
Structural Evolution of Primate Glutamate Dehydrogenase 2 as Revealed by In Silico Predictions and Experimentally Determined Structures

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
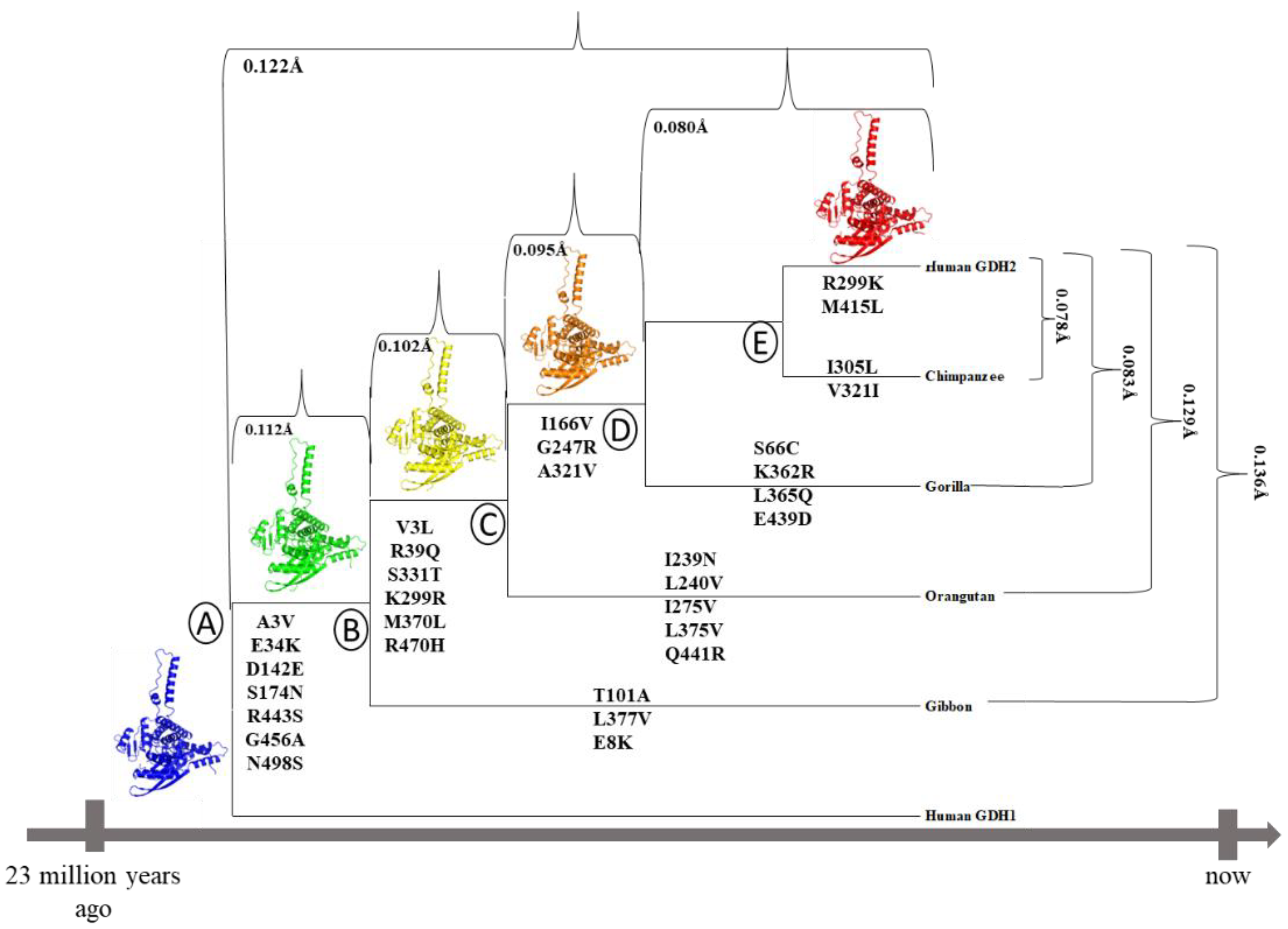
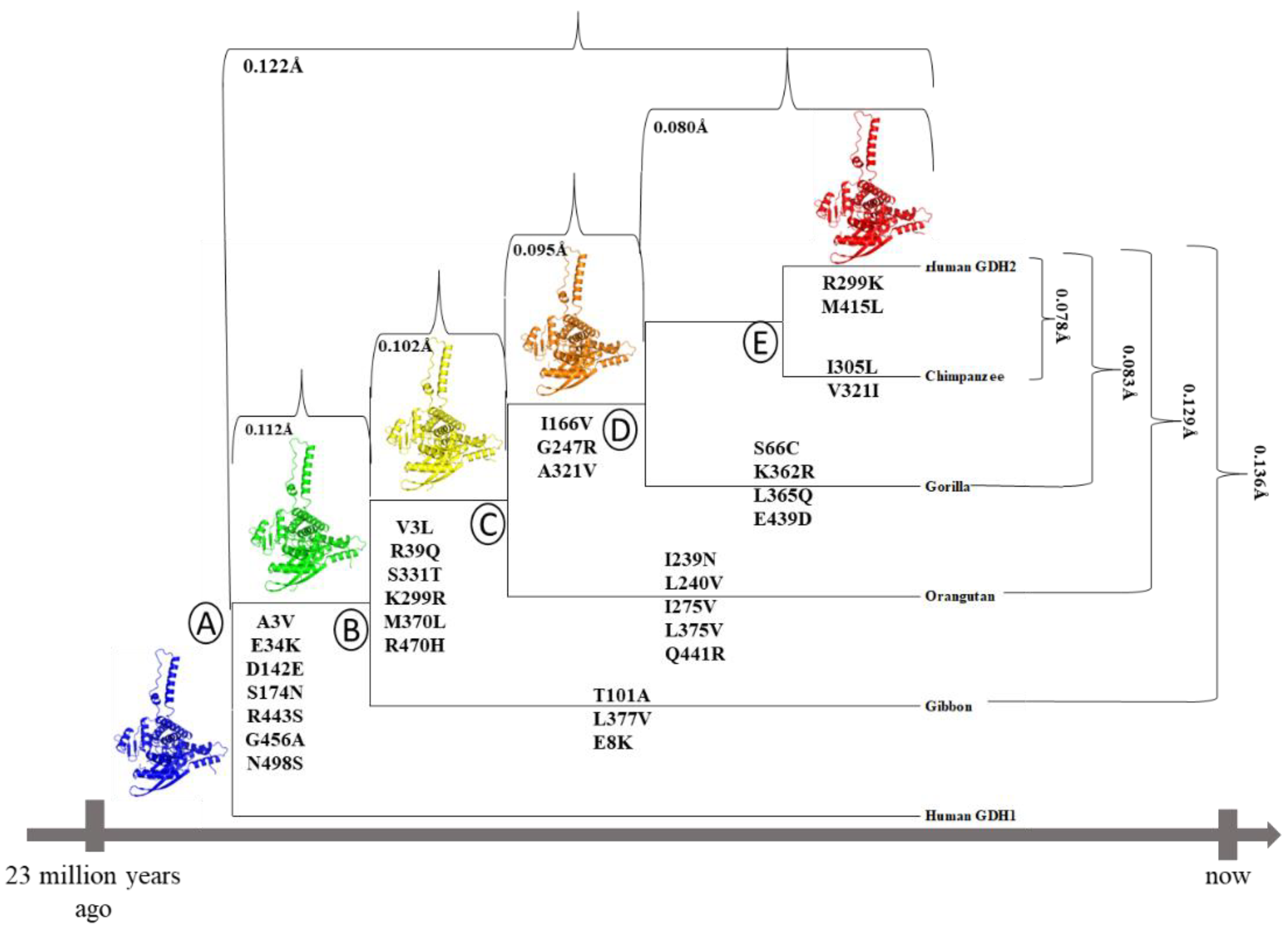
2.1. Phylogenetic Tree Analysis
2.2. Protein Structural Prediction and Analysis
2.3. Mutational Analysis
2.4. Sequence Alignment
3. Results
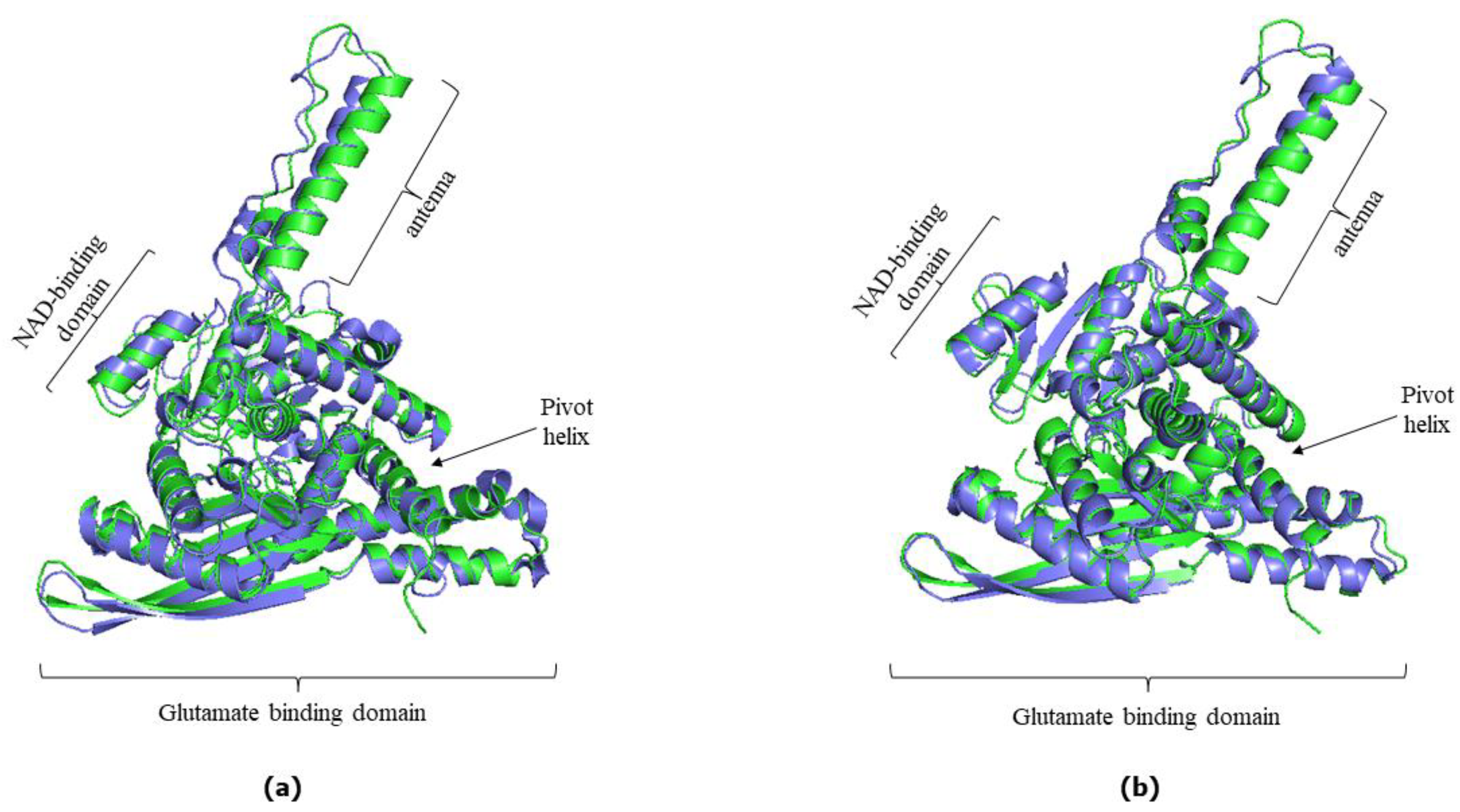
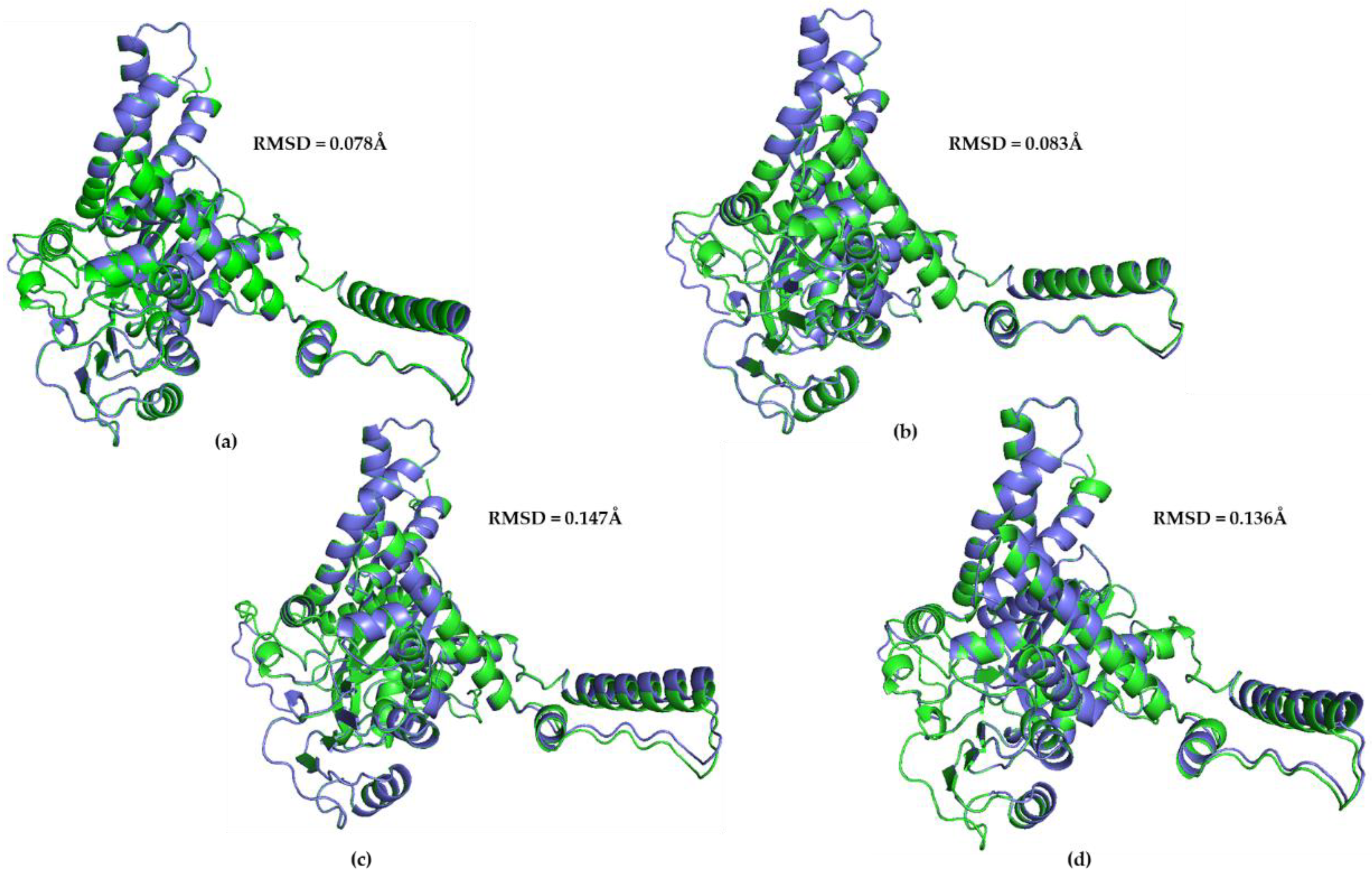
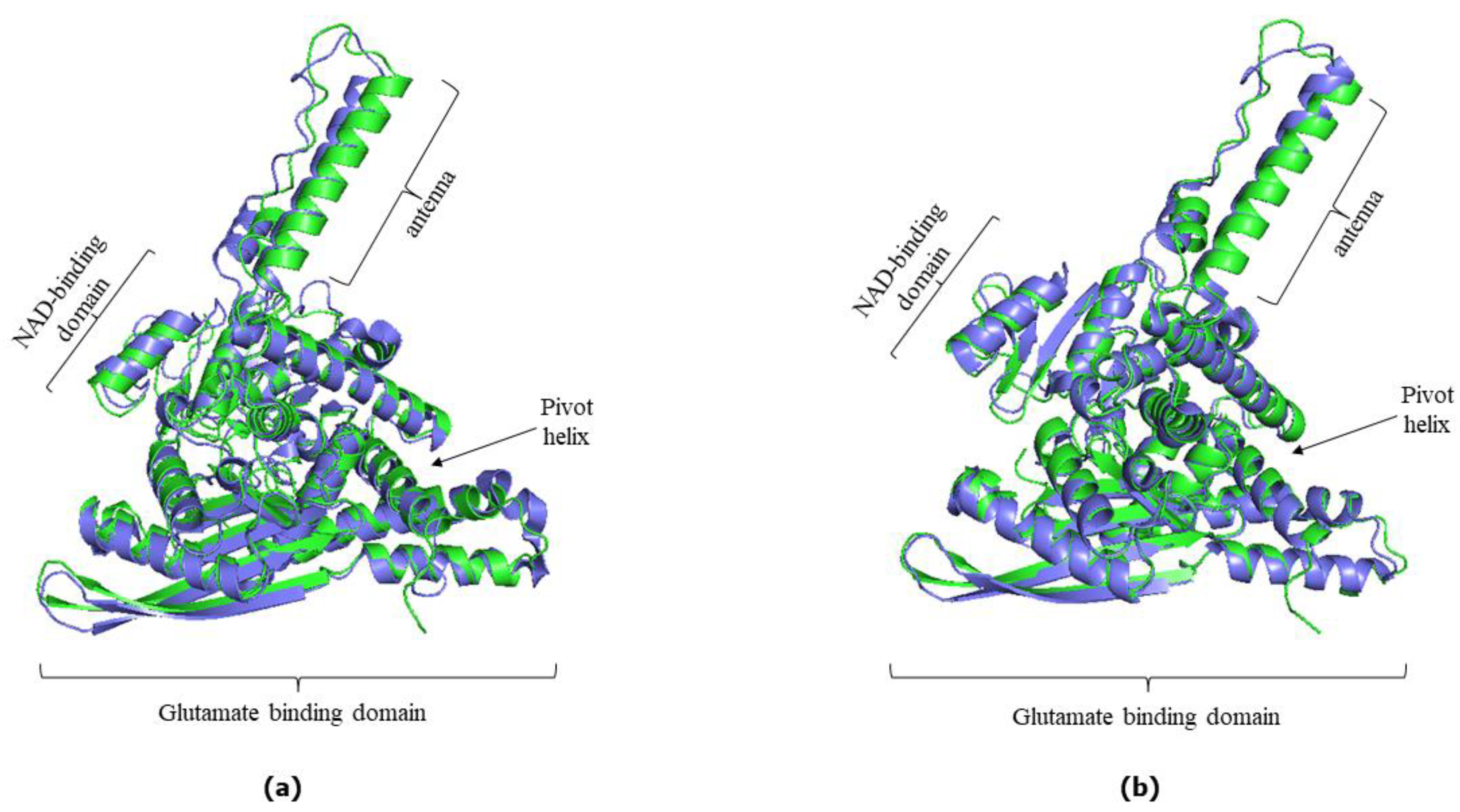
3.1. AlphaFold Predictions vs. Experimentally Determined hGDH1 and hGDH2 Structures
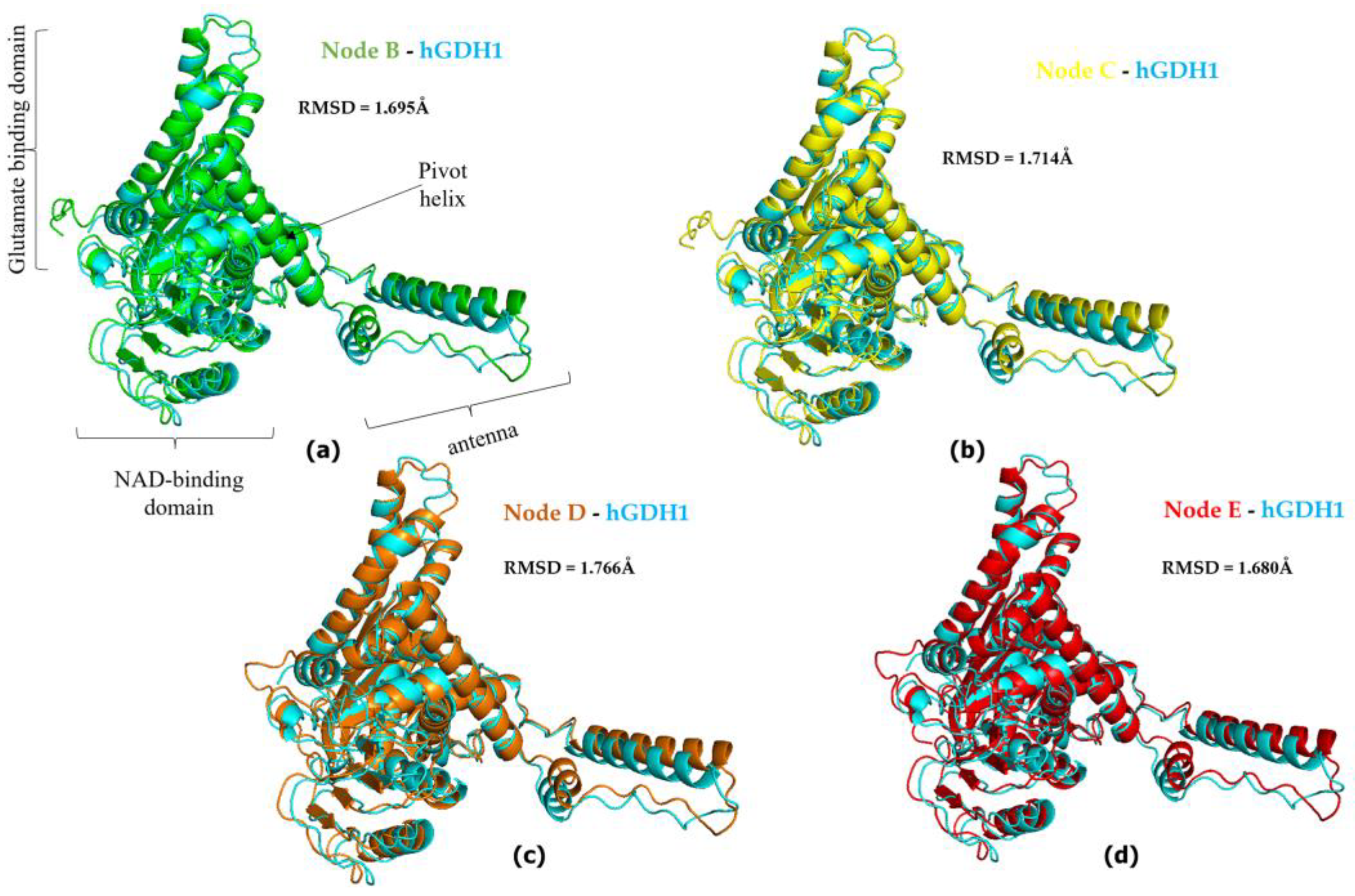
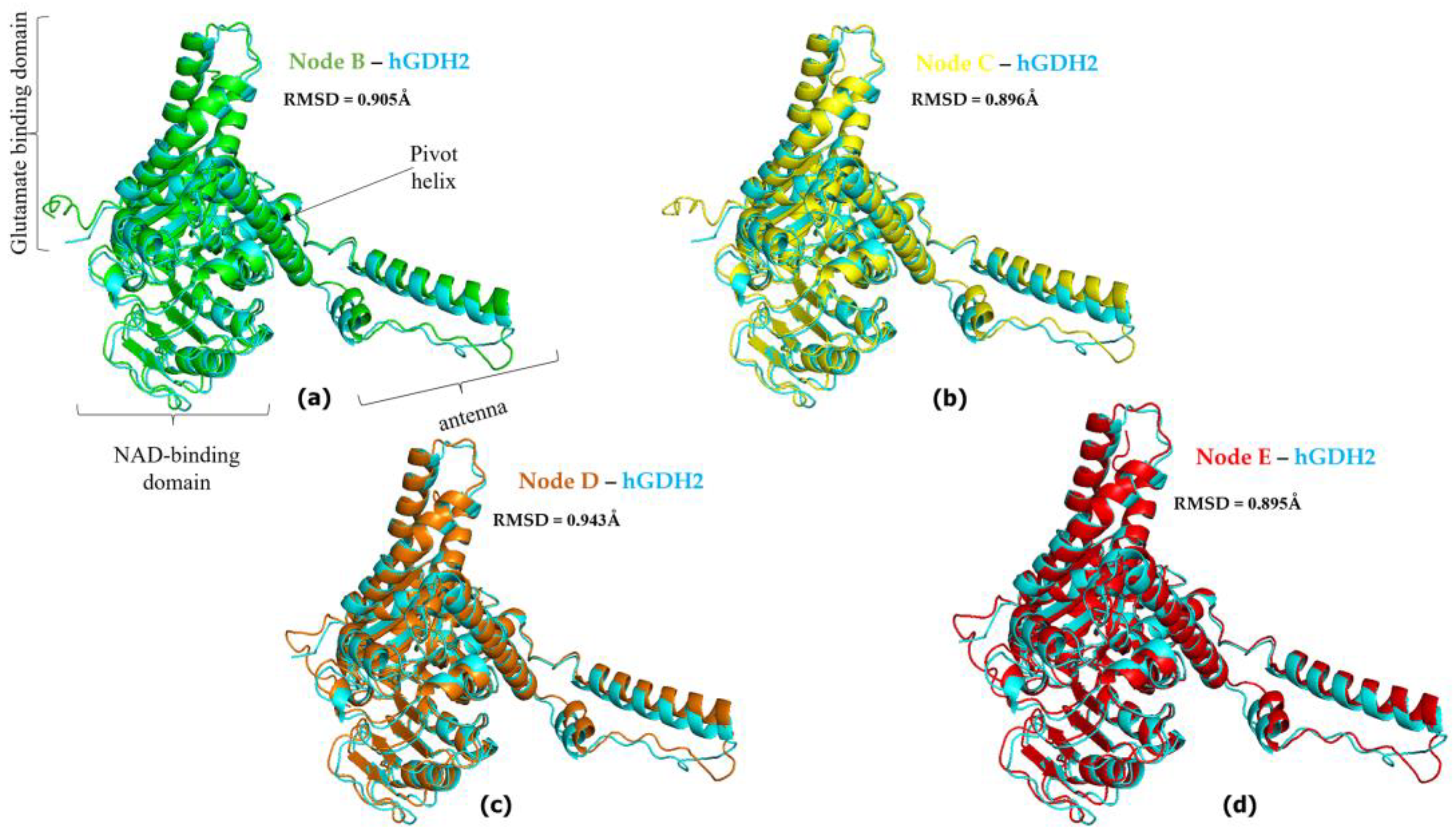
3.2. hGDH2 AlphaFold Colab Predicted Structures during Evolution That Led to Humans
3.3. Ape GDH2 AlphaFold Colab Predicted Structures and Comparison with Predicted hGDH2
3.4. Ape GDH2 AlphaFold Colab Predicted Structures during Evolution
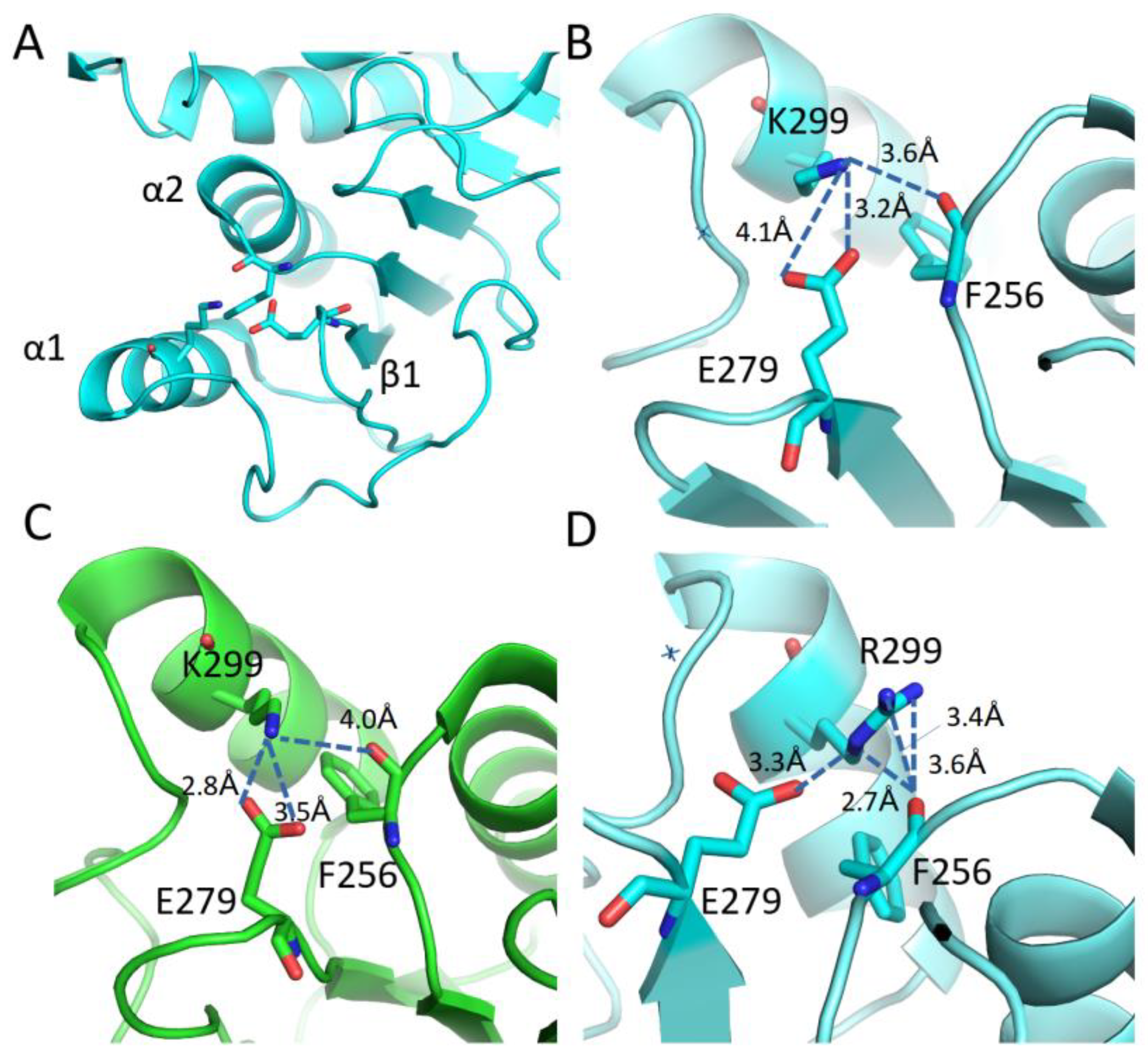
3.5. Mutational and Intramolecular Interactions Analysis
4. Discussion
Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Strecker, H.J. Glutamic Dehydrogenase. Arch. Biochem. Biophys. 1953, 46, 128–140. [Google Scholar] [CrossRef]
- Engel, P.C. Glutamate Dehydrogenases: The Why and How of Coenzyme Specificity. Neurochem. Res. 2014, 39, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Plaitakis, A.; Zaganas, I. Regulation of Human Glutamate Dehydrogenases: Implications for Glutamate, Ammonia and Energy Metabolism in Brain. J. Neurosci. Res. 2001, 66, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.Q.; Li, C.; Stanley, C.A.; Smith, T.J. Glutamate Dehydrogenase, a Complex Enzyme at a Crucial Metabolic Branch Point. Neurochem. Res. 2019, 44, 117–132. [Google Scholar] [CrossRef] [PubMed]
- Hudson, R.C.; Daniel, R.M. L-Glutamate Dehydrogenases: Distribution, Properties and Mechanism. Comp. Biochem. Physiol. B 1993, 106, 767–792. [Google Scholar] [CrossRef]
- Salganicoff, L.; Robertis, E.D. Subcellular Distribution of the Enzymes of the Glutamic Acid, Glutamine and Γ-Aminobutyric Acid Cycles in Rat Brain*. J. Neurochem. 1965, 12, 287–309. [Google Scholar] [CrossRef]
- Lai, J.C.K.; Sheu, K.-F.R.; Kim, Y.T.; Clarke, D.D.; Blass, J.P. The Subcellular Localization of Glutamate Dehydrogenase (GDH): Is GDH a Marker for Mitochondria in Brain? Neurochem. Res. 1986, 11, 733–744. [Google Scholar] [CrossRef]
- Fahien, L.A.; MacDonald, M.J. The Complex Mechanism of Glutamate Dehydrogenase in Insulin Secretion. Diabetes 2011, 60, 2450–2454. [Google Scholar] [CrossRef]
