Changes in Dendritic Spine Morphology and Density of Granule Cells in the Olfactory Bulb of Anguilla anguilla (L., 1758): A Possible Way to Understand Orientation and Migratory Behavior

 , ,
, ,  ,
,
Abstract
Simple Summary
Abstract
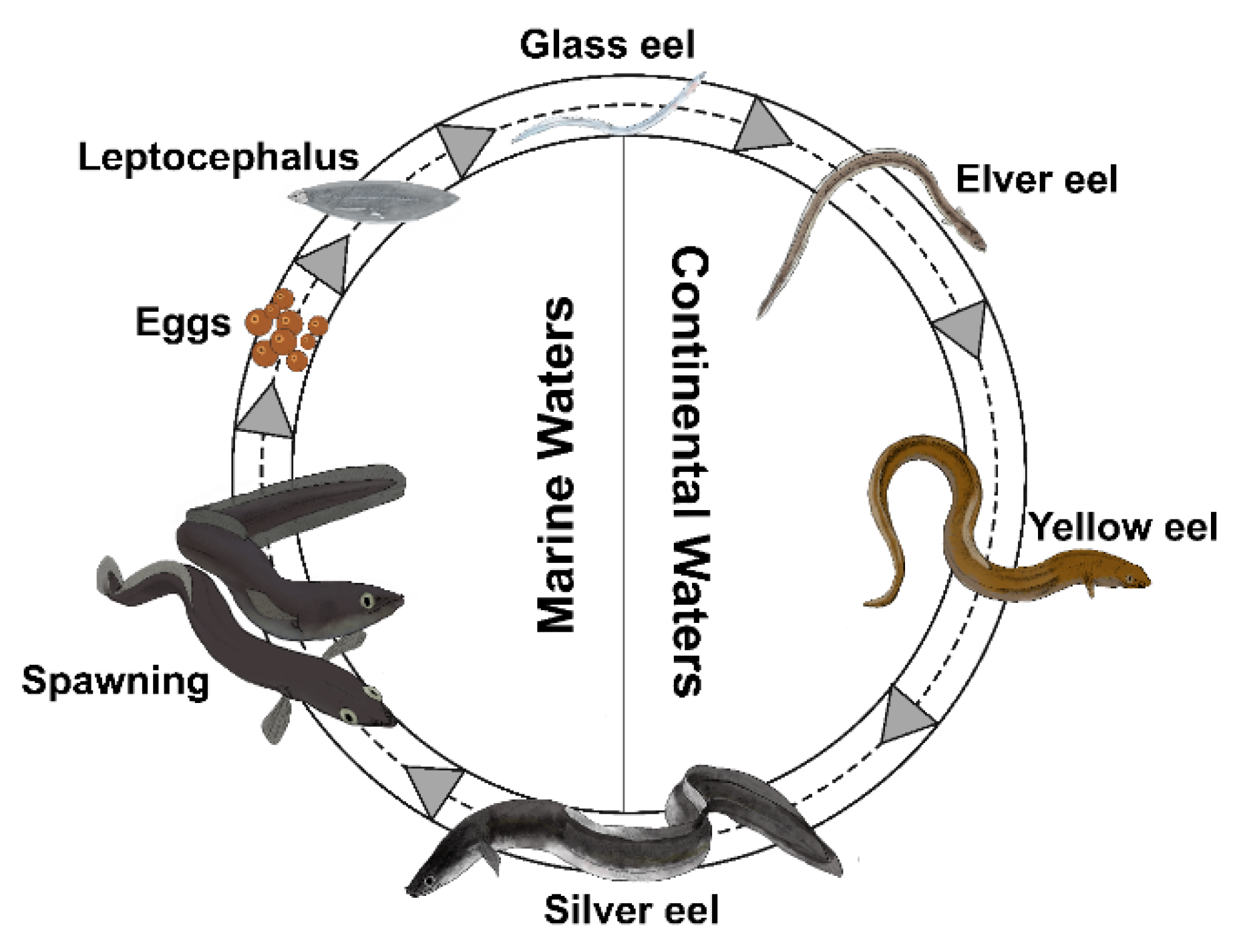
1. Introduction
2. Materials and Methods
2.1. Eel Samples
2.2. Golgi-Cox Processing
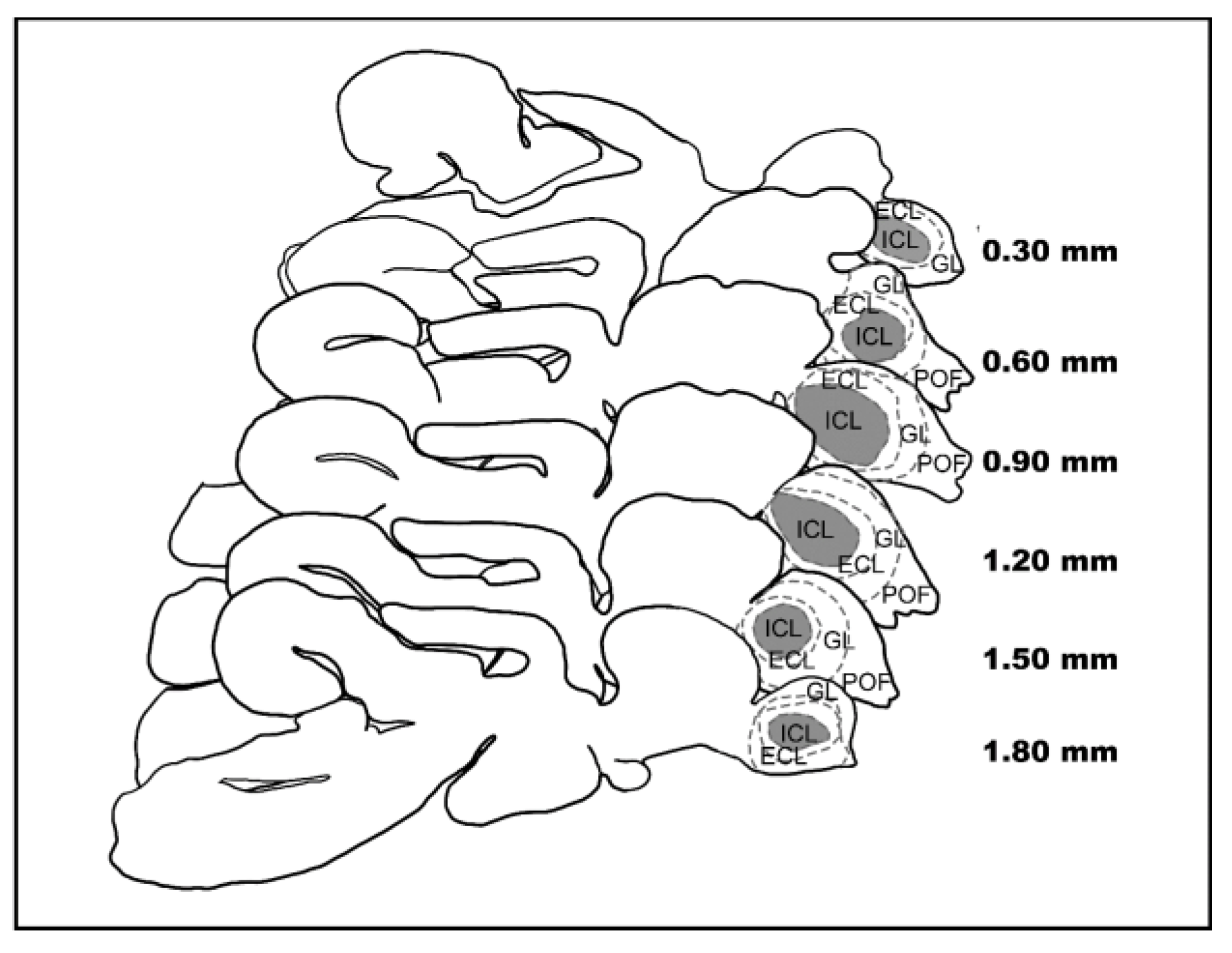
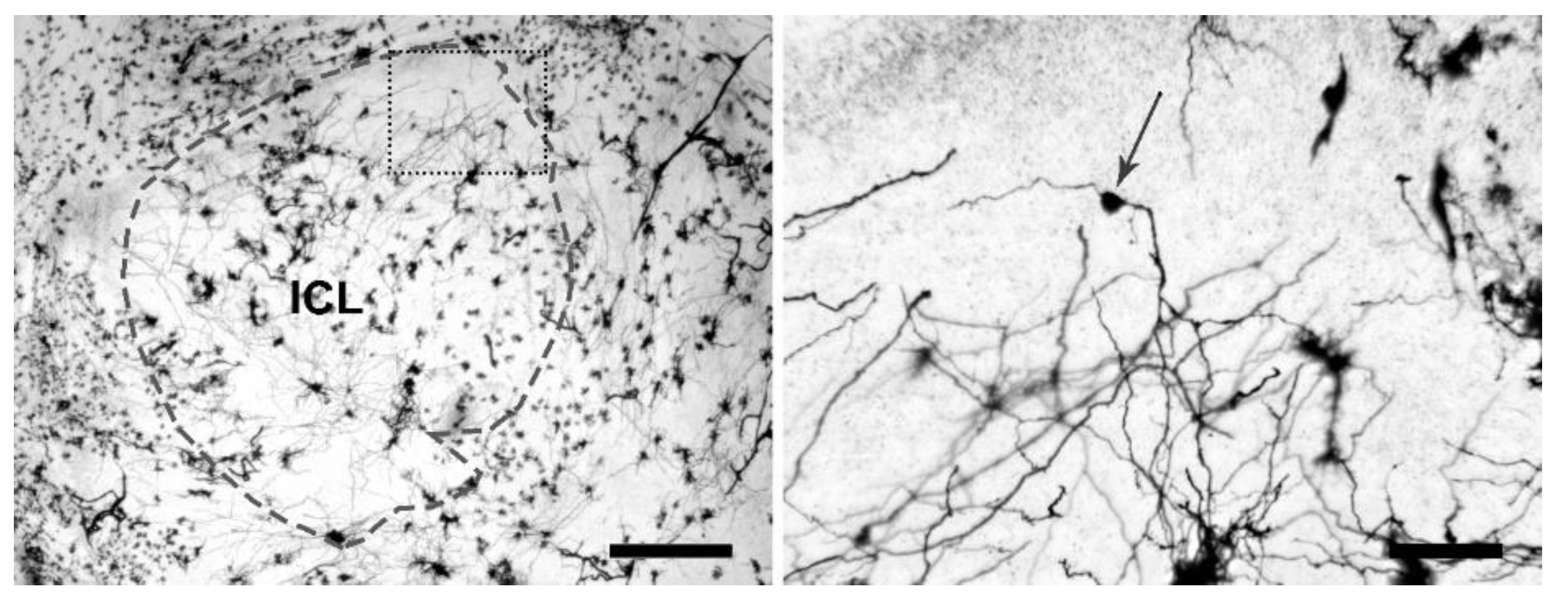
2.3. Laser Scanning Confocal Microscopy
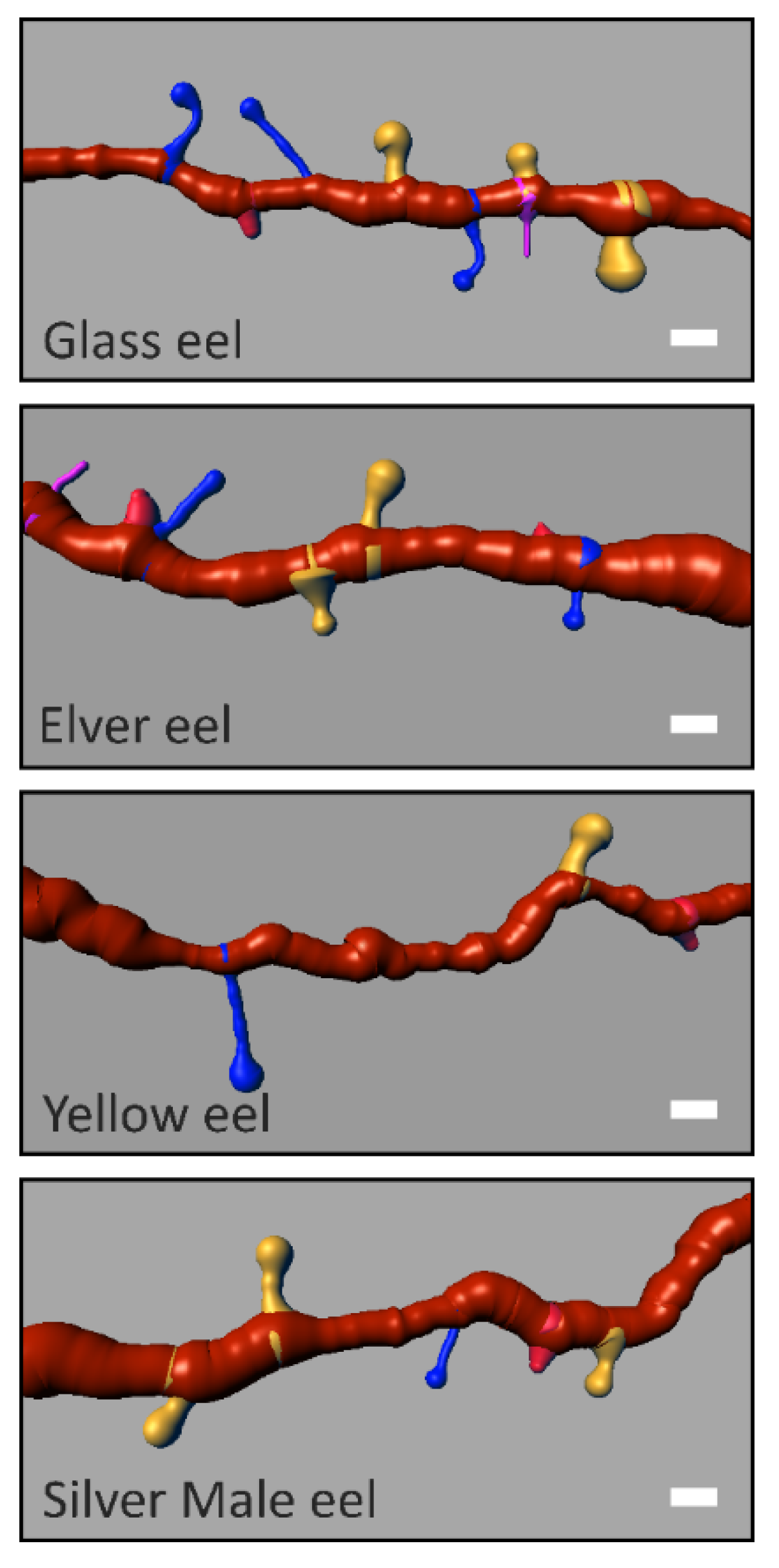
2.4. Rendering
2.5. Statistical Analysis
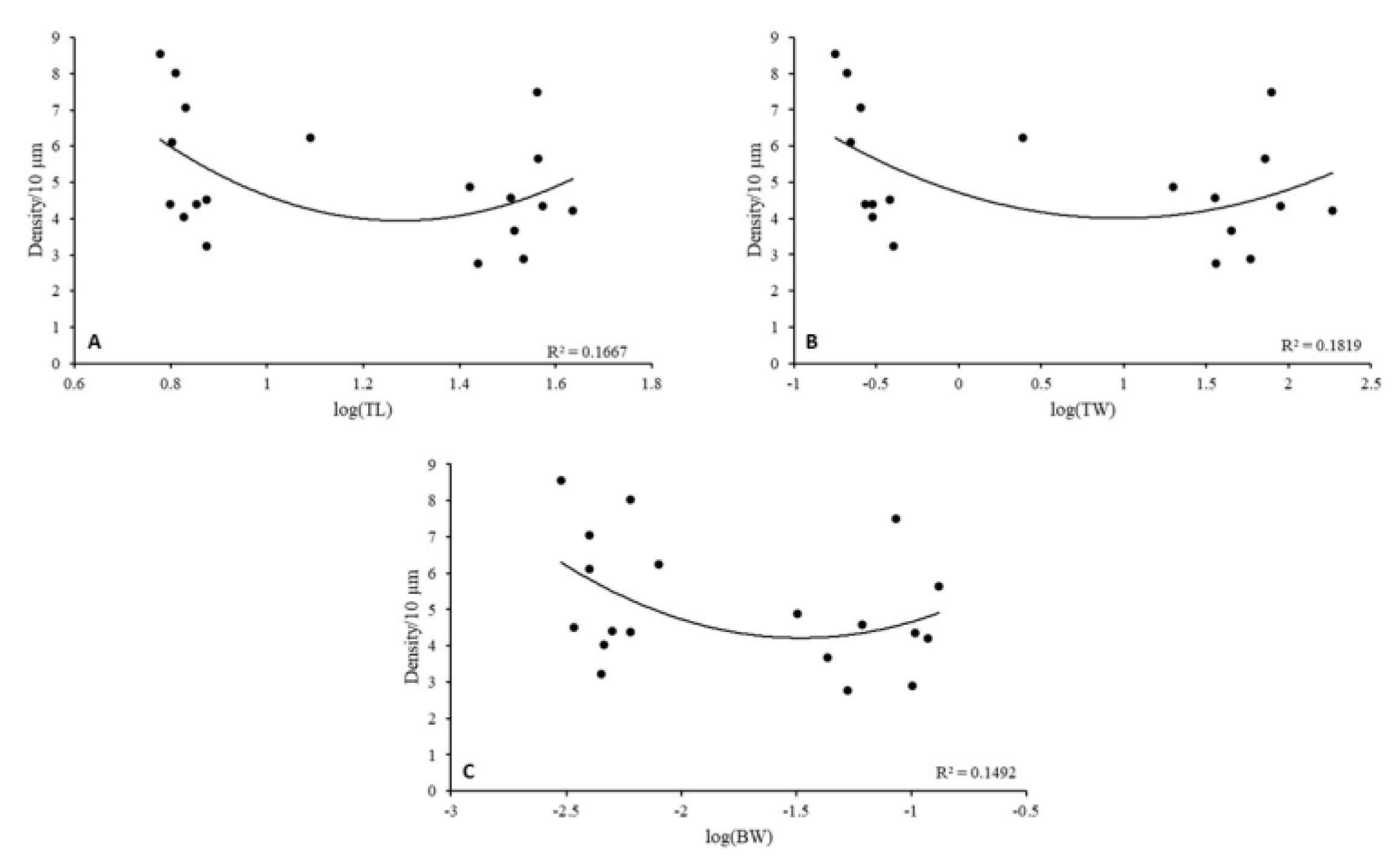
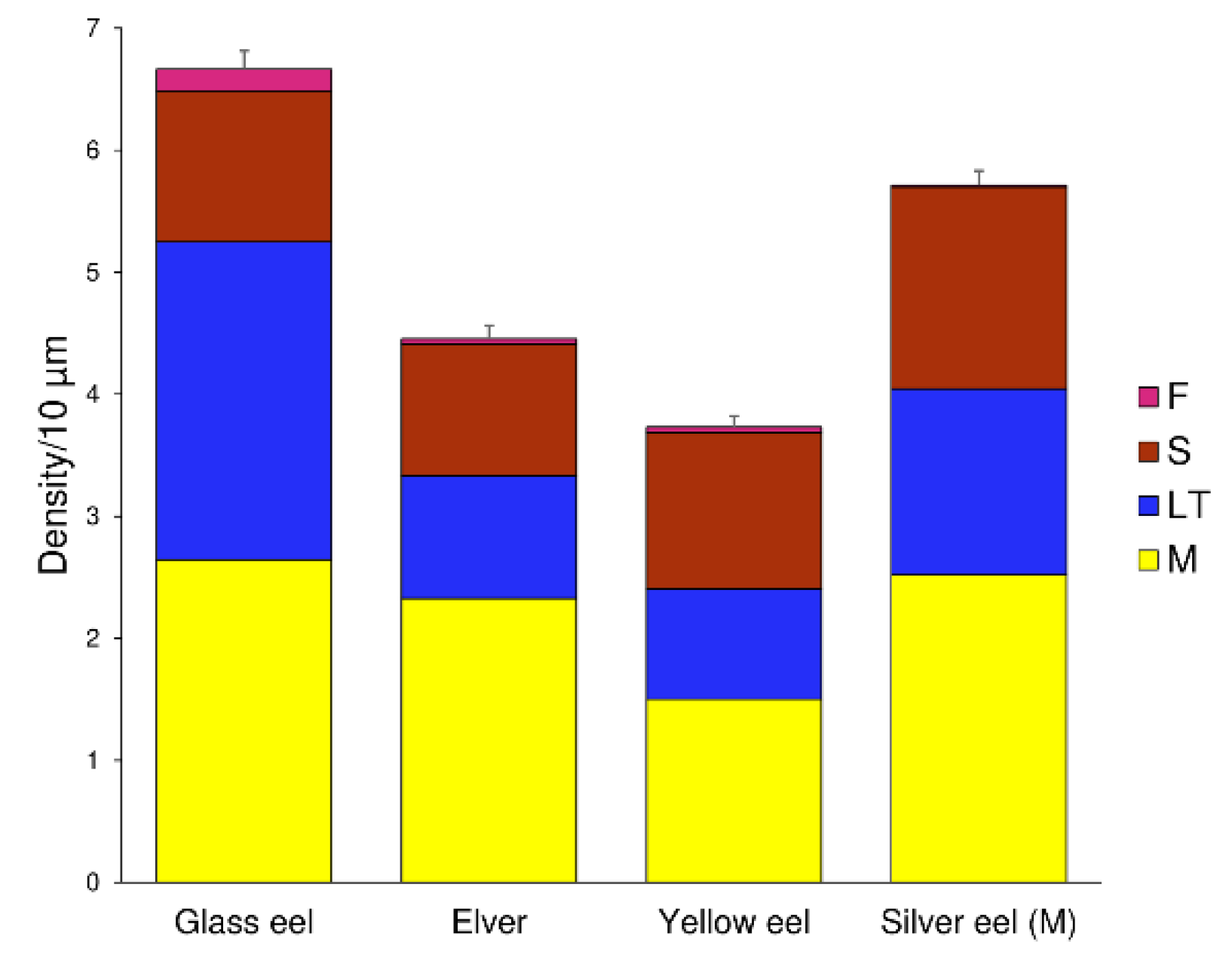
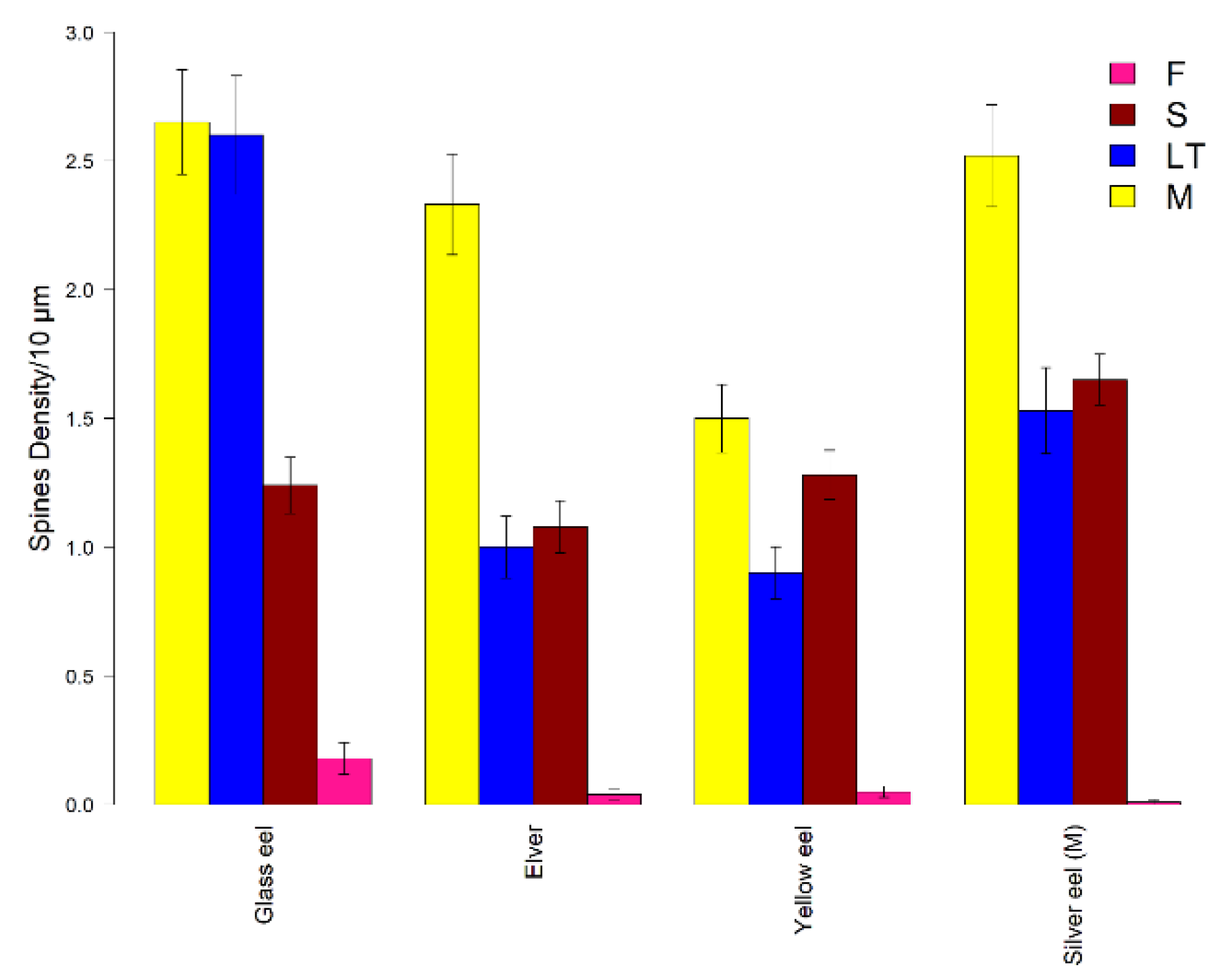
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kleerekoper, H. Some Aspects of Olfaction in Fishes, with Special Reference to Orientation. Am. Zool. 1967, 7, 385–395. [Google Scholar] [CrossRef]
- Hara, T.J. Olfaction in fish. Prog. Neurobiol. 1975, 5, 271–335. [Google Scholar] [CrossRef]
- Calvo-Ochoa, E.; Byrd-Jacobs, C.A. The Olfactory System of Zebrafish as a Model for the Study of Neurotoxicity and Injury: Implications for Neuroplasticity and Disease. Int. J. Mol. Sci. 2019, 20, 1639. [Google Scholar] [CrossRef]
- Gunning, G.E. The sensory basis for homing in the longear sunfish Lepomis megalotis megalotis (Rafinesque). Invest. Indiana Lakes Streams 1959, 5, 103–130. [Google Scholar]
- Creutzberg, F. On the orientation of migrating elvers (Anguilla vulgaris Tert.) in a tidal area. Neth. J. Sea Res. 1961, 1, 257–338. [Google Scholar] [CrossRef]
- Miles, S.G. Rheotaxis of elvers of the American eel (Anguilla rostrata) in the laboratory to water from different streams in Nova Scotia. J. Fish. Res. Bd. Can. 1968, 25, 1591–1601. [Google Scholar] [CrossRef]
- Miles, S.G. Laboratory experiments on the orientation of the adult American eel, Anguilla rostrata. J. Fish. Res. Bd. Can. 1968, 25, 2143–2155. [Google Scholar] [CrossRef]
- Thunberg, B.E. Olfaction in parent stream selection by the alewife (Alosa pseudoharengus). Anim. Behav. 1971, 19, 211–225. [Google Scholar] [CrossRef]
- Atema, J.; Jacobson, S.; Todd, J.; Baylan, D. The importance of chemical signals in stimulating behaviour of marine organisms: Effects of altered environmental chemistry on animal communication. In Bioassay Techniques and Environmental Chemistry; Ann Arbor Science: Ann Arbor, NY, USA, 1973; pp. 177–197. [Google Scholar]
- Ramenofsky, M.; Wingfield, J.C. Regulation of migration. Bioscience 2007, 57, 135–143. [Google Scholar] [CrossRef]
- Oka, Y. The origin of the centrifugal fibers to the olfactory bulb in the goldfish, Carassius auratus: An experimental study using the fluorescent dye primuline as a retrograde tracer. Brain Res. 1980, 185, 215–1225. [Google Scholar] [CrossRef]
