
Glycome Profiling of Cancer Cell Lines Cultivated in Physiological and Commercial Media



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Preparation of Plasmax Medium
2.3. Cell Line Culturing and Harvesting
2.4. Protein Extraction
2.5. Release and Permethylation of Glycans
2.6. Total RNA Extraction and Transcriptomics Analysis
2.7. LC-MS/MS Glycomics Analysis
2.8. Data Analysis
3. Results
3.1. Culturing Media Specification
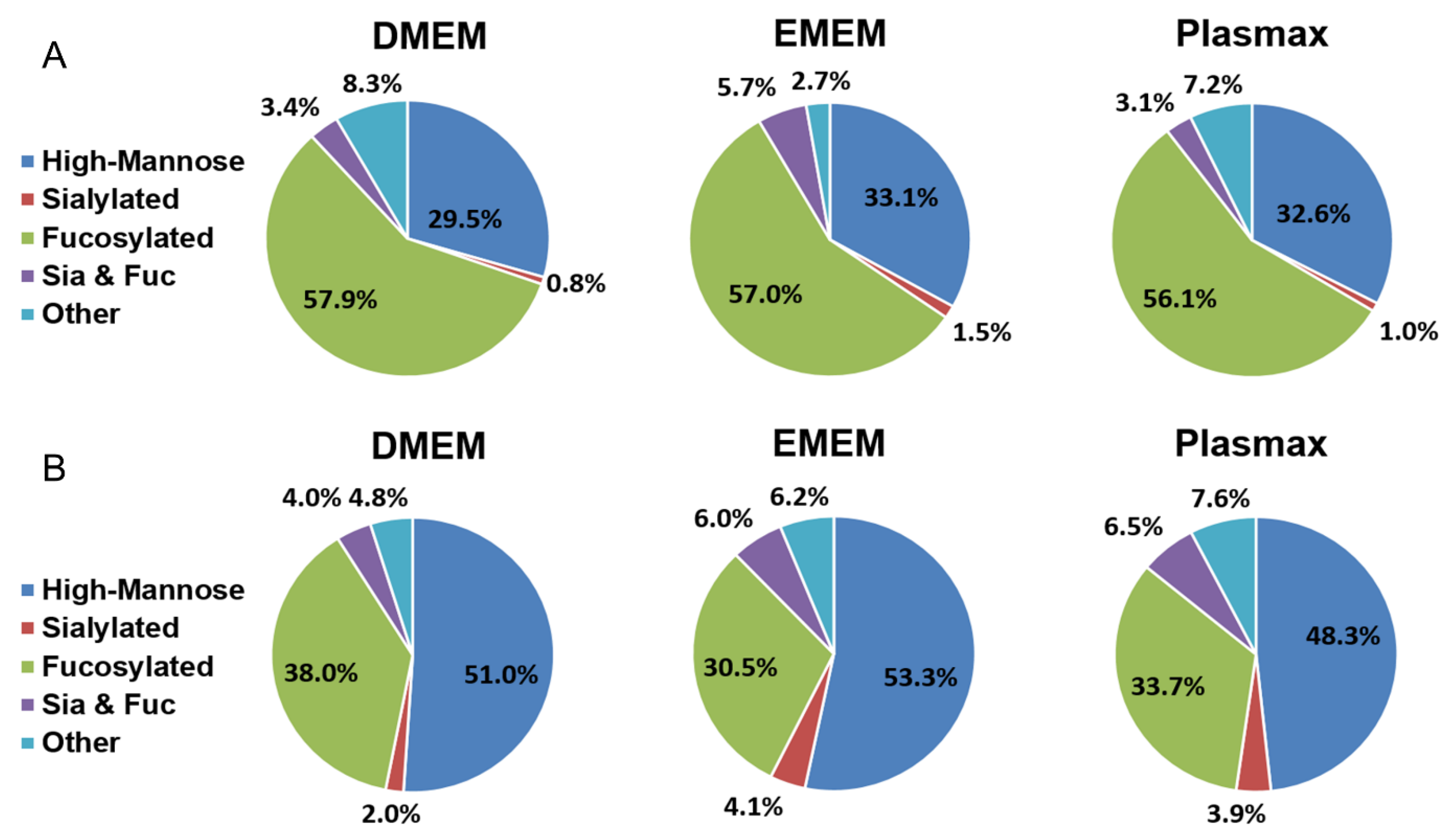
3.2. N-Glycan Identification
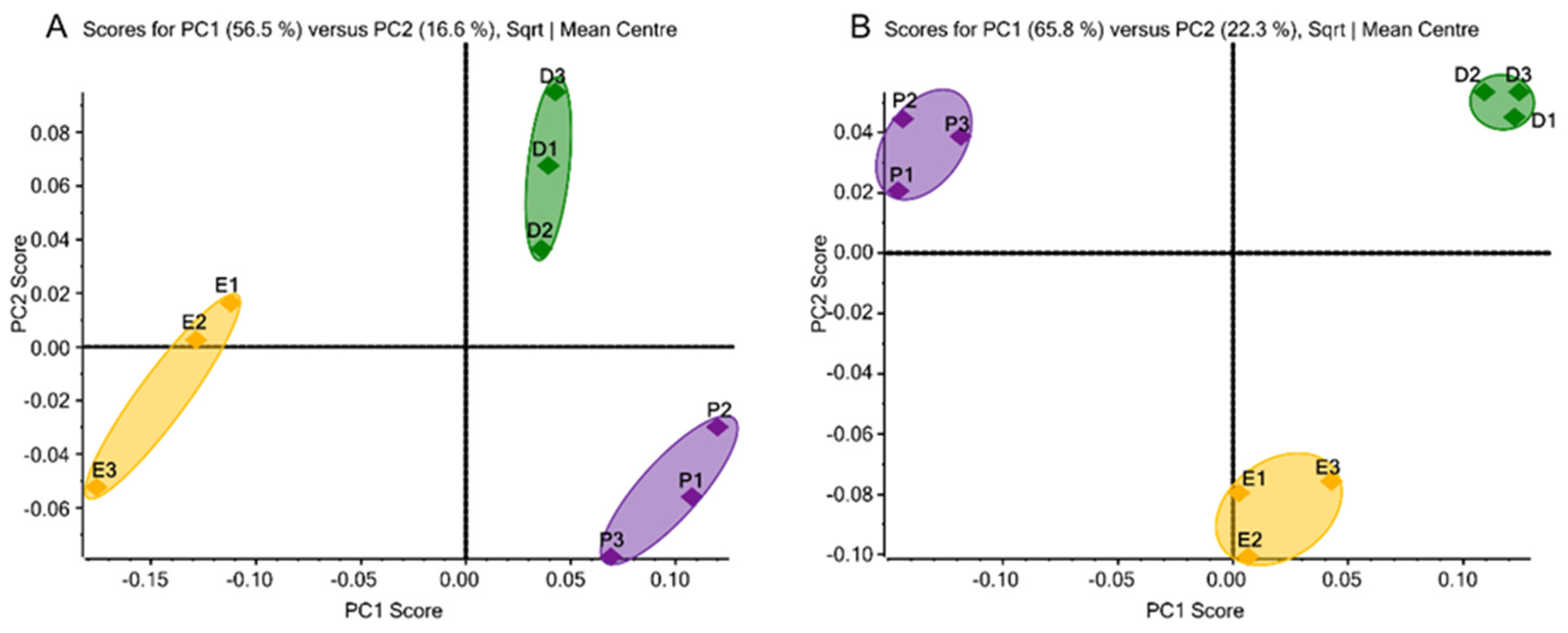
3.3. Chemometric Analysis of N-Glycans by Unsupervised Principal Component Analysis
3.4. N-Glycan Expression Changes among Plasmax, DMEM, and EMEM
3.5. Chemometric Analysis of O-Glycans by Unsupervised Principal Component Analysis
3.6. O-Glycan Expression Changes among Plasmax, DMEM, and EMEM
3.7. Expression Changes of Glycogenes in Plasmax
4. Discussion
5. Summary
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eagle, H. Amino acid metabolism in mammalian cell cultures. Science 1959, 130, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Yao, T.; Asayama, Y. Animal-cell culture media: History, characteristics, and current issues. Reprod. Med. Biol. 2017, 16, 99–117. [Google Scholar] [CrossRef] [PubMed]
- Dulbecco, R.; Freeman, G. Plaque production by the polyoma virus. Virology 1959, 8, 396–397. [Google Scholar] [CrossRef]
- Ben-David, U.; Siranosian, B.; Ha, G.; Tang, H.; Oren, Y.; Hinohara, K.; Strathdee, C.A.; Dempster, J.; Lyons, N.J.; Burns, R.; et al. Genetic and transcriptional evolution alters cancer cell line drug response. Nature 2018, 560, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Foldbjerg, R.; Dang, D.A.; Autrup, H. Cytotoxicity and genotoxicity of silver nanoparticles in the human lung cancer cell line, A549. Arch. Toxicol. 2011, 85, 743–750. [Google Scholar] [CrossRef]
- Adey, A.; Burton, J.N.; Kitzman, J.O.; Hiatt, J.B.; Lewis, A.P.; Martin, B.K.; Qiu, R.; Lee, C.; Shendure, J. The haplotype-resolved genome and epigenome of the aneuploid HeLa cancer cell line. Nature 2013, 500, 207–211. [Google Scholar] [CrossRef]
- Davidson, S.M.; Papagiannakopoulos, T.; Olenchock, B.A.; Heyman, J.E.; Keibler, M.A.; Luengo, A.; Bauer, M.R.; Jha, A.K.; O’Brien, J.P.; Pierce, K.A.; et al. Environment Impacts the Metabolic Dependencies of Ras-Driven Non-Small Cell Lung Cancer. Cell Metab. 2016, 23, 517–528. [Google Scholar] [CrossRef] [Green Version]
