
Early Biological Modulations Resulting from 1-Week Venlafaxine Exposure of Marine Mussels Mytilus galloprovincialis Determined by a Metabolomic Approach

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Venlafaxine Quantification
2.2. Metabolite Modulation and Pathway Analysis
2.3. Enrichment Analysis
3. Discussion
3.1. Early Responses after VLF Exposure
3.2. Sex-Dependent Responses
3.3. Temporal Variation in the Effects
3.4. Mode of Action of VLF on M. galloprovincialis, Impact on Neurotransmitters
4. Materials and Methods
4.1. Chemicals
4.2. Animals and Experimentation Design
4.3. Metabolomics Sample Analysis
4.3.1. Tissue Sample Preparation
4.3.2. Data Acquisition and Quality Control
4.3.3. Data Processing and Statistical Analysis
4.3.4. Metabolite Annotation and Identification
4.3.5. Enrichment Analysis
4.4. Venlafaxine Quantification
4.4.1. Sample Preparation
4.4.2. Data Acquisition
4.4.3. Identification and Quantification of VLF
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fernandes, M.J.; Paíga, P.; Silva, A.; Llaguno, C.P.; Carvalho, M.; Vázquez, F.M.; Delerue-Matos, C. Antibiotics and antidepressants occurrence in surface waters and sediments collected in the north of Portugal. Chemosphere 2020, 239, 124729. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, E.; Hernández, F.; Ibáñez, M.; Rico, A.; Pitarch, E.; Bijlsma, L. Occurrence and ecological risks of pharmaceuticals in a Mediterranean river in Eastern Spain. Environ. Int. 2020, 144, 106004. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Rubio, J.; Rodríguez-Gil, J.L.; Postigo, C.; Mastroianni, N.; López de Alda, M.; Barceló, D.; Valcárcel, Y. Psychoactive pharmaceuticals and illicit drugs in coastal waters of North-Western Spain: Environmental exposure and risk assessment. Chemosphere 2019, 224, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Desbiolles, F.; Malleret, L.; Tiliacos, C.; Wong-Wah-Chung, P.; Laffont-Schwob, I. Occurrence and ecotoxicological assessment of pharmaceuticals: Is there a risk for the Mediterranean aquatic environment? Sci. Total Environ. 2018, 639, 1334–1348. [Google Scholar] [CrossRef] [PubMed]
- Björlenius, B.; Ripszám, M.; Haglund, P.; Lindberg, R.H.; Tysklind, M.; Fick, J. Pharmaceutical residues are widespread in Baltic Sea coastal and offshore waters–Screening for pharmaceuticals and modelling of environmental concentrations of carbamazepine. Sci. Total Environ. 2018, 633, 1496–1509. [Google Scholar] [CrossRef]
- Gevao, B.; Uddin, S.; Dupont, S. Baseline concentrations of pharmaceuticals in Kuwait’s coastal marine environment. Mar. Pollut. Bull. 2021, 173, 113040. [Google Scholar] [CrossRef]
- Sousa, J.C.G.; Barbosa, M.O.; Ribeiro, A.R.L.; Ratola, N.; Pereira, M.F.R.; Silva, A.M.T. Distribution of micropollutants in estuarine and sea water along the Portuguese coast. Mar. Pollut. Bull. 2020, 154, 111120. [Google Scholar] [CrossRef]
- Álvarez-Muñoz, D.; Rodríguez-Mozaz, S.; Maulvault, A.L.; Tediosi, A.; Fernández-Tejedor, M.; Van den Heuvel, F.; Kotterman, M.; Marques, A.; Barceló, D. Occurrence of pharmaceuticals and endocrine disrupting compounds in macroalgaes, bivalves, and fish from coastal areas in Europe. Environ. Res. 2015, 143, 56–64. [Google Scholar] [CrossRef]
- Martínez Bueno, M.J.; Boillot, C.; Munaron, D.; Fenet, H.; Casellas, C.; Gómez, E. Occurrence of venlafaxine residues and its metabolites in marine mussels at trace levels: Development of analytical method and a monitoring program. Anal. Bioanal. Chem. 2014, 406, 601–610. [Google Scholar] [CrossRef]
- Bisesi, J.H.; Bridges, W.; Klaine, S.J. Reprint of: Effects of the antidepressant venlafaxine on fish brain serotonin and predation behavior. Aquat. Toxicol. 2014, 151, 88–96. [Google Scholar] [CrossRef]
