Abstract
Background/Objectives: Simple urinary tract infections (sUTIs) are common in women and increasingly affected by multidrug-resistant (MDR) Escherichia coli. Extended-spectrum β-lactamase (ESBL) and AmpC producers restrict oral treatment options and promote carbapenem use. This study aimed to (i) describe the etiology and antimicrobial susceptibility of sUTIs in women of reproductive age in Oman, (ii) determine the prevalence of ESBL/AmpC-producing E. coli, (iii) evaluate nitroxoline, fosfomycin, mecillinam, and temocillin against ESBL and non-ESBL E. coli, and (iv) characterize circulating clones and resistance/virulence determinants using whole-genome sequencing (WGS). Methods: In this multicentric study (September 2022–August 2023), 795 uropathogens from 762 women (15–50 years) with sUTI were collected from four Omani hospitals. Identification and susceptibility testing of E. coli (n = 489) and Klebsiella pneumoniae (n = 140) using BD Phoenix and MALDI-TOF MS was performed (CLSI 2022). Thirty ESBL-producing and 82 non-ESBL E. coli underwent phenotypic ESBL/AmpC testing and evaluation of mecillinam, temocillin, nitroxoline, and fosfomycin. WGS was performed on 26 isolates (23 ESBL, 3 wild type) and analyzed for MLST, and SNP phylogeny using ResFinder, CARD, PlasmidFinder, VirulenceFinder. Statistical significance was set at p < 0.05. Results: E. coli (62%) and K. pneumoniae (18%) were the predominant pathogens. E. coli showed high susceptibility to nitrofurantoin (~97%), carbapenems, aminoglycosides, and piperacillin–tazobactam, but reduced susceptibility to cephalosporins, fluoroquinolones, cotrimoxazole, and ampicillin. ESBL prevalence ranged from 38–51%; AmpC producers were rare (4.6%). Mecillinam, nitroxoline, and fosfomycin exhibited 100% activity against both ESBL and non-ESBL isolates; temocillin showed 89.3% activity in ESBL strains. WGS identified 15 sequence types dominated by ST-131, ST-1193, ST-73, and ST-174, with blaCTX-M-15 as the major ESBL genotype. Conclusions: sUTIs in Oman show a high burden of ESBL-producing E. coli. Nitrofurantoin, mecillinam, fosfomycin, temocillin, and nitroxoline would be effective carbapenem-sparing oral options. Continuous phenotypic and genomic surveillance are crucial to guide antimicrobial therapy and stewardship.
1. Introduction
Clinically, UTIs range from uncomplicated cystitis (simple urinary tract infections (sUTIs)), typically presenting with dysuria, urinary frequency, urgency, and suprapubic discomfort, to acute pyelonephritis, which may manifest with fever, flank pain, and systemic features. Diagnosis relies on clinical presentation supported by urinalysis and is confirmed by urine culture with antimicrobial susceptibility testing.
sUTIs are among the leading causes of bacterial infections [1]. They represent a significant global health concern, particularly among women. In fact, approximately 50% to 60% of women will experience at least one UTI in their lifetime, with nearly 10% reporting an infection annually [2]. Moreover, these infections affect about 8% of pregnancies, posing serious risks such as pyelonephritis, preterm birth, and low birth weight if left untreated [3]. Among sexually active women aged 15 to 50 years, the incidence rate of uncomplicated cystitis is estimated at approximately 2.91 per 100 person-years [4]. Additionally, the prevalence of UTIs increases with age, reaching about 20% in women aged 65 years and older [5].
Although oral agents remain effective for many sUTIs, the increasing prevalence of multidrug-resistant (MDR) uropathogens, particularly extended-spectrum β-lactamase (ESBL)-producing Escherichia coli, compromises empiric therapy and drives the use of broader-spectrum antibiotics. This challenge is especially important in pregnancy, where asymptomatic bacteriuria and symptomatic infection are associated with higher risks of progression to pyelonephritis and adverse maternal–fetal outcomes if not detected and treated promptly.
Most of these sUTIs are primarily managed in outpatient settings, where a large volume of antibiotics is dispensed, often in a largely unregulated manner. This is partly because antimicrobial stewardship efforts have traditionally focused on inpatient care [6].
Epidemiologically, E. coli remains the predominant uropathogen in Oman and globally, followed by organisms such as Klebsiella pneumoniae, Proteus mirabilis, and Enterococcus spp., with regional variation in resistance patterns. Across the Middle East and the Gulf region, E. coli remains the most common causative agent of both community- and hospital-acquired UTIs. There is growing global concern over the spread of multidrug-resistant (MDR) clones [7], with extended-spectrum β-lactamase (ESBL)–producing E. coli dominating clinical settings [8]. Of particular concern, the ESBL determinant blaCTX-M-15 is widely disseminated, often via transferable plasmids, and is frequently linked to successful extraintestinal pathogenic E. coli lineages, including ST-38, ST-405, and ST-69, which have been reported across diverse geographic settings. High-risk clones such as ST-131 have been reported across several countries in the region, including Oman, Kuwait, Qatar, and Saudi Arabia, highlighting their widespread dissemination and association with resistant infections [9,10]. In Saudi Arabia, this clone is widespread and often carries the blaCTX-M-15 genotype [9]. Other frequently reported clones are ST-38, ST-405, and ST-69, which are linked to fluoroquinolone and β-lactam resistance. Recently, the United Arab Emirates (UAE) has reported the emergence of ST-1193, a highly resistant clone in younger women [11]. The convergence of virulence and resistance in such lineages increases the risk of treatment failure, limits oral options, and amplifies the public-health burden, highlighting the need for improved diagnostic and antimicrobial stewardship strategies.