- Cantor, J.R.; Abu-Remaileh, M.; Kanarek, N.; Freinkman, E.; Gao, X.; Louissaint, A., Jr.; Lewis, C.A.; Sabatini, D.M. Physiologic Medium Rewires Cellular Metabolism and Reveals Uric Acid as an Endogenous Inhibitor of UMP Synthase. Cell 2017, 169, 258–272.e17. [Google Scholar] [CrossRef] [Green Version]
- Muir, A.; Danai, L.V.; Gui, D.Y.; Waingarten, C.Y.; Lewis, C.A.; Vander Heiden, M.G. Environmental cystine drives glutamine anaplerosis and sensitizes cancer cells to glutaminase inhibition. eLife 2017, 6, e27713. [Google Scholar] [CrossRef]
- Muir, A.; Vander Heiden, M.G. The nutrient environment affects therapy. Science 2018, 360, 962–963. [Google Scholar] [CrossRef]
- Birsoy, K.; Possemato, R.; Lorbeer, F.K.; Bayraktar, E.C.; Thiru, P.; Yucel, B.; Wang, T.; Chen, W.W.; Clish, C.B.; Sabatini, D.M. Metabolic determinants of cancer cell sensitivity to glucose limitation and biguanides. Nature 2014, 508, 108–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vande Voorde, J.; Ackermann, T.; Pfetzer, N.; Sumpton, D.; Mackay, G.; Kalna, G.; Nixon, C.; Blyth, K.; Gottlieb, E.; Tardito, S. Improving the metabolic fidelity of cancer models with a physiological cell culture medium. Sci. Adv. 2019, 5, eaau7314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, A.; Zhao, J.; Peng, W.; Banazadeh, A.; Williamson, S.D.; Goli, M.; Huang, Y.; Mechref, Y. Advances in mass spectrometry-based glycoproteomics. Electrophoresis 2018, 39, 3104–3122. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Zacharias, L.; Wooding, K.M.; Peng, W.; Mechref, Y. Glycoprotein enrichment analytical techniques: Advantages and disadvantages. Meth. Enzymol. 2017, 585, 397–429. [Google Scholar]
- Dong, X.; Huang, Y.; Cho, B.G.; Zhong, J.; Gautam, S.; Peng, W.; Williamson, S.D.; Banazadeh, A.; Torres-Ulloa, K.Y.; Mechref, Y. Advances in mass spectrometry-based glycomics. Electrophoresis 2018, 39, 3063–3081. [Google Scholar] [CrossRef]
- Mechref, Y.; Peng, W.; Banazadeh, A.; Huang, Y.; Zhou, S.; Hu, Y.; Hussein, A.; Tang, H. Quantitative Glycomics by Mass Spectrometry and Liquid Chromatography—Mass Spectrometry. In Encyclopedia of Analytical Chemistry: Applications, Theory and Instrumentation; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2006; pp. 1–31. [Google Scholar]