- McKenna, M.C. Glutamate Dehydrogenase in Brain Mitochondria: Do Lipid Modifications and Transient Metabolon Formation Influence Enzyme Activity? Neurochem. Int. 2011, 59, 525–533. [Google Scholar] [CrossRef]
- Shashidharan, P.; Michaelidis, T.M.; Robakis, N.K.; Kresovali, A.; Papamatheakis, J.; Plaitakis, A. Novel Human Glutamate Dehydrogenase Expressed in Neural and Testicular Tissues and Encoded by an X-Linked Intronless Gene. J. Biol. Chem. 1994, 269, 16971–16976. [Google Scholar] [CrossRef]
- Burki, F.; Kaessmann, H. Birth and Adaptive Evolution of a Hominoid Gene That Supports High Neurotransmitter Flux. Nat. Genet. 2004, 36, 1061–1063. [Google Scholar] [CrossRef] [PubMed]
- Varki, A. How to Make an Ape Brain. Nat. Genet. 2004, 36, 1034–1036. [Google Scholar] [CrossRef] [PubMed]
- Darling, A.E.; Miklós, I.; Ragan, M.A. Dynamics of Genome Rearrangement in Bacterial Populations. PLoS Genet. 2008, 4, e1000128. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Guo, S.; Jiang, X.; Bryk, J.; Naumann, R.; Enard, W.; Tomita, M.; Sugimoto, M.; Khaitovich, P.; Pääbo, S. Mice Carrying a Human GLUD2 Gene Recapitulate Aspects of Human Transcriptome and Metabolome Development. Proc. Natl. Acad. Sci. USA 2016, 113, 5358–5363. [Google Scholar] [CrossRef] [PubMed]
- Kanavouras, K.; Mastorodemos, V.; Borompokas, N.; Spanaki, C.; Plaitakis, A. Properties and Molecular Evolution of Human GLUD2 (Neural and Testicular Tissue-Specific) Glutamate Dehydrogenase. J. Neurosci. Res. 2007, 85, 3398–3406. [Google Scholar] [CrossRef] [PubMed]
- Kalef-Ezra, E.; Kotzamani, D.; Zaganas, I.; Katrakili, N.; Plaitakis, A.; Tokatlidis, K. Import of a Major Mitochondrial Enzyme Depends on Synergy between Two Distinct Helices of Its Presequence. Biochem. J. 2016, 473, 2813–2829. [Google Scholar] [CrossRef] [PubMed]
- Zaganas, I.; Spanaki, C.; Karpusas, M.; Plaitakis, A. Substitution of Ser for Arg-443 in the Regulatory Domain of Human Housekeeping (GLUD1) Glutamate Dehydrogenase Virtually Abolishes Basal Activity and Markedly Alters the Activation of the Enzyme by ADP and L-Leucine. J. Biol. Chem. 2002, 277, 46552–46558. [Google Scholar] [CrossRef]
- Zaganas, I.V.; Kanavouras, K.; Borompokas, N.; Arianoglou, G.; Dimovasili, C.; Latsoudis, H.; Vlassi, M.; Mastorodemos, V. The Odyssey of a Young Gene: Structure-Function Studies in Human Glutamate Dehydrogenases Reveal Evolutionary-Acquired Complex Allosteric Regulation Mechanisms. Neurochem. Res. 2014, 39, 471–486. [Google Scholar] [CrossRef]
- Zaganas, I.; Plaitakis, A. Single Amino Acid Substitution (G456A) in the Vicinity of the GTP Binding Domain of Human Housekeeping Glutamate Dehydrogenase Markedly Attenuates GTP Inhibition and Abolishes the Cooperative Behavior of the Enzyme *. J. Biol. Chem. 2002, 277, 26422–26428. [Google Scholar] [CrossRef]
