

The Extracellular Vesicles Proteome of Endometrial Cells Simulating the Receptive Menstrual Phase Differs from That of Endometrial Cells Simulating the Non-Receptive Menstrual Phase

 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Differential Centrifugation
2.3. Size Exclusion Chromatography
2.4. Nanoparticle Tracking Analysis
2.5. RL95-2 Cell Conditioned Media EV Isolation Optimisation
2.6. Hormone Treatment of RL95-2 Cells
2.7. Transmission Electron Microscopy (TEM)
2.8. Proteomic Analysi—Label-Free Liquid Chromatography Tandem Mass Spectrometry (LC-MS/MS)
2.9. Proteomic Data Analysis
2.10. Statistical Analysis
3. Results
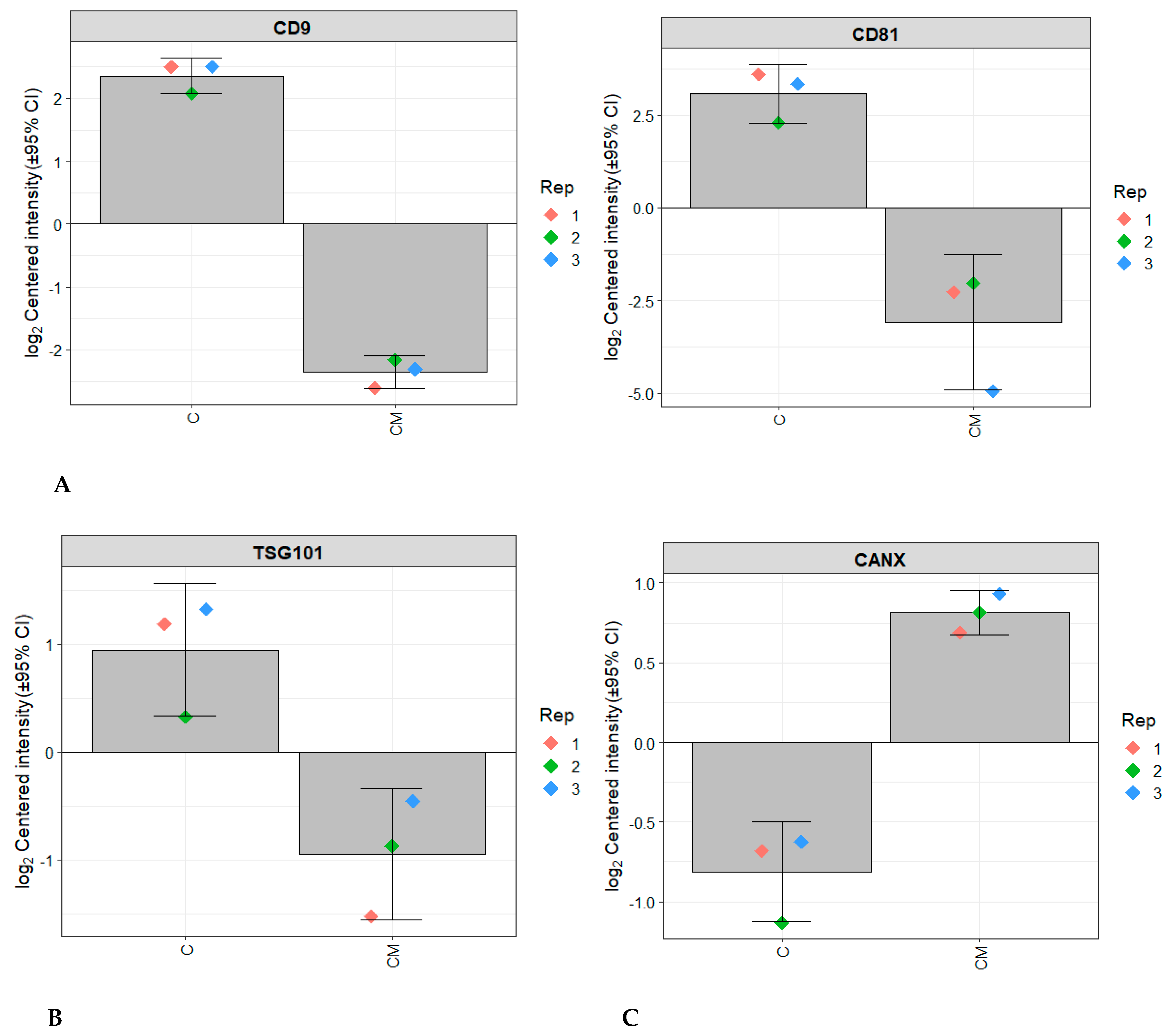
3.1. Characterisation of EVs Released from RL95-2 Cells
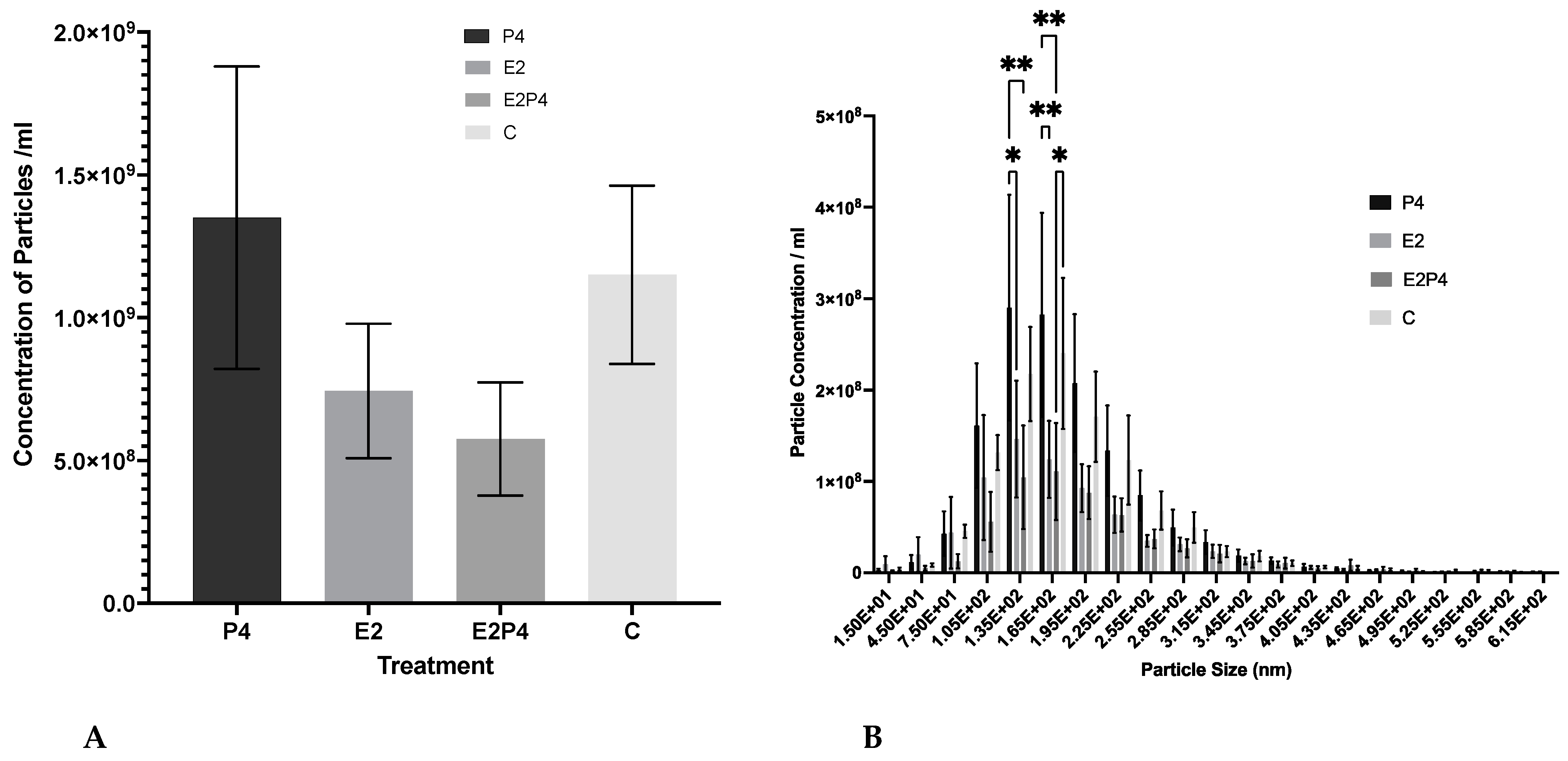
3.2. Concentration and Size Distribution of EVs Released from Hormone-Treated RL95-2 Cells
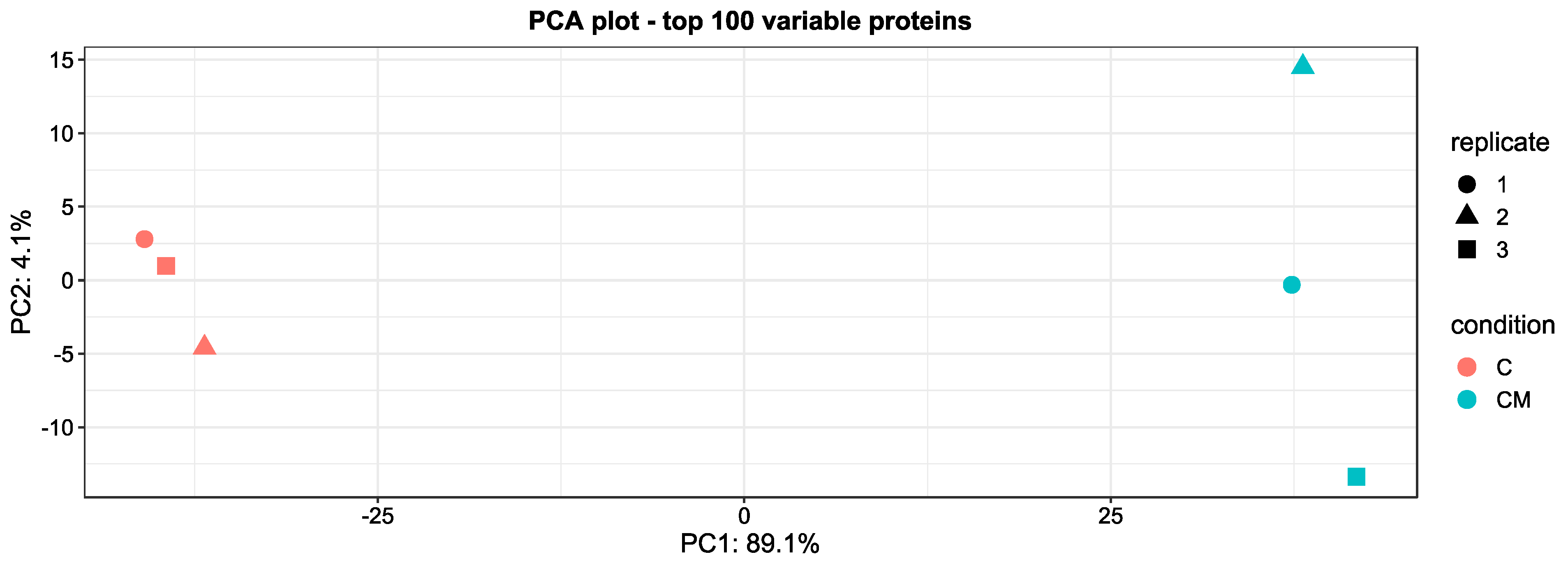
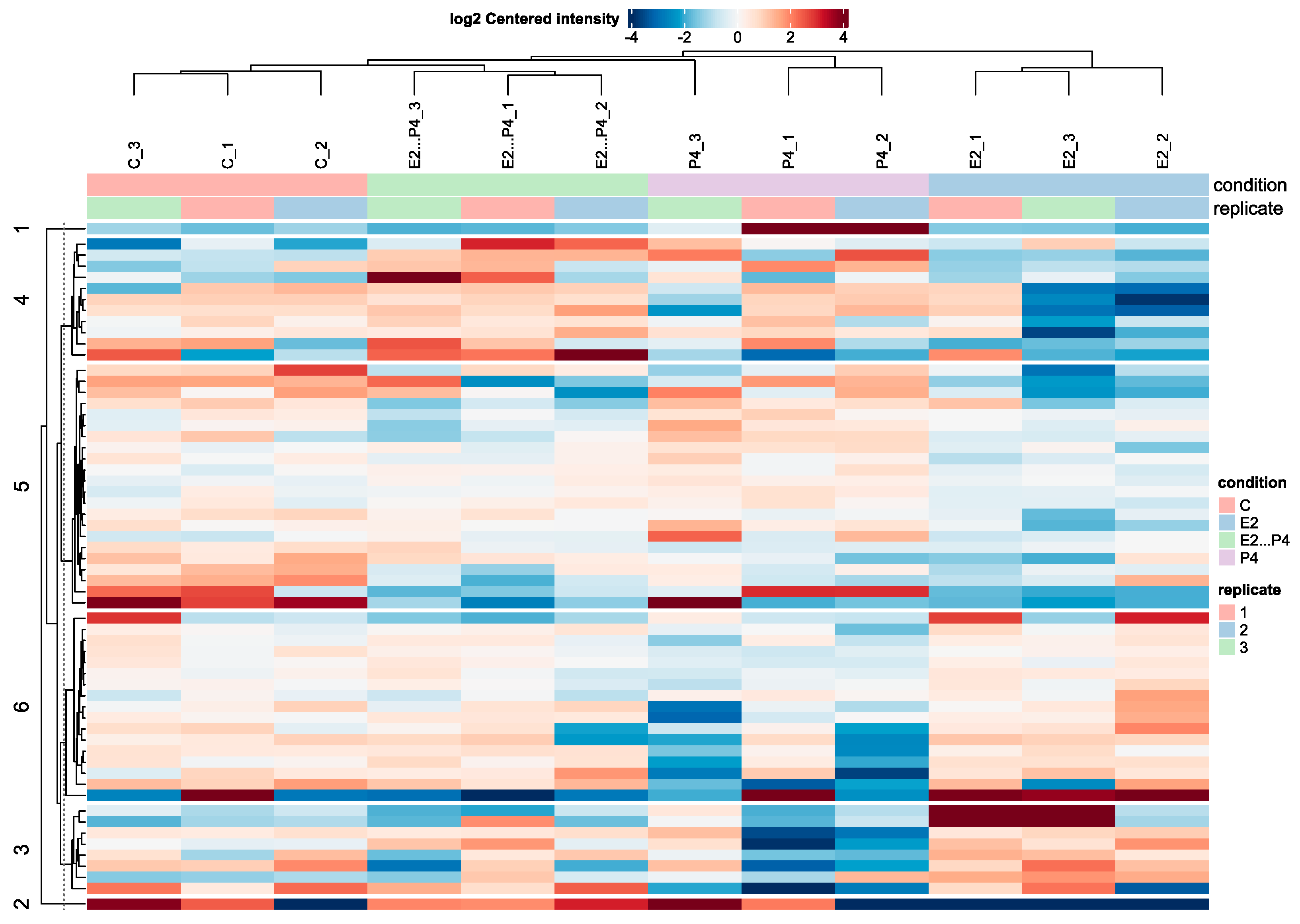
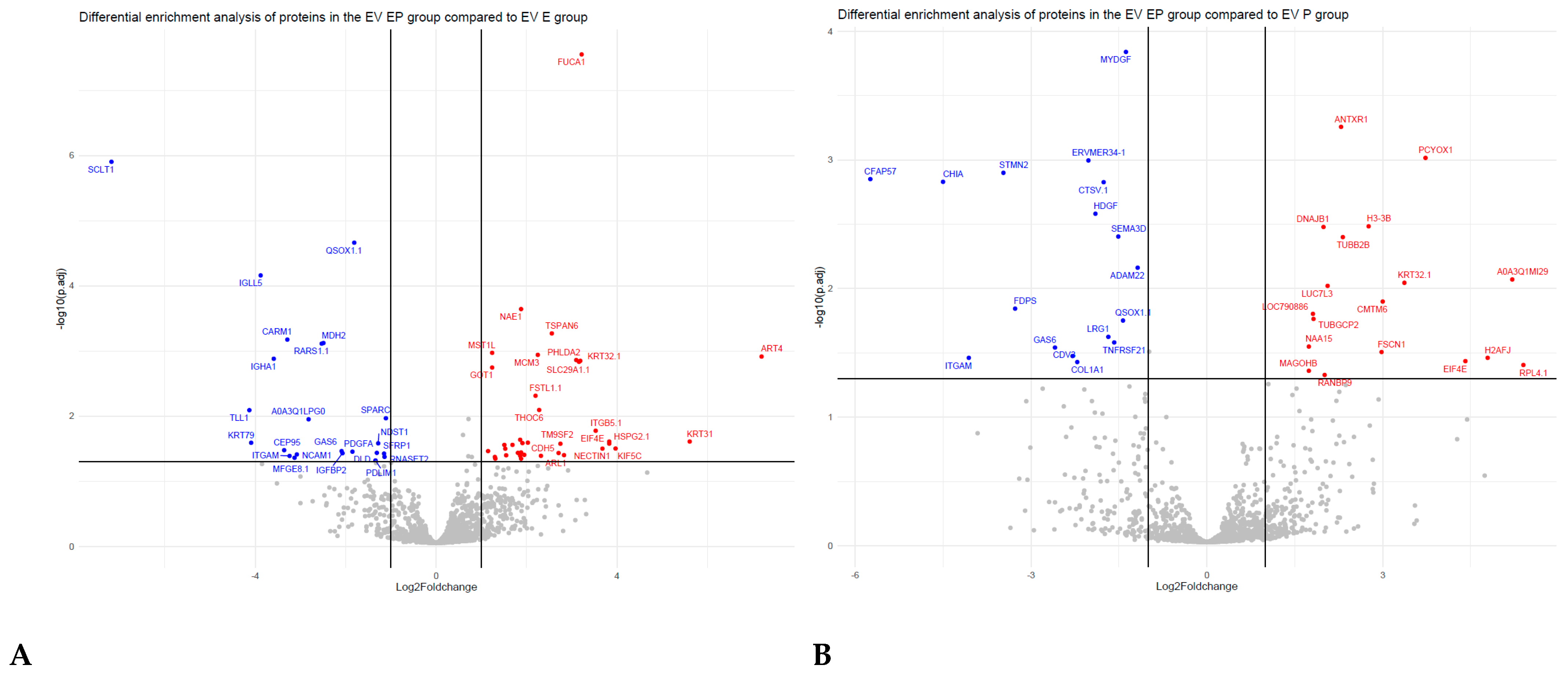
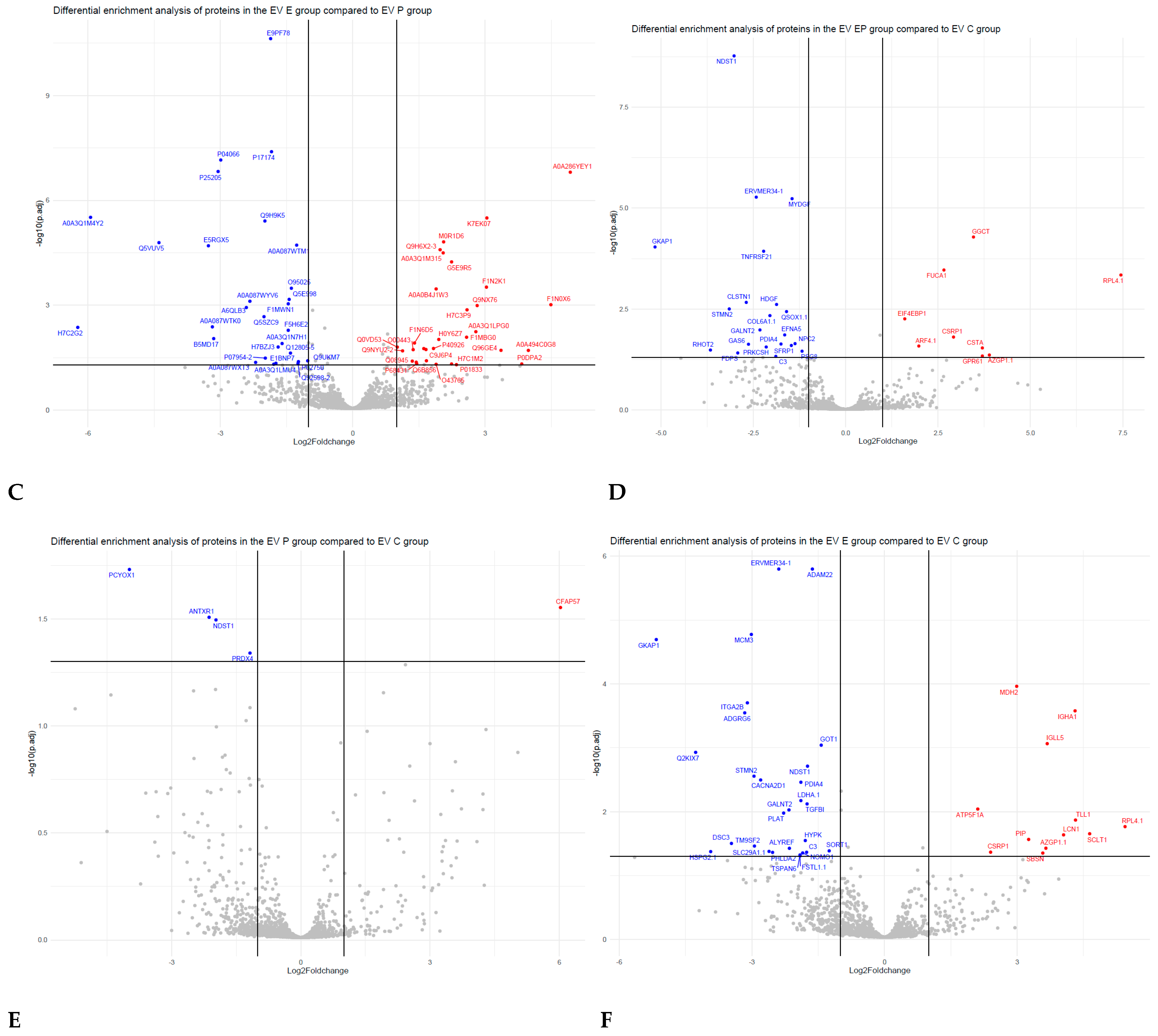
3.3. Comparative Proteomic Analysis of Endometrial EVs under Hormonal Treatment

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniProt Accession | Gene Name | Protein Description | Evidence | Reference |
|---|---|---|---|---|
| Embryo implantation | ||||
| Q6URK6 | CDH5 | Cadherin-5 | CDH5 has been shown to be expressed in the late proliferative phase in the endometrium. A mouse in vitro study has demonstrated that the expression of CDH5 in trophectoderm helps to facilitate embryo implantation. | [20,21] |
| F1MER7 | HSPG2 | Basement membrane-specific heparan sulfate proteoglycan core protein | Shown to be an abundant cargo of the luteal phase EVs and have a specific role in embryo implantation. HSPG2 within the uterine epithelium has been shown to facilitate trophoblast attachment and adhesion. | [19,22] |
| F6RAG5 | KIF5C | Kinesin heavy chain isoform 5C | Gene expression increased in embryo-endometrium contact co-cultures, as well as being shown to play a role during different morphogenetic processes in early embryo development. | [23,24] |
| A0A452DJ21 | EIF4E | Eukaryotic translation initiation factor 4E | The expression if EIF4E has been examined in porcine endometrium during implantation, with truncated EIF4E shown to particularly regulate protein synthesis during conceptus attachment at the time of implantation. | [25] |
| Q58D84 | FSTL1 | Follistatin-related protein 1 | Identified in menstrual fluid samples and is a common inflammatory and repair protein. | [26] |
| A6QLB3 | ITGA2B | Integrin subunit alpha 2b | ITGA2B has been identified during the stages of bovine trophectoderm cell migration and fusion with the uterine epithelial cells. | [27] |
| Q9NRN7 | AASDHPPT | l-aminoadipate-semialdehyde dehydrogenase-phosphopantetheinyl transferase | Found to be upregulated in the endometrium during pregnancy compared to genes during the estrous cycle. | [28] |
| Endometrial receptivity | ||||