- Rudd, P.M.; Wormald, M.R.; Stanfield, R.L.; Huang, M.; Mattsson, N.; Speir, J.A.; DiGennaro, J.A.; Fetrow, J.S.; Dwek, R.A.; Wilson, I.A. Roles for glycosylation of cell surface receptors involved in cellular immune recognition. J. Mol. Biol. 1999, 293, 351–366. [Google Scholar] [CrossRef]
- Iida, J.; Meijne, A.M.; Knutson, J.R.; Furcht, L.T.; McCarthy, J.B. Cell surface chondroitin sulfate proteoglycans in tumor cell adhesion, motility and invasion. Semin. Cancer Biol. 1996, 7, 155–162. [Google Scholar] [CrossRef]
- Fuster, M.M.; Esko, J.D. The sweet and sour of cancer: Glycans as novel therapeutic targets. Nat. Rev. Cancer 2005, 5, 526–542. [Google Scholar] [CrossRef]
- Julien, S.; Lagadec, C.; Krzewinski-Recchi, M.-A.; Courtand, G.; Le Bourhis, X.; Delannoy, P. Stable expression of sialyl-Tn antigen in T47-D cells induces a decrease of cell adhesion and an increase of cell migration. Breast Cancer Res. Treat. 2005, 90, 77–84. [Google Scholar] [CrossRef]
- Munkley, J. The Role of Sialyl-Tn in Cancer. Int. J. Mol. Sci. 2016, 17, 275. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, H.; Swoger, J.; Greene, S.; Saito, T.; Hurh, J.; Sweeley, C.; Leestma, J.; Mkrdichian, E.; Cerullo, L.; Nishikawa, A. β1, 6-N-acetylglucosamine-bearing N-glycans in human gliomas: Implications for a role in regulating invasivity. Cancer Res. 2000, 60, 134–142. [Google Scholar] [PubMed]
- Kim, Y.-S.; Ahn, Y.H.; Song, K.J.; Kang, J.G.; Lee, J.H.; Jeon, S.K.; Kim, H.-C.; Yoo, J.S.; Ko, J.-H. Overexpression and β-1, 6-N-Acetylglucosaminylation-initiated Aberrant Glycosylation of TIMP-1 A “Double Whammy” Strategy in Colon Cancer Progression. J. Biol. Chem. 2012, 287, 32467–32478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, A.; Zhao, J.; Zhong, J.; Wang, J.; Yadav, S.P.S.; Molitoris, B.A.; Wagner, M.C.; Mechref, Y. Altered O-glycomes of Renal Brush-Border Membrane in Model Rats with Chronic Kidney Diseases. Biomolecules 2021, 11, 1560. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.; Zhao, J.; Yadav, S.P.S.; Molitoris, B.A.; Wagner, M.C.; Mechref, Y. Changes in the Expression of Renal Brush Border Membrane N-Glycome in Model Rats with Chronic Kidney Diseases. Biomolecules 2021, 11, 1677. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Mirzaei, P.; Zhu, R.; Zhou, S.; Mechref, Y. Comparative Membrane N-Glycomics of Different Breast Cancer Cell Lines To Understand Breast Cancer Brain Metastasis. J. Proteome Res. 2020, 19, 854–863. [Google Scholar] [CrossRef]