- Stanley, C.A.; Lieu, Y.K.; Hsu, B.Y.; Burlina, A.B.; Greenberg, C.R.; Hopwood, N.J.; Perlman, K.; Rich, B.H.; Zammarchi, E.; Poncz, M. Hyperinsulinism and Hyperammonemia in Infants with Regulatory Mutations of the Glutamate Dehydrogenase Gene. N. Engl. J. Med. 1998, 338, 1352–1357. [Google Scholar] [CrossRef]
- Palladino, A.A.; Stanley, C.A. The Hyperinsulinism/Hyperammonemia Syndrome. Rev. Endocr. Metab. Disord. 2010, 11, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, R.R.; Flanagan, S.E.; Fulton, P.; Chakrapani, A.; Chadefaux, B.; Ben-Omran, T.; Banerjee, I.; Shield, J.P.; Ellard, S.; Hussain, K. Hyperinsulinism–Hyperammonaemia Syndrome: Novel Mutations in the GLUD1 Gene and Genotype–Phenotype Correlations. Eur. J. Endocrinol. 2009, 161, 731–735. [Google Scholar] [CrossRef] [PubMed]
- de Lonlay, P.; Benelli, C.; Fouque, F.; Ganguly, A.; Aral, B.; Dionisi-Vici, C.; Touati, G.; Heinrichs, C.; Rabier, D.; Kamoun, P.; et al. Hyperinsulinism and Hyperammonemia Syndrome: Report of Twelve Unrelated Patients. Pediatr. Res. 2001, 50, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Spanaki, C.; Zaganas, I.; Kleopa, K.A.; Plaitakis, A. Human GLUD2 Glutamate Dehydrogenase Is Expressed in Neural and Testicular Supporting Cells. J. Biol. Chem. 2010, 285, 16748–16756. [Google Scholar] [CrossRef] [PubMed]
- Mathioudakis, L.; Dimovasili, C.; Bourbouli, M.; Latsoudis, H.; Kokosali, E.; Gouna, G.; Vogiatzi, E.; Basta, M.; Kapetanaki, S.; Panagiotakis, S.; et al. Study of Alzheimer’s Disease- and Frontotemporal Dementia-Associated Genes in the Cretan Aging Cohort. Neurobiol. Aging 2022, 123, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, Y.; Nakayama, Y.; Fukusaki, E.; Irino, Y. Glutamate Production from Ammonia via Glutamate Dehydrogenase 2 Activity Supports Cancer Cell Proliferation under Glutamine Depletion. Biochem. Biophys. Res. Commun. 2018, 495, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.J.; Schmidt, T.; Fang, J.; Wu, J.; Siuzdak, G.; Stanley, C.A. The Structure of Apo Human Glutamate Dehydrogenase Details Subunit Communication and Allostery. J. Mol. Biol. 2002, 318, 765–777. [Google Scholar] [CrossRef]
- Smith, T.J.; Peterson, P.E.; Schmidt, T.; Fang, J.; Stanley, C.A. Structures of Bovine Glutamate Dehydrogenase Complexes Elucidate the Mechanism of Purine Regulation. J. Mol. Biol. 2001, 307, 707–720. [Google Scholar] [CrossRef]
- Li, M.; Li, C.; Allen, A.; Stanley, C.A.; Smith, T.J. The Structure and Allosteric Regulation of Mammalian Glutamate Dehydrogenase. Arch. Biochem. Biophys. 2012, 519, 69–80. [Google Scholar] [CrossRef]