The increasing prevalence of these MDR clones presents serious therapeutic challenges, which merit the introduction of newer antimicrobials. With the paucity of newer antimicrobials, nitroxoline (NI), mecillinam (MEC), temocillin (TMO), and fosfomycin (FOS) are gaining increasing traction due to their efficacy against MDR uropathogens in UTI [12].
Nitroxoline, an oral 8-hydroxyquinoline derivative, is currently prescribed in Germany, Poland, and Croatia, while mecillinam is commonly prescribed in Scandinavia and has recently been approved in the U.S. [13]. Temocillin is widely prescribed in the UK, Belgium, and France, targets MDR Gram-negative bacteria and is stable against ESBLs and AmpC enzymes, making it a practical carbapenem-sparing option [14]. Fosfomycin with single-dose oral therapy is suitable for uncomplicated UTIs against MDR pathogens [13].
This study explored the etiology, antimicrobial susceptibility profile, and prevalence of extended-spectrum β-lactamases (ESBL), AmpC β-lactamases (AmpC), and carbapenemases in women of reproductive age with UTI. Phenotypic detection of ESBLs and AmpCs was performed to assess the feasibility of conducting these tests in low-resource centers. Carbapenem resistance was assessed using complementary phenotypic and genomic approaches, rather than implying standalone phenotypic carbapenemase testing for all isolates. Finally, representative ESBL E. coli isolates were tested against NI, FOS, MEC, and TMO and subjected to whole genomic sequencing (WGS) to assess the circulating clones, resistance, and virulence gene profiles among the circulating uropathogenic strains.
2. Results
Most patients included in the study were from Sultan Qaboos University Hospital (SQUH), comprising 309 cases (41%). This was followed by healthcare centers affiliated with Al-Masarah Hospital with 230 patients (30%), Al-Nahda Hospital with 135 patients (18%), and Khoula Hospital with 88 patients (12%).
The distribution of uropathogens mirrored the patient’s distribution across these centers. SQUH yielded the highest number of isolates (342, 43%), followed by Al-Masarah (230, 29%), Al-Nahda Hospital (135, 17%), and Khoula Hospital (88, 11%). The slightly higher number of isolates from SQUH reflects multiple bacterial isolates recovered from some patients’ midstream urine (MSU) samples.
In total, 795 uropathogens were isolated from MSU cultures of sUTI patients across the four centers. Among the 14 identified species, Escherichia coli (n = 489, 62%) and Klebsiella pneumoniae (K. pneumoniae) (n = 140, 18%) were the predominant species. E. coli was the leading uropathogen in all centers, ranging from 8% in Khoula Hospital to 25% in SQUH, while K. pneumoniae exhibited variable prevalence, reaching its highest proportion (6.8%) at SQUH.
2.1. Microbial Etiology and Antimicrobial Susceptibility Profiles
Across all four centers, as seen in Table 1, E. coli isolates demonstrated a higher susceptibility rate to nitrofurantoin, averaging at 97.6%, in contrast to K. pneumoniae isolates, which exhibited a lower susceptibility rate of 82.3%. When considering both E. coli and K. pneumoniae, their susceptibility rates to cotrimoxazole were 66.4% and 82.1%, respectively. Similarly, their susceptibility to ciprofloxacin was 49.6% for E. coli and 59.3% for K. pneumoniae.

Table 1.
Prevalence of ESBL- and AmpC β-lactamase-producing E. coli across four hospitals.
Comparatively, the susceptibility to cephalosporins was somewhat lower among E. coli isolates (cefazolin at 45.8%, cefuroxime at 53.5%, ceftriaxone at 61.24%, ceftazidime at 66.4%, and cefepime at 41.7%) when compared to K. pneumoniae isolates (cefazolin at 47.9%, cefuroxime at 73.1%, ceftriaxone at 82.3%, ceftazidime at 85.3%, and cefepime at 59.7%). Additionally, the susceptibility rate to amoxicillin-clavulanate in E. coli was 85.9%, while in K. pneumoniae, it was 66.2%.
