Killing of Staphylococci by θ-Defensins Involves Membrane Impairment and Activation of Autolytic Enzymes

Abstract
:
1. Introduction
2. Results and Discussion
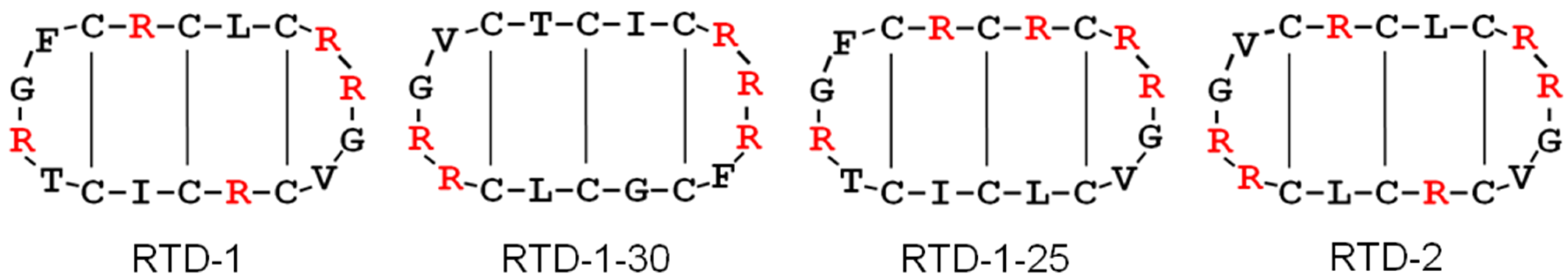

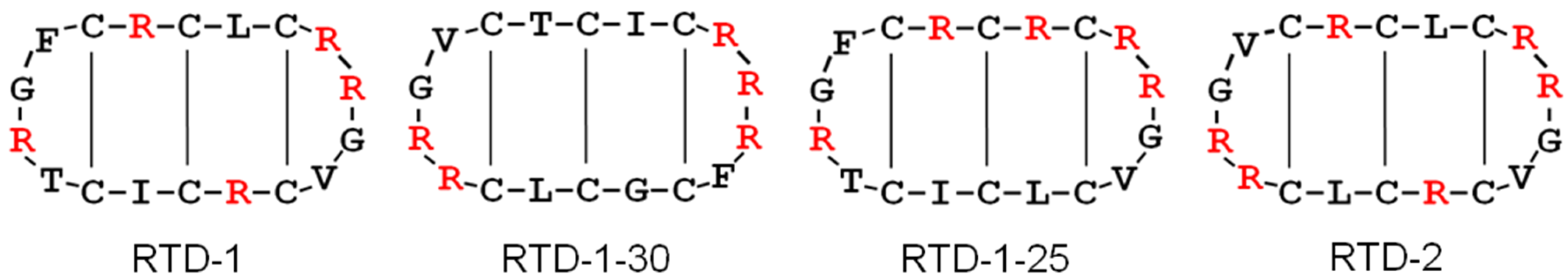
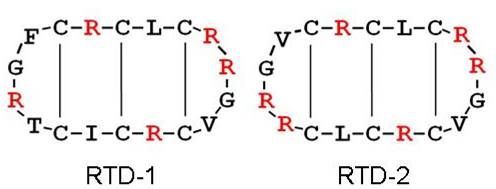{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RTD-1 | RTD-2 | RTD-1-30 | RTD-1-25 | |
|---|---|---|---|---|
| S. aureus SG511-Berlin | 6 ± 2 | 4 ± 0 | 6 ± 2 | 8 ± 4 |
| S. simulans 22 | 1.5 ± 0.5 | 1.5 ± 0.5 | 3 ± 1 | 3 ± 1 |
| S. carnosus TM300 | 0.75 ± 0.25 | 0.5 ± 0 | ND | ND |
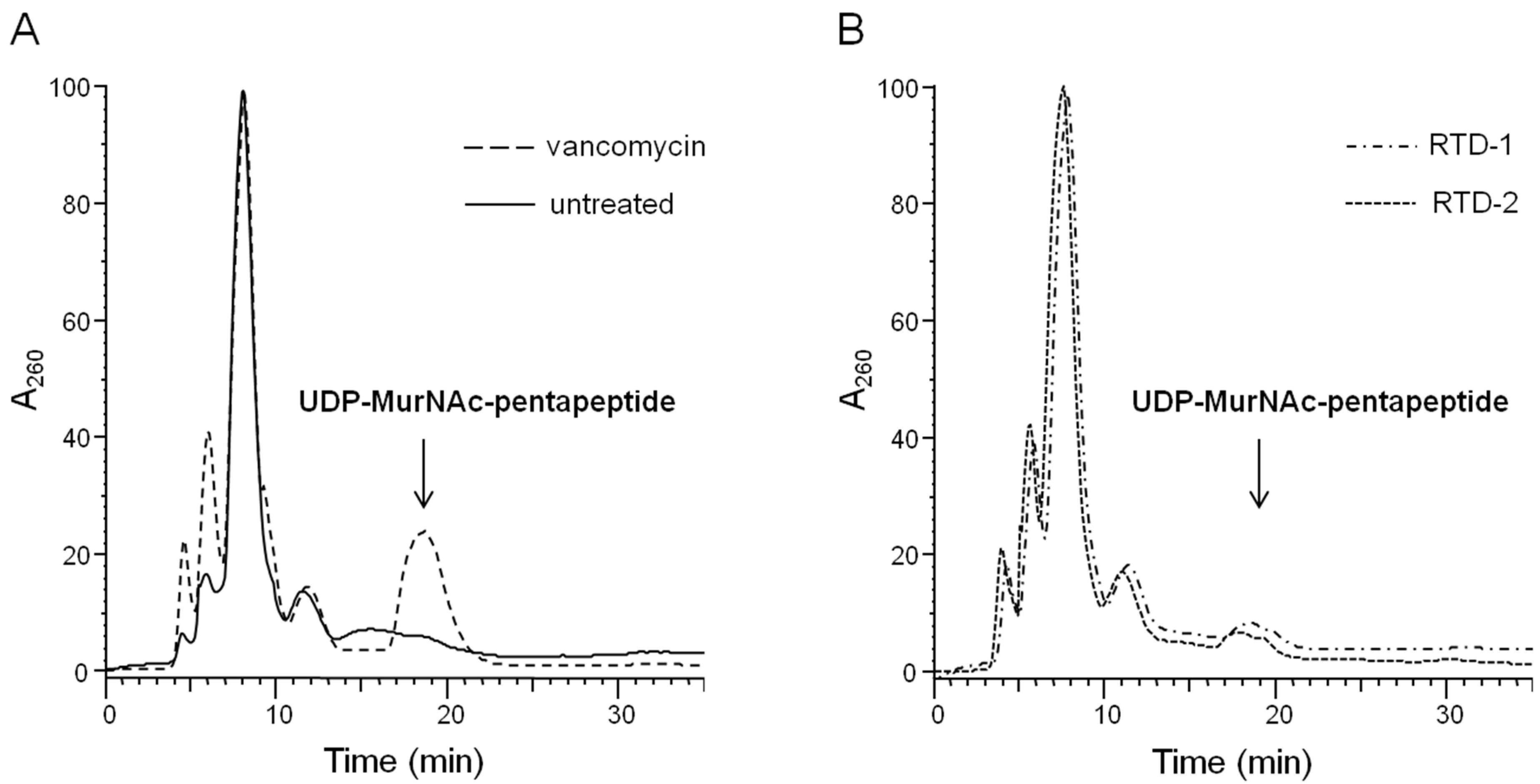
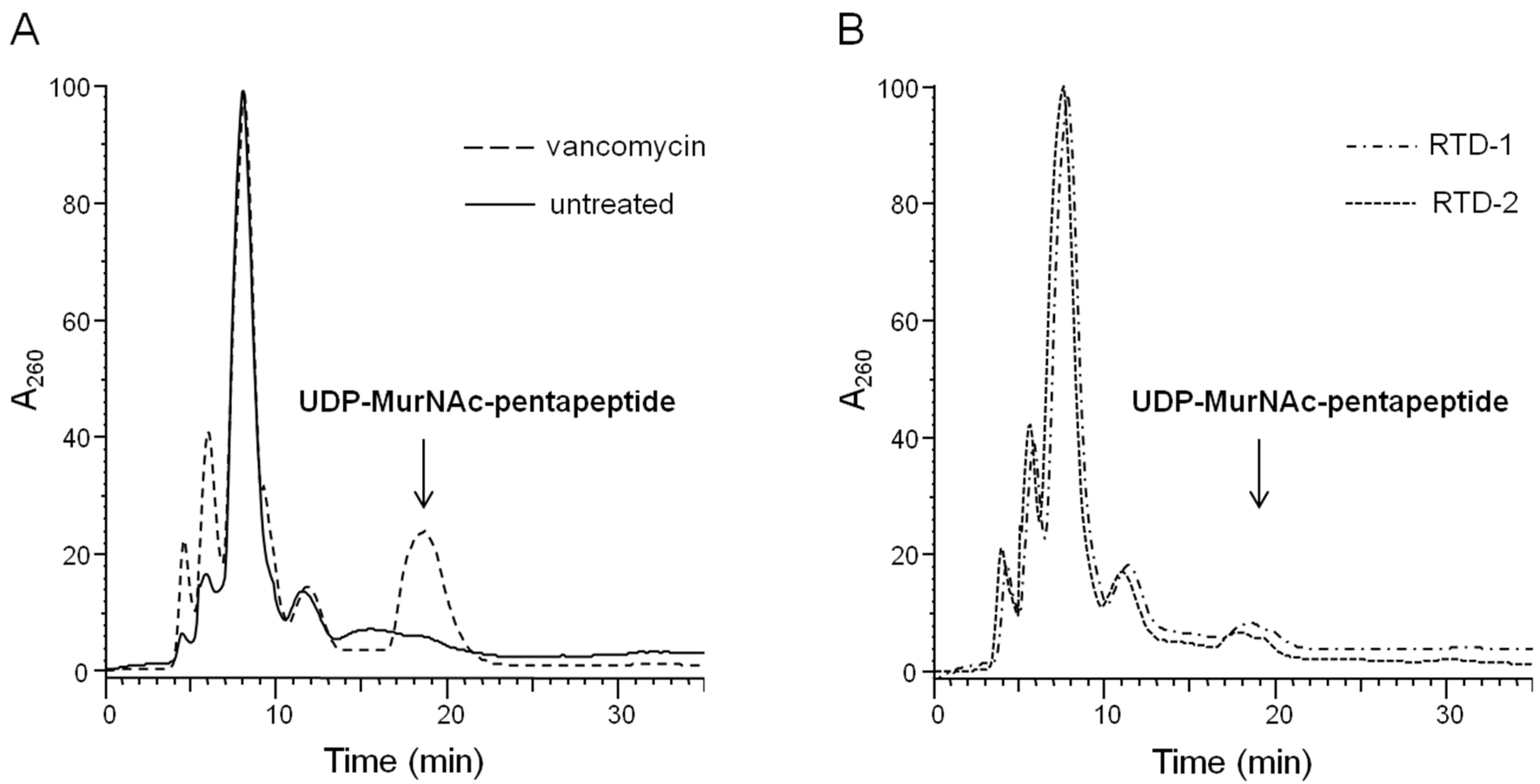
2.1. Impact on Bacterial Cell Wall Biosynthesis
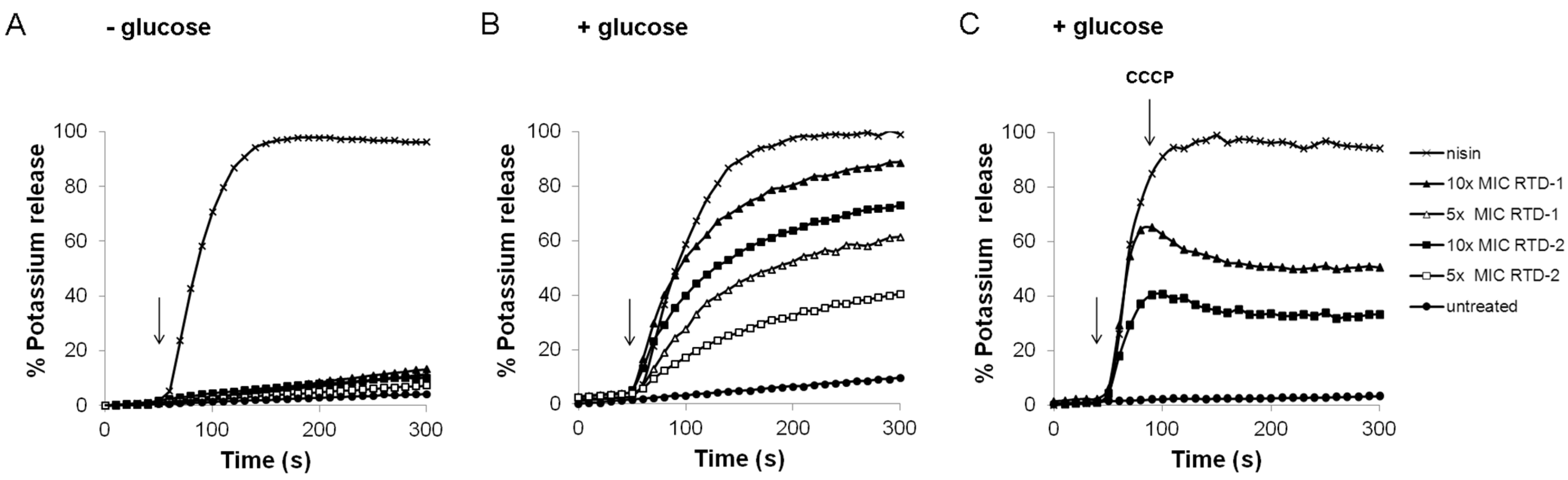
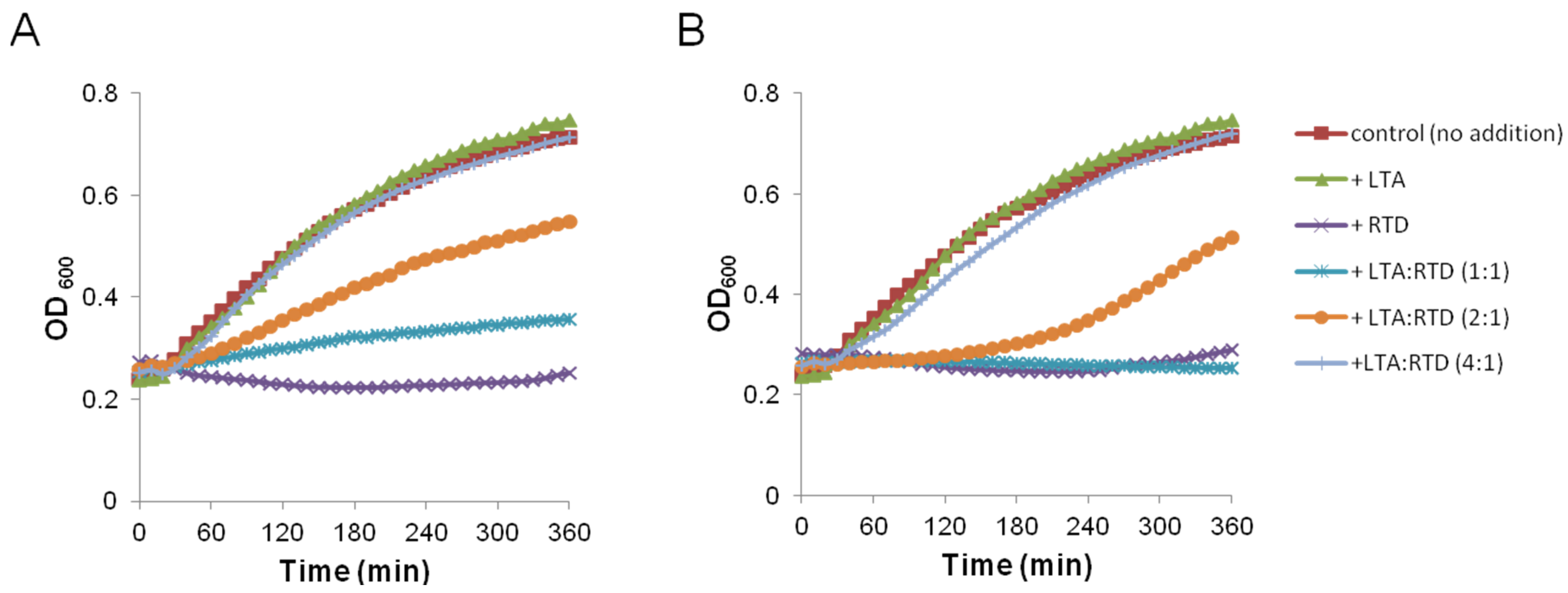
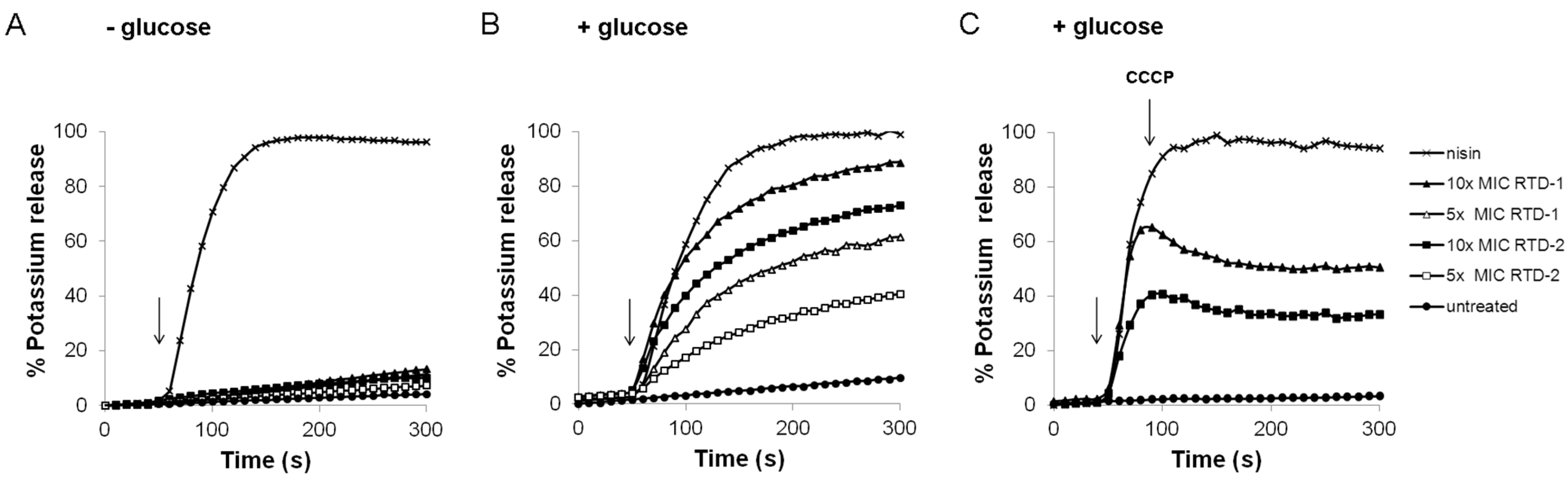
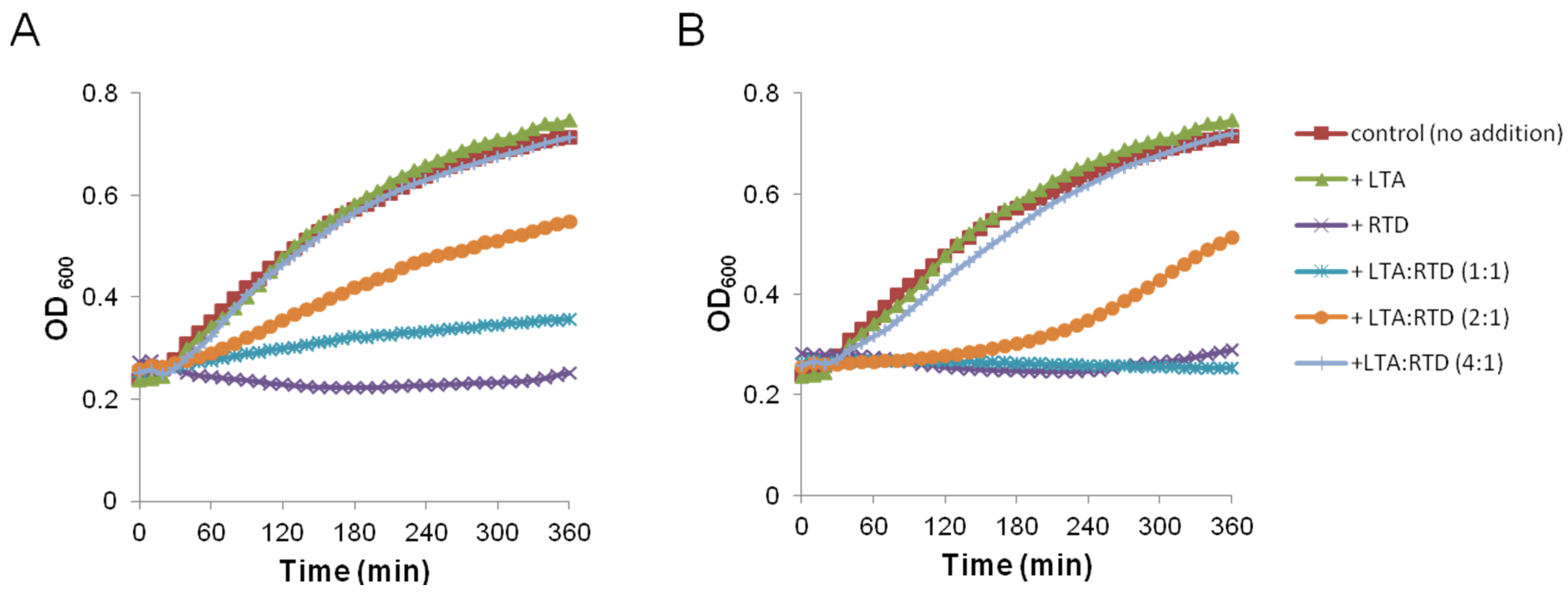
2.2. Impact on Membrane Integrity
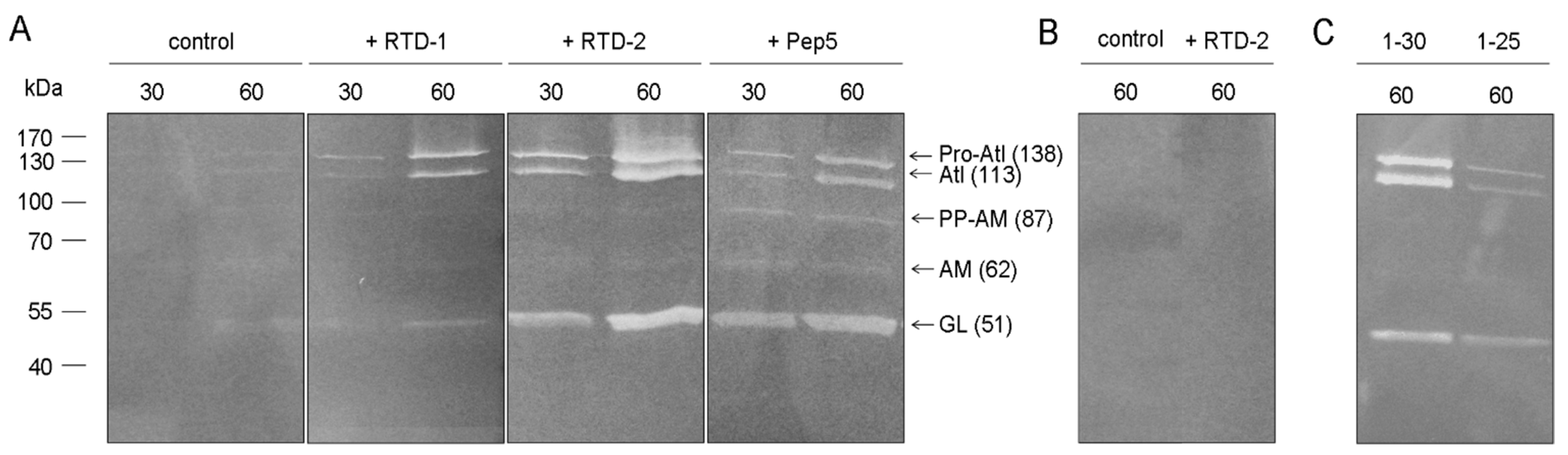
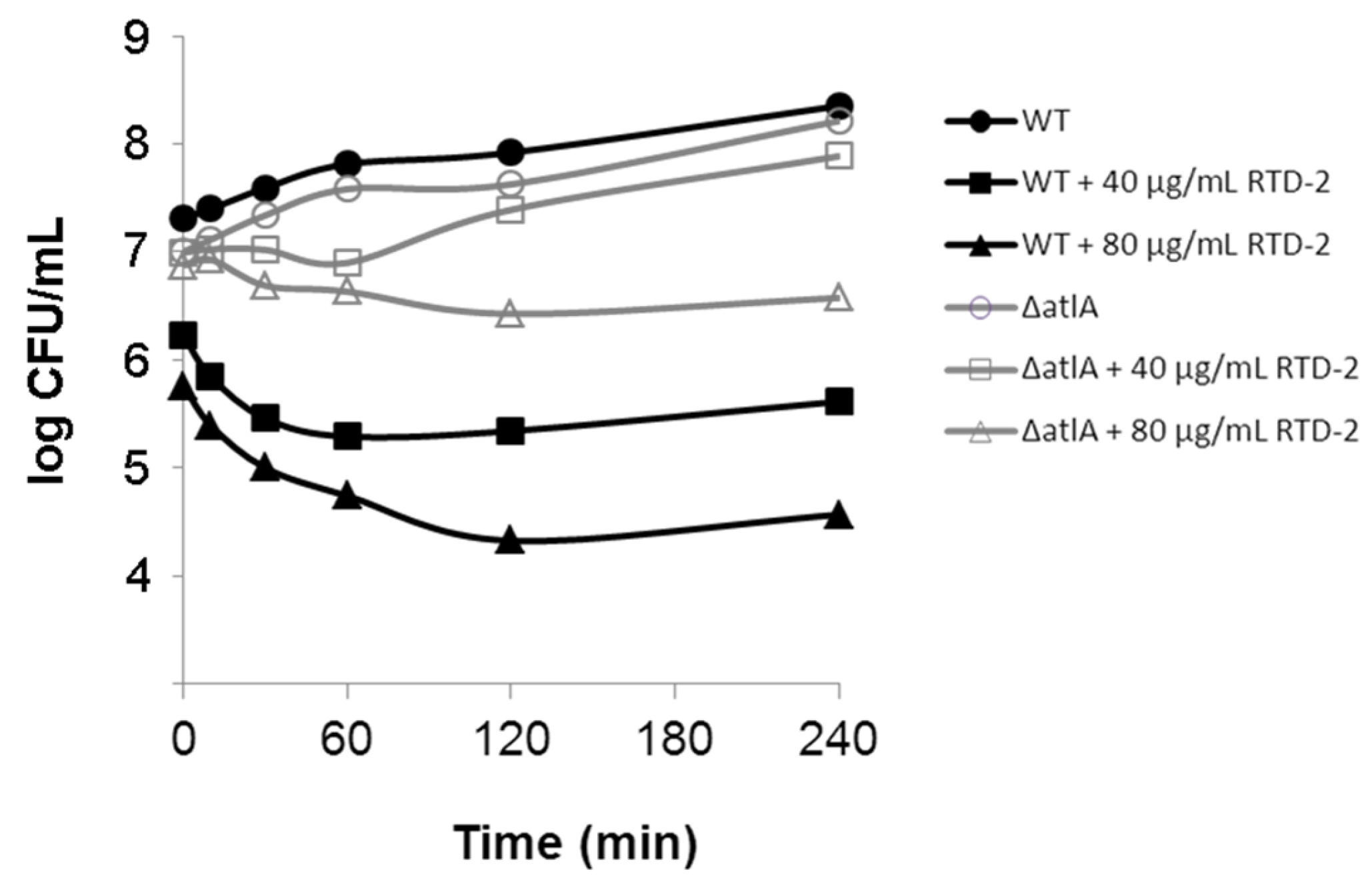
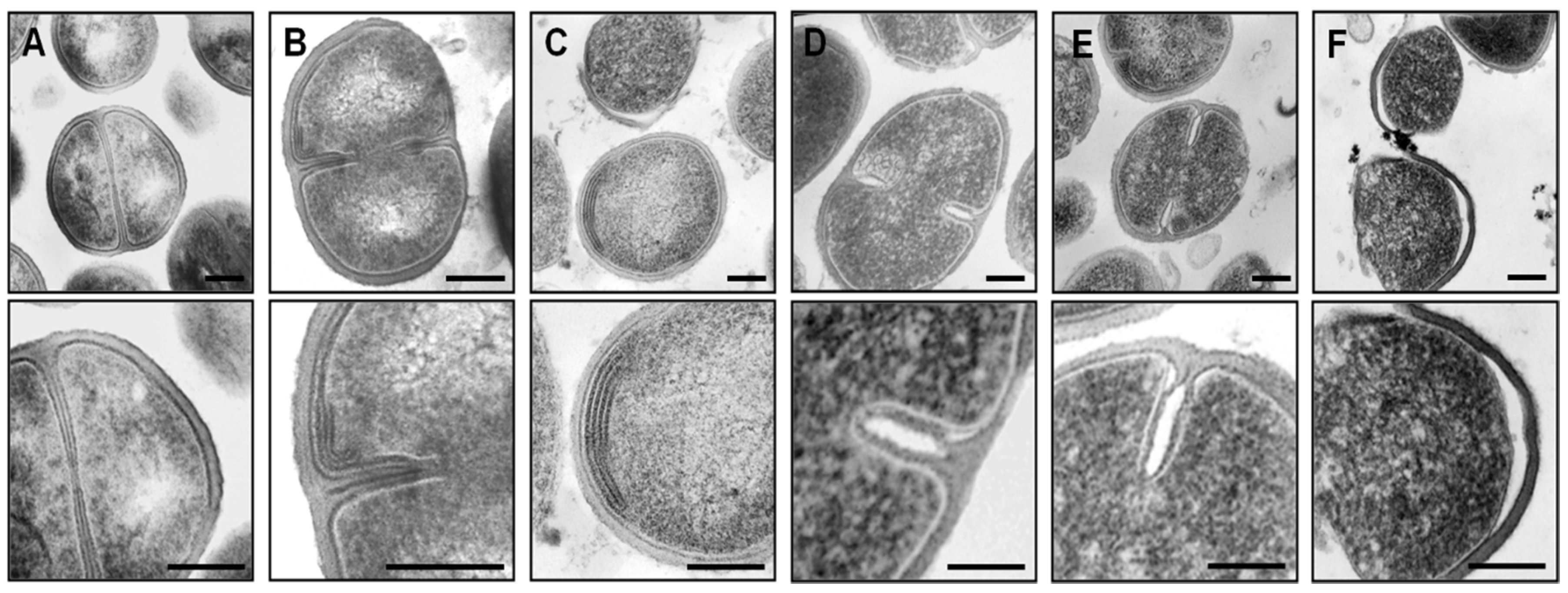
2.3. Impact on Autolytic Enzymes
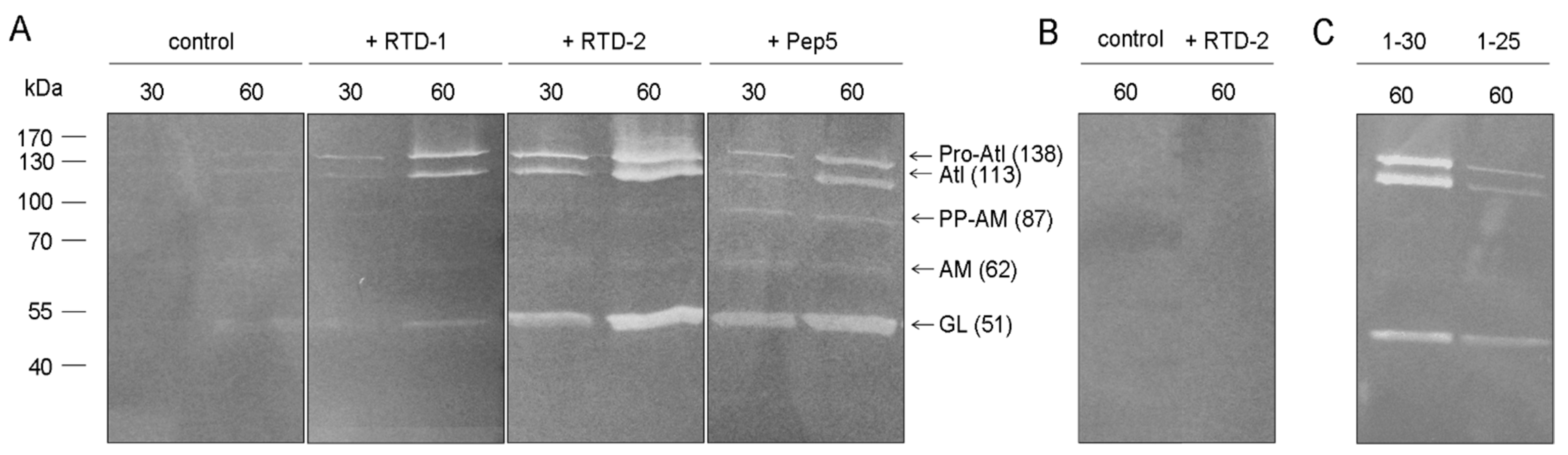
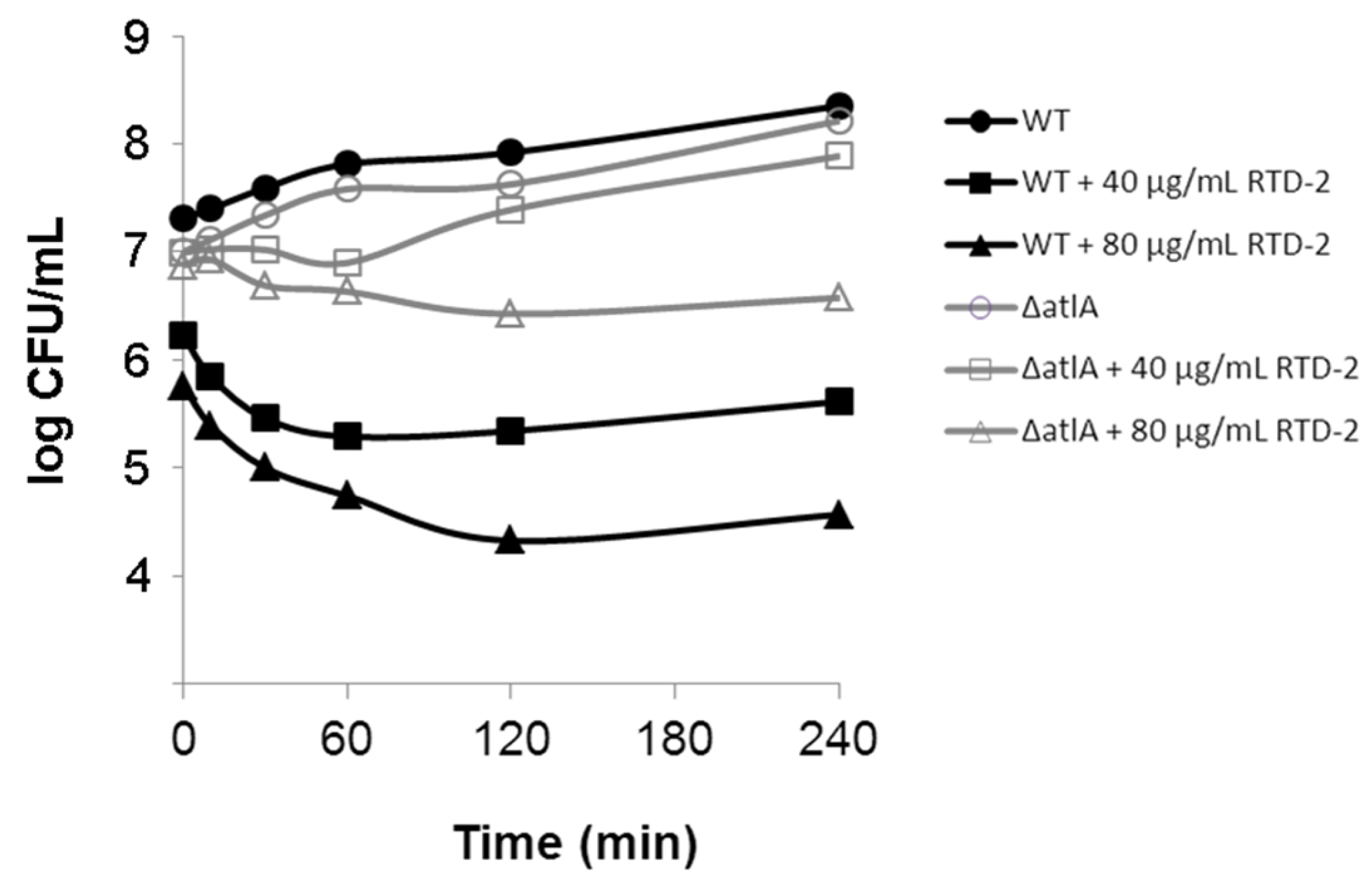
2.4. Activity against Gram-Negative Bacteria