- Fujita, I.; Satou, M.; Ueda, K. A field-potential study of centripetal and centrifugal connections of the olfactory bulb in the carp, Cyprinus carpio (L.). Brain Res. 1984, 321, 33–44. [Google Scholar] [CrossRef]
- Murakami, T.; Morita, Y.; Ito, H. Extrinsic and intrinsic fiber connections of the telencephalon in a teleost, Sebastiscus marmoratus. J. Comp. Neurol. 1983, 216, 115–131. [Google Scholar] [CrossRef] [PubMed]
- Rink, E.; Wullimann, M.F. Connections of the ventral telencephalon (subpallium) in the zebrafish (Danio rerio). Brain Res. 2004, 1011, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Anzai, S.; Satou, M. Long-term and short-term plasticity in the dendro-dendritic mitral-to-granule cell synapse of the teleost olfactory bulb. Neurosci. Res. 1996, 25 (Suppl. S1), S223. [Google Scholar] [CrossRef]
- Satou, M.; Anzai, S.; Huruno, M. Long-term potentiation and olfactory memory formation in the carp (Cyprinus carpio L.) olfactory bulb. J. Comp. Physiol. A 2005, 191, 421–434. [Google Scholar] [CrossRef]
- Satou, M.; Hoshikawa, R.; Sato, Y.; Okawa, K. An in vitro study of long-term potentiation in the carp (Cyprinus carpio L.) olfactory bulb. J. Comp. Physiol. A 2006, 192, 135–150. [Google Scholar] [CrossRef]
- Rehn, B.; Breipohl, W.; Mendoza, A.S.; Apfelbach, R. Changes in granule cells of the ferret olfactory bulb associated with imprinting on prey odours. Brain Res. 1986, 373, 114–125. [Google Scholar] [CrossRef]
- Matsutani, S.; Yamamoto, N. Postnatal development of dendritic spines on olfactory bulb granule cells in rats. J. Comp. Neurol. 2004, 473, 553–561. [Google Scholar] [CrossRef]
- Matsutani, S.; Yamamoto, N. Brain-derived neurotrophic factor induces rapid morphological changes in dendritic spines of olfactory bulb granule cells in cultured slices through the modulation of glutamatergic signaling. Neuroscience 2004, 123, 695–702. [Google Scholar] [CrossRef]
- Mizrahi, A. Dendritic development and plasticity of adult-born neurons in the mouse olfactory bulb. Nat. Neurosci. 2007, 10, 444–452. [Google Scholar] [CrossRef]
- Huang, Y.B.; Hu, C.R.; Zhang, L.; Yin, W.; Hu, B. In vivo study of dynamics and stability of dendritic spines on olfactory bulb interneurons in Xenopus laevis tadpoles. PLoS ONE 2015, 10, e0140752. [Google Scholar] [CrossRef] [PubMed]
- McDole, B.; Isgor, C.; Pare, C.; Guthrie, K. BDNF over-expression increases olfactory bulb granule cell dendritic spine density in vivo. Neuroscience 2015, 304, 146–160. [Google Scholar] [CrossRef][Green Version]