- Galus, M.; Jeyaranjaan, J.; Smith, E.; Li, H.; Metcalfe, C.; Wilson, J.Y. Chronic effects of exposure to a pharmaceutical mixture and municipal wastewater in zebrafish. Aquat. Toxicol. 2013, 132–133, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Melvin, S.D. Effect of antidepressants on circadian rhythms in fish: Insights and implications regarding the design of behavioural toxicity tests. Aquat. Toxicol. 2017, 182, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Canela, C.; Rovira García, X.; Martínez-Jerónimo, F.; Marcé, R.M.; Barata, C. Analysis of neurotransmitters in Daphnia magna affected by neuroactive pharmaceuticals using liquid chromatography-high resolution mass spectrometry. Environ. Pollut. 2019, 254, 113029. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.T.; Rudolph, R.L.; Preskorn, S.H. Evidence of the dual mechanisms of action of venlafaxine. Arch. Gen. Psychiatry 2000, 57, 503–509. [Google Scholar] [CrossRef]
- Best, C.; Melnyk-Lamont, N.; Gesto, M.; Vijayan, M.M. Environmental levels of the antidepressant venlafaxine impact the metabolic capacity of rainbow trout. Aquat. Toxicol. 2014, 155, 190–198. [Google Scholar] [CrossRef]
- Bundy, J.G.; Davey, M.P.; Viant, M.R. Environmental metabolomics: A critical review and future perspectives. Metabolomics 2009, 5, 3–21. [Google Scholar] [CrossRef]
- Tikunov, A.; Stoskopf, M.; Macdonald, J. Fluxomics of the Eastern Oyster for Environmental Stress Studies. Metabolites 2014, 4, 53–70. [Google Scholar] [CrossRef] [Green Version]
- Pomfret, S.M.; Brua, R.B.; Izral, N.M.; Yates, A.G. Metabolomics for biomonitoring: An evaluation of the metabolome as an indicator of aquatic ecosystem health. Environ. Rev. 2020, 28, 89–98. [Google Scholar] [CrossRef]
- Bonnefille, B.; Gomez, E.; Alali, M.; Rosain, D.; Fenet, H.; Courant, F. Metabolomics assessment of the effects of diclofenac exposure on Mytilus galloprovincialis: Potential effects on osmoregulation and reproduction. Sci. Total Environ. 2018, 613–614, 611–618. [Google Scholar] [CrossRef]
- Campillo, J.A.; Sevilla, A.; González-Fernández, C.; Bellas, J.; Bernal, C.; Cánovas, M.; Albentosa, M. Metabolomic responses of mussel Mytilus galloprovincialis to fluoranthene exposure under different nutritive conditions. Mar. Environ. Res. 2019, 144, 194–202. [Google Scholar] [CrossRef]
- Dumas, T.; Bonnefille, B.; Gomez, E.; Boccard, J.; Castro, N.A.; Fenet, H.; Courant, F. Metabolomics approach reveals disruption of metabolic pathways in the marine bivalve Mytilus galloprovincialis exposed to a WWTP effluent extract. Sci. Total Environ. 2020, 712, 136551. [Google Scholar] [CrossRef] [PubMed]
- Lubet, P. Exposé synoptique des données biologiques sur la moule Mytilus galloprovincialis (L.). Synop. FAO Pêches 1973, 88, 8. [Google Scholar]
- Mezzelani, M.; Fattorini, D.; Gorbi, S.; Nigro, M.; Regoli, F. Human pharmaceuticals in marine mussels: Evidence of sneaky environmental hazard along Italian coasts. Mar. Environ. Res. 2020, 162, 105137. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.J.G.; Martins, M.C.; Pereira, A.M.P.T.; Meisel, L.M.; Gonzalez-Rey, M.; Bebianno, M.J.; Lino, C.M.; Pena, A. Uptake, accumulation and metabolization of the antidepressant fluoxetine by Mytilus galloprovincialis. Environ. Pollut. 2016, 213, 432–437. [Google Scholar] [CrossRef] [Green Version]
- Farrington, J.W.; Tripp, B.W.; Tanabe, S.; Subramanian, A.; Sericano, J.L.; Wade, T.L.; Knap, A.H.; Edward, D. Goldberg’s proposal of “the Mussel Watch”: Reflections after 40 years. Mar. Pollut. Bull. 2016, 110, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Franzellitti, S.; Buratti, S.; Valbonesi, P.; Fabbri, E. The mode of action (MOA) approach reveals interactive effects of environmental pharmaceuticals on Mytilus galloprovincialis. Aquat. Toxicol. 2013, 140–141, 249–256. [Google Scholar] [CrossRef]
- Świacka, K.; Maculewicz, J.; Smolarz, K.; Szaniawska, A.; Caban, M. Mytilidae as model organisms in the marine ecotoxicology of pharmaceuticals–A review. Environ. Pollut. 2019, 254, 113082. [Google Scholar] [CrossRef]
- Ariza-Castro, N.; Courant, F.; Dumas, T.; Marion, B.; Fenet, H.; Gomez, E. Elucidating Venlafaxine Metabolism in the Mediterranean Mussel (Mytilus galloprovincialis) through Combined Targeted and Non-Targeted Approaches. Sci. Total Environ. 2021, 779, 146387. [Google Scholar] [CrossRef]