E. coli isolates displayed higher susceptibility rates to piperacillin-tazobactam at 98.9% when compared to K. pneumoniae isolates at 85.6%. Furthermore, E. coli isolates exhibited high susceptibility rates to carbapenems (ertapenem, imipenem, and meropenem, all at 100%), aminoglycosides (amikacin at 99.2% and gentamicin at 89.8%), and ceftazidime-avibactam (99.3%). In contrast, K. pneumoniae’s susceptibility to carbapenems (ertapenem at 94.3%, imipenem at 99%, and meropenem at 98.5%), aminoglycosides (amikacin at 96.5% and gentamicin at 94.8%), and ceftazidime-avibactam (97.3%) was slightly lower than that of E. coli.
The antimicrobial susceptibility profiles of E. coli and K. pneumoniae across the four participating centers are summarized in Table 2. It’s important to note that all these percentages represent average susceptibilities from all four centers. Overall, carbapenems exhibited excellent activity, with both meropenem and imipenem demonstrating 100% susceptibility for E. coli in all centers and nearly complete activity against K. pneumoniae (95.9–100%). Amikacin also maintained high efficacy, with susceptibility ranging from 98.8% to 100% across sites. Piperacillin–tazobactam showed similarly strong performance (97.5–100%), while nitrofurantoin remained highly effective against E. coli (96.5–98.5%) but demonstrated lower activity against K. pneumoniae (73–78%).
Table 2.
Antimicrobial susceptibility profile of E. coli (n = 489) and Klebsiella pneumoniae (n = 140) among 4 centers; SQUH, Khoula, Al-Masarrah, Al-Al-Nahdha.
Moderate susceptibility was observed for amoxicillin–clavulanate, particularly at Khoula Hospital (94.2%) and Al-Al-Nahdha Hospital (91.8%), with reduced rates at SQUH (79.3%) and Al-Masarrah Hospital (78.1%). Gentamicin displayed consistent activity (87.9–92.9%) across all centers. In contrast, the cephalosporins showed variable efficacy: ceftazidime ranged from 60.3% at Khoula Hospital to 77.6% at Al-Masarrah Hospital; ceftriaxone ranged from 59.6% to 61.8%; and cefazolin exhibited the lowest activity, especially at SQUH (15.6%), compared with 56–60% elsewhere. Cefepime susceptibility was moderate (30.4–63.6%), with the lowest value reported at Al-Masarrah Hospital.
Lower susceptibility rates were recorded for ampicillin (30.2–37.9%), cotrimoxazole (63.8–71.0%), and ciprofloxacin (58.6–64.2%) across all sites. Notably, ceftazidime–avibactam, tested exclusively at SQUH, displayed outstanding susceptibility (99.3%).
The prevalence of ESBL-producing E. coli varied among the four hospitals (Table 1). The highest prevalence was observed at Al-Masarrah Hospital, where 51.4% (73/142) of isolates were identified as ESBL producers. This was followed by SQUH, where 39.5% (79/200) of isolates exhibited the ESBL phenotype, and Al-Al-Nahdha Hospital, which showed a comparable rate of 39% (34/87). Khoula Hospital reported the lowest ESBL prevalence, with 38.3% (23/60) of isolates identified as ESBL producers.
Furthermore, AmpC β-lactamase-producing isolates were not detected in SQUH, Khoula, or Al-Masarrah Hospitals; however, four isolates (4/87; 4.6%) were identified at Al-Al-Nahdha Hospital.
Differences in resistance trends were observed between ESBL-producing and non-ESBL-producing strains. ESBL producers showed slightly reduced susceptibility to nitrofurantoin (p = 0.167), cotrimoxazole (p = 0.187), cefoxitin (p = 0.128), ampicillin (p = 0.083), amoxicillin/clavulanate, gentamicin, ciprofloxacin, and piperacillin/tazobactam compared to their non-ESBL counterparts. However, due to low number of isolates, the significance of this trends cannot be concluded.
2.2. Assessment of Various β-Lactam/β-Lactamase Inhibitors for ESBL Detection via the Double-Disk Synergy Test (DDST)
ESBL production was phenotypically evaluated using the double-disk synergy test (DDST), a well-established confirmatory method for detecting clavulanate- or tazobactam-inhibitable β-lactamases. In this assay, disks containing selected extended-spectrum cephalosporins were placed at a standardized distance from disks containing β-lactam/β-lactamase inhibitor combinations (amoxicillin–clavulanate or piperacillin–tazobactam) on Mueller–Hinton agar. Enhancement or distortion of the inhibition zone of the cephalosporin disk toward the inhibitor disk was interpreted as positive synergy, indicating ESBL activity (Figure 1).

Figure 1.
Double Disc Synergy Test (DDST). Detection of ESBL production in E. coli using several cephalosporins: cefepime (FEP), Cefpodoxime (PX), Ceftriaxone (CRO), Cefixime (CFM), and Cefaclor (CEC) against two different β-lactam/β-lactamase inhibitors: Amoxicillin-clavulanate (AMC) and Piperacillin/tazobactam (TZP). Black arrows indicate some synergistic pattern in the DDST.