| A0A452DJE0 | ACE2 | Angiotensin-converting enzyme 2 | ACE2 within the female reproductive system is abundantly expressed and is responsible for generating angiotensin, which has been shown to stimulate ovarian follicle growth, ovulation, and oocyte maturation, and is within the human endometrium. | [29] |
| H7BZJ3 | PDIA3 | Protein disulfide-isomerase A3 | PDIA3 upregulated in fertile endometriums during secretory phase as compared to proliferative phase. | [30] |
| P00750 | PLAT | Tissue-type plasminogen activator | In the reproductive system, PLAT mediates tissue remodelling required for ovulation and endometrial receptivity. | [31] |
| C9JPV1 | SLC6A6 | Sodium- and chloride-dependent taurine transporter | SLC6A6 gene expression has been shown to be greater in cow endometria in the large preovulatory follicle compared to the small preovulatory follicles. This gene is thought to have a role in amino acid metabolism which is linked to the receptive state of the endometrial tissue. | [32] |
| A0A087WYV6 | TSPAN6 | Tetraspanin-6 | Found in endometrial exosomes and thought to have functional and regulatory roles. | [33] |
| M0R1D6 | DNAJB1 | DnaJ homolog subfamily B member 1 | One of many novel genes identified and associated with the biological process of menstrual cycle. | [34] |
| D6RDI2 | LUC7L3 | Luc7-like protein 3 | Upregulated in human endometrial sample in women who exhibited implantation failure. | [35] |
| A0A3Q1LMU4 | INHBB | Inhibin beta B chain | Known marker gene of decidualization. | [36] |
| Embryo development | ||||
| P04066 | FUCA1 | Tissue alpha-l-fucosidase | FUCA1 was seen to be in higher abundance at elongation, as well as upregulated during day 15 of the cycle compared to day 7. | [37,38] |
| P00338 | LDHA | l-lactate dehydrogenase A chain | When LDHA is inhibited, histone lactylation level is reduced and an impaired rate of development of pre-implantation mice embryos is seen. | [39] |
4. Discussion
4.1. Receptive Phase Endometrial Cell EVs Were Enriched with Proteins Involved in Embryo Implantation
4.2. Receptive Phase Endometrial EVs Were Enriched in Proteins Involved in Embryo Development
4.3. Receptive Phase Endometrial EVs Were Enriched in Proteins Involved in Endometrial Receptivity
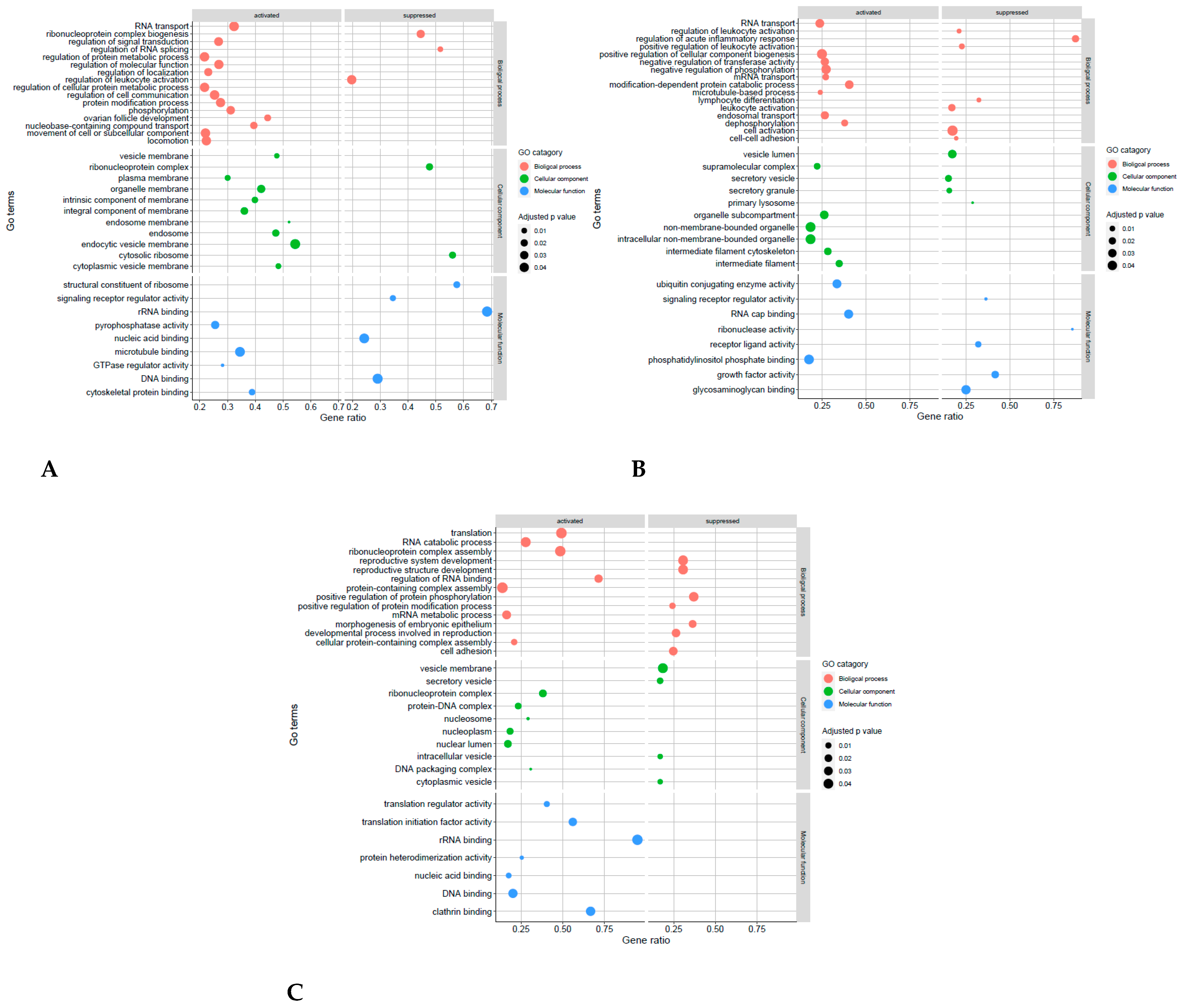
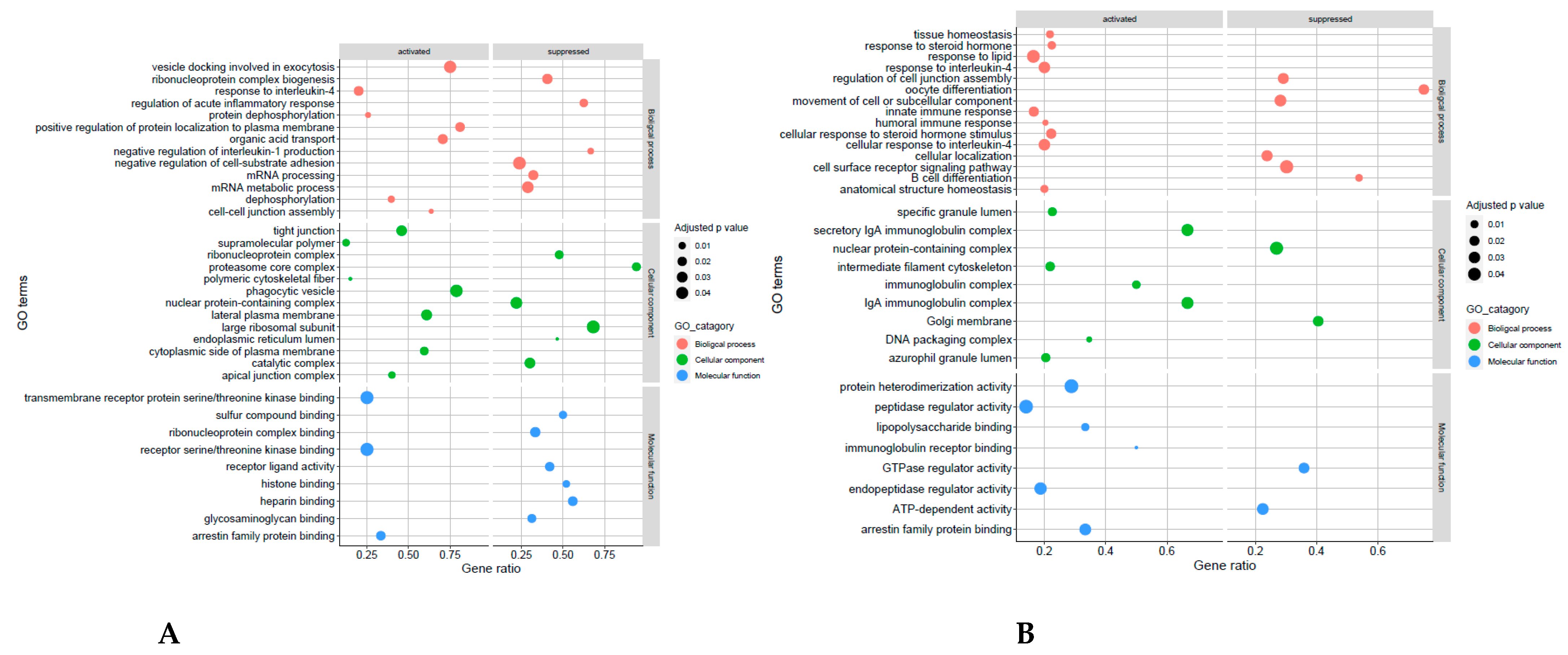
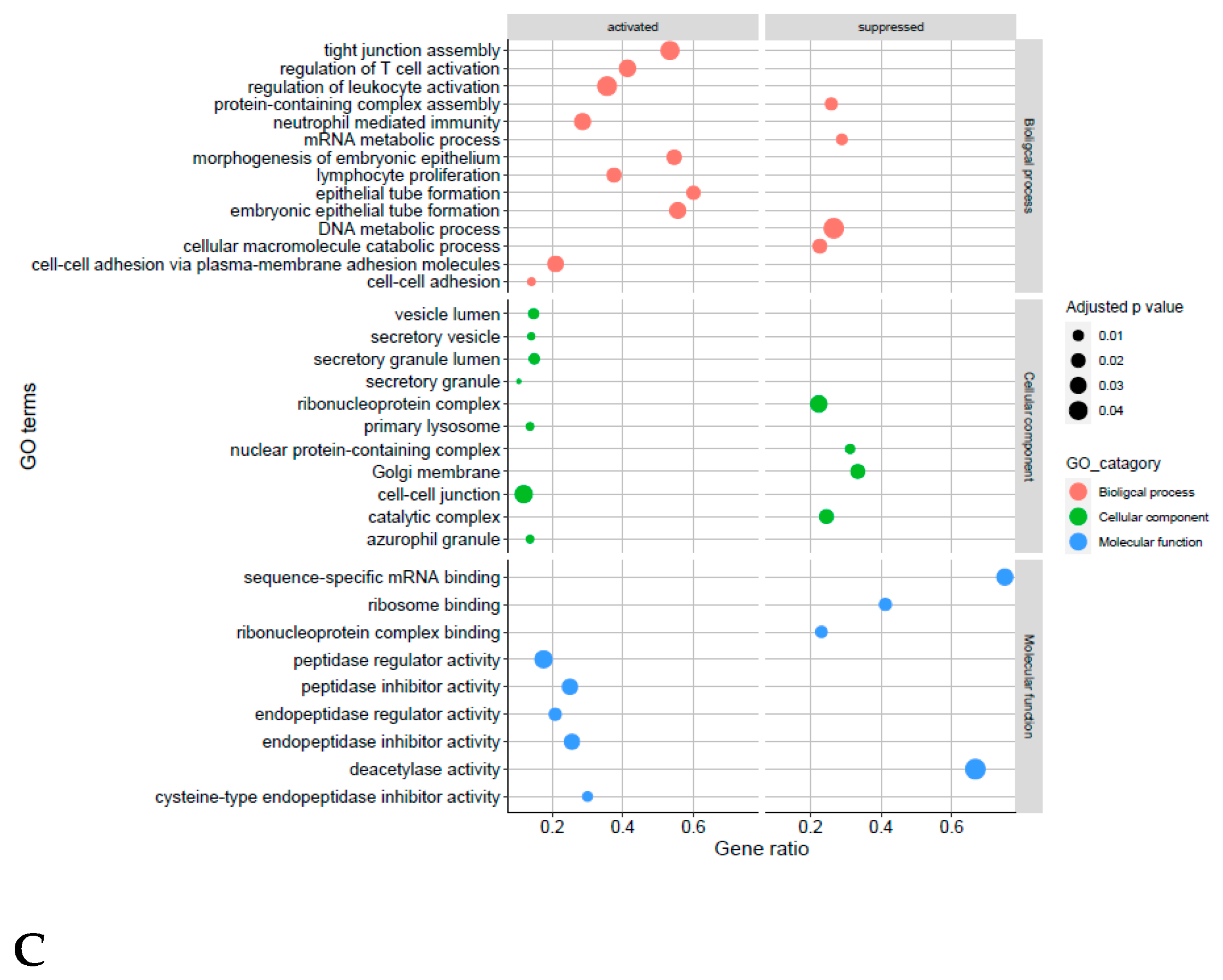
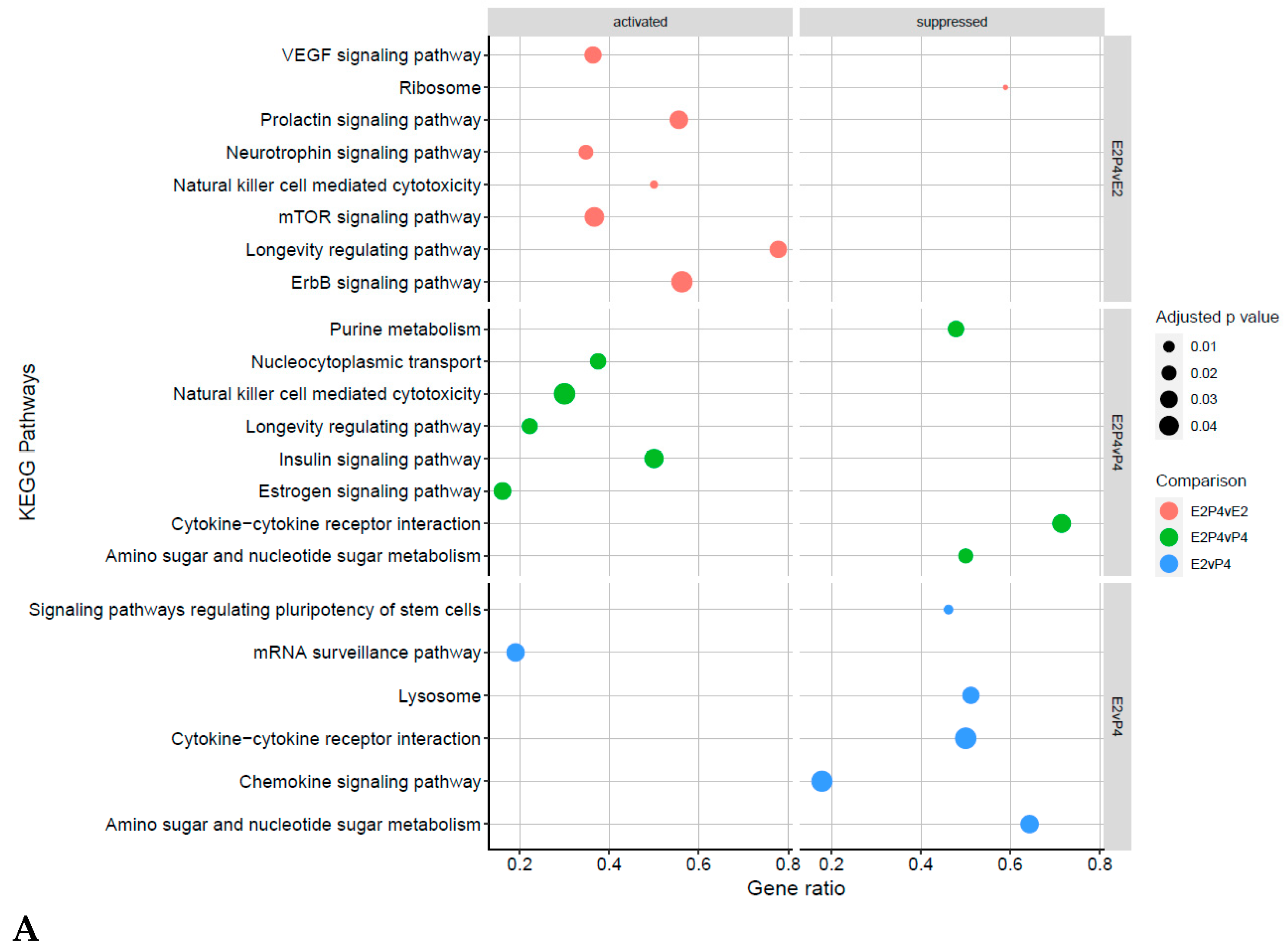
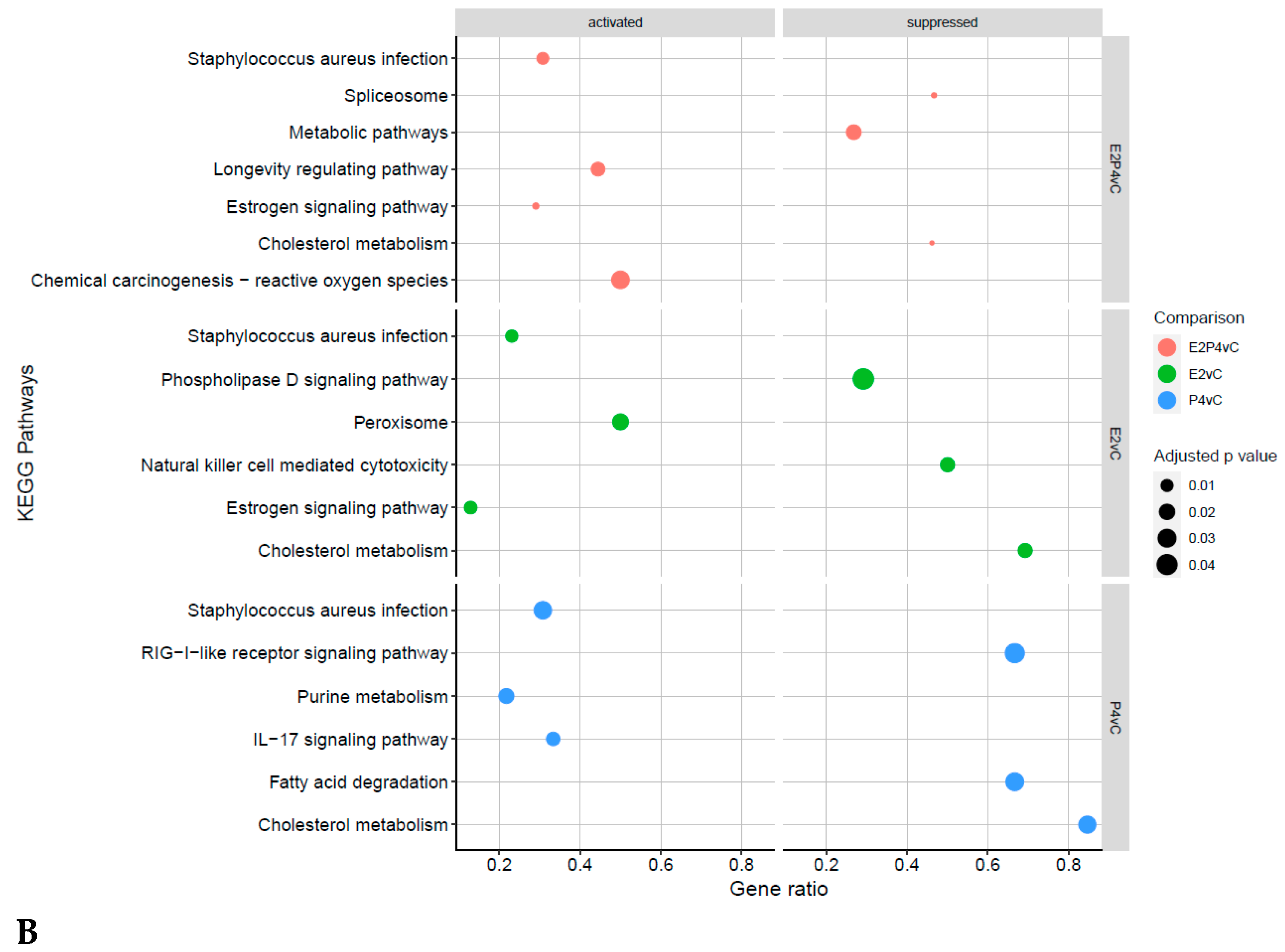
4.4. Receptive Phase Endometrial EVs GO Terms and KEGG Pathways Are Associated with Receptivity and Implantation Success
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reed, B.G.; Carr, B.R. The Normal Menstrual Cycle and the Control of Ovulation. In Endotext; MDText.com, Inc.: South Dartmouth, MA, USA, 2021. [Google Scholar]
- Taylor, A. ABC of Subfertility: Extent of the Problem. Br. Med. J. 2003, 327, 434. [Google Scholar] [CrossRef]
- Okada, H.; Tsuzuki, T.; Murata, H. Decidualization of the Human Endometrium. Reprod. Med. Biol. 2018, 17, 220–227. [Google Scholar] [CrossRef]