- Mandairon, N.; Kuczewski, N.; Kermen, F.; Forest, J.; Midroit, M.; Richard, M.; Thevenet, M.; Sacquet, J.; Linster, C.; Didier, A. Opposite regulation of inhibition by adult-born granule cells during implicit versus explicit olfactory learning. eLife 2018, 7, e34976. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, Y.; Hu, B. Olfactory experiences dynamically regulate plasticity of dendritic spines in granule cells of Xenopus tadpoles in vivo. Sci. Rep. 2016, 6, 35009. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, V.A.; Sabatini, B.L. Anatomical and physiological plasticity of dendritic spines. Annu. Rev. Neurosci. 2007, 30, 79–97. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, M.; Ellis-Davies, G.C.; Nemoto, T.; Miyashita, Y.; Iino, M.; Kasai, H. Dendritic spine geometry is critical for AMPA receptor expression in hippocampal CA1 pyramidal neurons. Nat. Neurosci. 2001, 4, 1086–1092. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, M.; Honkura, N.; Ellis-Davies, G.C.; Kasai, H. Structural basis of long-term potentiation in single dendritic spines. Nature 2004, 429, 761–766. [Google Scholar] [CrossRef]
- Ghani, M.U.; Mesadi, F.; Kanık, S.D.; Argunşah, A.Ö.; Hobbiss, A.F.; Israely, I.; Ünay, D.; Taşdizen, T. Çetin, M. Dendritic spine classification using shape and appearance features based on two-photon microscopy. J. Neurosci. Methods 2017, 279, 13–21. [Google Scholar] [CrossRef]
- Spiga, S.; Talani, G.; Mulas, G.; Licheri, V.; Fois, G.R.; Muggironi, G.; Masala, N.; Cannizzaro, C.; Biggio, G.; Sanna, E.; et al. Hampered long-term depression and thin spine loss in the nucleus accumbens of ethanol-dependent rats. Biol. Sci. 2014, 111, E3745–E3754. [Google Scholar] [CrossRef]
- Pchitskaya, E.; Bezprozvanny, I. Dendritic spines shape analysis—Classification or clusterization? Perspective. Front. Synaptic Neurosci. 2020, 12, 31. [Google Scholar] [CrossRef]
- Hayashi, Y.; Majewska, A.K. Dendritic spine geometry: Functional implication and regulation. Neuron 2005, 46, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Bourne, J.; Harris, K.M. Do thin spines learn to be mushroom spines that remember? Curr. Opin. Neurobiol. 2007, 17, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Hering, H.; Sheng, M. Dentritic spines: Structure, dynamics and regulation. Nat. Rev. Neurosci. 2001, 2, 880–888. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, Y.; De Roo, M.; Muller, D. Dendritic spine formation and stabilization. Curr. Opin. Neurobiol. 2009, 19, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Berry, K.P.; Nedivi, E. Spine dynamics: Are they all the same? Neuron 2017, 96, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Kasai, H.; Fukuda, M.; Watanabe, S.; Hayashi-Takagi, A.; Noguchi, J. Structural dynamics of dendritic spines in memory and cognition. Trends Neurosci. 2010, 33, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Lendvai, B.; Stern, E.A.; Chen, B.; Svoboda, K. Experience-dependent plasticity of dendritic spines in the developing rat barrel cortex in vivo. Nature 2000, 404, 876–881. [Google Scholar] [CrossRef]