A total of 30 phenotypically confirmed ESBL-producing E. coli isolates were evaluated. The frequency of observable synergy varied according to the cephalosporin tested. Cefepime showed the highest rate of synergy (83%), followed by cefixime (79%) and cefaclor (75%), suggesting that these substrates are particularly sensitive for detecting inhibitor-reversible hydrolysis by ESBL enzymes in this isolate set. In contrast, ceftriaxone demonstrated moderate sensitivity (50%), whereas synergy was less frequently observed with cefuroxime (63%) and cefazolin (88%).
2.3. Antimicrobial Susceptibility to Nitroxoline, Fosfomycin, Mecillinam and Temocillin in ESBL E. coli
Table 3 and Figure 2 summarize the antimicrobial susceptibility profiles of ESBL-producing and non-ESBL-producing E. coli isolates against mecillinam, temocillin, nitroxoline, and fosfomycin. Overall, all four agents demonstrated high in vitro activity against both isolate groups, with minimal differences observed between ESBL and non-ESBL phenotypes.
Table 3.
Antimicrobial susceptibility of ESBL- and non-ESBL producing E. coli to nitrofurantoin alternatives Mecillinam, Temocillin, Nitroxoline and Fosfomycin.

Figure 2.
Susceptibility of E. coli to nitroxoline, fosfomycin, temocillin, mecillinam and netimicin. Note: Arrows indicate potentiation between fosfomycin and temocillin and antagonism (Flattening) between fosfomycin and nitroxoline.
Mecillinam, nitroxoline, and fosfomycin showed complete activity, with 100% susceptibility among both ESBL-producing and non-ESBL-producing isolates. This uniform susceptibility across resistance profiles indicates that the activity of these agents remains largely unaffected by ESBL production, supporting their robustness against common β-lactamase-mediated resistance mechanisms in E. coli. The absence of resistant or intermediate isolates further highlights their potential reliability for empirical treatment of sUTIs.
In contrast, temocillin exhibited a modest reduction in activity among ESBL-producing isolates. While all non-ESBL isolates were fully susceptible (100%), susceptibility among ESBL producers was 89.3% (27/30), with the remaining 10.7% (3/30) categorized as intermediate. Notably, no temocillin resistance was observed in either group. This pattern suggests that, although temocillin retains substantial activity against ESBL-producing E. coli, its efficacy may be partially influenced by specific resistance mechanisms or strain-related factors present within the ESBL population.
2.4. Genotypic Characterization of ESBL Producing E. coli
The genetic diversity of ESBL-producing E. coli isolates was investigated using multi-locus sequence typing (MLST), a standardized genotyping method that classifies bacterial isolates into sequence types (STs) based on the allelic profiles of seven conserved housekeeping genes. Isolates sharing the same ST are considered closely related genetically and may represent successful lineages that circulate within communities or healthcare settings.
Among the 26 ESBL-producing E. coli isolates analyzed, 15 distinct sequence types were identified (Table 4), indicating a genetically diverse population rather than dominance by a single clone. Despite this diversity, several STs were repeatedly detected, suggesting the presence of locally prevalent or internationally disseminated lineages.
Table 4.
Multi-Locus Sequence Types of E. coli (n = 26).
The most frequently identified sequence types were ST-131, ST-174, ST-1193, and ST-73, each represented by three isolates (11.5% per ST). Collectively, these four STs accounted for nearly half of all ESBL-producing isolates, highlighting their epidemiological relevance. Notably, ST-131 is a globally recognized high-risk clone commonly associated with ESBL production and multidrug resistance, while ST-1193 and ST-73 have increasingly been reported in sUTIs and are known for their pathogenic potential. The detection of ST-174, a less frequently reported lineage, suggests possible regional circulation within the study setting.
Several additional STs were identified at lower frequencies. ST-38, ST-141, and ST-127 were each detected in two isolates, whereas eight STs (ST-69, ST-7401, ST-12, ST-410, ST-998, ST-10, ST-44, and ST-69) were each represented by a single isolate, further emphasizing the heterogeneity of ESBL-producing E. coli in this cohort. The presence of both globally recognized and less common STs suggests multiple independent acquisition events rather than expansion of a single dominant clone. Importantly, a novel sequence type was identified in isolate EC 1286, indicating the emergence of a previously unreported genetic lineage.
The phylogenetic analysis revealed three distinct primary clades (Figure 3). Clade one comprised clusters featuring sequence types ST-12, ST-141, and ST-998. Clade 2 encompassed a solitary cluster with sequence types ST-127 and ST-1193. Clade 3 exhibited five principal clusters, including sequence types ST-410, ST-44, ST-10, ST-7401, ST-69, Novel-ST, and ST-38. Notably, isolates with ST-998, ST-12, and ST-141 demonstrated close relatedness, as did ST-69 and ST-7401. Lastly, isolates with ST-410, ST-44, and ST-10 also showed a discernible level of relatedness within the phylogenetic structure.