- Want, E.J.; Masson, P.; Michopoulos, F.; Wilson, I.D.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Loftus, N.; Holmes, E.; Nicholson, J.K. Global metabolic profiling of animal and human tissues via UPLC-MS. Nat. Protoc. 2013, 8, 17–32. [Google Scholar] [CrossRef]
- Blanco, A.; Blanco, G. Amino Acid Metabolism. In Medical Biochemistry; Elsevier: Amsterdam, The Netherlands, 2017; pp. 367–399. [Google Scholar]
- Caylak, E.; Aytekin, M.; Halifeoglu, I. Antioxidant effects of methionine, α-lipoic acid, N-acetylcysteine and homocysteine on lead-induced oxidative stress to erythrocytes in rats. Exp. Toxicol. Pathol. 2008, 60, 289–294. [Google Scholar] [CrossRef]
- Asensi, M.; Sastre, J.; Pallardo, F.V.; Lloret, A.; Lehner, M.; Garcia-De-La Asuncion, J.; Viña, J. Ratio of reduced to oxidized glutathione as indicator of oxidative stress status and DNA damage. Methods Enzymol. 1999, 299, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Hooper, C.; Day, R.; Slocombe, R.; Handlinger, J.; Benkendorff, K. Stress and immune responses in abalone: Limitations in current knowledge and investigative methods based on other models. Fish Shellfish Immunol. 2007, 22, 363–379. [Google Scholar] [CrossRef] [PubMed]
- Lagos, L.; Herrera, M.; Sánchez-Lazo, C.; Martínez-Pita, I. Effect of larval stocking density on growth, survival and whole body cortisol of the Mediterranean mussel Mytilus galloprovincialis (Lamarck, 1819) larvae reared under laboratory conditions. Aquac. Res. 2015, 46, 1648–1656. [Google Scholar] [CrossRef]
- Ottaviani, E.; Franceschi, C. The invertebrate phagocytic immunocyte: Clues to a common evolution of immune and neuroendocrine systems. Immunol. Today 1997, 18, 169–174. [Google Scholar] [CrossRef]
- Shi, W.; Han, Y.; Guan, X.; Rong, J.; Su, W.; Zha, S.; Tang, Y.; Du, X.; Liu, G. Fluoxetine suppresses the immune responses of blood clams by reducing haemocyte viability, disturbing signal transduction and imposing physiological stress. Sci. Total Environ. 2019, 683, 681–689. [Google Scholar] [CrossRef]
- Calder, P.C. Branched-chain amino acids and immunity. J. Nutr. 2006, 136, 288S–293S. [Google Scholar] [CrossRef]
- Liu, X.; Sun, H.; Wang, Y.; Ma, M.; Zhang, Y. Gender-specific metabolic responses in hepatopancreas of mussel Mytilus galloprovincialis challenged by Vibrio harveyi. Fish Shellfish Immunol. 2014, 40, 407–413. [Google Scholar] [CrossRef]
- Lisowska, K.A.; Pietruczuk, K.; Szałach, Ł.P. Interlinking antidepressants and the immune system. In The Neuroscience of Depression; Elsevier: Amsterdam, The Netherlands, 2021; pp. 143–151. [Google Scholar]
- Aiello, E. Nervous and Chemical Stimulation of Gill Cilia in Bivalve Molluscs. Physiol. Zool. 1970, 43, 60–70. [Google Scholar] [CrossRef]
- Saimi, Y.; Murakami, A.; Takahashi, K. Electrophysiological correlates of nervous control of ciliary arrest response in the gill epithelial cells of Mytilus. Comp. Biochem. Physiol.-Part A Physiol. 1983, 74, 499–506. [Google Scholar] [CrossRef]
- Ram, J.L.; Moore, D.; Putchakayala, S.; Paredes, A.A.; Ma, D.; Croll, R.P. Serotonergic responses of the siphons and adjacent mantle tissue of the zebra mussel, Dreissena polymorpha. Comp. Biochem. Physiol.-C Pharmacol. Toxicol. Endocrinol. 1999, 124, 211–220. [Google Scholar] [CrossRef]
- Gex-Fabry, M.; Rudaz, S.; Balant-Gorgia, A.E.; Brachet, A.; Veuthey, J.L.; Balant, L.P.; Bertschy, G. Steady-state concentration of venlafaxine enantiomers: Model-based analysis of between-patient variability. Eur. J. Clin. Pharmacol. 2002, 58, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Physicians’ Desk Reference Inc. (Ed.) Physicians’ Desk Reference, 63rd ed.; Thomson Reuters: Montvale, NJ, USA, 2009; ISBN 978-1563637032. [Google Scholar]
- Bano, S.; Akhter, S.; Afridi, M.I. Gender based response to fluoxetine hydrochloride medication in endogenous depression. J. Coll. Physicians Surg. Pak. 2004, 14, 161–165. [Google Scholar] [PubMed]
- McGrath, P.J.; Stewart, J.W.; Quitkin, F.M.; Chen, Y.; Alpert, J.E.; Nierenberg, A.A.; Fava, M.; Cheng, J.; Petkova, E. Predictors of relapse in a prospective study of fluoxetine treatment of major depression. Am. J. Psychiatry 2006, 163, 1542–1548. [Google Scholar] [CrossRef] [PubMed]
