Efflux Activity Differentially Modulates the Levels of Isoniazid and Rifampicin Resistance among Multidrug Resistant and Monoresistant Mycobacterium tuberculosis Strains

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Results
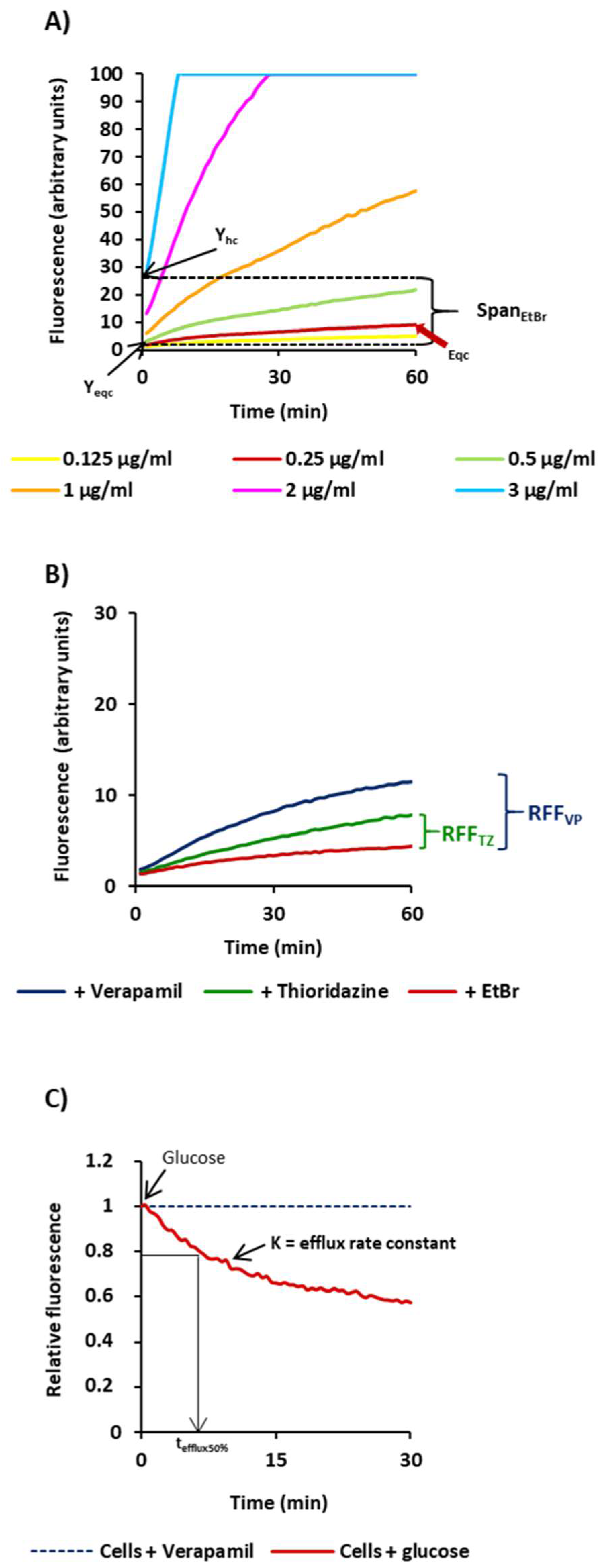
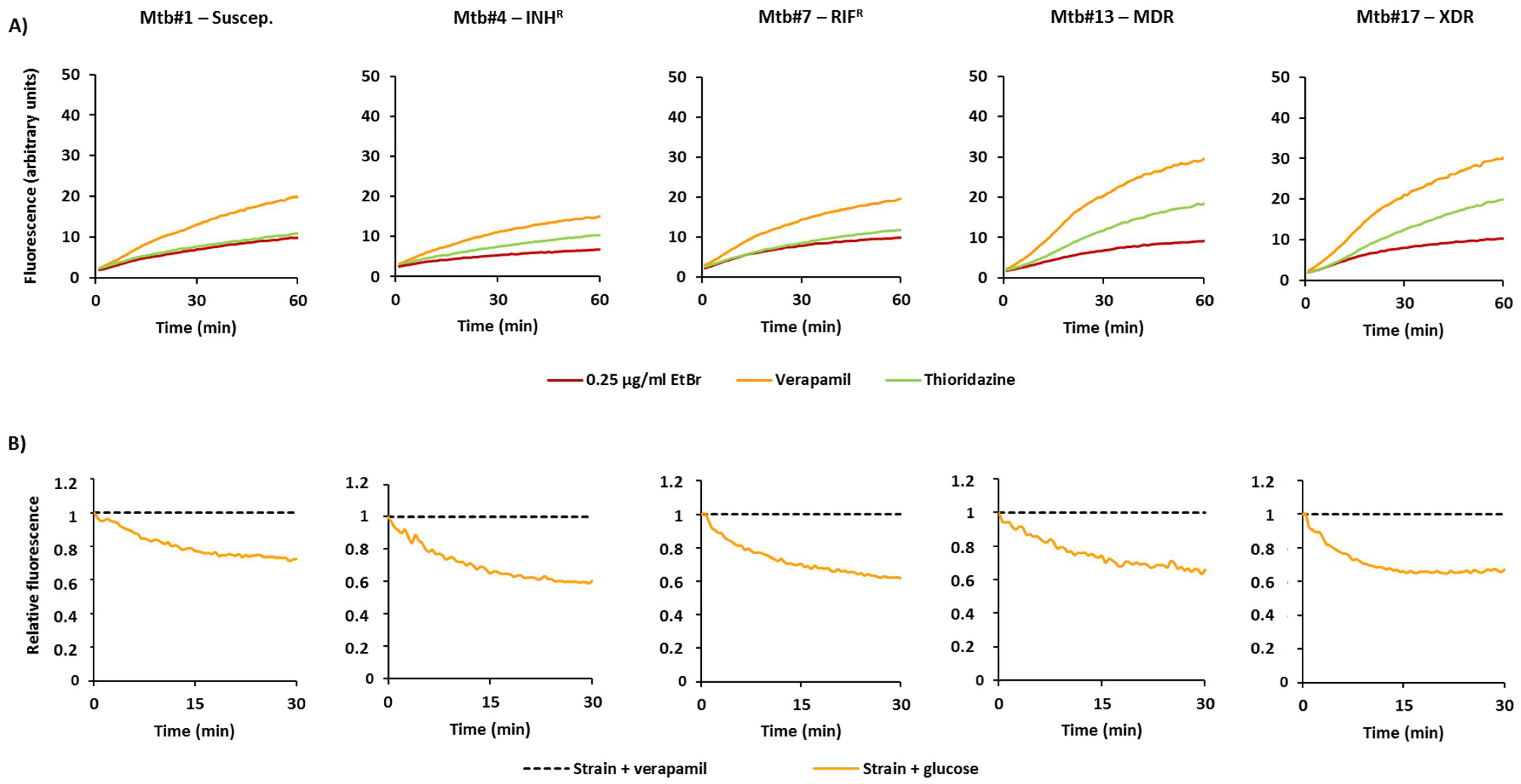
2.1. Characterization of the M. tuberculosis Strains According to Their Efflux Capacity
2.2. Effect of Efflux Inhibitors on the Resistance Levels to Rifampicin and Isoniazid
3. Discussion
4. Materials and Methods
4.1. M. tuberculosis Strains
4.2. Antimicrobials and Reagents
4.3. Susceptibility Testing
4.3.1. Growth of the Strains
4.3.2. First-Line Drug Susceptibility Testing
4.3.3. MIC Determination of Antibiotics and Efflux Inhibitors
4.3.4. Quantitative Drug Susceptibility Testing of Antibiotics in Presence and Absence of Inhibitors
4.4. Ethidium Bromide Real-Time Fluorometry
4.5. Genotypic Characterization of the Strains
- (i)
- DNA extraction: Genomic DNA was extracted using the QIAamp DNA mini kit (QIAGEN, GmbH, Hilden, Germany) according to the manufacturer’s instructions.
- (ii)
- Screening of mutations: The most common mutations in rpoB, katG, and the inhA promoter were screened using the system Genotype MTBDRplus V2 (Hain Lifescience GmbH, Nehren, Germany) according to the manufacturer’s instructions.
- (iii)
- DNA sequencing: The analysis of internal fragments of the genes katG, inhA, and rpoB was performed according to Machado et al. [43].