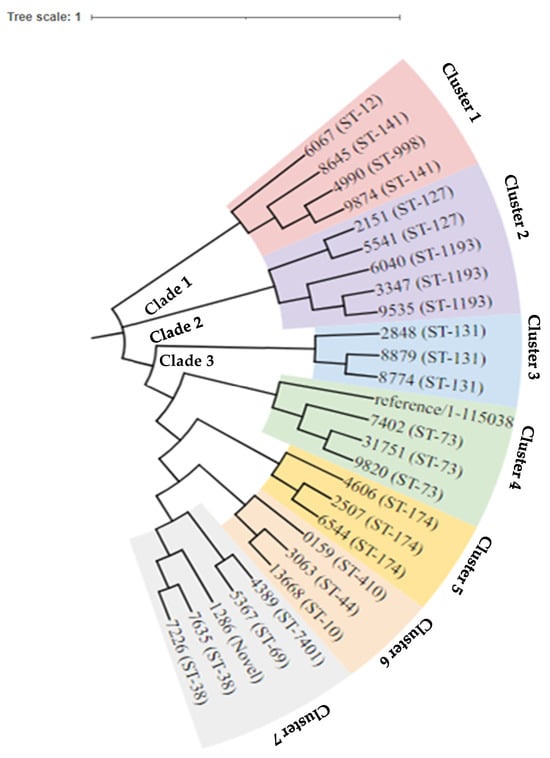
Figure 3.
Phylogeny tree of the 26 strains of E. coli with three main clades.
The distribution of key antimicrobial resistance genes across different E. coli sequence types varied. Amongst the ESBLs, blaCTX-M-15 predominated detected in 17 isolates, followed by blaCTX-M-14 and blaCTX-M-27 (4 and 3 isolates, respectively). blaTEM-1, was detected in four isolates as was blaOXA-1, was present in four isolates. The AmpC gene (DHA-1) was identified in two strains.
blaCTX-M-15, frequently co-occurred with cpxA. Many ESBL-producing strains also carried efflux pump regulators such as emrR, emrB, and mdtH, along with global regulators like marA which was present in 11 isolates. In addition, resistance determinants like aadA5 (aminoglycoside resistance) and QnrS1 (fluoroquinolone resistance) were frequently present in ESBL strains. Genes such as evgA and H-NS were found universally across all isolates while msbA and mdtN had a more limited distribution.
Table 5 summarizes the distribution of efflux resistance genes identified among 26 E. coli isolates. A total of 13 different efflux-associated genes were detected, belonging to various efflux pump families, including RND (Resistance-Nodulation-Division), ABC (ATP-Binding Cassette), MFS (Major Facilitator Superfamily), and regulatory systems.
Table 5.
Distribution of efflux resistance genes in E. coli n = 26.
Among these, the evgA and H-NS genes were universally present in all isolates, indicating their conserved role in global regulation and stress response. The marA regulator, another well-known global transcriptional activator associated with multiple antibiotic resistance, was detected in 11 isolates (42.3%), making it the most frequent regulator after evgA and H-NS.
Other efflux-related genes were detected at lower frequencies. The msbA and mdtN genes (ABC and RND families, respectively) were each identified in three isolates (11.5%), while the acrB–AcrE–mdtP cluster was found together in two isolates (7.7%), suggesting coordinated expression of multidrug efflux systems within these strains.
Several genes were detected only once, including yojI (ABC family), PmrF (LPS modification system), evgS (two-component system), acrD, AcrS (both RND family members), and emrA (MFS family). The presence of these unique genes highlights the genetic diversity and heterogeneity of efflux-mediated resistance mechanisms among the isolates studied.
Table 6 summarizes the distribution of antimicrobial resistance genes detected in 26 E. coli isolates, categorized according to their respective multi-locus sequence types (MLSTs). The results reveal a wide range of resistance determinants spanning multiple antibiotic classes, with notable variability between sequence types.
Table 6.
Distribution of resistance genes by ResFinder 4.0 tool, n = 26.
Resistance genes associated with aminoglycosides were among the most frequently observed, including aph(6)-Id, aph(3″)-Ib, aac(6′)-Ib-cr, aac(3)-IIa, and aadA variants (aadA1, aadA5), commonly present in high-risk lineages such as ST-131, ST-174, and ST-69. The coexistence of multiple aminoglycoside-modifying enzymes in several isolates suggests strong selective pressure from aminoglycoside exposure.
For quinolone resistance, mutations in gyrA and the plasmid-mediated gene qnrS1 were detected in a subset of isolates, particularly within ST-1193 and ST-131, two globally disseminated fluoroquinolone-resistant clones. These findings align with the phenotypic resistance typically associated with these sequence types.
Resistance to folate pathway antagonists was mediated by sul1, sul2, and dfrA (dfrA1, dfrA14, dfrA17) genes, with these determinants frequently co-occurring in isolates that also harbored aminoglycoside and β-lactamase genes, indicating the presence of multi-resistance plasmids.
Tetracycline resistance was conferred by tet(A) and tet(B), detected in a limited number of isolates, while macrolide resistance was attributed to mrx and mph(A), each identified sporadically.