- Dumas, T.; Boccard, J.; Gomez, E.; Fenet, H.; Courant, F. Multifactorial Analysis of Environmental Metabolomic Data in Ecotoxicology: Wild Marine Mussel Exposed to WWTP Effluent as a Case Study. Metabolites 2020, 10, 269. [Google Scholar] [CrossRef]
- Ji, C.; Wu, H.; Wei, L.; Zhao, J.; Yu, J. Proteomic and metabolomic analysis reveal gender-specific responses of mussel Mytilus galloprovincialis to 2,2’,4,4’-tetrabromodiphenyl ether (BDE 47). Aquat. Toxicol. 2013, 140–141, 449–457. [Google Scholar] [CrossRef]
- Burg, M.B.; Ferraris, J.D. Intracellular organic osmolytes: Function and regulation. J. Biol. Chem. 2008, 283, 7309–7313. [Google Scholar] [CrossRef] [Green Version]
- Holbrook, D.J. Effects of toxicants on nucleic acid and protein metabolism. In Introduction to Biochemical Toxicology; Hodgson, E., Smart, R., Eds.; John Wiley & Sons: New York, NY, USA, 1987; pp. 325–342. [Google Scholar]
- Gomez, E.; Boillot, C.; Martinez Bueno, M.J.; Munaron, D.; Mathieu, O.; Courant, F.; Fenet, H. In vivo exposure of marine mussels to venlafaxine: Bioconcentration and metabolization. Environ. Sci. Pollut. Res. 2021, 28, 68862–68870. [Google Scholar] [CrossRef]
- Cappello, T.; De Marco, G.; Oliveri Conti, G.; Giannetto, A.; Ferrante, M.; Mauceri, A.; Maisano, M. Time-dependent metabolic disorders induced by short-term exposure to polystyrene microplastics in the Mediterranean mussel Mytilus galloprovincialis. Ecotoxicol. Environ. Saf. 2021, 209, 111780. [Google Scholar] [CrossRef]
- Brew, D.W.; Black, M.C.; Santos, M.; Rodgers, J.; Henderson, W.M. Metabolomic Investigations of the Temporal Effects of Exposure to Pharmaceuticals and Personal Care Products and Their Mixture in the Eastern Oyster (Crassostrea virginica). Environ. Toxicol. Chem. 2020, 39, 419–436. [Google Scholar] [CrossRef]
- Caccia, S. Metabolism of the newer antidepressants: An overview of the pharmacological and pharmacokinetic implications. Clin. Pharmacokinet. 1998, 34, 281–302. [Google Scholar] [CrossRef]
- David, D.J.; Gardier, A.M. Les bases de pharmacologie fondamentale du système sérotoninergique: Application à la réponse antidépressive [The pharmacological basis of the serotonin system: Application to antidepressant response]. Encephale 2016, 42, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Haddjeri, N.; Blier, P. Effects of sustained (±) pindolol administration on serotonin neurotransmission in rats. J. Psychiatry Neurosci. 2000, 25, 378–388. [Google Scholar] [PubMed]
- Tierney, A.J. Structure and function of invertebrate 5-HT receptors: A review. Comp. Biochem. Physiol.-A Mol. Integr. Physiol. 2001, 128, 791–804. [Google Scholar] [CrossRef]
- Ram, J.L.; Fong, P.; Croll, R.P.; Nichols, S.J.; Wall, D. The zebra mussel (dreissena polymorpha), a new pest in North America: Reproductive mechanisms as possible targets of control strategies. Invertebr. Reprod. Dev. 1992, 22, 77–86. [Google Scholar] [CrossRef]
- Hirai, S.; Kishimoto, T.; Kadam, A.L.; Kanatani, H.; Koide, S.S. Induction of spawning and oocyte maturation by 5-hydroxytryptamine in the surf clam. J. Exp. Zool. 1988, 245, 318–321. [Google Scholar] [CrossRef]
- Kadam, A.L.; Koide, S.S. Serotonin analogs and spisula oocyte maturation. Invertebr. Reprod. Dev. 1989, 15, 225–227. [Google Scholar] [CrossRef]
- Osanai, K.; Kuraishi, R. Response of oocytes to meiosis-inducing agents in pelecypods [Crassostrea gigas, Tapes philippinarum and Mytilus edulis]. Bull. Mar. Biol. Stn. Asamushi-Tohoku Univ. (Jpn.) 1988, 18, 45–56. [Google Scholar]
- Gibbons, M.C.; Castagna, M. Serotonin as an inducer of spawning in six bivalve species. Aquaculture 1984, 40, 189–191. [Google Scholar] [CrossRef]
- Fong, P.P. Zebra mussel spawning is induced in low concentrations of putative serotonin reuptake inhibitors. Biol. Bull. 1998, 194, 143–149. [Google Scholar] [CrossRef]