- (iv)
- Strain typing: Spoligotyping was performed as previously described [44]. Detection of the hybridization patterns was carried out using the ECL Chemiluminescence Detection System (GE Healthcare, Cleveland, OH, USA).
4.6. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AST | antibiotic susceptibility testing |
| d | days |
| EtBr | ethidium bromide |
| h | hours |
| INH | isoniazid |
| K | efflux rate constant |
| LAM | Latin American-Mediterranean |
| MDR | multidrug resistant |
| qDST | quantitative drug susceptibility testing |
| R | resistant |
| RIF | rifampicin |
| Suscep/S | susceptible |
| SIT | spoligotype international type |
| TB | tuberculosis |
| TTD | time to detection |
| TZ | thioridazine |
| VP | verapamil |
| wt | wild-type sequence |
| XDR | extensively drug resistant. |
References
- World Health Organization. Global Tuberculosis Report 2017; WHO/HTM/TB/2017.23; World Health Organization: Geneva, Switzerland, 2017; pp. 1–262. [Google Scholar]
- Böttger, E. The ins and outs of Mycobacterium tuberculosis drug susceptibility testing. Clin. Microbiol. Infect. 2011, 17, 1128–1134. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H. Preventing drug access to targets: Cell surface permeability barriers and active efflux in bacteria. Semin. Cell Dev. Biol. 2001, 12, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Viveiros, M.; Leandro, C.; Amaral, L. Mycobacterial efflux pumps and chemotherapeutic implications. Int. J. Antimicrob. Agents 2003, 22, 274–278. [Google Scholar] [CrossRef]
- Rossi, E.D.; Aínsa, J.A.; Riccardi, G. Role of mycobacterial efflux transporters in drug resistance: An unresolved question. FEMS Microbiol. Rev. 2006, 30, 36–52. [Google Scholar] [CrossRef] [PubMed]
- Louw, G.E.; Warren, R.M.; Gey van Pittius, N.C.; McEvoy, C.R.; van Helden, P.D.; Victor, T.C. A balancing act: Efflux/influx in mycobacterial drug resistance. Antimicrob. Agents Chemother. 2009, 53, 3181–3189. [Google Scholar] [CrossRef] [PubMed]
- Viveiros, M.; Martins, M.; Rodrigues, L.; Machado, D.; Couto, I.; Ainsa, J.; Amaral, L. Inhibitors of mycobacterial efflux pumps as potential boosters for anti-tubercular drugs. Expert Rev. Anti. Infect. Ther. 2012, 10, 983–998. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, P.E.A.; Machado, D.; Ramos, D.; Couto, I.; von Groll, A.; Viveiros, M. Efflux pumps in mycobacteria: Antimicrobial resistance, physiological functions, and role in pathogenicity. In Efflux-Mediated Antimicrobial Resistance in Bacteria, 1st ed.; Li, X.-Z., Elkins, C.A., Zgurskaya, H.I., Eds.; Springer International Publishing: Basel, Switzerland, 2016; pp. 527–559. [Google Scholar] [CrossRef]
- Li, X.-Z.; Elkins, C.A.; Zgurskaya, H.I. Efflux-Mediated Antimicrobial Resistance in Bacteria, 1st ed.; Springer International Publishing: Basel, Switzerland, 2016; pp. 1–848. ISBN 978-3-319-39658-3. [Google Scholar] [CrossRef]
- Webber, M.A.; Piddock, L.J. The importance of efflux pumps in bacterial antibiotic resistance. J. Antimicrob. Chemother. 2003, 51, 9–11. [Google Scholar] [CrossRef] [PubMed]