- Muller, D.; Nikonenko, I. Dendritic spines. Neural Circuit Development and Function in the Healthy and Diseased Brain. In Comprehensive Developmental Neuroscience; Rubenstein, J., Rakic, P., Eds.; Academic Press: Cambridge, MA, USA, 2013; Volume 3, p. 95. [Google Scholar]
- Fu, M.; Zuo, Y. Experience-dependent structural plasticity in the cortex. Trends Neurosci. 2011, 34, 177–187. [Google Scholar] [CrossRef]
- Yang, G.; Pan, F.; Gan, W.B. Stably maintained dendritic spines are associated with lifelong memories. Nature 2009, 462, 920–924. [Google Scholar] [CrossRef]
- Zhou, Q.; Homma, K.J.; Poo, M.M. Shrinkage of dendritic spines associated with long-term depression of hippocampal synapses. Neuron 2004, 44, 749–757. [Google Scholar] [CrossRef]
- Bailey, C.H.; Kandel, E.R.; Harris, K.M. Structural components of synaptic plasticity and memory consolidation. Cold Spring Harb. Perspect. Biol. 2015, 7, a021758. [Google Scholar] [CrossRef] [PubMed]
- Tesch, F.W. The Eel; Willey-Blackwell Publishing: Oxford, UK, 2003; p. 416. [Google Scholar] [CrossRef]
- Schmidt, J. Breeding Places and Migrations of the Eel. Nature 1923, 111, 51–54. [Google Scholar] [CrossRef]
- Miller, M.J.; Westerberg, H.; Sparholt, H.; Wysujack, K.; Sørensen, S.R.; Marohn, L.; Jacobsen, M.W.; Freese, M.; Ayala, D.J.; Pohlmann, J.D.; et al. Spawning by the European eel across 2000 km of the Sargasso Sea. Biol. Lett. 2019, 15, 20180835. [Google Scholar] [CrossRef] [PubMed]
- Tesch, F.W. The Eel Biology and Management of Anguillid Eels; John Wiley & Sons: New York, NY, USA, 1977; p. 434. [Google Scholar]
- Bonhommeau, S.; Castonguay, M.; Rivot, E.; Sabatié, R.; Le Pape, O. The duration of migration of Atlantic Anguilla larvae. Fish Fish. 2010, 11, 289–306. [Google Scholar] [CrossRef]
- Hanel, R.; Stepputtis, D.; Bonhommeau, S.; Castonguay, M.; Schaber, M.; Wysujack, K.; Vobach, M.; Miller, M.J. Low larval abundance in the Sargasso Sea: New evidence about reduced recruitment of the Atlantic eels. Naturwissenschaften 2014, 101, 1041–1054. [Google Scholar] [CrossRef]
- Miller, M.J.; Bonhommeau, S.; Munk, P.; Castonguay, M.; Hanel, R.; McCleave, J.D. A century of research on the larval distributions of the Atlantic eels: A re-examination of the data. Biol. Rev. 2015, 90, 1035–1064. [Google Scholar] [CrossRef]
- Tesch, F.W. Occurrence of Eel Anguilla anguilla Larvae West of the European Continental Shelf, 1971–1977. Environ. Biol. Fishes 1980, 5, 185–190. [Google Scholar] [CrossRef]
- Deelder, C.L. On the migration of the elver (Anguilla vulgaris Turt.) at sea. J. Cons. Int. Explor. Mer. 1952, 20, 187–218. [Google Scholar] [CrossRef]