- Lazzara, R.; Blázquez, M.; Porte, C.; Barata, C. Low environmental levels of fluoxetine induce spawning and changes in endogenous estradiol levels in the zebra mussel Dreissena polymorpha. Aquat. Toxicol. 2012, 106–107, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Starovlah, I.M.; Radovic Pletikosic, S.M.; Kostic, T.S.; Andric, S.A. Reduced spermatozoa functionality during stress is the consequence of adrenergic-mediated disturbance of mitochondrial dynamics markers. Sci. Rep. 2020, 10, 16813. [Google Scholar] [CrossRef] [PubMed]
- Bymaster, F.P.; Dreshfield-Ahmad, L.J.; Threlkeld, P.G.; Shaw, J.L.; Thompson, L.; Nelson, D.L.; Hemrick-Luecke, S.K.; Wong, D.T. Comparative affinity of duloxetine and venlafaxine for serotonin and norepinephrine transporters in vitro and in vivo, human serotonin receptor subtypes, and other neuronal receptors. Neuropsychopharmacology 2001, 25, 871–880. [Google Scholar] [CrossRef]
- Roeder, T. Octopamine in invertebrates. Prog. Neurobiol. 1999, 59, 533–561. [Google Scholar] [CrossRef]
- Cappello, T.; Fernandes, D.; Maisano, M.; Casano, A.; Bonastre, M.; Bebianno, M.J.; Mauceri, A.; Fasulo, S.; Porte, C. Sex steroids and metabolic responses in mussels Mytilus galloprovincialis exposed to drospirenone. Ecotoxicol. Environ. Saf. 2017, 143, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.; Gong, Z.; Kelly, B.C. Assessing biological effects of fluoxetine in developing zebrafish embryos using gas chromatography-mass spectrometry based metabolomics. Chemosphere 2017, 188, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Faggio, C.; Tsarpali, V.; Dailianis, S. Mussel digestive gland as a model tissue for assessing xenobiotics: An overview. Sci. Total Environ. 2018, 636, 220–229. [Google Scholar] [CrossRef]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A Cross-Platform Toolkit for Mass Spectrometry and Proteomics. Nature Biotechnology 2012, 30, 918–920. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis: Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Wishart, D.S. MetPA: A Web-Based Metabolomics Tool for Pathway Analysis and Visualization. Bioinformatics 2010, 26, 2342–2344. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| Pathway | Metabolite | Adduct | m/z | RT (min) | Male Mussels | Female Mussels | Annotation Level | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 3 | 7 | 1 | 3 | 7 | ||||||
| Acetylcholine synthesis | Glycerophosphorylcholine | M+H+ | 258.1101 | 1.53 | - | - | - |  | | - | 1 |
| Propionylcholine | M+ | 160.1337 | 4.46 |  | ↘ | - | - | - | ↗ | 2 | |
| Alanine, aspartate and glutamate metabolism | Aspartylphenylalanine † | M+H+ | 281.1132 | 11.03 | |  | | | | ↘ | 2 |
| Gamma-l-glutamyl-l-leucine | M+H+ | 261.1445 | 9.63 | - | - | - | |  | ↗ | 1 | |
| Isovalerylalanine | M+H+ | 174.1125 | 9.44 | - | - | - | | | - | 2 | |
| L-Alanine †,° | M+H+ | 90.0546 | 1.45 | - | ↘ | - | - | - | | 1 | |
| L-Arginine † | M+H+ | 175.1190 | 2.63 | - | - | - | ↗ | | | 1 | |
| N-acetyl-L-aspartic acid | M−H− | 174.0408 | 1.48 | | - | ↗ | - | | | 1 | |
| N-Acetyl-L-glutamic acid | M−H− | 188.0565 | 1.85 | - | - | - | | - | - | 1 | |
| Pyrrolidin-2-one † | M+Na+ | 108.0420 | 1.85 | | - | ↘ | - | - | - | 1 | |
| Succinic acid † | M−H− | 117.0193 | 1.89 | ↘ | - | | ↘ | - | ↗ | 1 | |
| Amino Sugar Metabolism | Glucosamine | M+H+ | 180.0867 | 2.07 | - | - | - | - | - | | 2 |
| N-Acetyl-D-glucosamine | M+FA−H− | 266.0881 | 1.21 | - | - | | - | - | ↗ | 1 | |
| or N-Acetylmannosamine | |||||||||||
| Arginine and Proline Metabolism and derivatives | 2-Oxoarginine | M−H2O−H− | 154.0617 | 1.92 | - | - | - | - | | - | 2 |
| 4-(Glutamylamino) butanoate | M+H+ | 233.1132 | 3.13 | - | - | - | - | | | 2 | |
| Gamma-glutamyl-L-putrescine | M−H2O+H+ | 200.1388 | 3.94 | ↗ | | - | - | ↘ | ND | 2 | |
| L-4-Hydroxyglutamate semialdehyde | M−H2O−H− | 128.0348 | 1.56 | - | ↘ | - | - | ↗ | | 2 | |
| L-Arginine † | M+H+ | 175.1190 | 2.63 | - | - | - | ↗ | | | 1 | |
| L-Aspartate-semialdehyde † | M−H− | 116.0353 | 1.44 | ↘ | - | ↗ | - | | - | 2 | |
| L-Glutamine † | M+H+ | 147.0764 | 1.26 | - | ↘ | - | - | | | 2 | |
| N-alpha-acetylcitrulline | M−H− | 216.0990 | 1.47 | ND | ND | ND | ND | ND | | 1 | |
| Pyrrolidin-2-one † | M+Na+ | 108.0420 | 1.85 | | - | ↘ | - | - | - | 1 | |