- Piddock, L.J. Clinically relevant chromosomally encoded multidrug resistance efflux pumps in bacteria. Clin. Microbiol. Rev. 2006, 19, 382–402. [Google Scholar] [CrossRef] [PubMed]
- Ryan, B.; Dougherty, T.; Beaulieu, D.; Chuang, J.; Dougherty, B.; Barrett, J. Efflux in bacteria: What do we really know about it? Expert Opin. Investig. Drugs. 2001, 10, 1409–1422. [Google Scholar] [CrossRef] [PubMed]
- Viveiros, M.; Dupont, M.; Rodrigues, L.; Couto, I.; Davin-Regli, A.; Martins, M.; Pagès, J.; Amaral, L. Antibiotic stress, genetic response and altered permeability of E. coli. PLoS ONE 2007, 2, e365. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, P.E.; Palomino, J.C. Molecular basis and mechanisms of drug resistance in Mycobacterium tuberculosis: Classical and new drugs. J. Antimicrob. Chemother. 2011, 66, 1417–1430. [Google Scholar] [CrossRef] [PubMed]
- Black, P.A.; Warren, R.M.; Louw, G.E.; van Helden, P.D.; Victor, T.C.; Kana, B.D. Energy metabolism and drug efflux in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2014, 58, 2491–2503. [Google Scholar] [CrossRef] [PubMed]
- Andries, K.; Villellas, C.; Coeck, N.; Thys, K.; Gevers, T.; Vranckx, L.; Lounis, N.; de Jong, B.; Koul, A. Acquired resistance of Mycobacterium tuberculosis to bedaquiline. PLoS ONE 2014, 9, e102135. [Google Scholar] [CrossRef] [PubMed]
- Machado, D.; Pires, D.; Perdigão, P.; Couto, I.; Portugal, I.; Martins, M.; Amaral, L.; Anes, E.; Viveiros, M. Ion channel blockers as antimicrobial agents, efflux inhibitors, and enhancers of macrophage killing activity against drug resistant Mycobacterium tuberculosis. PLoS ONE 2016, 11, e0149326. [Google Scholar] [CrossRef] [PubMed]
- Machado, D.; Coelho, T.S.; Perdigão, J.; Pereira, C.; Couto, I.; Portugal, I.; Maschmann, R.; Ramos, D.; von Groll, A.; Rossetti, M.; et al. Interplay between mutations and efflux in drug resistant clinical isolates of Mycobacterium tuberculosis. Front. Microbiol. 2017, 8, 711. [Google Scholar] [CrossRef] [PubMed]
- Machado, D.; Couto, I.; Perdigão, J.; Rodrigues, L.; Portugal, I.; Baptista, P.; Veigas, B.; Amaral, L.; Viveiros, M. Contribution of efflux to the emergence of isoniazid and multidrug resistance in Mycobacterium tuberculosis. PLoS ONE 2012, 7, e34538. [Google Scholar] [CrossRef] [PubMed]
- Schmalstieg, A.M.; Srivastava, S.; Belkaya, S.; Deshpande, D.; Meek, C.; Leff, R.; van Oers, N.S.C.; Gumbo, T. The antibiotic resistance arrow of time: Efflux pump induction is a general first step in the evolution of mycobacterial drug resistance. Antimicrob. Agents Chemother. 2012, 56, 4806–4815. [Google Scholar] [CrossRef] [PubMed]
- Amaral, L.; Martins, M.; Viveiros, M. Enhanced killing of intracellular multidrug-resistant Mycobacterium tuberculosis by compounds that affect the activity of efflux pumps. J. Antimicrob. Chemother. 