- Tosi, L.; Sala, L.; Sola, C.; Spampanato, A.; Tongiorgi, P. Experimental analysis of the thermal and salinity preferences of glass-eels, Anguilla anguilla (L.), before and during the upstream migration. Fish Biol. 1988, 33, 721–733. [Google Scholar] [CrossRef]
- Crnjar, R.; Slcalera, G.; Bigiani, A.; Tomassini Barbarossa, I.; Magherini, P.C.; Pietra, P. Olfactory sensitivity to amino acids in the juvenile stages of the European eel Anguilla anguilla (L.). J. Fish Biol. 1992, 40, 567–576. [Google Scholar] [CrossRef]
- Sola, C.; Tosi, L. Bile salts and taurine as chemical stimuli for glass eels, Anguilla anguilla: A behavioural study. Environ. Biol. Fishes 1993, 37, 197–204. [Google Scholar] [CrossRef]
- Sola, C. Chemoattraction of upstream migrating glass eels Anguilla anguilla to earthy and green odorants. Environ. Biol. Fishes 1995, 43, 179–185. [Google Scholar] [CrossRef]
- Schmucker, A.K.; Johnson, N.S.; Galbraith, H.S.; Li, W. Glass-eel-stage American eels respond to conspecific odor as a function of concentration. Trans. Am. Fish. Soc. 2016, 145, 712–722. [Google Scholar] [CrossRef]
- Cresci, A.; Paris, C.B.; Durif, C.M.; Shema, S.; Bjelland, R.M.; Skiftesvik, A.B.; Browman, H.I. Glass eels (Anguilla anguilla) have a magnetic compass linked to the tidal cycle. Sci. Adv. 2017, 3, e1602007. [Google Scholar] [CrossRef] [PubMed]
- Cresci, A.; Paris, C.B.; Foretich, M.A.; Durif, C.M.; Shema, S.D.; O’Brien, C.E.; Vikebø, F.B.; Skiftesvik, A.B.; Browman, H.I. Atlantic haddock (Melanogrammus aeglefinus) larvae have a magnetic compass that guides their orientation. iScience 2019, 19, 1173–1178. [Google Scholar] [CrossRef]
- Cresci, A.; Durif, C.M.; Paris, C.B.; Shema, S.D.; Skiftesvik, A.B.; Browman, H.I. Glass eels (Anguilla anguilla) imprint the magnetic direction of tidal currents from their juvenile estuaries. Commun. Biol. 2019, 2, 366. [Google Scholar] [CrossRef]
- Tzeng, W.N.; Wang, C.H.; Wickström, M.H.; Reizenstein, M. Occurrence of the semi-catadromous European eel Anguilla anguilla in the Baltic Sea. Mar. Biol. 2000, 137, 93–98. [Google Scholar] [CrossRef]
- Wood, P.; Partridge, J.C.; Grip, W.J. Rod visual pigment changes in the elver of the eel Anguilla anguilla L. measured by microspectrophotometry. J. Fish Biol. 1992, 41, 601–611. [Google Scholar] [CrossRef]
- Ciccotti, B.E.; Macchi, E.; Rossi, A.; Cataldi, E.; Cataudella, S. Glass eel (Anguilla anguilla) acclimation to freshwater and seawater: Morphological changes of the digestive tract. J. Appl. Ichthyol. 1993, 9, 74–81. [Google Scholar] [CrossRef]
- Durif, C.M.F.; Van Ginneken, V.; Dufour, S.; Müller, T.; Elie, P. Seasonal evolution and individual differences in silvering eels from different locations. In Spawning Migration of the European Eel; Springer: Dordrecht, The Netherlands, 2009; pp. 13–38. [Google Scholar]