- Peterson, P.E.; Pierce, J.; Smith, T.J. Crystallization and Characterization of Bovine Liver Glutamate Dehydrogenase. J. Struct. Biol. 1997, 120, 73–77. [Google Scholar] [CrossRef]
- Peterson, P.E.; Smith, T.J. The Structure of Bovine Glutamate Dehydrogenase Provides Insights into the Mechanism of Allostery. Structure 1999, 7, 769–782. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Schmidt, T.; Fang, J.; Stanley, C.A.; Smith, T.J. Structural Studies on ADP Activation of Mammalian Glutamate Dehydrogenase and the Evolution of Regulation. Biochemistry 2003, 42, 3446–3456. [Google Scholar] [CrossRef] [PubMed]
- Grzechowiak, M.; Sliwiak, J.; Jaskolski, M.; Ruszkowski, M. Structural Studies of Glutamate Dehydrogenase (Isoform 1) From Arabidopsis Thaliana, an Important Enzyme at the Branch-Point Between Carbon and Nitrogen Metabolism. Front. Plant Sci. 2020, 11, 754. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Wang, W.; Zeng, X.; Liu, M.; Li, M.; Li, C.; Wang, M. Crystal Structure of Glutamate Dehydrogenase 3 from Candida Albicans. Biochem. Biophys. Res. Commun. 2021, 570, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Britton, K.L.; Yip, K.S.; Sedelnikova, S.E.; Stillman, T.J.; Adams, M.W.; Ma, K.; Maeder, D.L.; Robb, F.T.; Tolliday, N.; Vetriani, C.; et al. Structure Determination of the Glutamate Dehydrogenase from the Hyperthermophile Thermococcus Litoralis and Its Comparison with That from Pyrococcus Furiosus. J. Mol. Biol. 1999, 293, 1121–1132. [Google Scholar] [CrossRef]
- Yip, K.S.; Stillman, T.J.; Britton, K.L.; Artymiuk, P.J.; Baker, P.J.; Sedelnikova, S.E.; Engel, P.C.; Pasquo, A.; Chiaraluce, R.; Consalvi, V. The Structure of Pyrococcus Furiosus Glutamate Dehydrogenase Reveals a Key Role for Ion-Pair Networks in Maintaining Enzyme Stability at Extreme Temperatures. Structure 1995, 3, 1147–1158. [Google Scholar] [CrossRef]
- Knapp, S.; de Vos, W.M.; Rice, D.; Ladenstein, R. Crystal Structure of Glutamate Dehydrogenase from the Hyperthermophilic Eubacterium Thermotoga Maritima at 3.0 A Resolution. J. Mol. Biol. 1997, 267, 916–932. [Google Scholar] [CrossRef]
- Li, M.; Li, C.; Allen, A.; Stanley, C.A.; Smith, T.J. Glutamate Dehydrogenase: Structure, Allosteric Regulation, and Role in Insulin Homeostasis. Neurochem. Res. 2014, 39, 433–445. [Google Scholar] [CrossRef]
