. [Google Scholar] [CrossRef]
- Antoniewicz, M.R. A Guide to Deciphering Microbial Interactions and Metabolic Fluxes in Microbiome Communities. Curr. Opin. Biotechnol. 2020, 64, 230–237. [Google Scholar] [CrossRef]
- Sieow, B.F.; Nurminen, T.J.; Ling, H.; Chang, M.W. Meta-Omics- and Metabolic Modeling-Assisted Deciphering of Human Microbiota Metabolism. Biotechnol. J. 2019, 14, e1800445. [Google Scholar] [CrossRef] [PubMed]
- Arikan, M.; Muth, T. Integrated Multi-Omics Analyses of Microbial Communities: A Review of the Current State and Future Directions. Mol. Omi. 2023, 19, 607–623. [Google Scholar] [CrossRef]
| Year of Study | Nature of Study | Participants | Aim and Main Findings | Refs. |
|---|---|---|---|---|
| 2019 | Experimental and observational study | PDAC patients; mice | The study aimed to investigate the role of the tumor microbiota and the immune system in influencing long-term survival (LTS) in patients with PDAC. The main findings reveal that patients with long-term survival had higher alpha-diversity in their tumor microbiome. An intra-tumoral microbiome signature (Pseudoxanthomonas-Streptomyces-Saccharopolyspora-Bacillus clausii) was identified as highly predictive of long-term survival. FMT experiments demonstrated that modulating the tumor microbiome could affect tumor growth and immune infiltration. | [57] |
| 2018 | Experimental study | PDAC patients; mice | The study aimed to investigate the role of the microbiome in PDAC and its potential as a therapeutic target. The study revealed that the cancerous pancreas harbors a significantly more abundant microbiome compared to the normal pancreas, with specific bacteria increased in tumorous pancreas compared to the gut. Ablation of the microbiome protects against preinvasive and invasive PDA, while transfer of bacteria from PDA-bearing hosts reverses tumor protection. | [58] |
| 2022 | Experimental study | PDAC cells; mice; PDAC patients | The study aimed to investigate the mechanisms by which type 2 immune cells traffic to the tumor microenvironment in PDAC and identify potential therapeutic targets. Intratumoral fungal mycobiome plays a crucial role in IL-33 secretion, and targeting this pathway shows therapeutic potential in reducing Th2 cells and innate lymphoid cells 2 (ILC2) infiltration and promoting tumor regression in PDAC. | [59] |
| 2022 | Experimental study | PCA patients; mice | The study aimed to investigate the role and mechanism of P. gingivalis in promoting PCA oncogenesis. The study explores a significant association between P. gingivalis and PCA, demonstrating its presence in both oral cavity and tumor tissues of PCA patients. Exposure to P. gingivalis accelerates tumor development in mouse models of PCA, fostering a neutrophil-dominated proinflammatory TME through elevated secretion of neutrophilic chemokines and neutrophil elastase (NE). | [60] |
| 2024 | Observational and experimental study | PDAC patients | The study aimed to elucidate the impact of intratumoral bacteria on the pathophysiology and prognosis of PDAC patients. The study identifies the presence of intratumoral bacteria, particularly anaerobic species such as Bacteroides, Lactobacillus, and Peptoniphilus, in human PDAC tissue, which is associated with suppressed anti-PDAC immunity and poorer prognosis. | [56] |
| 2023 | Experimental study with bioinformatics analysis | PDAC patients (data from the TCGA); mice | The study aimed to explore whether intervention with butyrate-producing probiotics can limit PDAC progression. The study demonstrates that intratumoral butyrate-producing microbiota is associated with favorable outcomes in PDAC. Intervention with Clostridium butyricum or its metabolite butyrate induces superoxidative stress and intracellular lipid accumulation, enhancing susceptibility to ferroptosis and inhibiting PDAC progression. | [61] |
| 2023 | Experimental and observational study | Mice | The study characterized the fecal and intratumoral microbiome of mouse models of PDAC and found significant differences compared to healthy controls. The fecal microbiome of KPC mice resembled that of human PDAC patients, and KPC tumors harbored more bacterial components compared to healthy pancreas tissue. | [62] |
| 2020 | Observational study using bioinformatics analysis | Pancreatic adenocarcinoma patients (data from the TCGA) | The study aimed to characterize the intra-pancreatic microbiome in pancreatic adenocarcinoma and explore its association with prognosis, smoking, and gender. The presence of specific bacterial species within pancreatic adenocarcinoma tumors was correlated with metastasis and immune suppression. Additionally, the study highlights the link between the increased prevalence and poorer prognosis of pancreatic adenocarcinoma in males and smokers with the presence of potentially cancer-promoting or immune-inhibiting microbes, emphasizing the importance of understanding and targeting the pro-TME for therapeutic interventions. | [63] |