- Bhagwat, S.R.; Chandrashekar, D.S.; Kakar, R.; Davuluri, S.; Bajpai, A.K.; Nayak, S.; Bhutada, S.; Acharya, K.; Sachdeva, G. Endometrial Receptivity: A Revisit to Functional Genomics Studies on Human Endometrium and Creation of HGEx-ERdb. PLoS ONE 2013, 8, e58419. [Google Scholar] [CrossRef]
- Es-Haghi, M.; Godakumara, K.; Häling, A.; Lättekivi, F.; Lavrits, A.; Viil, J.; Andronowska, A.; Nafee, T.; James, V.; Jaakma, Ü.; et al. Specific Trophoblast Transcripts Transferred by Extracellular Vesicles Affect Gene Expression in Endometrial Epithelial Cells and May Have a Role in Embryo-Maternal Crosstalk. Cell Commun. Signal. 2019, 17, 146. [Google Scholar] [CrossRef]
- O’Neil, E.V.; Burns, G.W.; Ferreira, C.R.; Spencer, T.E. Characterization and Regulation of Extracellular Vesicles in the Lumen of the Ovine Uterus. Biol. Reprod. 2020, 102, 1020–1032. [Google Scholar] [CrossRef]
- Nakamura, K.; Kusama, K.; Ideta, A.; Kimura, K.; Hori, M.; Imakawa, K. Effects of MiR-98 in Intrauterine Extracellular Vesicles on Maternal Immune Regulation during the Peri-Implantation Period in Cattle. Sci. Rep. 2019, 9, 20330. [Google Scholar] [CrossRef]
- Dissanayake, K.; Nõmm, M.; Lättekivi, F.; Ord, J.; Ressaissi, Y.; Godakumara, K.; Reshi, Q.U.A.; Viil, J.; Jääger, K.; Velthut-Meikas, A.; et al. Oviduct as a Sensor of Embryo Quality: Deciphering the Extracellular Vesicle (EV)-Mediated Embryo-Maternal Dialogue. J. Mol. Med. 2021, 99, 685–697. [Google Scholar] [CrossRef]
- Caponnetto, F.; Manini, I.; Skrap, M.; Palmai-Pallag, T.; Di Loreto, C.; Beltrami, A.P.; Cesselli, D.; Ferrari, E. Size-Dependent Cellular Uptake of Exosomes. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 1011–1020. [Google Scholar] [CrossRef]
- Li, T.; Greenblatt, E.M.; Shin, M.E.J.; Brown, T.J.; Chan, C. Cargo Small Non-Coding RNAs of Extracellular Vesicles Isolated from Uterine Fluid Associate with Endometrial Receptivity and Implantation Success. Fertil. Steril. 2021, 115, 1327–1336. [Google Scholar] [CrossRef]
- Segura-Benítez, M.; Carbajo-García, M.C.; Corachán, A.; Faus, A.; Pellicer, A.; Ferrero, H. Proteomic Analysis of Extracellular Vesicles Secreted by Primary Human Epithelial Endometrial Cells Reveals Key Proteins Related to Embryo Implantation. Reprod. Biol. Endocrinol. 2022, 20, 3. [Google Scholar] [CrossRef]
- Giacomini, E.; Scotti, G.M.; Vanni, V.S.; Lazarevic, D.; Makieva, S.; Privitera, L.; Signorelli, S.; Cantone, L.; Bollati, V.; Murdica, V.; et al. Global Transcriptomic Changes Occur in Uterine Fluid-Derived Extracellular Vesicles during the Endometrial Window for Embryo Implantation. Hum. Reprod. 2021, 36, 2249–2274. [Google Scholar] [CrossRef]
- Burns, G.W.; Brooks, K.E.; Spencer, T.E. Extracellular Vesicles Originate from the Conceptus and Uterus during Early Pregnancy in Sheep. Biol. Reprod. 2016, 94, 56. [Google Scholar] [CrossRef]
- Almiñana, C.; Vegas, A.R.; Tekin, M.; Hassan, M.; Uzbekov, R.; Fröhlich, T.; Bollwein, H.; Bauersachs, S. Isolation and Characterization of Equine Uterine Extracellular Vesicles: A Comparative Methodological Study. Int. J. Mol. Sci. 2021, 22, 979. [Google Scholar] [CrossRef]