- Peng, W.; Zhu, R.; Zhou, S.; Mirzaei, P.; Mechref, Y. Integrated Transcriptomics, Proteomics, and Glycomics Reveals the Association between Up-regulation of Sialylated N-glycans/Integrin and Breast Cancer Brain Metastasis. Sci. Rep. 2019, 9, 17361. [Google Scholar] [CrossRef]
- Clémençon, B.; Lüscher, B.P.; Fine, M.; Baumann, M.U.; Surbek, D.V.; Bonny, O.; Hediger, M.A. Expression, purification, and structural insights for the human uric acid transporter, GLUT9, using the Xenopus laevis oocytes system. PLoS ONE 2014, 9, e108852. [Google Scholar] [CrossRef] [Green Version]
- Drew, D.; Newstead, S.; Sonoda, Y.; Kim, H.; Von Heijne, G.; Iwata, S. GFP-based optimization scheme for the overexpression and purification of eukaryotic membrane proteins in Saccharomyces cerevisiae. Nat. Protoc. 2008, 3, 784. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Mechref, Y. Comparing MALDI-MS, RP-LC-MALDI-MS and RP-LC-ESI-MS glycomic profiles of permethylated N-glycans derived from model glycoproteins and human blood serum. Electrophoresis 2012, 33, 1768–1777. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Mondello, S.; Kobeissy, F.; Talih, F.; Ferri, R.; Mechref, Y. LC-MS/MS glycomics of idiopathic rapid eye movement sleep behavior disorder. Electrophoresis 2018, 39, 3096–3103. [Google Scholar] [CrossRef]
- Dong, X.; Peng, W.; Yu, C.-Y.; Zhou, S.; Donohoo, K.B.; Tang, H.; Mechref, Y. 8-plex LC–MS/MS Analysis of Permethylated N-Glycans Achieved by Using Stable Isotopic Iodomethane. Anal. Chem. 2019, 91, 11794–11802. [Google Scholar] [CrossRef] [PubMed]
- Banazadeh, A.; Peng, W.; Veillon, L.; Mechref, Y. Carbon Nanoparticles and Graphene Nanosheets as MALDI Matrices in Glycomics: A New Approach to Improve Glycan Profiling in Biological Samples. J. Am. Soc. Mass Spectrom. 2018, 29, 1892–1900. [Google Scholar] [CrossRef] [PubMed]
- Mechref, Y.; Kang, P.; Novotny, M.V. Solid-phase permethylation for glycomic analysis. Methods Mol. Biol. 2009, 534, 53–64. [Google Scholar]