| 2021 | Observational study | PDAC patients | This study investigated the role of clinical factors in bacterial colonization within PDAC. Findings reveal that biliary stent placement and neoadjuvant chemotherapy were associated with increased intratumor bacterial colonization, particularly from the Enterobacteriaceae family. | [64] |
| 2023 | Observational study | Patients with obstructive jaundice | This study aimed to define bile microbiome in patients with obstructive jaundice caused by PDAC compared to benign pancreaticobiliary diseases. Using 16S rRNA sequencing, distinct microbial signatures were identified, with PDAC patients exhibiting altered bile microbiome composition characterized by lower abundance of Escherichia and two unclassified genera, and increased abundance of Streptococcus. | [65] |
| 2022 | Case-control study | PDAC, pancreatic cyst(s), and normal pancreata patients | The case-control study aimed to investigate the duodenal fluid microbiome profiles in patients with PDAC, pancreatic cyst(s), and normal pancreata. Patients with PDAC exhibited diminished alpha diversity and enrichment of Bifidobacterium genera compared to control subjects and those with pancreatic cyst(s). | [66] |
| 2023 | Observational study | Intraductal papillary mucinous neoplasms (IPMNs) patients | This study investigated the association between specific microbiota and clinicopathologic characteristics of IPMNs of the pancreas. It found a higher relative abundance of Bacteroidetes and Fusobacteria in invasive IPMNs compared to noninvasive IPMNs, suggesting a potential role of intratumoral microbiota in the progression of IPMNs. | [67] |
| 2022 | Prospective pilot study | PDAC patients | This prospective pilot study demonstrates the feasibility of using endoscopic ultrasound-guided fine-needle biopsy to obtain adequate fresh tumor tissue for intratumoral microbial research in patients with PDAC. The intratumoral microbiome profiles generated from tissues obtained by EUS-FNB were comparable to those obtained by surgical biopsy, suggesting that EUS-FNB could serve as a valid and valuable tool for studying the intratumoral microbiome in both resectable and unresectable PDAC. | [68] |
| 2022 | Comparative study | PDAC patients | The study aimed to investigate the association between intratumor microbiome composition and patient survival, as well as the effect of specific microorganisms on tumor growth inhibition. The composition of the intratumor microbiome differs significantly between long-term and short-term survivors of PDAC in Chinese patients, with differential bacterial composition associated with metabolic pathways in the TME. Additionally, administration of Megasphaera enhances tumor growth inhibition when combined with anti-programmed cell death-1 (anti-PD-1) treatment in mice bearing 4T1 tumors. | [69] |
| 2022 | Experimental study | Normal pancreatic epithelial cells and PDAC cells | The study aimed to explore how F. nucleatum infection influences PDAC progression. F. nucleatum infection induces normal pancreatic epithelial cells and PDAC cells to secrete cytokines promoting tumor progression, including increased proliferation and invasive cell motility. Blocking GM-CSF signaling significantly limits the proliferative gains induced by infection. | [70] |
| 2023 | Observational study | PCA patients | The study aimed to identify prognostic microbes and their impact on PCA. The main findings reveal 26 prognostic genera and two microbiome-related subtypes (Mcluster A and B). Patients in Mcluster B had a worse prognosis and higher TNM stage and pathological grade. Patients in Mcluster A were more likely to benefit from CTLA-4 blockers and various chemotherapeutic agents. Additionally, a microbe-derived model for assessing outcomes showed good predictive performance, and the expression of LAMA3 and LIPH was associated with advanced stage and poor prognosis in PCA. | [71] |
| 2023 | Observational study | Pancreatic adenocarcinoma patients | The study aimed to understand how the presence of P. gingivalis in pancreatic adenocarcinoma affects the tumor microenvironment and immune response. The presence of P. gingivalis in pancreatic adenocarcinoma samples is strongly associated with specific immune cell gene expression patterns, as well as reduced chemical complementarity between P. gingivalis antigen and T-cell receptor sequences recovered from tumor samples. | [72] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bi, X.; Wang, J.; Liu, C. Intratumoral Microbiota: Metabolic Influences and Biomarker Potential in Gastrointestinal Cancer. Biomolecules 2024, 14, 917. https://doi.org/10.3390/biom14080917
Bi X, Wang J, Liu C. Intratumoral Microbiota: Metabolic Influences and Biomarker Potential in Gastrointestinal Cancer. Biomolecules. 2024; 14(8):917. https://doi.org/10.3390/biom14080917
Chicago/Turabian StyleBi, Xueyuan, Jihan Wang, and Cuicui Liu. 2024. "Intratumoral Microbiota: Metabolic Influences and Biomarker Potential in Gastrointestinal Cancer" Biomolecules 14, no. 8: 917. https://doi.org/10.3390/biom14080917
APA StyleBi, X., Wang, J., & Liu, C. (2024). Intratumoral Microbiota: Metabolic Influences and Biomarker Potential in Gastrointestinal Cancer. Biomolecules, 14(8), 917. https://doi.org/10.3390/biom14080917