Among β-lactamase genes, multiple classes were detected. The most prevalent ESBLs included blaCTX-M variants (blaCTX-M-14b, blaCTX-M-15, blaCTX-M-27), particularly within ST-131 and ST-1193. blaTEM-1B and blaSHV-12 were also detected, often co-carried with blaOXA-1, highlighting the multidrug-resistant potential of these clones. A single isolate harbored an AmpC (blaDHA-k) gene, whereas OXA-type β-lactamases were detected in a few isolates belonging to high-risk lineages.
Table 7 presents the distribution of virulence genes detected among 26 E. coli isolates representing 15 distinct STs. A wide range of virulence-associated factors were identified, encompassing genes involved in adhesion, iron acquisition, toxin production, capsule formation, and immune evasion, reflecting the genetic and functional diversity of the isolates.
Table 7.
Distribution of virulence genes detected in E. coli, n = 26.
Among adhesion-related genes, fimH, encoding type 1 fimbriae, was the most prevalent, detected in 13 isolates (50%), followed by focG, papC, and members of the sfa and yeh fimbrial clusters, each detected in fewer isolates. Genes linked to iron acquisition such as fyuA, irp2, sitA, and iss were widely distributed, each detected in up to 11 isolates, underscoring the importance of siderophore-mediated iron uptake in E. coli virulence.
Capsule-associated genes, including kpsE, kpsMII, and neuC, were also found in several isolates, suggesting that polysialic acid capsule synthesis may contribute to serum resistance and persistence in the urinary tract. Toxin-related genes, such as hlyA, cnf1, usp, vat, and sat, were present in a smaller subset (3–4 isolates), typically associated with ExPEC pathotypes.
Notably, high-risk global lineages such as ST-131, ST-1193, and ST-73 carried multiple virulence factors, particularly those involved in adhesion and iron acquisition, highlighting their pathogenic potential. Less common sequence types (e.g., ST-998, ST-410, ST-44) carried fewer virulence genes, indicating variability in virulence gene content across clonal lineages.
3. Discussion
Epidemiological studies estimate that approximately 50–60% of women in the reproductive age group (15–49 years) experience at least one UTI during their lifetime, and a significant proportion will have recurrent infections, due to a combination of anatomical, hormonal, behavioral, and microbiological factors. Anatomically, the shorter female urethra and its proximity to the anus facilitate ascending bacterial entry, particularly by uropathogenic E. coli. Hormonal fluctuations during the reproductive years, including menstrual cycling and pregnancy, influence the vaginal microbiota and may reduce protective Lactobacillus species, thereby increasing susceptibility to colonization. Sexual activity, contraceptive practices, hydration habits, and delayed voiding further contribute to UTI risk in this age group. Recurrent infections are additionally driven by the ability of uropathogens to adhere to uroepithelial cells, persist as intracellular bacterial communities, and re-emerge after treatment, as well as by host susceptibility and repeated antimicrobial exposure [15,16]. Among the reproductive group, pregnant women are particularly vulnerable due to physiological changes in the urinary tract, with UTIs occurring in 2–10% of all pregnancies [3]. In contrast, non-pregnant women in the same age range have a UTI prevalence ranging from 20–30%, depending on factors such as sexual activity, contraception use, and personal hygiene practices [17].
We evaluated the susceptibility profiles of E. coli and K. pneumoniae in pregnant and non-pregnant women to promote antimicrobial stewardship. In addition, we explored the interplay between AMR and virulence genes in whole-genome sequenced representative E. coli isolates. A study analyzed 1798 urine cultures and found that E. coli and K. pneumoniae were the predominant uropathogens, accounting for 60% and 33.2% of isolates, respectively. Notably, E. coli isolates exhibited high susceptibility to fosfomycin (98.6%) and nitrofurantoin (80%), whereas K. pneumoniae showed lower susceptibility to nitrofurantoin (17.2%) but remained relatively susceptible to fosfomycin (73.9%) and amikacin (50.2%) [18].
Although overall antimicrobial susceptibility did not differ significantly according to pregnancy status, wild-type E. coli isolates—defined as strains lacking acquired resistance mechanisms and exhibiting standard antibiotic susceptibility—were more frequently identified among pregnant women (31%) than non-pregnant women (24%) [18]. Similar patterns have been reported in previous studies, where lower rates of antimicrobial resistance among uropathogens isolated from pregnant women were observed compared with non-pregnant populations [19,20,21]. These authors attributed this trend to differences in healthcare exposure and antibiotic selection pressure. Pregnant women are more likely to undergo routine antenatal screening, enabling early detection and treatment of bacteriuria, often before repeated antibiotic exposure occurs [21]. In contrast, non-pregnant women may experience recurrent or self-managed UTIs, which are associated with higher cumulative antibiotic use and increased selection of resistant strains [19]. Moreover, antibiotic prescribing during pregnancy is typically more restricted and guideline-driven, which may further limit resistance development [20]. Similar observations have also been reported in studies from Europe and Asia, where ESBL-producing E. coli were less prevalent among pregnant women than among non-pregnant women [22,23], supporting the interpretation that pregnancy-related clinical practices may favor the persistence of wild-type strains.