- Zhou, S.; Huang, Y.; Dong, X.; Peng, W.; Veillon, L.; Kitagawa, D.A.S.; Aquino, A.J.A.; Mechref, Y. Isomeric Separation of Permethylated Glycans by Porous Graphitic Carbon (PGC)-LC-MS/MS at High Temperatures. Anal. Chem. 2017, 89, 6590–6597. [Google Scholar] [CrossRef] [PubMed]
- Kang, P.; Mechref, Y.; Klouckova, I.; Novotny, M.V. Solid-phase permethylation of glycans for mass spectrometric analysis. Rapid Commun. Mass Spectrom. 2005, 19, 3421–3428. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Banazadeh, A.; Peng, W.; Mechref, Y. A carbon nanoparticles-based solid-phase purification method facilitating sensitive MALDI-MS analysis of permethylated N-glycans. Electrophoresis 2018, 39, 3087–3095. [Google Scholar] [CrossRef]
- Goetz, J.A.; Novotny, M.V.; Mechref, Y. Enzymatic/chemical release of O-glycans allowing MS analysis at high sensitivity. Anal. Chem. 2009, 81, 9546–9552. [Google Scholar] [CrossRef]
- Yu, C.Y.; Mayampurath, A.; Hu, Y.; Zhou, S.; Mechref, Y.; Tang, H. Automated annotation and quantification of glycans using liquid chromatography-mass spectrometry. Bioinformatics 2013, 29, 1706–1707. [Google Scholar] [CrossRef] [Green Version]
- Psychogios, N.; Hau, D.D.; Peng, J.; Guo, A.C.; Mandal, R.; Bouatra, S.; Sinelnikov, I.; Krishnamurthy, R.; Eisner, R.; Gautam, B. The human serum metabolome. PLoS ONE 2011, 6, e16957. [Google Scholar] [CrossRef] [Green Version]
- Abdi, H.; Williams, L.J. Principal component analysis. Wiley Interdiscip. Rev. Comput. Stat. 2010, 2, 433–459. [Google Scholar] [CrossRef]
- Hennet, T. The galactosyltransferase family. Cell. Mol. Life Sci. 2002, 59, 1081–1095. [Google Scholar] [CrossRef] [PubMed]
- Abdel Rahman, A.M.; Ryczko, M.; Nakano, M.; Pawling, J.; Rodrigues, T.; Johswich, A.; Taniguchi, N.; Dennis, J.W. Golgi N-glycan branching N-acetylglucosaminyltransferases I, V and VI promote nutrient uptake and metabolism. Glycobiology 2015, 25, 225–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.-Y.; Osada, H.; Santamaria-Babi, L.F.; Kannagi, R. Interaction of GATA-3/T-bet transcription factors regulates expression of sialyl Lewis X homing receptors on Th1/Th2 lymphocytes. Proc. Natl. Acad. Sci. USA 2006, 103, 16894–16899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishihara, S.; Iwasaki, H.; Nakajima, K.; Togayachi, A.; Ikehara, Y.; Kudo, T.; Kushi, Y.; Furuya, A.; Shitara, K.; Narimatsu, H. α1, 3-Fucosyltransferase IX (Fut9) determines Lewis X expression in brain. Glycobiology 2003, 13, 445–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuhrer, M. Glycosylation profiling in clinical proteomics: Heading for glycan biomarkers. Expert Rev. Proteomics 2007, 4, 135–136. [Google Scholar] [CrossRef]