- Dimovasili, C.; Fadouloglou, V.E.; Kefala, A.; Providaki, M.; Kotsifaki, D.; Kanavouras, K.; Sarrou, I.; Plaitakis, A.; Zaganas, I.; Kokkinidis, M. Crystal Structure of Glutamate Dehydrogenase 2, a Positively Selected Novel Human Enzyme Involved in Brain Biology and Cancer Pathophysiology. J. Neurochem. 2021, 157, 802–815. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Senior, A.W.; Evans, R.; Jumper, J.; Kirkpatrick, J.; Sifre, L.; Green, T.; Qin, C.; Žídek, A.; Nelson, A.W.R.; Bridgland, A.; et al. Improved Protein Structure Prediction Using Potentials from Deep Learning. Nature 2020, 577, 706–710. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Tamura, K.; Nei, M. MEGA3: Integrated Software for Molecular Evolutionary Genetics Analysis and Sequence Alignment. Brief. Bioinform. 2004, 5, 150–163. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-W.; Lin, J.; Chu, Y.-W. iStable: Off-the-Shelf Predictor Integration for Predicting Protein Stability Changes. BMC Bioinform. 2013, 14 (Suppl. 2), S5. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Lu, H.; Zhang, N.; Zhu, Z.; Wang, S.; Li, M. PremPS: Predicting the Impact of Missense Mutations on Protein Stability. PLoS Comput. Biol. 2020, 16, e1008543. [Google Scholar] [CrossRef] [PubMed]
- Laimer, J.; Hiebl-Flach, J.; Lengauer, D.; Lackner, P. MAESTROweb: A Web Server for Structure-Based Protein Stability Prediction. Bioinformatics 2016, 32, 1414–1416. [Google Scholar] [CrossRef] [PubMed]
- Pandurangan, A.P.; Ochoa-Montaño, B.; Ascher, D.B.; Blundell, T.L. SDM: A Server for Predicting Effects of Mutations on Protein Stability. Nucleic Acids Res. 2017, 45, W229–W235. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.H.; Pires, D.E.; Ascher, D.B. DynaMut: Predicting the Impact of Mutations on Protein Conformation, Flexibility and Stability. Nucleic Acids Res. 2018, 46, W350–W355. [Google Scholar] [CrossRef]
- Grant, B.J.; Rodrigues, A.P.C.; ElSawy, K.M.; McCammon, J.A.; Caves, L.S.D. Bio3d: An R Package for the Comparative Analysis of Protein Structures. Bioinformatics 2006, 22, 2695–2696. [Google Scholar] [CrossRef]
- Frappier, V.; Najmanovich, R.J. A Coarse-Grained Elastic Network Atom Contact Model and Its Use in the Simulation of Protein Dynamics and the Prediction of the Effect of Mutations. PLoS Comput. Biol. 2014, 10, e1003569. [Google Scholar] [CrossRef]
- McKenna, M.C.; Stridh, M.H.; McNair, L.F.; Sonnewald, U.; Waagepetersen, H.S.; Schousboe, A. Glutamate Oxidation in Astrocytes: Roles of Glutamate Dehydrogenase and Aminotransferases. J. Neurosci. Res. 2016, 94, 1561–1571. [Google Scholar] [CrossRef]
- Burley, S.K.; Arap, W.; Pasqualini, R. Predicting Proteome-Scale Protein Structure with Artificial Intelligence. N. Engl. J. Med. 2021, 385, 2191–2194. [Google Scholar] [CrossRef] [PubMed]