- Nakamura, K.; Kusama, K.; Ideta, A.; Imakawa, K.; Hori, M. IFNT-Independent Effects of Intrauterine Extracellular Vesicles (EVs) in Cattle. Reproduction 2020, 159, 503–511. [Google Scholar] [CrossRef]
- Thippabhotla, S.; Zhong, C.; He, M. 3D Cell Culture Stimulates the Secretion of in Vivo like Extracellular Vesicles. Sci. Rep. 2019, 9, 13012. [Google Scholar] [CrossRef]
- Ng, Y.H.; Rome, S.; Jalabert, A.; Forterre, A.; Singh, H.; Hincks, C.L.; Salamonsen, L.A. Endometrial Exosomes/Microvesicles in the Uterine Microenvironment: A New Paradigm for Embryo-Endometrial Cross Talk at Implantation. PLoS ONE 2013, 8, e58502. [Google Scholar] [CrossRef]
- Yang, H.; Lei, C.; Cheng, C.; Feng, Y.; Zhang, W.; Petracco, R.G.; Sak, S. The Antiapoptotic Effect of Galectin-3 in Human Endometrial Cells under the Regulation of Estrogen and Progesterone. Biol. Reprod. 2012, 87, 39. [Google Scholar] [CrossRef]
- Greening, D.W.; Nguyen, H.P.T.; Elgass, K.; Simpson, R.J.; Salamonsen, L.A. Human Endometrial Exosomes Contain Hormone-Specific Cargo Modulating Trophoblast Adhesive Capacity: Insights into Endometrial-Embryo Interactions. Biol. Reprod. 2016, 94, 38. [Google Scholar] [CrossRef]
- Zhou, W.; Santos, L.; Dimitriadis, E. Characterization of the Role for Cadherin 6 in the Regulation of Human Endometrial Receptivity. Reprod. Biol. Endocrinol. 2020, 18, 66. [Google Scholar] [CrossRef]
- Vilella, F.; Moreno-Moya, J.M.; Balaguer, N.; Grasso, A.; Herrero, M.; Martínez, S.; Marcilla, A.; Simón, C. Hsa-MiR-30d, Secreted by the Human Endometrium, Is Taken up by the Pre-Implantation Embryo and Might Modify Its Transcriptome. Development 2015, 142, 3210–3221. [Google Scholar] [CrossRef]
- Slater, M.; Murphy, C.R. Chondroitin Sulphate and Heparan Sulfate Proteoglycan Are Sequentially Expressed in the Uterine Extracellular Matrix during Early Pregnancy in the Rat. Matrix Biol. 1999, 18, 125–131. [Google Scholar] [CrossRef]
- Sponchiado, M.; Marei, W.F.A.; Beemster, G.T.S.; Bols, P.E.J.; Binelli, M.; Leroy, J.L.M.R. Molecular Interactions at the Bovine Embryo–Endometrial Epithelium Interface. Reproduction 2020, 160, 887–903. [Google Scholar] [CrossRef]
- Dathe, V.; Pröls, F.; Brand-Saberi, B. Expression of Kinesin Kif5c during Chick Development. Anat. Embryol. 2004, 207, 475–480. [Google Scholar] [CrossRef]
- Wollenhaupt, K.; Reinke, K.; Brüssow, K.P.; Albrecht, D.; Tiemann, U.; Seyfert, H.M.; Tomek, W. Natural Occurrence and Physiological Role of a Truncated EIF4E in the Porcine Endometrium during Implantation. Biochem. J. 2010, 432, 353–363. [Google Scholar] [CrossRef]
- Salamonsen, L.A. Menstrual Fluid Factors Mediate Endometrial Repair. Front. Reprod. Health 2021, 3, 779979. [Google Scholar] [CrossRef]
- Imakawa, K.; Bai, R.; Fujiwara, H.; Kusama, K. Conceptus Implantation and Placentation: Molecules Related to Epithelial–Mesenchymal Transition, Lymphocyte Homing, Endogenous Retroviruses, and Exosomes. Reprod. Med. Biol. 2016, 15, 1–11. [Google Scholar] [CrossRef]
- Kim, M.; Seo, H.; Choi, Y.; Shim, J.; Kim, H.; Lee, C.K.; Ka, H. Microarray Analysis of Gene Expression in the Uterine Endometrium during the Implantation Period in Pigs. Asian-Australas. J. Anim. Sci. 2012, 25, 1102–1116. [Google Scholar] [CrossRef]