- Amilhat, E.; Aarestrup, K.; Faliex, E.; Simon, G.; Westerberg, H.; Righton, D. First evidence of European eels exiting the Mediterranean Sea during their spawning migration. Sci. Rep. 2016, 6, 21817. [Google Scholar] [CrossRef]
- Righton, D.; Westerberg, H.; Feunteun, E.; Okland, F.; Gargan, P.; Amilhat, E.; Metcalfe, J.; Lobon-Cervia, J.; Sjo Berg, N.; Simon, J.; et al. Empirical observations of the spawning migration of European eels: The long and dangerous road to the Sargasso Sea. Sci. Adv. 2016, 2, e1501694. [Google Scholar] [CrossRef] [PubMed]
- Béguer-Pon, M.; Dodson, J.J.; Castonguay, M.; Jellyman, D.; Aarestrup, K.; Tsukamoto, K. Tracking anguillid eels: Five decades of telemetry-based research. Mar. Freshw. Res. 2018, 69, 199. [Google Scholar] [CrossRef]
- Huertas, M.; Canário, A.V.M.; Hubbard, P.C. Chemical Communication in the Genus Anguilla: A Minireview. Behaviour 2008, 145, 1389–1407. [Google Scholar] [CrossRef]
- Westin, L. Orientation mechanisms in migrating European silver eel (Anguilla anguilla): Temperature and olfaction. Mar. Biol. 1990, 106, 175–179. [Google Scholar] [CrossRef]
- Westin, L. The spawning migration of European silver eel (Anguilla anguilla L.) with particular reference to stocked eel in the Baltic. Fish. Res. 1998, 38, 257–270. [Google Scholar] [CrossRef]
- Westin, L. Migration failure in stocked eels Anguilla anguilla. Mar. Ecol. Prog. Ser. 2003, 254, 307–311. [Google Scholar] [CrossRef]
- De Waele, J.; Martina, M.L.V.; Sanna, L.; Cabras, S.; Cossu, Q.A. Flash flood hydrology in karstic terrain: Flumineddu Canyon, 446 central-east Sardinia. Geomorphology 2010, 120, 162–173. [Google Scholar] [CrossRef]
- Sabatini, A.; Podda, C.; Frau, G.; Cani, M.V.; Musu, A.; Serra, M.; Palmas, F. Restoration of native Mediterranean brown trout Salmo cettii Rafinesque, 1810 (Actinopterygii: Salmonidae) populations using an electric barrier as a mitigation tool. Eur. Zool. J. 2018, 85, 138–150. [Google Scholar] [CrossRef]
- Palmas, F.; Righi, T.; Musu, A.; Frongia, C.; Podda, C.; Serra, M.; Splendiani, A.; Caputo Barucchi, V.; Sabatini, A. Pug-headed-ness anomaly in a wild and isolated population of native mediterranean trout Salmo trutta L.; 1758 complex (Osteichthyes: Salmonidae). Dyversity 2020, 12, 353. [Google Scholar] [CrossRef]
- Podda, C.; Palmas, F.; Pusceddu, A.; Sabatini, A. When the eel meets dams: Larger dams’ long-term impacts on Anguilla anguilla (L., 1758). Front. Environ. Sci. 2022, 10, 876369. [Google Scholar] [CrossRef]
- Regione Autonoma della Sardegna. Carta Ittica Della Sardegna—D.G.R. N. 2/28, 428. del 20/01/2022; Università degli Studi di Cagliari (DISVA): Sardinia, Italy, 2022; Available online: https://delibere.regione.sardegna.it/protected/58916/0/def/ref/DBR58914/ (accessed on 21 March 2022).