3. Experimental Section
3.1. Peptide Synthesis and Purification
3.2. Determination of Minimal Inhibitory Concentration (MIC)
3.3. Bacterial Killing Kinetics
3.4. Intracellular Accumulation of the Final Soluble Cell Wall Precursor UDP-MurNAc-Pentapeptide
3.5. Potassium Release from Whole Cells
3.6. Estimation of Membrane Potential
3.7. Zymogram Analysis
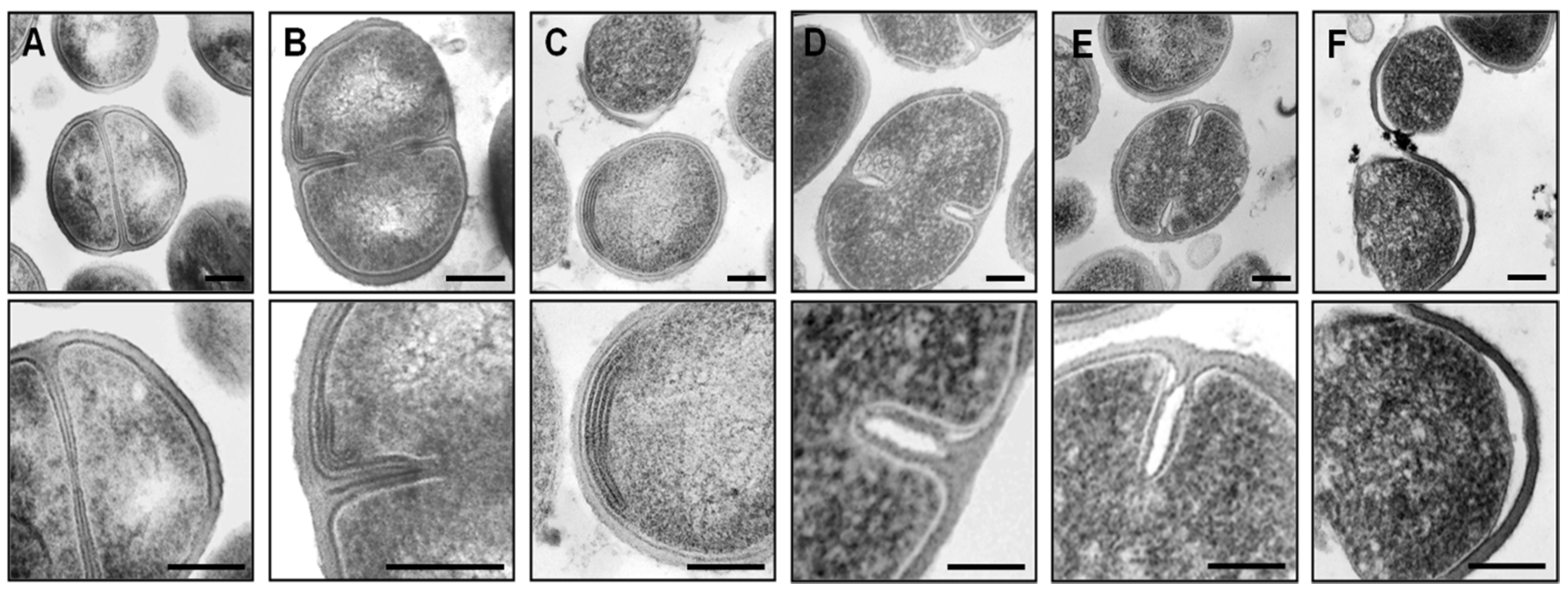
3.8. Transmission Electron Microscopy
3.9. Growth Kinetic in Presence of LTA
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jenssen, H.; Hamill, P.; Hancock, R.E. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar]
- Yeung, A.T.; Gellatly, S.L.; Hancock, R.E. Multifunctional cationic host defence peptides and their clinical applications. Cell. Mol. Life Sci. 2011, 68, 2161–2176. [Google Scholar] [CrossRef] [PubMed]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Hilchie, A.L.; Wuerth, K.; Hancock, R.E. Immune modulation by multifaceted cationic host defense (antimicrobial) peptides. Nat. Chem. Biol. 2013, 9, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Selsted, M.E.; Ouellette, A.J. Mammalian defensins in the antimicrobial immune response. Nat. Immunol. 