- Tunyasuvunakool, K.; Adler, J.; Wu, Z.; Green, T.; Zielinski, M.; Žídek, A.; Bridgland, A.; Cowie, A.; Meyer, C.; Laydon, A.; et al. Highly Accurate Protein Structure Prediction for the Human Proteome. Nature 2021, 596, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Conery, J.S. The Evolutionary Fate and Consequences of Duplicate Genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, L.S.; Pessoa, D.M.A.; Mountford, J.K.; Davies, W.I.L.; Hunt, D.M. The Genetic and Evolutionary Drives behind Primate Color Vision. Front. Ecol. Evol. 2017, 5, 34. [Google Scholar] [CrossRef]
- Glazko, G.; Veeramachaneni, V.; Nei, M.; Makałowski, W. Eighty Percent of Proteins Are Different between Humans and Chimpanzees. Gene 2005, 346, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Khaitovich, P.; Hellmann, I.; Enard, W.; Nowick, K.; Leinweber, M.; Franz, H.; Weiss, G.; Lachmann, M.; Pääbo, S. Parallel Patterns of Evolution in the Genomes and Transcriptomes of Humans and Chimpanzees. Science 2005, 309, 1850–1854. [Google Scholar] [CrossRef] [PubMed]
- Janiak, M.C. Digestive Enzymes of Human and Nonhuman Primates: Digestive Enzymes of Human and Nonhuman Primates. Evol. Anthropol. 2016, 25, 253–266. [Google Scholar] [CrossRef]
- Gröhs Ferrareze, P.A.; Zimerman, R.A.; Franceschi, V.B.; Caldana, G.D.; Netz, P.A.; Thompson, C.E. Molecular Evolution and Structural Analyses of the Spike Glycoprotein from Brazilian SARS-CoV-2 Genomes: The Impact of Selected Mutations. J. Biomol. Struct. Dyn. 2022, 41, 3110–3128. [Google Scholar] [CrossRef]
- Vlassi, M.; Cesareni, G.; Kokkinidis, M. A Correlation between the Loss of Hydrophobic Core Packing Interactions and Protein Stability. J. Mol. Biol. 1999, 285, 817–827. [Google Scholar] [CrossRef]
- Choi, M.-M.; Kim, E.-A.; Yang, S.-J.; Choi, S.Y.; Cho, S.-W.; Huh, J.-W. Amino Acid Changes within Antenna Helix Are Responsible for Different Regulatory Preferences of Human Glutamate Dehydrogenase Isozymes *. J. Biol. Chem. 2007, 282, 19510–19517. [Google Scholar] [CrossRef]
- Choi, M.-M.; Hwang, E.Y.; Kim, E.-A.; Huh, J.-W.; Cho, S.-W. Identification of Amino Acid Residues Responsible for Different GTP Preferences of Human Glutamate Dehydrogenase Isozymes. Biochem. Biophys. Res. Commun. 2008, 368, 742–747. [Google Scholar] [CrossRef] [PubMed]
- Kotzamani, D.; Plaitakis, A. Alpha Helical Structures in the Leader Sequence of Human GLUD2 Glutamate Dehydrogenase Responsible for Mitochondrial Import. Neurochem. Int. 2012, 61, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Beniashvili, D.S. An Overview of the World Literature on Spontaneous Tumors in Nonhuman Primates. J. Med. Primatol. 1989, 18, 423–437. [Google Scholar] [CrossRef] [PubMed]