2007, 59, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- Adams, K.N.; Takaki, K.; Connolly, L.E.; Wiedenhoft, H.; Winglee, K.; Humbert, O.; Edelstein, P.H.; Cosma, C.L.; Ramakrishnan, L. Drug tolerance in replicating mycobacteria mediated by a macrophage-induced efflux mechanism. Cell 2011, 45, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Adams, K.N.; Szumowski, J.D.; Ramakrishnan, L. Verapamil, and its metabolite norverapamil, inhibit macrophage-induced, bacterial efflux pump-mediated tolerance to multiple anti-tubercular drugs. J. Infect. Dis. 2014, 210, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Van Soolingen, D.; Hernandez-Pando, R.; Orozco, H.; Aguilar, D.; Magis-Escurra, C.; Amaral, L.; van Ingen, J.; Boeree, M.J. The antipsychotic thioridazine shows promising therapeutic activity in a mouse model of multidrug-resistant tuberculosis. PLoS ONE 2010, 5, e12640. [Google Scholar] [CrossRef] [PubMed]
- Abbate, E.; Vescovo, M.; Natiello, M.; Cufre, M.; Garcia, A.; Montaner, P.G.; Ambroggi, M.; Ritacco, V.; van Soolingen, D. Successful alternative treatment of extensively drug-resistant tuberculosis in Argentina with a combination of linezolid, moxifloxacin and thioridazine. J. Antimicrob. Chemother. 2012, 67, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Louw, G.E.; Warren, R.M.; Gey van Pittius, N.C.; Leon, R.; Jimenez, A.; Hernandez-Pando, R.; McEvoy, C.R.; Grobbelaar, M.; Murray, M.; van Helden, P.D.; Victor, T.C. Rifampicin reduces susceptibility to ofloxacin in rifampicin-resistant Mycobacterium tuberculosis through efflux. Am. J. Respir. Crit. Care Med. 2011, 184, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Cohen, K.A.; Winglee, K.; Maiga, M.; Diarra, B.; Bishai, W.R. Efflux inhibition with verapamil potentiates bedaquiline in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2014, 58, 574–576. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Tyagi, S.; Almeida, D.V.; Maiga, M.C.; Ammerman, N.C.; Bishai, W.R. Acceleration of tuberculosis treatment by adjunctive therapy with verapamil as an efflux inhibitor. Am. J. Respir. Crit. Care Med. 2013, 188, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Ordway, D.; Viveiros, M.; Leandro, C.; Bettencourt, R.; Almeida, J.; Martins, M.; Kristiansen, J.E.; Molnar, J.; Amaral, L. Clinical concentrations of thioridazine kill intracellular multidrug-resistant Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2003, 47, 917–922. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.; Viveiros, M.; Amaral, L. The TB laboratory of the future: Macrophage-based selection of XDR-TB therapeutics. Future Microbiol. 2008, 3, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Viveiros, M.; Rodrigues, L.; Martins, M.; Couto, I.; Spengler, G.; Martins, A.; Amaral, L. Evaluation of efflux activity of bacteria by a semi-automated fluorometric system. In Antibiotic Resistance Protocols, 2nd ed.; Gillespie, S.H., McHugh, T.D., Eds.; Humana Press: New York, NY, USA, 2010; pp. 159–172. [Google Scholar] [CrossRef]
- Costa, S.; Lopes, E.; Azzali, E.; Machado, D.; Coelho, T.; da Silva, P.E.; Viveiros, M.; Pieroni, M.; Couto, I. An experimental model for the rapid screening of compounds with potential use against mycobacteria. Assay Drug Dev. Technol. 2016, 14, 524–534. [Google Scholar] [CrossRef] [PubMed]