2005, 6, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T.; Metcalf, J.A.; Gallin, J.I.; Boxer, L.A.; Lehrer, R.I. Microbicidal/cytotoxic proteins of neutrophils are deficient in two disorders: Chediak-Higashi syndrome and “specific” granule deficiency. J. Clin. Invest. 1988, 82, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Wehkamp, J.; Salzman, N.H.; Porter, E.; Nuding, S.; Weichenthal, M.; Petras, R.E.; Shen, B.; Schaeffeler, E.; Schwab, M.; Linzmeier, R.; et al. Reduced Paneth cell alpha-defensins in ileal Crohn’s disease. Proc. Natl. Acad. Sci. USA 2005, 102, 18129–18134. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T.; Lehrer, R.I. Antimicrobial peptides of vertebrates. Curr. Opin. Immunol. 1998, 10, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Defensins: Antimicrobial peptides of innate immunity. Nat. Rev. Immunol. 2003, 3, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.; Diamond, G. The role of cationic antimicrobial peptides in innate host defences. Trends Microbiol. 2000, 8, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Rodriguez Mdel, C.; Lanz-Mendoza, H.; Zhu, S. AdDLP, a bacterial defensin-like peptide, exhibits anti-Plasmodium activity. Biochem. Biophys. Res. Commun. 