- Toledano, A.; Álvarez, M.I.; López-Rodríguez, A.B.; Toledano-Díaz, A.; Fernández-Verdecia, C.I. [Does Alzheimer’s disease exist in all primates? Alzheimer pathology in non-human primates and its pathophysiological implications (II)]. Neurologia 2014, 29, 42–55. [Google Scholar] [CrossRef]
- Kim, A.Y.; Baik, E.J. Glutamate Dehydrogenase as a Neuroprotective Target Against Neurodegeneration. Neurochem. Res. 2019, 44, 147–153. [Google Scholar] [CrossRef]
- Kim, A.Y.; Jeong, K.-H.; Lee, J.H.; Kang, Y.; Lee, S.H.; Baik, E.J. Glutamate Dehydrogenase as a Neuroprotective Target against Brain Ischemia and Reperfusion. Neuroscience 2017, 340, 487–500. [Google Scholar] [CrossRef]
- Li, M.; Allen, A.; Smith, T.J. High Throughput Screening Reveals Several New Classes of Glutamate Dehydrogenase Inhibitors. Biochemistry 2007, 46, 15089–15102. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
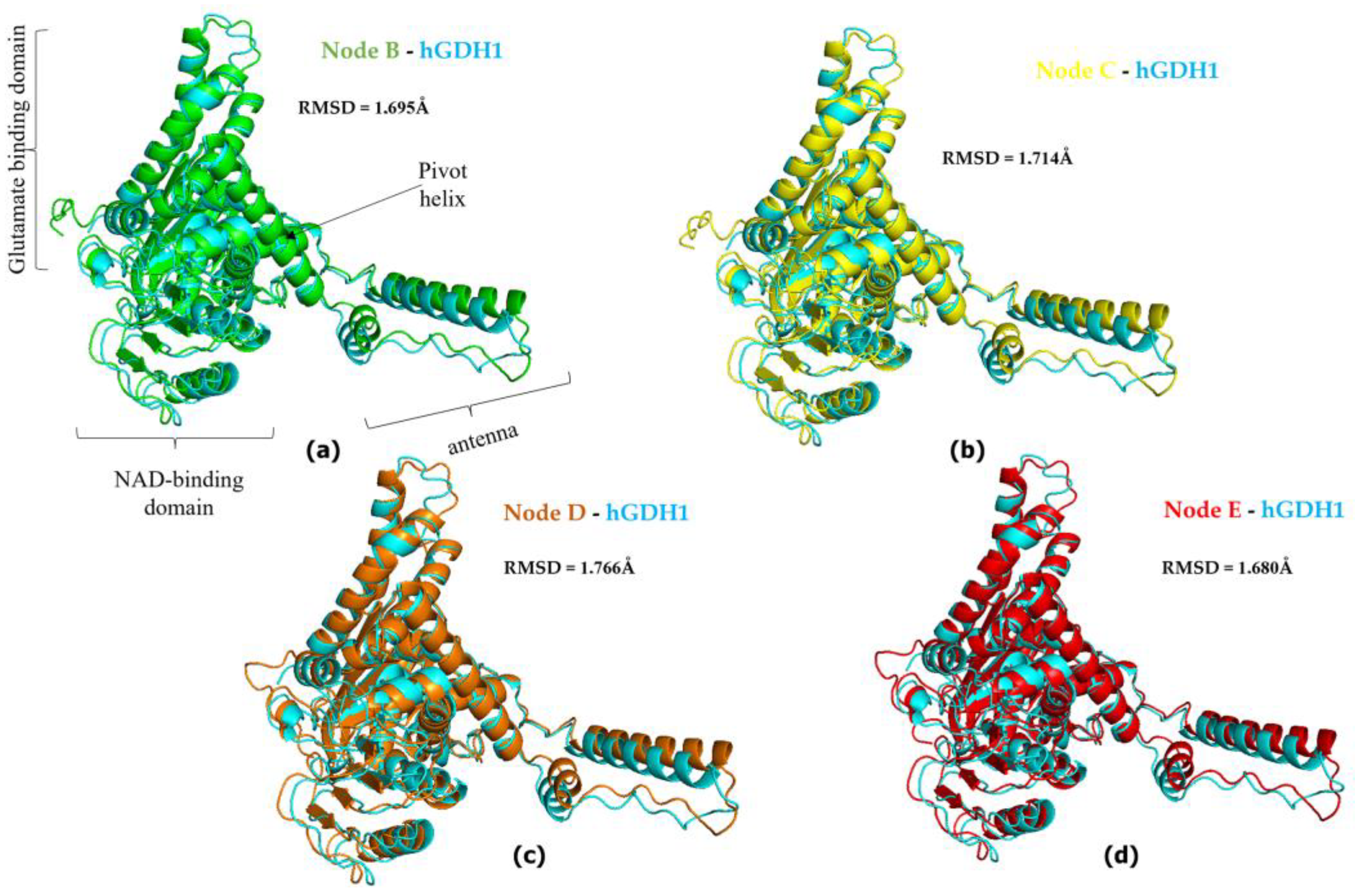{kind=link}
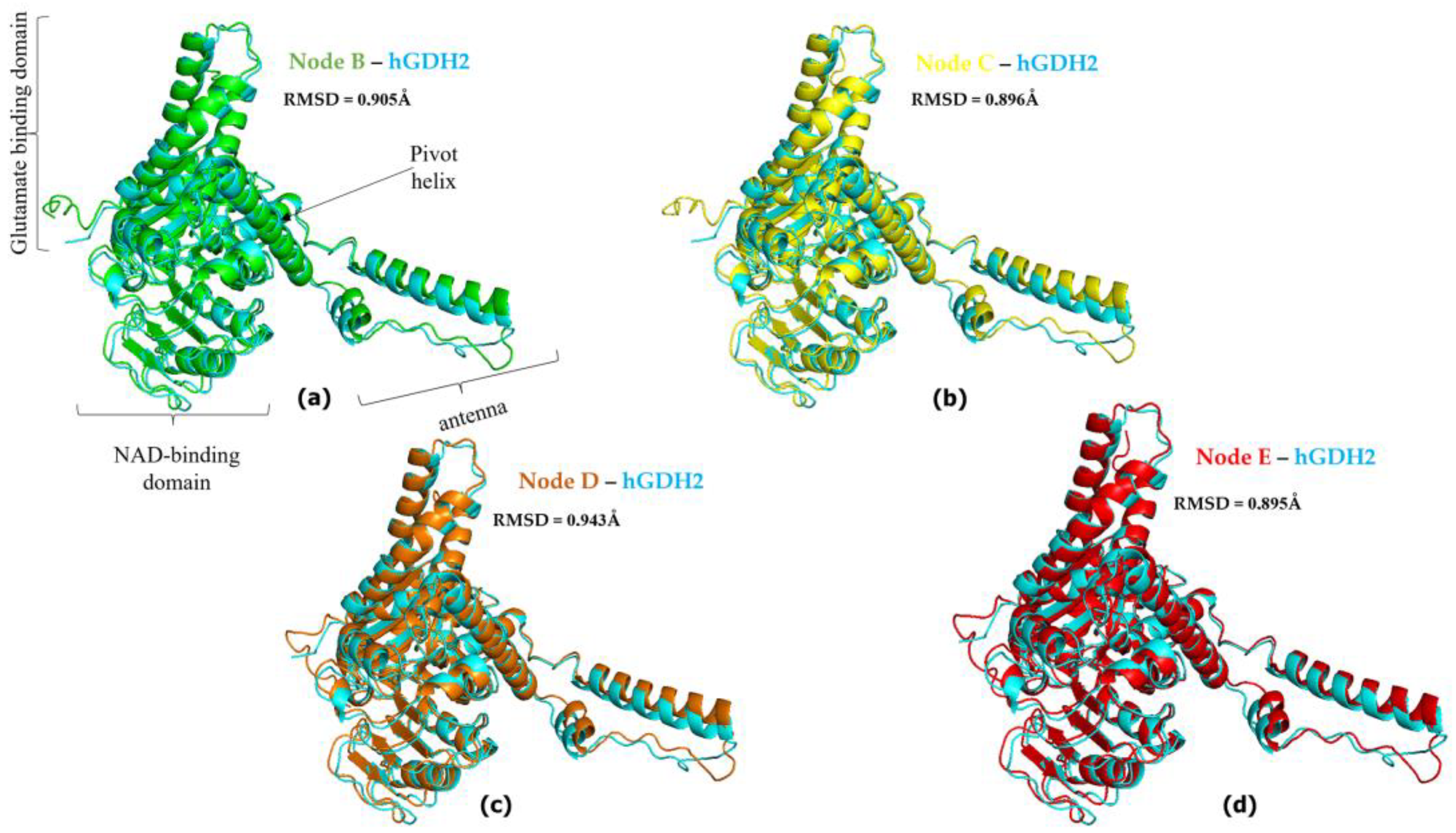{kind=link}
{kind=link}
{kind=link}
| Protein | AlphaFold pLDDT |
|---|---|
| Node A (=hGDH1) | 93.79 |
| Node B | 93.38 |
| Node C | 93.80 |
| Node D | 93.36 |
| Node E (=hGDH2) | 93.85 |
| Chimpanzee (Node E) | 93.67 |
| Gorilla (Node D) | 93.52 |
| Orangutan (Node C) | 93.86 |
| Gibbon (Node B) | 93.44 |
| DynaMut | iSTABLE | PremPS | MaestroWEB | SDM | ||
|---|---|---|---|---|---|---|
| Human Node A-B | A3V | S | D | S | S | S |
| Human Node A-B | E34K | S | D | D | D | D |
| Human Node A-B | D142E | S | S | S | S | S |
| Human Node A-B | S174N | S | S | D | S | S |
| Human Node A-B | R443S | D | D | D | S | D |
| Human Node A-B | G456A | S | S | D | S | S |
| Human Node A-B | N498S | D | D | D | S | D |
| Human Node B-C | V3L | S | D | S | D | D |
| Human Node B-C | R39Q | S | D | D | S | D |
| Human Node B-C | K299R | D | D | D | S | S |
| Human Node B-C | S331T | S | D | D | D | S |
| Human Node B-C | M370L | S | D | S | S | S |
| Human Node B-C | R470H | S | D | D | S | D |
| Human Node C-D | I166V | D | D | D | S | S |
| Human Node C-D | G247R | S | S | D | D | D |
| Human Node C-D | A321V | D | S | S | S | D |
| Human Node D-E | R299K | D | D | S | S | D |
| Human Node D-E | M415L | D | D | D | D | S |
| Chimpanzee Node E | I305L | D | D | S | D | S |
| Chimpanzee Node E | V321I | S | D | D | S | D |
| Gorilla Node D | S66C | D | D | D | S | S |
| Gorilla Node D | K362R | D | S | D | S | S |
| Gorilla Node D | L365Q | D | D | D | S | S |
| Gorilla Node D | E439D | D | S | D | S | D |
| Orangutan Node C | I239N | D | D | D | S | D |
| Orangutan Node C | L240V | D | D | S | S | D |
| Orangutan Node C | I275V | D | D | D | D | D |
| Orangutan Node C | L375V | D | D | D | S | D |
| Orangutan Node C | Q441R | D | S | D | D | D |