- Pieroni, M.; Machado, D.; Azzali, E.; Costa, S.S.; Couto, I.; Costantino, G.; Viveiros, M. Rational design and synthesis of thioridazine analogues as enhancers of the antituberculosis therapy. J. Med. Chem. 2015, 58, 5842–5853. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Wu, X. Development of efflux pump inhibitors in antituberculosis therapy. Int. J. Antimicrob. Agents. 2016, 47, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Pym, A.S.; Saint-Joanis, B.; Cole, S.T. Effect of katG mutations on the virulence of Mycobacterium tuberculosis and the implication for transmission in humans. Infect. Immun. 2002, 70, 4955–4960. [Google Scholar] [CrossRef] [PubMed]
- Gagneux, S.; Burgos, M.V.; DeRiemer, K.; Enciso, A.; Muñoz, S.; Hopewell, P.C.; Small, P.; Pym, A.S. Impact of bacterial genetics on the transmission of isoniazid-resistant Mycobacterium tuberculosis. PLoS Pathog. 2006, 2, e61. [Google Scholar] [CrossRef] [PubMed]
- Perdigão, J.; Silva, H.; Machado, D.; Macedo, R.; Maltez, F.; Silva, C.; Jordão, L.; Couto, I.; Mallard, K.; Coll, F.; et al. Unraveling Mycobacterium tuberculosis genomic diversity and evolution in Lisbon, Portugal, a highly drug resistant setting. BMC Genom. 2014, 15, 991. [Google Scholar] [CrossRef] [PubMed]
- Cambau, E.; Viveiros, M.; Machado, D.; Raskine, L.; Ritter, C.; Tortoli, E.; Fauville-Dufaux, M.; Hoffner, S.; Richter, E.; Perez del Molino, M.L.; et al. Revisiting susceptibility testing in MDR-TB by a standardized quantitative phenotypic assessment in a European multicentre study. J. Antimicrob. Chemother. 2015, 70, 686–696. [Google Scholar] [CrossRef] [PubMed]
- Springer, B.; Lucke, K.; Calligaris-Maibach, R.; Ritter, C.; Böttger, E.C. Quantitative drug susceptibility testing of Mycobacterium tuberculosis using MGIT960 and the EpiCenter instrumentation. J. Clin. Microbiol. 2009, 47, 1773–1780. [Google Scholar] [CrossRef] [PubMed]
- Paixão, L.; Rodrigues, L.; Couto, I.; Martins, M.; Fernandes, P.; de Carvalho, C.C.; Monteiro, G.A.; Sansonetty, F.; Amaral, L.; Viveiros, M. Fluorometric determination of ethidium bromide efflux kinetics in Escherichia coli. J. Biol. Eng. 2009, 3, 18. [Google Scholar] [CrossRef] [PubMed]
- Machado, L.; Spengler, G.; Evaristo, M.; Handzlik, J.; Molnar, J.; Viveiros, M.; Kiec-Kononowicz, K.; Amaral, L. Biological activity of twenty-three hydantoin derivatives on intrinsic efflux pump system of Salmonella enterica serovar Enteritidis NCTC 13349. In Vivo 2011, 25, 769–772. [Google Scholar] [PubMed]
- Iyer, R.; Ferrari, A.; Rijnbrand, R.; Erwin, A.L. A fluorescent microplate assay quantifies bacterial efflux and demonstrates two distinct compound binding sites in AcrB. Antimicrob. Agents Chemother. 2015, 59, 2388–2397. [Google Scholar] [CrossRef] [PubMed]
- Machado, D.; Perdigão, J.; Ramos, J.; Couto, I.; Portugal, I.; Ritter, C.; Boettger, E.C.; Viveiros, M. High-level resistance to isoniazid and ethionamide in multidrug-resistant Mycobacterium tuberculosis of the Lisboa family is associated with inhA double mutations. J. Antimicrob. Chemother. 2013, 68, 1728–1732. [Google Scholar] [CrossRef] [PubMed]
- Kamerbeek, J.; Schouls, L.; Kolk, A.; van Agterveld, M.; van Soolingen, D.; Kuijper, S.; Bunschoten, A.; Molhuizen, H.; Shaw, R.; Goyal, M.; et al. Simultaneous detection and strain differentiation of Mycobacterium tuberculosis for diagnosis and epidemiology. J. Clin. Microbiol. 1997, 35, 907–914. [Google Scholar]