2009, 387, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Yeaman, M.R.; Yount, N.Y. Unifying themes in host defence effector polypeptides. Nat. Rev. Microbiol. 2007, 5, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.E.; Osapay, G.; Tran, P.A.; Yuan, J.; Selsted, M.E. Isolation, synthesis, and antimicrobial activities of naturally occurring theta-defensin isoforms from baboon leukocytes. Infect. Immun. 2008, 76, 5883–5891. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.Q.; Yuan, J.; Osapay, G.; Osapay, K.; Tran, D.; Miller, C.J.; Ouellette, A.J.; Selsted, M.E. A cyclic antimicrobial peptide produced in primate leukocytes by the ligation of two truncated alpha-defensins. Science 1999, 286, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Tran, D.; Tran, P.A.; Tang, Y.Q.; Yuan, J.; Cole, T.; Selsted, M.E. Homodimeric theta-defensins from rhesus macaque leukocytes: isolation, synthesis, antimicrobial activities, and bacterial binding properties of the cyclic peptides. J. Biol. Chem. 2002, 277, 3079–3084. [Google Scholar] [CrossRef] [PubMed]
- Tongaonkar, P.; Tran, P.; Roberts, K.; Schaal, J.; Osapay, G.; Tran, D.; Ouellette, A.J.; Selsted, M.E. Rhesus macaque theta-defensin isoforms: expression, antimicrobial activities, and demonstration of a prominent role in neutrophil granule microbicidal activities. J. Leukoc. Biol. 2011, 89, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Schaal, J.B.; Tran, D.; Tran, P.; Osapay, G.; Trinh, K.; Roberts, K.D.; Brasky, K.M.; Tongaonkar, P.; Ouellette, A.J.; Selsted, M.E. Rhesus macaque theta defensins suppress inflammatory cytokines and enhance survival in mouse models of bacteremic sepsis. PLoS One 2012, 7, e51337. [Google Scholar] [CrossRef] [PubMed]
- Cole, A.M.; Hong, T.; Boo, L.M.; Nguyen, T.; Zhao, C.; Bristol, G.; Zack, J.A.; Waring, A.J.; Yang, O.O.; Lehrer, R.I. Retrocyclin: A primate peptide that protects cells from infection by T- and M-tropic strains of HIV-1. Proc. Natl. Acad. Sci. USA 2002, 99, 1813–1818. [Google Scholar] [CrossRef] [PubMed]
- Gallo, S.A.; Wang, W.; Rawat, S.S.; Jung, G.; Waring, A.J.; Cole, A.M.; Lu, H.; Yan, X.; Daly, N.L.; Craik, D.J.; et al. Theta-defensins prevent HIV-1 Env-mediated fusion by binding gp41 and blocking 6-helix bundle formation. J. Biol. Chem. 2006, 281, 18787–18792. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, N.; Cole, A.L.; Ruchala, P.; Waring, A.J.; Lehrer, R.I.; Stuchlik, O.; Pohl, J.; Cole, A.M. Reawakening retrocyclins: ancestral human defensins active against HIV-1. PLoS Biol. 2009, 7, e95. [Google Scholar] [CrossRef] [PubMed]
- De Leeuw, E.; Li, C.; Zeng, P.; Diepeveen-de Buin, M.; Lu, W.Y.; Breukink, E.; Lu, W. Functional interaction of human neutrophil peptide-1 with the cell wall precursor lipid II. FEBS Lett. 2010, 584, 1543–1548. [Google Scholar] [CrossRef] [PubMed]
- Oeemig, J.S.; Lynggaard, C.; Knudsen, D.H.; Hansen, F.T.; Norgaard, K.D.; Schneider, T.; Vad, B.S.; Sandvang, D.H.; Nielsen, L.A.; Neve, S.; et al. Eurocin, a new fungal defensin: Structure, lipid binding, and its mode of action. J. Biol. Chem. 2012, 287, 42361–42372. [Google Scholar] [CrossRef]
- Sass, V.; Schneider, T.; Wilmes, M.; Korner, C.; Tossi, A.; Novikova, N.; Shamova, O.; Sahl, H.G. Human beta-defensin 3 inhibits cell wall biosynthesis in Staphylococci. Infect. Immun. 2010, 78, 2793–2800. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, P.; Wilmes, M.; Pugniere, M.; Aumelas, A.; Bachere, E.; Sahl, H.G.; Schneider, T.; Destoumieux-Garzon, D. Insight into invertebrate defensin mechanism of action: Oyster defensins inhibit peptidoglycan biosynthesis by binding to lipid II. J. Biol. Chem. 2010, 285, 29208–29216. [Google Scholar] [CrossRef] [PubMed]