- de Freitas Junior, J.C.M.; Morgado-Díaz, J.A. The role of N-glycans in colorectal cancer progression: Potential biomarkers and therapeutic applications. Oncotarget 2016, 7, 19395. [Google Scholar] [CrossRef] [Green Version]
- Arnold, J.N.; Saldova, R.; Galligan, M.C.; Murphy, T.B.; Mimura-Kimura, Y.; Telford, J.E.; Godwin, A.K.; Rudd, P.M. Novel glycan biomarkers for the detection of lung cancer. J. Proteome Res. 2011, 10, 1755–1764. [Google Scholar] [CrossRef]
- Chik, J.H.; Zhou, J.; Moh, E.S.; Christopherson, R.; Clarke, S.J.; Molloy, M.P.; Packer, N.H. Comprehensive glycomics comparison between colon cancer cell cultures and tumours: Implications for biomarker studies. J. Proteomics 2014, 108, 146–162. [Google Scholar] [CrossRef]
- Goso, Y. Malonic acid suppresses mucin-type O-glycan degradation during hydrazine treatment of glycoproteins. Anal. Biochem. 2016, 496, 35–42. [Google Scholar] [CrossRef]
- Kozak, R.P.; Royle, L.; Gardner, R.A.; Bondt, A.; Fernandes, D.L.; Wuhrer, M. Improved nonreductive O-glycan release by hydrazinolysis with ethylenediaminetetraacetic acid addition. Anal. Biochem. 2014, 453, 29–37. [Google Scholar] [CrossRef]
- Nishikaze, T. Sialic acid derivatization for glycan analysis by mass spectrometry. Proc. Jpn. Acad. Ser. B 2019, 95, 523–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.; Veillon, L.; Dong, X.; Huang, Y.; Mechref, Y. Direct comparison of derivatization strategies for LC-MS/MS analysis of N-glycans. Analyst 2017, 142, 4446–4455. [Google Scholar] [CrossRef] [PubMed]
- Petit, D.; Teppa, E.; Mir, A.-M.; Vicogne, D.; Thisse, C.; Thisse, B.; Filloux, C.; Harduin-Lepers, A. Integrative view of α2, 3-sialyltransferases (ST3Gal) molecular and functional evolution in deuterostomes: Significance of lineage-specific losses. Mol. Biol. Evol. 2015, 32, 906–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez Aguilar, A.; Meng, L.; Hou, X.; Li, W.; Moremen, K.W.; Wu, P. Sialyltransferase-based chemoenzymatic histology for the detection of N-and O-glycans. Bioconjugate Chem. 2018, 29, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Wopereis, S.; Lefeber, D.J.; Morava, E.; Wevers, R.A. Mechanisms in protein O-glycan biosynthesis and clinical and molecular aspects of protein O-glycan biosynthesis defects: A review. Clin. Chem. 2006, 52, 574–600. [Google Scholar] [CrossRef] [Green Version]