- Reis, F.M.; Reis, A.M. Angiotensin-Converting Enzyme 2 (ACE2), Angiotensin-(1-7) and Mas Receptor in Gonadal and Reproductive Functions. Clin. Sci. 2020, 134, 2929–2941. [Google Scholar] [CrossRef]
- Rai, P.; Kota, V.; Sundaram, C.S.; Deendayal, M.; Shivaji, S. Proteome of Human Endometrium: Identification of Differentially Expressed Proteins in Proliferative and Secretory Phase Endometrium. Proteom. Clin. Appl. 2010, 4, 48–59. [Google Scholar] [CrossRef]
- Hoirisch-Clapauch, S.; Brenner, B. The Role of the Fibrinolytic System in Female Reproductive Disorders and Depression. Thromb. Update 2020, 1, 100004. [Google Scholar] [CrossRef]
- França, M.R.; da Silva, M.I.S.; Pugliesi, G.; Van Hoeck, V.; Binelli, M. Evidence of Endometrial Amino Acid Metabolism and Transport Modulation by Peri-Ovulatory Endocrine Profiles Driving Uterine Receptivity. J. Anim. Sci. Biotechnol. 2017, 8, 54. [Google Scholar] [CrossRef]
- Koh, Y.Q.; Peiris, H.N.; Vaswani, K.; Reed, S.; Rice, G.E.; Salomon, C.; Mitchell, M.D. Characterization of Exosomal Release in Bovine Endometrial Intercaruncular Stromal Cells. Reprod. Biol. Endocrinol. 2016, 14, 78. [Google Scholar] [CrossRef] [PubMed]
- Groothuis, P.G.; Dassen, H.H.N.M.; Romano, A.; Punyadeera, C. Estrogen and the Endometrium: Lessons Learned from Gene Expression Profiling in Rodents and Human. Hum. Reprod. Update 2007, 13, 405–417. [Google Scholar] [CrossRef]
- Tapia-Pizarro, A.; Figueroa, P.; Brito, J.; Marín, J.C.; Munroe, D.J.; Croxatto, H.B. Endometrial Gene Expression Reveals Compromised Progesterone Signaling in Women Refractory to Embryo Implantation. Reprod. Biol. Endocrinol. 2014, 12, 92. [Google Scholar] [CrossRef]
- Griffiths IV, R.M.; Pru, C.A.; Behura, S.K.; Cronrath, A.R.; McCallum, M.L.; Kelp, N.C.; Winuthayanon, W.; Spencer, T.E.; Pru, J.K. AMPK Is Required for Uterine Receptivity and Normal Responses to Steroid Hormones. Reproduction 2020, 159, 707–717. [Google Scholar] [CrossRef]
- Van der Weijden, V.A.; Bick, J.T.; Bauersachs, S.; Arnold, G.J.; Fröhlich, T.; Drews, B.; Ulbrich, S.E. Uterine Fluid Proteome Changes during Diapause and Resumption of Embryo Development in Roe Deer (Capreolus capreolus). Reproduction 2019, 158, 13–24. [Google Scholar] [CrossRef]
- Faulkner, S.; Elia, G.; O’Boyle, P.; Dunn, M.; Morris, D. Composition of the Bovine Uterine Proteome Is Associated with Stage of Cycle and Concentration of Systemic Progesterone. Proteomics 2013, 13, 3333–3353. [Google Scholar] [CrossRef]
- Yang, W.; Wang, P.; Cao, P.; Wang, S.; Yang, Y.; Su, H.; Nashun, B. Hypoxic in Vitro Culture Reduces Histone Lactylation and Impairs Pre-Implantation Embryonic Development in Mice. Epigenet. Chromatin 2021, 14, 57. [Google Scholar] [CrossRef]
- Legato, M.J. Principles of Gender-Specific Medicine: Gender in the Genomic Era, 3rd ed.; Academic Press: Cambridge, MA, USA, 2017. [Google Scholar]