{kind=link}
{kind=link}
| Strain | Spoligotype SIT/Clade | AST | Drug Resistance Mutational Background | MICs (µg/mL) | ||||
|---|---|---|---|---|---|---|---|---|
| Antibiotics | Efflux Inhibitors | |||||||
| INH | RIF | INH | RIF | VP | TZ | |||
| H37Rv | H37Rv | Suscep. | wt | wt | 0.1 | 1 | 256 | 15 |
| Mtb1 | LAM9—Orphan | Suscep. | wt | wt | 0.1 | 1 | 256 | 30 |
| Mtb2 | Unknown—SIT105 | Suscep. | wt | wt | 0.1 | 1 | 256 | 15 |
| Mtb3 | T1—SIT612 | INHR | katG S315T | wt | 10 | 1 | 256 | 15 |
| Mtb4 | LAM1—SIT20 | INHR | inhA C-15T | wt | 0.4 | 1 | 256 | 15 |
| Mtb5 | LAM2—SIT53 | RIFR | wt | rpoB H526D | 0.1 | 320 | 256 | 15 |
| Mtb6 | Beijing—SIT1 | RIFR | wt | rpoB H526Y | 0.1 | 320 | 128 | 15 |
| Mtb7 | LAM1—SIT2576 | RIFR | wt | rpoB S531L | 0.1 | 320 | 256 | 15 |
| Mtb8 | LAM1—SIT2576 | RIFR | wt | rpoB S531L | 0.1 | 640 | 256 | 15 |
| Mtb9 | LAM1—SIT20 | MDR | inhA C-15T/S94A | rpoB D516V | 3 | 20 | 256 | 15 |
| Mtb10 | T1—SIT53 | MDR | katG S315N | rpoB H526Y | 3 | 20 | 256 | 15 |
| Mtb11 | T1—SIT53 | MDR | katG S315T | rpoB S531L | 10 | 640 | 256 | 15 |
| Mtb12 | T2—Orphan | MDR | katG S315T | rpoB S531L | 10 | 160 | 128 | 15 |
| Mtb13 | LAM1—SIT20 | MDR | inhA C-15T/S94A | rpoB S531L | 3 | 320 | 256 | 15 |
| Mtb14 | Beijing—SIT1 | MDR | katG S315T | rpoB S531L | 20 | 320 | 256 | 15 |
| Mtb15 | LAM1—SIT20 | XDR | inhA C-15T/S94A | rpoB S531L | 20 | 80 | 128 | 15 |
| Mtb16 | LAM4—SIT1106 | XDR | inhA C-15T/I194T | rpoB S531L | 3 | 320 | 256 | 15 |
| Mtb17 | LAM4—SIT1106 | XDR | inhA C-15T/I194T | rpoB S531L | 3 | 320 | 256 | 15 |
| Strains | Efflux Activity | ||||
|---|---|---|---|---|---|
| SpanEtBr | RFFVP | RFFTZ | K | tefflux50% | |
| Suscep. | |||||
| H37Rv | 26.10 | 1.57 ± 0.02 | 0.77 ± 0.01 | 0.09 ± 0.008 | 7.94 ± 0.75 |
| Mtb#1 | 29.42 | 1.04 ± 0.03 | 0.13 ± 0.02 | 0.09 ± 0.086 | 8.06 ± 0.63 |
| Mtb#2 | 28.51 | 2.07 ± 0.09 | 1.18 ± 0.03 | 0.09 ± 0.003 | 7.77 ± 0.30 |
| INHR | |||||
| Mtb#3 | 14.07 | 0.59 ± 0.02 | 0.90 ± 0.04 | 0.06 ± 0.008 | 11.98 ± 1.81 |
| Mtb#4 | 13.50 | 1.21 ± 0.02 | 0.13 ± 0.05 | 0.08 ± 0.004 | 8.95 ± 0.48 |
| RIFR | |||||
| Mtb#5 | 12.13 | 1.00 ± 0.03 | 0.41 ± 0.01 | 0.07 ± 0.009 | 8.55 ± 0.47 |
| Mtb#6 | 13.50 | 2.31 ± 0.02 | 1.11 ± 0.01 | 0.12 ± 0.004 | 5.95 ± 0.21 |
| Mtb#7 | 10.46 | 1.26 ± 0.06 | 0.13 ± 0.05 | 0.10 ± 0.0004 | 6.67 ± 0.02 |
| Mtb#8 | 13.16 | 1.16 ± 0.03 | 0.28 ± 0.01 | 0.14 ± 0.004 | 4.39 ± 0.41 |
| MDR | |||||
| Mtb#9 | 22.39 | 1.62 ± 0.09 | 0.79 ± 0.04 | 0.11 ± 0.0006 | 6.49 ± 0.04 |
| Mtb#11 | 7.45 | 1.31 ± 0.36 | 0.39 ± 0.06 | 0.12 ± 0.0057 | 5.90 ± 0.42 |
| Mtb#12 | 8.86 | 0.71 ± 0.00 | 0.30 ± 0.09 | 0.05 ± 0.0011 | 14.06 ± 0.46 |
| Mtb#13 | 11.74 | 2.18 ± 0.01 | 0.98 ± 0.02 | 0.10 ± 0.0095 | 7.25 ± 0.72 |
| Mtb#14 | 12.19 | 1.21 ± 0.15 | 0.89 ± 0.06 | 0.09 ± 0.0036 | 8.08 ± 0.33 |
| XDR | |||||
| Mtb#16 | 13.56 | 1.15 ± 0.17 | 1.05 ± 0.01 | 0.10 ± 0.007 | 6.89 ± 0.69 |
| Mtb#17 | 14.80 | 1.78 ± 0.22 | 0.91 ± 0.23 | 0.14 ± 0.022 | 4.99 ± 0.78 |