- Podda, C.; Palmas, F.; Frau, G.; Chessa, G.; Culurgioni, J.; Diciotti, R.; Fois, N.; Sabatini, A. Environmental influences on the ecruitment dynamics of juvenile European eels, Anguilla anguilla, in a small estuary of the Tyrrhenian Sea, Sardinia, Italy. Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 30, 1638–1648. [Google Scholar] [CrossRef]
- Gilderhus, P.A.; Marking, L.L. Comparative efficacy of 16 anesthetic chemicals on rainbow trout. N. Am. J. Fish. Manag. 1987, 7, 288–292. [Google Scholar] [CrossRef]
- Cox, W.H. Imprägnation des centralen Nervensystemsmit Quecksilbersalzen. Arch. Mikrosk. Anat. 1891, 37, 16–21. [Google Scholar] [CrossRef]
- Glaser, E.M.; Van der Loos, H. Analysis of thick brain sections by obverse—Reverse computer microscopy: Application of a new, high clarity Golgi—Nissl stain. J. Neurosci. Methods 1981, 4, 117–125. [Google Scholar] [CrossRef]
- Zaqout, S.; Kaindl, A.M. Golgi-Cox staining step by step. Front. Neuroanat. 2016, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Wullimann, M.; Rupp, B.; Reichert, H. Neuroanatomy of the Zebrafish Brain. A Topological Atlas, 1st ed.; Birkhäuser: Basel, Switzerland, 1996; p. 144. [Google Scholar] [CrossRef]
- Kolb, B.; McClimans, J. Cryostat sectioning of Golgi-Cox tissue. Stain Technol. 1986, 61, 379–380. [Google Scholar] [CrossRef]
- Medina, M.; Reperant, J.; Dufour, S.; Ward, R.; Le Belle, N.; Miceli, D. The distribution of GABA-immunoreactive neurons in the brain of the silver eel (Anguilla anguilla L.). Anat. Embryol. 1994, 189, 25–39. [Google Scholar] [CrossRef]
- Alonso, J.R.; Lara, J.; Miguel, J.J.; ON, J. A Golgi study of the granule cells in the olfactory bulb of Cyprinus carpio L. and Barbus meridionalis Risso. Z. Mikrosk.-Anat. Forsch. 1986, 100, 224–232. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.r-project.org (accessed on 1 April 2022).
- Døving, K.B. Functional Properties of the Fish Olfactory System. In Progress in Sensory Physiology 6. Progress in Sensory Physiology; Autrum, H., Ottoson, D., Perl, E.R., Schmidt, R.F., Shimazu, H., Willis, W.D., Eds.; Springer: Berlin/Heidelberg, Germany, 1986; Volume 6. [Google Scholar] [CrossRef]
- Okabe, S.; Kim, H.D.; Miwa, A.; Kuriu, T.; Okado, H. Continual remodeling of postsynaptic density and its regulation by synaptic activity. Nat. Neurosci. 1999, 2, 804–811. [Google Scholar] [CrossRef]
- Dickstein, D.L.; Weaver, C.M.; Luebke, J.I.; Hof, P.R. Dendritic spine changes associated with normal aging. Neuroscience 2013, 251, 21–32. [Google Scholar] [CrossRef]
- Peters, A.; Kaiserman-Abramof, I.R. The small pyramidal neuron of the rat cerebral cortex. The perikaryon, dendrites and spines. Am. J. Anat. 1970, 127, 321–355. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.M.; Jensen, F.E.; Tsao, B. Three-dimensional structure of dendritic spines and synapses in rat hippocampus (CA1) at postnatal day 15 and adult ages: Implications for the maturation of synaptic physiology and long-term potentiation. J. Neurosci. 1992, 12, 2685–2705. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.G.; Medvedev, N.I.; Popov, V.I.; Schoepfer, R.; Davies, H.A.; Murphy, K.; Dallérac, G.M.; Kraev, I.V.; Rodrìguez, J.J. Chemically induced long-term potentiation increases the number of perforated and complex postsynaptic densities but does not alter dendritic spine volume in CA1 of adult mouse hippocampal slices. Eur. J. Neurosci. 2005, 21, 3368–3378. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.M. Structure, development, and plasticity of dendritic spines. Curr. Opin. Neurobiol. 1999, 9, 343–348. [Google Scholar] [CrossRef]
- Fiala, J.C.; Allwardt, B.; Harris, K.M. Dendritic spines do not split during hippocampal LTP or maturation. Nat. Neurosci. 2002, 5, 297–298. [Google Scholar] [CrossRef]
- Brocco, M.A.; Fernández, M.E.; Frasch, A.C. Filopodial protrusions induced by glycoprotein M6a exhibit high motility and aids synapse formation. Eur. J. Neurosci. 2010, 31, 195–202. [Google Scholar] [CrossRef]
- Fiala, J.C.; Feinberg, M.; Popov, V.; Harris, K.M. Synaptogenesis via dendritic filopodia in developing hippocampal area CA1. J. Neurosci. 1998, 18, 8900–8911. [Google Scholar] [CrossRef]
- Ziv, N.E.; Smith, S.J. Evidence for a role of dendritic filopodia in synaptogenesis and spine formation. Neuron 1996, 17, 91–102. [Google Scholar] [CrossRef]
- Sorra, K.E.; Harris, K.M. Overview on the structure, composition, function, development, and plasticity of hippocampal dendritic spines. Hippocampus 2000, 10, 501–511. [Google Scholar] [CrossRef]
- Churcher, A.M.; Hubbard, P.C.; Marques, J.P.; Canário, A.V.; Huertas, M. Deep sequencing of the olfactory epithelium reveals specific chemosensory receptors are expressed at sexual maturity in the European eel Anguilla anguilla. Mol. Ecol. 2015, 24, 822–834. [Google Scholar] [CrossRef]
- Coss, R.G. Delayed plasticity of an instinct: Recognition and avoidance of 2 facing eyes by the jewel fish. Dev. Psychobiol. 1979, 12, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Coss, R.G.; Globus, A. Spine stems on tectal interneurons in jewel fish are shortened by social stimulation. Science 1978, 200, 787–789. [Google Scholar] [CrossRef] [PubMed]
- Coss, R.G.; Globus, A. Social experience affects the development of dendritic spines and branches on tectal interneurons in the jewel fish. Dev. Psychobiol. 1979, 12, 347–358. [Google Scholar] [CrossRef]