| Sarcosine †,° | M+H+ | 90.05495 | 1.45 | - | ↘ | - | - | - | | 1 | |
| Succinic acid † | M−H− | 117.0193 | 1.89 | ↘ | - | | ↘ | - | ↗ | 1 | |
| Symmetric dimethylarginine | M+H+ | 203.1503 | 3.45 | - | | ↘ | - | | | 2 | |
| beta-Alanine metabolism | Hydroxypropionic acid † | M−H− | 89.0244 | 1.55 | - | - | | - | - | ↘ | 2 |
| Pyrrolidin-2-one † | M+Na+ | 108.0420 | 1.85 | | - | ↘ | - | - | - | 1 | |
| Uracil † | M−H− | 111.0200 | 1.52 | - | - | - | | - | - | 1 | |
| Biotin metabolism | L-Lysine † | M+H+ | 147.1128 | 2.50 | - | ↘ | - | - | | ↘ | 1 |
| Pimelic acid | M−H− | 159.0663 | 8.54 | ↗ | ↘ | - | | ↗ | - | 1 | |
| Cysteine and methionine metabolism | 2-Hydroxyphenethylamine | M−H2O+H+ | 120.0802 | 7.81 | | | - | | | | 2 |
| L-Alanine † | M+H+ | 90.0550 | 1.45 | - | ↘ | - | - | - | | 1 | |
| L-Aspartate-semialdehyde † | M−H− | 116.0353 | 1.44 | ↘ | - | ↗ | - | | - | 2 | |
| L-Methionine † | M+H+ | 150.0583 | 2.57 | | - | - | | | ↘ | 1 | |
| Glutathione metabolism | 5-l-glutamyl-l-alanine | M+H+ | 219.0976 | 1.69 | - | - | - | | - | ND | 2 |
| Cysteineglutathione disulfide | M−H− | 425.0806 | 1.30 | ND | ND | ND | | | ND | 2 | |
| Pyroglutamic acid | M−H− | 128.0353 | 1.56 | - | ↘ | - | - | ↗ | | 1 | |
| Glycine, serine and threonine metabolism | Aminoacetone | M+FA−H− | 118.0510 | 1.33 | - | - | - | - | | - | 2 |
| DL-2-hydroxybutyric acid | M−H− | 103.0401 | 2.09 | ↗ | - | - | | - | - | 1 | |
| Glycine † | M+H+ | 76.0393 | 1.34 | ↘ | - | | - | | - | 2 | |
| Hydroxypyruvic acid | M−H− | 103.0037 | 1.52 | - | - | - | ↗ | | ↘ | 2 | |
| L-allothreonine | M−H− | 118.0510 | 1.29 | - | - | - | - | | - | 1 | |
| L-Arginine † | M+H+ | 175.1190 | 2.63 | - | - | - | ↗ | | | 1 | |
| L-Aspartate-semialdehyde † | M−H− | 116.0353 | 1.44 | ↘ | - | ↗ | - | | - | 2 | |
| L-Homoserine | M+H+ | 120.0655 | 1.38 | | ↘ | - | - | - | - | 1 | |
| L-Methionine † | M+H+ | 150.0583 | 2.57 | | - | - | | | ↘ | 1 | |
| L-Tryptophan † | M+H+ | 205.0972 | 11.58 | - | | - | - | | - | 1 | |
| N-Acetyl-L-threonine | M−H− | 160.0615 | 1.50 | ↘ | - | | | | - | 1 | |
| Histidine metabolism | Carnosine | M+H+ | 227.1139 | 3.56 | ND | ND | ND | - | | ND | 2 |
| Histamine | M+H+ | 112.0869 | 5.84 | | | ↗ | | ↘ | ↗ | 1 | |
| Methylimidazoleacetic acid | M+H+ | 141.0659 | 3.10 | - | - | - | - | | ND | 1 | |
| Lysine biosynthesis or degradation | Aminoadipic acid | M+H+ | 162.0761 | 1.59 | - | - | - | | - | - | 1 |
| Glutaric acid | M−H− | 131.0350 | 2.48 | ↗ | - | - | | ↗ | - | 1 | |
| L-Aspartate-semialdehyde | M−H− | 116.0353 | 1.44 | ↘ | - | ↗ | - | | - | 2 | |
| L-Lysine † | M+H+ | 147.1128 | 2.50 | - | ↘ | - | - | | ↘ | 1 | |
| N-Succinyl-2-amino-6-ketopimelate | M+H+ | 290.0870 | 2.24 | - | ↘ | - | | | - | 2 | |
| N6-Acetyl-L-lysine | M+H+ | 189.1234 | 1.89 | - | - | ↗ | - | | - | 1 | |
| Pipecolic acid | M+H+ | 130.0863 | 2.04 | ↗ | - | ↗ | ↘ | ↘ | | 1 | |
| Nicotinate and Nicotinamide Metabolism | Niacinamide | M+H+ | 123.0553 | 2.46 | ↗ | | - | | - | - | 1 |
| Nicotinamide N-oxide | M+H+ | 139.0502 | 3.46 | ↗ | - | | - | - | | 2 | |
| Pyrrolidin-2-one † | M+Na+ | 108.0420 | 1.85 | | - | ↘ | - | - | - | 1 | |
| Trigonelline | M+H+ | 138.0550 | 1.45 | - | | - | - | - | - | 2 | |
| Pentose phosphate pathway | Deoxyribose | M+FA−H− | 179.0561 | 1.21 | - | ↗ | | | - | - | 2 |
| Gluconic acid | M−H− | 195.0510 | 1.15 | - | - | | - | - | ↘ | 1 | |
| Ribose 1-phosphate † | M−H2O+H+ | 213.0153 | 2.93 | ↗ | - | - | | ↗ | - | 2 | |
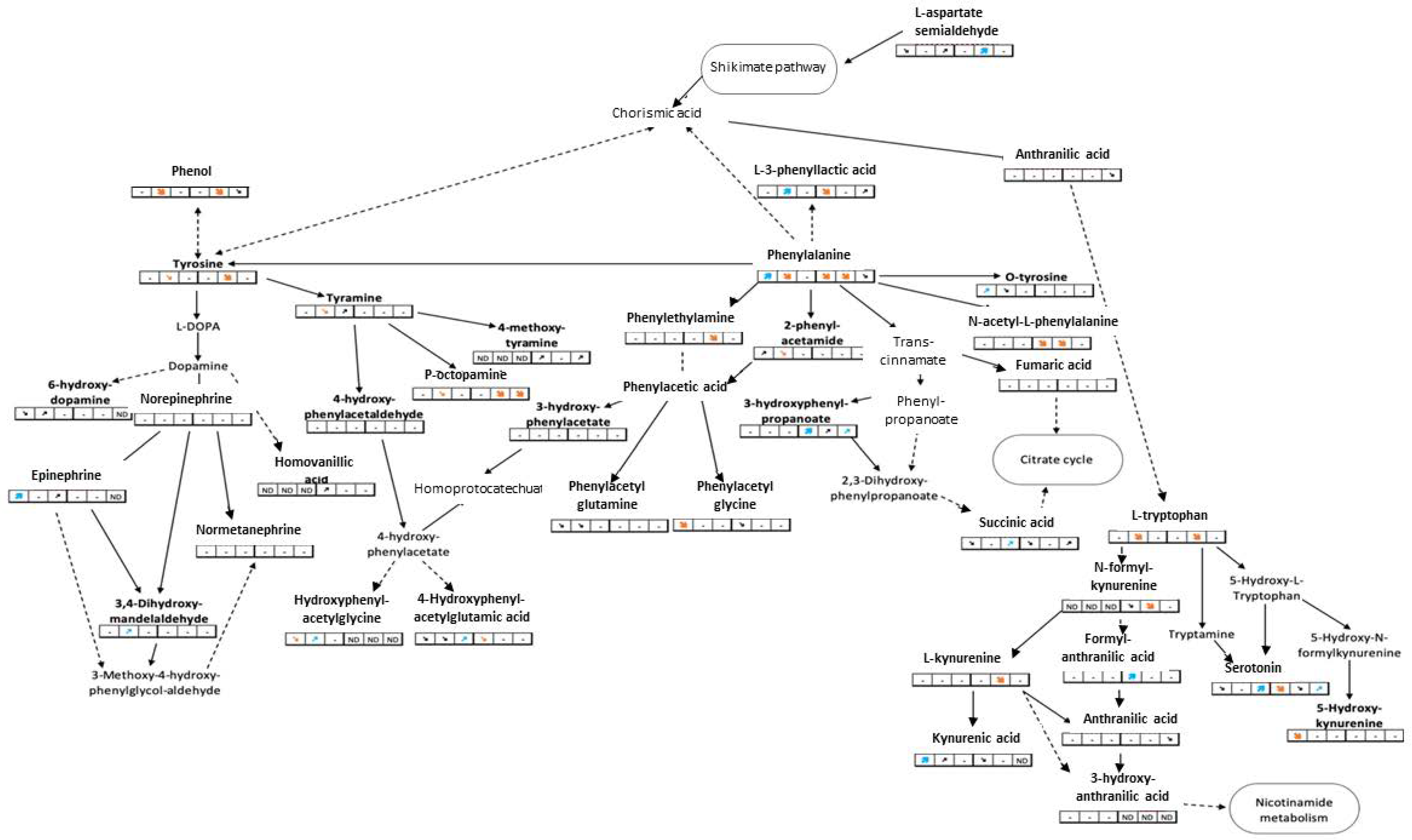
| Phenylalanine metabolism | 2-Phenylacetamide | M+H+ | 136.0757 | 7.97 | ↗ | | - | - | - | - | 1 |
| 3-(3-Hydroxyphenyl)propanoic acid | M−H− | 165.0557 | 10.79 | - | - | - | | ↗ | | 1 | |
| Aspartylphenylalanine † | M+H+ | 281.1132 | 11.03 | | | | | | ↘ | 2 | |
| L-3-phenyllactic acid | M−H− | 165.0557 | 10.48 | - | | - | | - | ↗ | 1 | |
| L-Phenylalanine | M+H+ | 166.0863 | 7.81 | | | - | | | ↘ | 1 | |
| L-Tyrosine † | M+H+ | 182.0812 | 3.54 | - | | - | - | | - | 1 | |
| N-Acetyl-L-phenylalanine | M−H− | 206.0823 | 10.66 | - | - | - | | | - | 1 | |
| Phenylacetylglycine | M−H− | 192.0666 | 9.53 | | - | - | ↘ | - | - | 1 | |