Our study confirms that nitrofurantoin remains a reliable empirical option for the treatment of uncomplicated urinary tract infections (sUTIs), demonstrating high susceptibility rates in both pregnant (93.5%) and non-pregnant women (97.6%), along with favorable pharmacokinetic properties, including oral availability, urinary tract specificity, and minimal impact on the gut microbiota. Nitrofurantoin is active against a broad range of Gram-negative and Gram-positive uropathogens, including E. coli, supporting its role as a cephalosporin- and fluoroquinolone-sparing agent in sUTI management [24]. In non-pregnant women, nitrofurantoin is generally well tolerated, with the most commonly reported adverse effects being mild gastrointestinal symptoms such as nausea and abdominal discomfort; rare but recognized complications include pulmonary hypersensitivity reactions, hepatotoxicity, and peripheral neuropathy, particularly with prolonged or repeated use. In pregnancy, nitrofurantoin is considered safe during the first and second trimesters and is widely recommended in clinical guidelines; however, its use is generally avoided near term (after 38 weeks’ gestation) and during labor due to the risk of neonatal hemolytic anemia, especially in infants with glucose-6-phosphate dehydrogenase (G6PD) deficiency. Despite these considerations, when appropriately prescribed, nitrofurantoin offers a favorable balance between efficacy and safety, reinforcing its value as a first-line therapy for sUTI while reducing reliance on broader-spectrum antibiotics [25,26,27].
Moderate susceptibility rates (50–80%) observed for fluoroquinolones, cephalosporins, and cotrimoxazole reflect the increasing impact of well-characterized genetic resistance mechanisms identified through WGS. Reduced susceptibility to fluoroquinolones is commonly associated with chromosomal point mutations in the quinolone resistance-determining regions (QRDRs) of the gyrA and parC genes, which encode DNA gyrase and topoisomerase IV, respectively. In addition, WGS frequently detects plasmid-mediated quinolone resistance (PMQR) genes, such as qnr variants and aac(6′)-Ib-cr, which confer low-level resistance but facilitate the selection of higher-level resistance when combined with chromosomal mutations. The presence of these determinants explains the intermediate susceptibility patterns observed in this study and is consistent with global reports linking fluoroquinolone resistance to prior antibiotic exposure and horizontal gene transfer [28,29].
For cephalosporins, reduced susceptibility is primarily driven by the presence of ESBL genes, particularly blaCTX-M variants (notably blaCTX-M-15), as well as blaTEM and blaSHV. These enzymes hydrolyze third- and fourth-generation cephalosporins, leading to diminished activity despite in vitro susceptibility in some isolates. Additional mechanisms, including porin loss or alteration and increased expression of efflux pumps, can further reduce cephalosporin permeability and enhance resistance, particularly in ESBL-producing strains. Similar genetic profiles have been widely reported in uropathogenic E. coli from both community and hospital settings.
Reduced susceptibility to cotrimoxazole (trimethoprim–sulfamethoxazole) is typically associated with the acquisition of dihydrofolate reductase (dfr) genes conferring trimethoprim resistance and sulfonamide resistance genes (sul1, sul2), which encode altered dihydropteroate synthase. These genes are often carried on integrons and plasmids, facilitating their spread among uropathogens and explaining the persistence of moderate susceptibility rates despite reduced clinical use in some regions.
When compared with other studies, our findings align with global trends showing declining susceptibility to these historically first-line agents. However, these findings contrast with previous studies that described slightly higher susceptibility rates to cephalosporins and fluoroquinolones [30,31,32]. Such differences likely reflect regional variation in antibiotic prescribing practices, local resistance gene pools, and clonal population structures, as revealed by WGS. Together, these results emphasize the value of integrating genomic data with phenotypic susceptibility testing to better understand resistance mechanisms and to inform context-specific empirical treatment guidelines.
Excellent activity was observed against aminoglycosides and piperacillin-tazobactam, making them effective carbapenem-sparing alternatives against ESBL-producing uropathogens.
ESBLs present a formidable challenge in healthcare settings, imparting resistance to multiple antibiotics and thereby limiting treatment options. The differences in susceptibility profiles between ESBL and non-ESBL producers underscore the importance of being aware of local ESBL rates when prescribing antibiotics.
Carbapenems, with 100% susceptibility, underscore their potency against extended-spectrum β-lactamase (ESBL) and AmpC-producing isolates. However, their use should be restricted to prevent the acceleration of carbapenem resistance [33]. It is essential to identify carbapenem-sparing options for managing ESBL-producing uropathogens. Notably, our study also demonstrated 100% susceptibility to nitroxoline among 30 ESBL-producing E. coli isolates, consistent with findings from a larger study of 394 multidrug-resistant Enterobacterales urinary isolates, where nitroxoline showed the highest activity: 99% susceptibility among ESBL producers, 98% among AmpC, and 100% among carbapenemase-producing and ESBL + AmpC isolates.