- Fazeli, A.; Pewsey, E. Maternal Communication with Gametes and Embryos: A Complex Interactome. Brief. Funct. Genom. Proteom. 2008, 7, 111–118. [Google Scholar] [CrossRef]
- Fazeli, A.; Holt, W.V. Cross Talk during the Periconception Period. Theriogenology 2016, 86, 438–442. [Google Scholar] [CrossRef]
- Fazeli, A.; Moein Vaziri, N.; Holt, W.V. Proteomics of the Periconception Milieu. Proteomics 2015, 15, 649–655. [Google Scholar] [CrossRef]
- Hart, A.R.; Khan, N.L.A.; Godakumara, K.; Dissanayake, K.; Piibor, J.; Muhandiram, S.; Eapen, S.; Heath, P.R.; Fazeli, A. The Role of Extracellular Vesicles in Endometrial Receptivity and Their Potential in Reproductive Therapeutics and Diagnosis. Reprod. Biol. 2022, 22, 100645. [Google Scholar] [CrossRef]
- Godakumara, K.; Ord, J.; Lättekivi, F.; Dissanayake, K.; Viil, J.; Boggavarapu, N.R.; Faridani, O.R.; Jääger, K.; Velthut-Meikas, A.; Jaakma, Ü.; et al. Trophoblast Derived Extracellular Vesicles Specifically Alter the Transcriptome of Endometrial Cells and May Constitute a Critical Component of Embryo-Maternal Communication. Reprod. Biol. Endocrinol. 2021, 19, 115. [Google Scholar] [CrossRef]
- Way, D.L.; Grosso, D.S.; Davis, J.R.; Surwit, E.A.; Christian, C.D. Characterization of a New Human Endometrial Carcinoma (RL95-2) Established in Tissue Culture. In Vitro 1983, 19, 147–158. [Google Scholar] [CrossRef]
- Harduf, H.; Goldman, S.; Shalev, E. Human Uterine Epithelial RL95-2 and HEC-1A Cell-Line Adhesiveness: The Role of Plexin B1. Fertil. Steril. 2007, 87, 1419–1427. [Google Scholar] [CrossRef]
- Thie, M.; Denker, H.W. In Vitro Studies on Endometrial Adhesiveness for Trophoblast: Cellular Dynamics in Uterine Epithelial Cells. Cells Tissues Organs 2002, 172, 237–252. [Google Scholar] [CrossRef]
- Harduf, H.; Goldman, S.; Shalev, E. Progesterone Receptor A and C-Met Mediates Spheroids-Endometrium Attachment. Reprod. Biol. Endocrinol. 2009, 7, 14. [Google Scholar] [CrossRef]
- Thie, M.; Herter, P.; Pommerenke, H.; Dürr, F.; Sieckmann, F.; Nebe, B.; Rychly, J.; Denker, H.W. Adhesiveness of the Free Surface of a Human Endometrial Monolayer for Trophoblast as Related to Actin Cytoskeleton. Mol. Hum. Reprod. 1997, 3, 275–283. [Google Scholar] [CrossRef]
- Heneweer, C.; Kruse, L.H.; Kindháuser, F.; Schmidt, M.; Jakobs, K.H.; Denker, H.W.; Thie, M. Adhesiveness of Human Uterine Epithelial RL95-2 Cells to Trophoblast: Rho Protein Regulation. Mol. Hum. Reprod. 2002, 8, 1014–1022. [Google Scholar] [CrossRef]
- Burns, G.W.; Brooks, K.E.; O’Neil, E.V.; Hagen, D.E.; Behura, S.K.; Spencer, T.E. Progesterone Effects on Extracellular Vesicles in the Sheep Uterus. Biol. Reprod. 2018, 98, 612–622. [Google Scholar] [CrossRef]
- Dassen, H.; Punyadeera, C.; Kamps, R.; Klomp, J.; Dunselman, G.; Dijcks, F.; De Goeij, A.; Ederveen, A.; Groothuis, P. Progesterone Regulation of Implantation-Related Genes: New Insights into the Role of Oestrogen. Cell. Mol. Life Sci. 2007, 64, 1009–1032. [Google Scholar] [CrossRef]