- Schneider, T.; Kruse, T.; Wimmer, R.; Wiedemann, I.; Sass, V.; Pag, U.; Jansen, A.; Nielsen, A.K.; Mygind, P.H.; Raventos, D.S.; et al. Plectasin, a fungal defensin, targets the bacterial cell wall precursor Lipid II. Science 2010, 328, 1168–1172. [Google Scholar] [CrossRef] [PubMed]
- Thevissen, K.; Warnecke, D.C.; Francois, I.E.; Leipelt, M.; Heinz, E.; Ott, C.; Zahringer, U.; Thomma, B.P.; Ferket, K.K.; Cammue, B.P. Defensins from insects and plants interact with fungal glucosylceramides. J. Biol. Chem. 2004, 279, 3900–3905. [Google Scholar] [CrossRef] [PubMed]
- Thevissen, K.; de Mello Tavares, P.; Xu, D.; Blankenship, J.; Vandenbosch, D.; Idkowiak-Baldys, J.; Govaert, G.; Bink, A.; Rozental, S.; de Groot, P.W.; et al. The plant defensin RsAFP2 induces cell wall stress, septin mislocalization and accumulation of ceramides in Candida albicans. Mol. Microbiol. 2012, 84, 166–180. [Google Scholar] [CrossRef] [PubMed]
- Wilmes, M.; Sahl, H.G.; University of Bonn, Bonn, Germany. Unpublished data. 2012.