| Strain | MIC (µg/mL) | MIC (µg/mL) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| INH | INH + VP | INH + TZ | RIF | RIF + VP | RIF + TZ | |||||||||||
| 0.1 | 1 | 3 | 10 | 0.1 | 1 | 3 | 10 | 1 | 4 | 20 | 1 | 4 | 20 | |||
| Suscep. | ||||||||||||||||
| H37Rv | 0.1 | S | - | - | - | - | - | - | - | 1 | S | - | - | - | - | - |
| Mtb1 | 0.1 | S | - | - | - | - | - | - | - | 1 | S | - | - | - | - | - |
| Mtb2 | 0.1 | S | - | - | - | - | - | - | - | 1 | S | - | - | - | - | - |
| INHR | ||||||||||||||||
| Mtb3 | 10 | R | R | R | S | R | R | R | S | 1 | - | - | - | - | - | - |
| Mtb4 | 1 | R | S | S | S | R | S | S | S | 1 | - | - | - | - | - | - |
| RIFR | ||||||||||||||||
| Mtb5 | 0.1 | - | - | - | - | - | - | - | - | 340 | R | R | R | R | R | R |
| Mtb6 | 0.1 | - | - | - | - | - | - | - | - | 320 | R | R | R | R | R | R |
| Mtb7 | 0.1 | - | - | - | - | - | - | - | - | 320 | R | R | R | R | R | R |
| Mtb8 | 0.1 | - | - | - | - | - | - | - | - | 640 | R | R | R | R | R | R |
| MDR | ||||||||||||||||
| Mtb9 | 3 | R | S | S | - | R | S | S | - | 20 | R | S | S | R | S | S |
| Mtb10 | 3 | R | R | S | - | R | R | S | - | 20 | S | S | S | S | S | S |
| Mtb11 | 10 | R | R | S | S | R | R | R | S | 640 | R | R | R | R | R | R |
| Mtb12 | 10 | R | R | S | S | R | R | S | S | 160 | R | R | S | R | R | R |
| Mtb13 | 3 | R | S | S | - | R | S | S | - | 320 | R | R | S | R | R | R |
| Mtb14 | 20 | R | R | R | S | R | R | S | S | 320 | R | R | S | R | R | R |
| XDR | ||||||||||||||||
| Mtb15 | 20 | R | S | S | S | R | R | R | R | 80 | R | R | S | R | R | S |
| Mtb16 | 3 | R | S | S | - | R | S | S | - | 320 | R | R | S | R | R | R |
| Mtb17 | 3 | R | S | S | - | R | R | S | - | 320 | R | R | S | R | R | R |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Machado, D.; Perdigão, J.; Portugal, I.; Pieroni, M.; Silva, P.A.; Couto, I.; Viveiros, M. Efflux Activity Differentially Modulates the Levels of Isoniazid and Rifampicin Resistance among Multidrug Resistant and Monoresistant Mycobacterium tuberculosis Strains. Antibiotics 2018, 7, 18. https://doi.org/10.3390/antibiotics7010018
Machado D, Perdigão J, Portugal I, Pieroni M, Silva PA, Couto I, Viveiros M. Efflux Activity Differentially Modulates the Levels of Isoniazid and Rifampicin Resistance among Multidrug Resistant and Monoresistant Mycobacterium tuberculosis Strains. Antibiotics. 2018; 7(1):18. https://doi.org/10.3390/antibiotics7010018
Chicago/Turabian StyleMachado, Diana, João Perdigão, Isabel Portugal, Marco Pieroni, Pedro A. Silva, Isabel Couto, and Miguel Viveiros. 2018. "Efflux Activity Differentially Modulates the Levels of Isoniazid and Rifampicin Resistance among Multidrug Resistant and Monoresistant Mycobacterium tuberculosis Strains" Antibiotics 7, no. 1: 18. https://doi.org/10.3390/antibiotics7010018
APA StyleMachado, D., Perdigão, J., Portugal, I., Pieroni, M., Silva, P. A., Couto, I., & Viveiros, M. (2018). Efflux Activity Differentially Modulates the Levels of Isoniazid and Rifampicin Resistance among Multidrug Resistant and Monoresistant Mycobacterium tuberculosis Strains. Antibiotics, 7(1), 18. https://doi.org/10.3390/antibiotics7010018