- Coss, R.G.; Burgess, J.W. Jewel fish retain juvenile schooling pattern after crowded development. Dev. Psychobiol. 1981, 14, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Berard, D.R.; Burgess, J.W.; Coss, R.G. Plasticity of dendritic spine formation: A state-dependent stochastic process. Int. J. Neurosci. 1981, 13, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Burgess, J.W.; Coss, R.G. Crowded jewel fish show changes in dendritic spine density and spine morphology. Neurosci. Lett. 1980, 17, 277–281. [Google Scholar] [CrossRef]
- Plata, A.L.D.; Robles, E. NMDA Receptor Antagonist MK801 Reduces Dendritic Spine Density and Stability in Zebrafish Pyramidal Neurons. Neuroscience 2022, 498, 50–63. [Google Scholar] [CrossRef]
- Song, C.; Liu, B.P.; Zhang, Y.P.; Peng, Z.; Wang, J.; Collier, A.D.; Echevarria, D.J.; Savelieva, K.V.; Lawrence, R.F.; Rex, C.S.; et al. Modeling consequences of prolonged strong unpredictable stress in zebrafish: Complex effects on behavior and physiology. Prog. Neuropsychopharmacol. Biol. Psychiatry. 2018, 81, 384–394. [Google Scholar] [CrossRef]
- Bevacqua, D.; Melià, P.; Gatto, M.; De Leo, G.A. A global viability assessment of the European eel. Glob. Change Biol. 2015, 21, 3323–3335. [Google Scholar] [CrossRef]
- Podda, C.; Palmas, F.; Pusceddu, A.; Sabatini, A. Hard times for catadromous fish: The case of the European eel Anguilla anguilla (L. 1758). Adv. Oceanogr. Limnol. 2021, 12, 9997. [Google Scholar] [CrossRef]
- Durif, C.M.F.; Gjosaeter, J.; Vollestad, L.A. Influence of oceanic factors on Anguilla anguilla (L.) over the twentieth century in coastal habitats of the Skagerrak, Southern Norway. Proc. R. Soc. B 2011, 278, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Pike, C.; Crook, V.; Gollock, M. Anguilla anguilla. The IUCN Red List of Threatened Species. 2020. Available online: https://doi.org/10.2305/IUCN.UK.2020-2.RLTS.T60344A152845178.en (accessed on 1 April 2022).
- Lauder, G.V.; Liem, K.F. Patterns of diversity and evolution in ray-finned fishes. In Fish Neurobiology. Brain Stem and Sense Organs; Northcutt, R.G., Davis, R.E., Eds.; University of Michigan Press: Ann Arbor, MI, USA, 1983; Volume I, pp. 2–24. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Life Stage | TL (cm) | TW (g) | BW (g) |
|---|---|---|---|
| Glass eel | 6.53 ± 0.43 | 0.23 ± 0.04 | 0.004 ± 0.001 |
| Elver | 8.06 ± 2.43 | 0.77 ± 0.94 | 0.005 ± 0.002 |
| Yellow eel | 30.56 ± 3.41 | 39.04 ± 14.04 | 0.058 ± 0.026 |
| Silver eel | 38.40 ± 3.23 | 105.75 ± 52.22 | 0.11 ± 0.049 |
| Glass eel | Elver | Yellow eel | Silver eel (M) | |||||
|---|---|---|---|---|---|---|---|---|
| K-W = 130.19 p < 0.001 | K-W= 134.86 p < 0.001 | K-W= 122.46 p < 0.001 | K-W= 154.93 p < 0.001 | |||||
| Paired groups | p value | significance | p value | significance | p value | significance | p value | significance |
| M-S | <0.0001 | **** | <0.0001 | **** | 0.4912 | Ns | 0.0014 | ** |
| M-LT | 0.6921 | ns | <0.0001 | **** | <0.0001 | **** | <0.0001 | **** |
| M-F | <0.0001 | **** | <0.0001 | **** | <0.0001 | **** | <0.0001 | **** |
| LT-S | <0.0001 | **** | 0.3539 | ns | 0.0028 | **** | 0.051 | ns |
| LT-F | <0.0001 | **** | <0.0001 | **** | <0.0001 | **** | <0.0001 | **** |
| S-F | <0.0001 | **** | <0.0001 | **** | <0.0001 | **** | <0.0001 | **** |
| Mushroom (M) | Stubby (S) | Long Thin (LT) | Filopodia (F) | |||||
|---|---|---|---|---|---|---|---|---|
| K-W = 23.52 p < 0.001 | K-W = 130.19 p < 0.001 | K-W = 47.23 p < 0.001 | K-W = 7.90 p = 0.048 | |||||
| Paired groups | p value | significance | p value | significance | p value | significance | p value | significance |
| Glass eel-Elver | 0.2479 | ns | 0.3944 | ns | <0.0001 | **** | 0.0793 | ns |
| Glass eel-Yellow eel | <0.0001 | **** | 0.5585 | ns | <0.0001 | **** | 0.1318 | ns |
| Glass eel-Silver eel (M) | 0.6584 | ns | 0.0042 | ** | <0.0001 | **** | 0.0128 | * |
| Elver-Yellow eel | 0.0013 | ** | 0.0953 | ns | 0.7401 | ns | 0.7591 | ns |
| Elver-Silver eel (M) | 0.3719 | ns | <0.0001 | **** | 0.0145 | * | 0.3738 | ns |
| Yellow eel-Silver eel (M) | <0.0001 | **** | 0.0097 | ** | 0.0040 | ** | 0.2338 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porceddu, R.; Podda, C.; Mulas, G.; Palmas, F.; Picci, L.; Scano, C.; Spiga, S.; Sabatini, A. Changes in Dendritic Spine Morphology and Density of Granule Cells in the Olfactory Bulb of Anguilla anguilla (L., 1758): A Possible Way to Understand Orientation and Migratory Behavior. Biology 2022, 11, 1244. https://doi.org/10.3390/biology11081244
Porceddu R, Podda C, Mulas G, Palmas F, Picci L, Scano C, Spiga S, Sabatini A. Changes in Dendritic Spine Morphology and Density of Granule Cells in the Olfactory Bulb of Anguilla anguilla (L., 1758): A Possible Way to Understand Orientation and Migratory Behavior. Biology. 2022; 11(8):1244. https://doi.org/10.3390/biology11081244
Chicago/Turabian StylePorceddu, Riccardo, Cinzia Podda, Giovanna Mulas, Francesco Palmas, Luca Picci, Claudia Scano, Saturnino Spiga, and Andrea Sabatini. 2022. "Changes in Dendritic Spine Morphology and Density of Granule Cells in the Olfactory Bulb of Anguilla anguilla (L., 1758): A Possible Way to Understand Orientation and Migratory Behavior" Biology 11, no. 8: 1244. https://doi.org/10.3390/biology11081244
APA StylePorceddu, R., Podda, C., Mulas, G., Palmas, F., Picci, L., Scano, C., Spiga, S., & Sabatini, A. (2022). Changes in Dendritic Spine Morphology and Density of Granule Cells in the Olfactory Bulb of Anguilla anguilla (L., 1758): A Possible Way to Understand Orientation and Migratory Behavior. Biology, 11(8), 1244. https://doi.org/10.3390/biology11081244