- Gargett, C.E.; Schwab, K.E.; Deane, J.A. Endometrial Stem/Progenitor Cells: The First 10 Years. Hum. Reprod. Update 2016, 22, 137–163. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Wang, A.; Feng, L.; Wang, Y.; Zhang, H.; Zhang, I.; Bany, B.M.; Ma, L. Heparan Sulfate Proteoglycan Sulfation Regulates Uterine Differentiation and Signaling during Embryo Implantation. Endocrinology 2018, 159, 2459–2472. [Google Scholar] [CrossRef]
- Carson, D.D.; Tang, J.P.; Julian, J. Heparan Sulfate Proteoglycan (Perlecan) Expression by Mouse Embryos during Acquisition of Attachment Competence. Dev. Biol. 1993, 155, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Henarejos-Castillo, I.; Sebastian-Leon, P.; Devesa-Peiro, A.; Pellicer, A.; Diaz-Gimeno, P. SARS-CoV-2 Infection Risk Assessment in the Endometrium: Viral Infection-Related Gene Expression across the Menstrual Cycle. Fertil. Steril. 2020, 114, 223–232. [Google Scholar] [CrossRef]
- Stelzig, K.E.; Canepa-Escaro, F.; Schiliro, M.; Berdnikovs, S.; Prakash, Y.S.; Chiarella, S.E. Estrogen Regulates the Expression of SARS-CoV-2 Receptor ACE2 in Differentiated Airway Epithelial Cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2020, 318, L1280–L1281. [Google Scholar] [CrossRef]
- McConaha, M.E.; Eckstrum, K.; An, J.; Steinle, J.J.; Bany, B.M. Microarray Assessment of the Influence of the Conceptus on Gene Expression in the Mouse Uterus during Decidualization. Reproduction 2011, 141, 511–527. [Google Scholar] [CrossRef]
- Horne, A.W.; Van Den Driesche, S.; King, A.E.; Burgess, S.; Myers, M.; Ludlow, H.; Lourenco, P.; Ghazal, P.; Williams, A.R.; Critchley, H.O.D.; et al. Endometrial Inhibin/Activin β-B Subunit Expression Is Related to Decidualization and Is Reduced in Tubal Ectopic Pregnancy. J. Clin. Endocrinol. Metab. 2008, 93, 2375–2382. [Google Scholar] [CrossRef]
- Xu, B.; Wang, J.; Xia, L.; Zhang, D.; Wu, X.; Zhang, A. Increased Uterine NK Cell Numbers and Perforin Expression during the Implantation Phase in IVF Cycles with GnRH Antagonist Protocol. Sci. Rep. 2017, 7, 39912. [Google Scholar] [CrossRef]
- Bulmer, J.N.; Morrison, L.; Longfellow, M.; Ritson, A.; Pace, D. Granulated Lymphocytes in Human Endometrium: Histochemical and Immunohistochemical Studies. Hum. Reprod. 1991, 6, 791–798. [Google Scholar] [CrossRef]
- Lash, G.E.; Otun, H.A.; Innes, B.A.; Percival, K.; Searle, R.F.; Robson, S.C.; Bulmer, J.N. Regulation of Extravillous Trophoblast Invasion by Uterine Natural Killer Cells Is Dependent on Gestational Age. Hum. Reprod. 2010, 25, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- El-Azzamy, H.; Dambaeva, S.V.; Katukurundage, D.; Garcia, M.D.S.; Skariah, A.; Hussein, Y.; Germain, A.; Fernandez, E.; Gilman-Sachs, A.; Beaman, K.D.; et al. Dysregulated Uterine Natural Killer Cells and Vascular Remodeling in Women with Recurrent Pregnancy Losses. Am. J. Reprod. Immunol. 2018, 80, e13024. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.; Li, X.; Sun, R.; Tong, X.; Ling, B.; Tian, Z.; Wei, H. Natural Killer Cells Promote Immune Tolerance by Regulating Inflammatory TH17 Cells at the Human Maternal-Fetal Interface. Proc. Natl. Acad. Sci. USA 2013, 110, E231–E240. [Google Scholar] [CrossRef] [PubMed]