| Phenylethylamine | M+H+ | 122.0964 | 13.93 | - | - | - | - | | - | 1 | |
| Purine metabolism | 2-Hydroxyadenine | M+Na+ | 174.0386 | 2.34 | ↗ | | - | - | - | ↗ | 2 |
| 2′-Deoxyguanosine | M+H+ | 268.1040 | 2.27 | | ↘ | - | ↘ | | ND | 1 | |
| Cyclic AMP | M−H− | 328.0452 | 1.51 | | - | - | | ↘ | ↘ | 1 | |
| Glycine † | M+H+ | 76.0393 | 1.34 | ↘ | - | | - | | - | 2 | |
| Guanine | M+H+ | 152.0567 | 2.33 | | - | - | - | - | - | 1 | |
| Inosine-5′-monophosphate | M−H− | 347.0398 | 1.18 | - | - | ↗ | - | | ND | 1 | |
| Ribose 1-phosphate † | M−H2O+H+ | 213.0153 | 2.93 | ↗ | - | - | | ↗ | - | 2 | |
| Succinyladenosine | M+H+ | 384.1150 | 3.60 | - | ↗ | - | | - | ↘ | 2 | |
| Xanthine | M−H− | 151.0262 | 1.64 | - | | - | | - | - | 1 | |
| Pyrimidine metabolism | 5-Methylcytosine | M−H− | 124.0516 | 2.31 | | | ↗ | ↗ | - | - | 2 |
| Cytosine | M+H+ | 112.0505 | 2.41 | - | ↗ | - | | - | - | 1 | |
| Deoxycytidine | M+H+ | 228.0979 | 3.10 | - | - | ↘ | | | - | 1 | |
| Deoxyinosine | M−H− | 251.0786 | 1.87 | | ↘ | ↘ | | | ↘ | 1 | |
| Dihydrothymine | M+H+ | 129.0659 | 1.97 | ↘ | ↘ | | - | | - | 1 | |
| Hydroxypropionic acid † | M−H− | 89.0244 | 1.55 | - | - | | - | - | ↘ | 1 | |
| L-Glutamine † | M+H+ | 147.0764 | 1.26 | - | ↘ | - | - | | | 1 | |
| Pseudouridine | M−H− | 243.0623 | 1.31 | ND | ND | ND | - | | | 2 | |
| Thymidine | M+FA−H− | 287.0885 | 2.39 | | | - | | | - | 1 | |
| Uracil † | M−H− | 111.0200 | 1.52 | - | - | - | | - | - | 1 | |
| DL-Lactic Acid † | M−H− | 89.0244 | 6.38 | - | - | | - | | ↘ | 1 | |
| Steroid hormone biosynthesis | 4-Methylpentanal | M+ACN+H+ | 142.1226 | 10.07 | - | | - | - | - | - | 2 |
| Cortolone | 2M+H+ | 227.1251 | 13.07 | | - | - | - | - | - | 1 | |
| Tetrahydrocortisol | M+FA−H− | 411.2388 | 12.24 | | - | - | - | - | - | 1 | |
| Tryptophan metabolism | 5-Hydroxykynurenine | M+H+ | 225.0870 | 8.49 | | - | - | - | - | - | 2 |
| DL-kynurenine | M+H+ | 209.0921 | 8.73 | - | - | - | - | | - | 1 | |
| Formylanthranilic acid | M−H− | 164.0353 | 11.46 | - | - | - | | - | - | 1 | |
| Indoleacrylic acid | M+H+ | 188.0706 | 11.58 | - | - | - | - | | - | 2 | |
| Kynurenic acid | M+H+ | 190.0499 | 10.27 | | ↗ | - | ↘ | - | ND | 1 | |
| L-Tryptophan † | M+H+ | 205.0972 | 11.58 | - | | - | - | | - | 1 | |
| N′-Formylkynurenine | M−H− | 235.0724 | 7.23 | ND | ND | ND | ↘ | | - | 1 | |
| Serotonin | M+H+ | 177.1022 | 10.46 | ↘ | - | | | ↘ | | 1 | |
| Tyrosine metabolism | 1,2-Dehydrosalsolinol | M+H+ | 178.0863 | 9.84 | ↗ | ↗ | - | ↗ | | ↗ | 2 |
| 3,4-dihydroxymandelaldehyde | M−H− | 167.0350 | 4.49 | - | | - | - | - | - | 1 | |
| 4-Hydroxyphenylacetylglutamic acid | M+H+ | 282.0972 | 8.24 | ↘ | ↘ | | | - | - | 2 | |
| Epinephrine | M+H+ | 184.0968 | 3.67 | | - | ↗ | - | - | ND | 1 | |
| Gamma-glutamyltyrosine | M−H− | 309.1092 | 6.98 | - | - | - | | | | 1 | |
| Hydroxyphenylacetylglycine | M−H− | 208.0615 | 5.02 | | | - | ND | ND | ND | 1 | |
| L-Tyrosine † | M+H+ | 182.0812 | 3.54 | - | | - | - | | - | 1 | |
| N-Acetyl-L-tyrosine | M−H− | 222.0772 | 7.29 | - | - | - | | | - | 1 | |
| O-tyrosine | M+H+ | 182.0812 | 7.40 | | ↘ | - | - | - | - | 1 | |
| P-octopamine | M−H2O+H+ | 136.0751 | 3.54 | - | | - | - | | | 1 | |
| Phenol | M−H2O+H+ | 77.0380 | 7.81 | - | | - | - | | ↘ | 2 | |
| Tyramine | M+H+ | 138.0913 | 8.11 | - | | ↗ | - | - | - | 1 | |
| Valine, leucine and isoleucine metabolism and derivatives | DL-isoleucine | M+H+ | 132.1019 | 3.92 | - | - | - | | - | ↘ | 2 |
| or L-Alloisoleucine | |||||||||||
| or L-Isoleucine | |||||||||||
| or L-Leucine | |||||||||||
| Erythronilic acid | M−H2O−H− | 99.0446 | 2.71 | ↗ | ↘ | - | | - | - | 2 | |
| Hydroxyisocaproic acid | M−H− | 131.0714 | 8.74 | - | | - | | ↘ | - | 1 | |
| L-Threonine † | M−H− | 118.0510 | 1.31 | - | - | - | - | | - | 1 | |
| N-acetylisoleucine | M−H− | 172.0979 | 9.44 | - | - | - | | | - | 1 | |
| N-Acetylvaline | M−H− | 158.0823 | 4.80 | | - | - | | - | - | 1 | |
| Vitamin B6 metabolism | 4-Pyridoxic acid | M−H− | 182.0460 | 1.44 | - | | - | - | - | - | 1 |
| 5-Pyridoxolactone | M−H− | 164.0353 | 3.25 | - | - | - | | - | - | 2 | |