- Mollicone, R.; Moore, S.E.; Bovin, N.; Garcia-Rosasco, M.; Candelier, J.-J.; Martinez-Duncker, I.; Oriol, R. Activity, splice variants, conserved peptide motifs, and phylogeny of two new α1, 3-fucosyltransferase families (FUT10 and FUT11). J. Biol. Chem. 2009, 284, 4723–4738. [Google Scholar] [CrossRef] [Green Version]
 , N-acetylglucosamine (GlcNAc);
, N-acetylglucosamine (GlcNAc);  , Galactose (Gal);
, Galactose (Gal);  , Fucose (Fuc);
, Fucose (Fuc);  , Mannose (Man);
, Mannose (Man);  , N-acetylneuraminic acid (NeuAc/Sialic Acid). *: p-value < 0.05; **: p-value < 0.005; ***: p-value < 0.0005.
, N-acetylglucosamine (GlcNAc); , Galactose (Gal); , Fucose (Fuc); , Mannose (Man); , N-acetylneuraminic acid (NeuAc/Sialic Acid). *: p-value < 0.05; **: p-value < 0.005; ***: p-value < 0.0005.
, N-acetylneuraminic acid (NeuAc/Sialic Acid). *: p-value < 0.05; **: p-value < 0.005; ***: p-value < 0.0005.
, N-acetylglucosamine (GlcNAc); , Galactose (Gal); , Fucose (Fuc); , Mannose (Man); , N-acetylneuraminic acid (NeuAc/Sialic Acid). *: p-value < 0.05; **: p-value < 0.005; ***: p-value < 0.0005.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glycan Type | Plasmax-DMEM | p-Value (Plasmax/DMEM) | Plasmax-EMEM | p-Value (Plasmax/EMEM) | EMEM-DMEM | p-Value (EMEM/DMEM) |
|---|---|---|---|---|---|---|
| Expression Discrepancies of Different Types of N-Glycans Derived from MDA-MB-231BR Cell Line | ||||||
| High Mannose | +3.2% (± 3.3%) | 0.36 | −0.6% (±2.5%) | 0.86 | +3.8% (±2.1%) | 0.02 |
| Sialylated | +0.1% (±0.4%) | 0.68 | −0.6% (±0.6%) | 0.38 | +0.7% (±1.1%) | 0.28 |
| Fucosylated | −1.7% (±0.1%) | 0.61 | −0.5% (±2.6%) | 0.90 | −1.2% (±2.9%) | 0.63 |
| Sia & Fuc | −0.5% (±0.4%) | 0.65 | −2.9% (±2.6%) | 0.26 | +2.4% (±4.3%) | 0.34 |
| Other | −1.2% (±0.6%) | 0.09 | +4.6% (±0.4%) | 2 × 10−4 | −5.7% (±0.6%) | 2 × 10−4 |
| Expression Discrepancies of Different Types of N-Glycans Derived from CRL-1620 Cell Line | ||||||
| High Mannose | −2.7% (±3.1%) | 0.20 | −5.0% (±0.7%) | 3 × 10−4 | +2.4% (±1.5%) | 0.25 |