- Manaster, I.; Mizrahi, S.; Goldman-Wohl, D.; Sela, H.Y.; Stern-Ginossar, N.; Lankry, D.; Gruda, R.; Hurwitz, A.; Bdolah, Y.; Haimov-Kochman, R.; et al. Endometrial NK Cells Are Special Immature Cells That Await Pregnancy. J. Immunol. 2008, 181, 1869–1876. [Google Scholar] [CrossRef]
- Laplante, M.; Sabatini, D.M. Regulation of MTORC1 and Its Impact on Gene Expression at a Glance. J. Cell Sci. 2013, 126, 1712–1719. [Google Scholar] [CrossRef]
- Chen, X.; He, J.; Ding, Y.; Zeng, L.; Gao, R.; Cheng, S.; Liu, X.; Wang, Y. The Role of MTOR in Mouse Uterus during Embryo Implantation. Reproduction 2009, 138, 351–356. [Google Scholar] [CrossRef]
- Salmasi, S.; Sharifi, M.; Rashidi, B. Evaluating the Effect of Ovarian Stimulation and Exogenous Progesterone on CD31-Positive Cell Density, VEGF Protein, and MiR-17-5p Expression of Endometrium Immediately before Implantation. Biomed. Pharmacother. 2021, 133, 110922. [Google Scholar] [CrossRef]
- Rabbani, M.L.; Rogers, P.A.W. Role of Vascular Endothelial Growth Factor in Endometrial Vascular Events before Implantation in Rats. Reproduction 2001, 122, 85–90. [Google Scholar] [CrossRef]
- Shao, L.; Wang, H.; Duan, E.; Zhu, C. Expression of Vascular Endothelial Growth Factor in Rat Uterus during Peri-Implantation. Chin. Sci. Bull. 2001, 46, 1178–1181. [Google Scholar] [CrossRef]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE Database Resources in 2022: A Hub for Mass Spectrometry-Based Proteomics Evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef] [PubMed]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hart, A.R.; Khan, N.L.A.; Dissanayake, K.; Godakumara, K.; Andronowska, A.; Eapen, S.; Heath, P.R.; Fazeli, A. The Extracellular Vesicles Proteome of Endometrial Cells Simulating the Receptive Menstrual Phase Differs from That of Endometrial Cells Simulating the Non-Receptive Menstrual Phase. Biomolecules 2023, 13, 279. https://doi.org/10.3390/biom13020279
Hart AR, Khan NLA, Dissanayake K, Godakumara K, Andronowska A, Eapen S, Heath PR, Fazeli A. The Extracellular Vesicles Proteome of Endometrial Cells Simulating the Receptive Menstrual Phase Differs from That of Endometrial Cells Simulating the Non-Receptive Menstrual Phase. Biomolecules. 2023; 13(2):279. https://doi.org/10.3390/biom13020279
Chicago/Turabian StyleHart, Amber Rose, Norhayati Liaqat Ali Khan, Keerthie Dissanayake, Kasun Godakumara, Aneta Andronowska, Saji Eapen, Paul R Heath, and Alireza Fazeli. 2023. "The Extracellular Vesicles Proteome of Endometrial Cells Simulating the Receptive Menstrual Phase Differs from That of Endometrial Cells Simulating the Non-Receptive Menstrual Phase" Biomolecules 13, no. 2: 279. https://doi.org/10.3390/biom13020279
APA StyleHart, A. R., Khan, N. L. A., Dissanayake, K., Godakumara, K., Andronowska, A., Eapen, S., Heath, P. R., & Fazeli, A. (2023). The Extracellular Vesicles Proteome of Endometrial Cells Simulating the Receptive Menstrual Phase Differs from That of Endometrial Cells Simulating the Non-Receptive Menstrual Phase. Biomolecules, 13(2), 279. https://doi.org/10.3390/biom13020279