| L-Glutamine † | M+H+ | 147.0764 | 1.26 | - | ↘ | - | - | | | 1 | |
| Pyridoxine | M−H2O−H− | 150.0555 | 10.81 | - | - | - | | - | ↗ | 2 | |
| Other metabolites | 2-Ketohexanoic acid | M+H+ | 131.0703 | 3.56 | - | - | - | | - | - | 2 |
| DL-Lactic Acid † | M−H− | 89.0244 | 6.38 | - | - | | - | | ↘ | 1 | |
| Ethyladipic acid | M−H− | 173.0819 | 10.35 | | ↘ | - | | ↗ | - | 2 | |
| g-glutamyl-ornithine | M−H2O+H+ | 244.1286 | 3.19 | ND | ND | ND | - | - | | 2 | |
| L-Acetylcarnitine | M+H+ | 204.1230 | 5.78 | - | - | | ↘ | - | - | 1 | |
| O-Propanoyl-D-carnitine | M+H+ | 218.1387 | 10.38 | ↗ | ↗ | | ↗ | - | ↘ | 2 | |
| Salsoline-1-carboxylate | M+H+ | 238.1074 | 9.94 | | - | - | | - | - | 2 | |
| Vanillic acid 4-sulfate | M+FA−H− | 293.1143 | 10.65 | - | - | | | | | 2 | |
| Day | Metabolic Pathways | |
|---|---|---|
| Male Mussels | Female Mussels | |
| 1 | Phenylalanine † (1) Phenylalanine, tyrosine & tryptophan biosynthesis † (1) | Valine, leucine & isoleucine degradation * (2) Valine, leucine & isoleucine biosynthesis * (2) Glycerophospholipid * (3) Pyrimidine * (4) Pentose phosphate pathway * Glycerolipid † Ether lipid † (5) Pyruvate † (6) Glycolysis/gluconeogenesis † (7) |
| 3 | Steroid hormone biosynthesis * Vitamin B6 * Phenylalanine, tyrosine & tryptophan biosynthesis * (1) Phenylalanine * (1) Glycine, serine & threonine † (8) Aminoacyl-tRNA biosynthesis † (9) Tryptophan † (1) Ether lipid † (5) Biotin † (10) | Phenylalanine * (1) Phenylalanine, Tyrosine & Tryptophan biosynthesis * (1) Aminoacyl-tRNA biosynthesis * (9) Tyrosine * (1) D-arginine & D-ornithine * (11) Glycine, serine & threonine * (8) Cysteine & methionine * Biotin * (10) Primary bile acid biosynthesis † Glycerophospholipid † (3) Pyrimidine † (4) Ether lipid † (5) |
| 7 | Glycolysis/Gluconeogenesis * (7) Pyruvate * (6) beta-Alanine * (12) Alanine, aspartate & glutamate * (12) Propanoate * Nicotinate & nicotinamide * Pantothenate & CoA biosynthesis * Citrate cycle * Butanoate * Histidine † Aminoacyl-tRNA biosynthesis † (9) Arginine biosynthesis † (11) Valine, leucine & isoleucine degradation † (2) Amino sugar & nucleotide sugar † | Purine * Glutathione * Valine, leucine & isoleucine degradation * (2) Valine, leucine & isoleucine biosynthesis * (2) Nitrogen * D-glutamine & D-glutamate * (12) Aminoacyl-tRNA biosynthesis * (9) Glycolysis/gluconeogenesis † (7) Pyruvate † (6) Arginine & proline † (11) Biotin † (10) |
| Day | Male Mussels | Female Mussels | Undifferentiated | Dead | ||||
|---|---|---|---|---|---|---|---|---|
| Controls | Exposed | Controls | Exposed | Controls | Exposed | Controls | Exposed | |
| 1 | 13 | 7 (11) | 9 | 14 | 2 | / | 1 | / |
| 3 | 8 | 11 (12) | 15 | 13 | 1 | / | 1 | / |
| 7 | 11 | 15 | 12 | 6 (7) | / | 3 | 2 | / |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramirez, G.; Gomez, E.; Dumas, T.; Rosain, D.; Mathieu, O.; Fenet, H.; Courant, F. Early Biological Modulations Resulting from 1-Week Venlafaxine Exposure of Marine Mussels Mytilus galloprovincialis Determined by a Metabolomic Approach. Metabolites 2022, 12, 197. https://doi.org/10.3390/metabo12030197
Ramirez G, Gomez E, Dumas T, Rosain D, Mathieu O, Fenet H, Courant F. Early Biological Modulations Resulting from 1-Week Venlafaxine Exposure of Marine Mussels Mytilus galloprovincialis Determined by a Metabolomic Approach. Metabolites. 2022; 12(3):197. https://doi.org/10.3390/metabo12030197
Chicago/Turabian StyleRamirez, Gaëlle, Elena Gomez, Thibaut Dumas, David Rosain, Olivier Mathieu, Hélène Fenet, and Frédérique Courant. 2022. "Early Biological Modulations Resulting from 1-Week Venlafaxine Exposure of Marine Mussels Mytilus galloprovincialis Determined by a Metabolomic Approach" Metabolites 12, no. 3: 197. https://doi.org/10.3390/metabo12030197
APA StyleRamirez, G., Gomez, E., Dumas, T., Rosain, D., Mathieu, O., Fenet, H., & Courant, F. (2022). Early Biological Modulations Resulting from 1-Week Venlafaxine Exposure of Marine Mussels Mytilus galloprovincialis Determined by a Metabolomic Approach. Metabolites, 12(3), 197. https://doi.org/10.3390/metabo12030197