| Sialylated | +1.9% (±0.3%) | 1 × 10−3 | −0.1% (±0.2%) | 0.80 | +2.0% (±0.3%) | 1 × 10−3 |
| Fucosylated | −4.3% (±2.3%) | 0.04 | +3.2% (±1.3%) | 0.04 | −7.5% (±3.6%) | 0.01 |
| Sia & Fuc | +2.5% (±0.6%) | 9 × 10−4 | +0.5% (±0.3%) | 0.06 | +1.9% (±0.3%) | 2 × 10−3 |
| Other | +2.8% (±0.4%) | 1 × 10−3 | +1.5% (±0.4%) | 0.02 | +1.3% (±0.2%) | 0.06 |
| Glycan Type | Plasmax-DMEM | p-Value (Plasmax/DMEM) | Plasmax-EMEM | p-Value (Plasmax/EMEM) | EMEM-DMEM | p-Value (EMEM/DMEM) |
|---|---|---|---|---|---|---|
| Expression Discrepancies of Different Types of O-Glycans Derived from MDA-MB-231BR Cell Line | ||||||
| Sialylated | −6.4% (±1.7%) | 4 × 10−3 | +3.4% (±0.9%) | 0.05 | −9.8% (±1.0%) | 7 × 10−4 |
| Fucosylated | −1.4% (±0.8%) | 0.40 | +8.1% (±1.0%) | 1 × 10−4 | −9.6% (±2.0%) | 3 × 10−3 |
| Sia & Fuc | +16.3% (±7.9%) | 0.01 | +2.8% (±8.7%) | 0.74 | +13.2% (±11.7%) | 0.15 |
| Other | −8.2% (±4.1%) | 0.06 | −14.3% (±13.8%) | 0.16 | +6.1% (±12.2%) | 0.48 |
| Expression Discrepancies of Different Types of O-Glycans Derived from CRL-1620 Cell Line | ||||||
| Sialylated | +15.4% (±6.0%) | 0.01 | +6.7% (±6.0%) | 0.24 | +8.7% (±6.1%) | 0.06 |
| Fucosylated | −6.2% (±2.2%) | 4 × 10−3 | −6.3% (±0.7%) | 5 × 10−5 | +0.1% (±0.7%) | 0.95 |
| Sia & Fuc | −10.1% (±3.9%) | 0.02 | −27.9% (±1.6%) | 1 × 10−4 | +17.8% (±5.0%) | 4 × 10−3 |
| Other | +1.0% (±1.0%) | 0.86 | +27.5% (±5.3%) | 3 × 10−3 | −26.5% (±4.4%) | 3 × 10−3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Peng, W.; Yu, A.; Fokar, M.; Mechref, Y. Glycome Profiling of Cancer Cell Lines Cultivated in Physiological and Commercial Media. Biomolecules 2022, 12, 743. https://doi.org/10.3390/biom12060743
Wang J, Peng W, Yu A, Fokar M, Mechref Y. Glycome Profiling of Cancer Cell Lines Cultivated in Physiological and Commercial Media. Biomolecules. 2022; 12(6):743. https://doi.org/10.3390/biom12060743
Chicago/Turabian StyleWang, Junyao, Wenjing Peng, Aiying Yu, Mohamed Fokar, and Yehia Mechref. 2022. "Glycome Profiling of Cancer Cell Lines Cultivated in Physiological and Commercial Media" Biomolecules 12, no. 6: 743. https://doi.org/10.3390/biom12060743
APA StyleWang, J., Peng, W., Yu, A., Fokar, M., & Mechref, Y. (2022). Glycome Profiling of Cancer Cell Lines Cultivated in Physiological and Commercial Media. Biomolecules, 12(6), 743. https://doi.org/10.3390/biom12060743