- Sahl, H.G. Influence of the staphylococcinlike peptide Pep 5 on membrane potential of bacterial cells and cytoplasmic membrane vesicles. J. Bacteriol. 1985, 162, 833–836. [Google Scholar] [PubMed]
- Bierbaum, G.; Sahl, H.G. Induction of autolysis of staphylococci by the basic peptide antibiotics Pep 5 and nisin and their influence on the activity of autolytic enzymes. Arch. Microbiol. 1985, 141, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Bierbaum, G.; Sahl, H.G. Autolytic system of Staphylococcus simulans 22: Influence of cationic peptides on activity of N-acetylmuramoyl-L-alanine amidase. J. Bacteriol. 1987, 169, 5452–5458. [Google Scholar] [PubMed]
- Lahav, M.; Ginsburg, I. Effect of leukocyte hydrolases on bacteria. X. The role played by leukocyte factors, cationic polyelectrolytes, and by membrane-damaging agents in the lysis of Staphylococcus aureus: Relation to chronic inflammatory processes. Inflammation 1977, 2, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Ginsburg, I.; Lahav, M. Lysis and biodegradation of microorganisms in infectious sites may involve cooperation between leukocyte, serum factors and bacterial wall autolysins: A working hypothesis. Eur. J. Clin. Microbiol. 1983, 2, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Bierbaum, G.; Sahl, H.G. Induction of autolysis of Staphylococcus simulans 22 by Pep5 and nisin and influence of the cationic peptides on the activity of the autolytic enzymes. In Nisin and Novel Lantibiotics; Jung, G., Sahl, H.G., Eds.; ESCOM Science Publisher: Leiden, The Netherlands, 1991; pp. 386–396. [Google Scholar]
- Albrecht, T.; Raue, S.; Rosenstein, R.; Nieselt, K.; Götz, F. Phylogeny of the staphylococcal major autolysin and its use in genus and species typing. J. Bacteriol. 2012, 194, 2630–2636. [Google Scholar] [CrossRef] [PubMed]
- Schlag, M.; Biswas, R.; Krismer, B.; Kohler, T.; Zoll, S.; Yu, W.; Schwarz, H.; Peschel, A.; Götz, F. Role of staphylococcal wall teichoic acid in targeting the major autolysin Atl. Mol. Microbiol. 2010, 75, 864–873. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Sugai, M.; Komatsuzawa, H.; Nakashima, S.; Oshida, T.; Matsumoto, A.; Suginaka, H. An autolysin ring associated with cell separation of Staphylococcus aureus. J. Bacteriol. 1996, 178, 1565–1571. [Google Scholar] [PubMed]
- Zoll, S.; Schlag, M.; Shkumatov, A.V.; Rautenberg, M.; Svergun, D.I.; Götz, F.; Stehle, T. Ligand-binding properties and conformational dynamics of autolysin repeat domains in staphylococcal cell wall recognition. J. Bacteriol. 2012, 194, 3789–3802. [Google Scholar] [CrossRef] [PubMed]
- Sass, V.; Pag, U.; Tossi, A.; Bierbaum, G.; Sahl, H.G. Mode of action of human β-defensin 3 against Staphylococcus aureus and transcriptional analysis of responses to defensin challenge. Int. J. Med. Microbiol. 2008, 298, 619–633. [Google Scholar] [CrossRef] [PubMed]
- Wiedemann, I.; Breukink, E.; van Kraaij, C.; Kuipers, O.P.; Bierbaum, G.; de Kruijff, B.; Sahl, H.G. Specific binding of nisin to the peptidoglycan precursor lipid II combines pore formation and inhibition of cell wall biosynthesis for potent antibiotic activity. J. Biol. Chem. 2001, 276, 1772–1779. [Google Scholar] [CrossRef] [PubMed]
- Tran, D.; Tran, P.A.; Tang, Y.Q.; Yuan, J.; Cole, T.; Selsted, M.E. Microbicidal properties and cytocidal selectivity of rhesus macaque theta defensins. Antimicrob. Agents Chemother. 2008, 52, 944–953. [Google Scholar] [CrossRef] [PubMed]
- Wilmes, M.; Sahl, H.G. Defensin-based anti-infective strategies. Int. J. Med. Microbiol. 2014, 304, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Sahl, H.G.; Brandis, H. Production, purification and chemical properties of an antistaphylococcal agent produced by Staphylococcus epidermidis. J. Gen. Microbiol. 1981, 127, 377–384. [Google Scholar] [PubMed]
- Burianek, L.L.; Yousef, A.E. Solvent extraction of bacteriocins from liquid cultures. Lett. Appl. Microbiol. 2000, 31, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Orlov, D.S.; Nguyen, T.; Lehrer, R.I. Potassium release, a useful tool for studying antimicrobial peptides. J. Microbiol. Methods 2002, 49, 325–328. [Google Scholar] [CrossRef] [PubMed]
- MagellanTM Software V7.0, Tecan: Crailsheim, Germany, 2010.
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilmes, M.; Stockem, M.; Bierbaum, G.; Schlag, M.; Götz, F.; Tran, D.Q.; Schaal, J.B.; Ouellette, A.J.; Selsted, M.E.; Sahl, H.-G. Killing of Staphylococci by θ-Defensins Involves Membrane Impairment and Activation of Autolytic Enzymes. Antibiotics 2014, 3, 617-631. https://doi.org/10.3390/antibiotics3040617
Wilmes M, Stockem M, Bierbaum G, Schlag M, Götz F, Tran DQ, Schaal JB, Ouellette AJ, Selsted ME, Sahl H-G. Killing of Staphylococci by θ-Defensins Involves Membrane Impairment and Activation of Autolytic Enzymes. Antibiotics. 2014; 3(4):617-631. https://doi.org/10.3390/antibiotics3040617
Chicago/Turabian StyleWilmes, Miriam, Marina Stockem, Gabriele Bierbaum, Martin Schlag, Friedrich Götz, Dat Q. Tran, Justin B. Schaal, André J. Ouellette, Michael E. Selsted, and Hans-Georg Sahl. 2014. "Killing of Staphylococci by θ-Defensins Involves Membrane Impairment and Activation of Autolytic Enzymes" Antibiotics 3, no. 4: 617-631. https://doi.org/10.3390/antibiotics3040617
APA StyleWilmes, M., Stockem, M., Bierbaum, G., Schlag, M., Götz, F., Tran, D. Q., Schaal, J. B., Ouellette, A. J., Selsted, M. E., & Sahl, H.-G. (2014). Killing of Staphylococci by θ-Defensins Involves Membrane Impairment and Activation of Autolytic Enzymes. Antibiotics, 3(4), 617-631. https://doi.org/10.3390/antibiotics3040617