| Gibbon Node B | E8K | S | D | S | D | D |
| Gibbon Node B | T101A | D | D | D | D | S |
| Gibbon Node B | L377V | D | D | D | S | D |
| Evolutionary Step | Amino Acid Substitutions | Bonds Lost | Bonds Gained | Interactions Lost | Interactions Gained |
|---|---|---|---|---|---|
| Node A-B | A3V | Ser1 | Ser1, Ala5 | ||
| Node A-B | E34K | Lys31 | Lys31, Asp30 | Leu32 | |
| Node A-B | D142E | Gln144, Glu146 | Arg178, Gln146, Arg178 | Trp182 | |
| Node A-B | S174N | Tyr99 | Tyr99 | Pro137 | |
| Node A-B | R443S | Ala447, Phe440, Glu439 | Phe440, Ala447 | Gln441, Ser445 | |
| Node A-B | G456A | His454, Tyr459, Thr460 | Val453, His454, Tys459, Ile452 | Phe387 | |
| Node A-B | N498S | Gly501, Ala500, Phe494 | Val496, Phe494, Ile52 | ||
| Node B-C | V3L | Ser1 | |||
| Node B-C | R39Q | ||||
| Node B-C | K299R | Glu296, His302, Gln301, Glu296 | Leu295, Glu296, | Phe256 | Phe256, Leu295, Ile305 |
| Node B-C | S331T | Lost: Gln334 | |||
| Node B-C | M370L | Ile347 | Ile347 | Ile347, Phe230, Met237 | Tyr236, Leu479, Leu481 |
| Node B-C | R470H | Met473, Ala472 | |||
| Node C-D | I166V | Pro92 | Gly163, Gly160, Ile162 | ||
| Node C-D | G247R | Lys249 | |||
| Node C-D | A321V | Ile318 | Cys323, Lys344 | Tyr314, Ile318, Val252, Cys323 | |
| Node D-E | R299K | His302, Glu296, Gln301, Asp297 | Glu279, Ile305 | Gln301, Phe256 | |
| Node D-E | M415L | Gln418, His412, Val417 | Val417 | Leu413 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Litso, I.; Plaitakis, A.; Fadouloglou, V.E.; Providaki, M.; Kokkinidis, M.; Zaganas, I. Structural Evolution of Primate Glutamate Dehydrogenase 2 as Revealed by In Silico Predictions and Experimentally Determined Structures. Biomolecules 2024, 14, 22. https://doi.org/10.3390/biom14010022
Litso I, Plaitakis A, Fadouloglou VE, Providaki M, Kokkinidis M, Zaganas I. Structural Evolution of Primate Glutamate Dehydrogenase 2 as Revealed by In Silico Predictions and Experimentally Determined Structures. Biomolecules. 2024; 14(1):22. https://doi.org/10.3390/biom14010022
Chicago/Turabian StyleLitso, Ionela, Andreas Plaitakis, Vasiliki E. Fadouloglou, Mary Providaki, Michael Kokkinidis, and Ioannis Zaganas. 2024. "Structural Evolution of Primate Glutamate Dehydrogenase 2 as Revealed by In Silico Predictions and Experimentally Determined Structures" Biomolecules 14, no. 1: 22. https://doi.org/10.3390/biom14010022
APA StyleLitso, I., Plaitakis, A., Fadouloglou, V. E., Providaki, M., Kokkinidis, M., & Zaganas, I. (2024). Structural Evolution of Primate Glutamate Dehydrogenase 2 as Revealed by In Silico Predictions and Experimentally Determined Structures. Biomolecules, 14(1), 22. https://doi.org/10.3390/biom14010022