Fosfomycin is an excellent option for sUTIs as an empirical choice, as it has a broad spectrum of activity against both Gram-negative and some Gram-positive bacteria and retains efficacy in multidrug-resistant isolates. Importantly, in our study, E. coli isolates showed 100% susceptibility to fosfomycin. Its ability to achieve high concentrations in the urinary tract enhances its effectiveness in treating UTIs. The single-dose oral formulation improves patient compliance and is a safe choice during pregnancy. Fosfomycin’s unique mechanism of action reduces the likelihood of cross-resistance with other commonly used antibiotics [31]. Prior studies have demonstrated that fosfomycin and nitrofurantoin remain appropriate agents for the empirical management of sUTI [32,33,34,35].
Mecillinam represents a valuable empiric option for sUTIs due to its high and selective activity against E. coli, including many ESBL-producing strains, and its minimal impact on the intestinal microbiota [12,13]. This limited microbiota disruption is primarily explained by the drug’s pharmacokinetic properties: after oral administration as its prodrug pivmecillinam, the active compound is rapidly absorbed and preferentially excreted into the urine, resulting in low intestinal exposure and reduced selective pressure on commensal gut bacteria. In addition, mecillinam is a narrow-spectrum β-lactam with minimal activity against anaerobes and many non-target Gram-negative organisms that dominate the gut microbiota, further limiting ecological disturbance.
The strong activity of mecillinam against E. coli is attributed to its unique mechanism of action, as it selectively binds to penicillin-binding protein 2 (PBP2), which plays a critical role in maintaining the rod shape and cell wall integrity of E. coli. This target is distinct from those of many other β-lactams, and importantly, most ESBL enzymes do not efficiently hydrolyze mecillinam, allowing the drug to retain activity against ESBL-producing isolates. Furthermore, resistance to mecillinam typically requires specific chromosomal mutations affecting PBP2 or associated pathways, which occur relatively infrequently and often impose a fitness cost on the bacterium, limiting widespread dissemination.
The oral availability of pivmecillinam enhances its practicality for outpatient therapy and it is considered safe for use during pregnancy [36]. Consistently high susceptibility rates exceeding 95% across multiple European surveillance studies [37,38], along with guideline endorsement in several countries [39,40], support its role as a narrow-spectrum, carbapenem-sparing agent. In line with these reports, all E. coli isolates in the present study demonstrated 100% susceptibility to mecillinam, reinforcing its suitability for empirical treatment of sUTIs in settings facing increasing resistance to broader-spectrum antibiotics.
Temocillin is an effective oral agent for managing ESBL UTIs [39]. Its stability against ESBL and AmpC enzymes makes it a good carbapenem-sparing agent [40]. In our study, 89.3% of E. coli isolates were susceptible to temocillin, supporting its utility in treating ESBL-related UTIs. EUCAST breakpoints recommend high-exposure dosing (2 g q8h), which achieves urinary concentrations well above clinical MICs [39,40].
The MLST analysis revealed substantial genetic diversity among E. coli isolates, with 15 distinct sequence types (STs) identified, indicating that UTIs in this population are caused by multiple unrelated lineages rather than a single dominant clone. Such diversity is commonly reported in sUTIs and reflects the broad ecological reservoir of E. coli, ongoing horizontal gene transfer, and repeated introduction of strains from different sources. Despite this heterogeneity, the predominance of ST-131, ST-1193, ST-73, and ST-174 suggests selective expansion of well-adapted lineages with enhanced capacity for colonization, persistence, and, in some cases, antimicrobial resistance. ST-131 and ST-1193 are globally disseminated high-risk clones frequently associated with ESBL production and fluoroquinolone resistance, and their dominance has been widely reported in studies from Europe, Asia, and the Middle East, supporting the global success of these lineages. ST-73, in contrast, is typically linked to extraintestinal pathogenicity rather than extensive resistance, indicating that virulence, in addition to resistance, contributes to clonal success. The presence of ST-174, which has been less frequently reported in international studies, may reflect regional circulation or local selective pressures, such as antibiotic prescribing patterns. Compared with studies reporting clonal dominance by one or two STs, the broader ST distribution observed here suggests a dynamic population structure, emphasizing the importance of local genomic surveillance to capture regional differences in circulating uropathogenic E. coli and to inform empirical treatment and public health strategies [41].
ST-131, the leading sequence type is widely recognized for its pandemic multidrug-resistant lineage, often associated with the extended-spectrum β-lactamase gene blaCTX-M-15. It is frequently isolated from urinary tract and bloodstream infections in both community and hospital settings and combines both antimicrobial resistance and virulence traits [41].
The prevalence of ST-1193, which clusters with ST-127, is increasing globally, and its presence in our study suggests it may be gaining a foothold in local populations. It is primarily associated with fluoroquinolone resistance and has increasingly been detected in clinical isolates worldwide. Emerging evidence indicates that ST-1193 is increasingly supplanting ST-131 in certain regions, with respect to both prevalence and antimicrobial resistance, particularly among uropathogenic Escherichia coli strains [42].
The identification of ST-73 in this study reflects the continued circulation of a E. coli lineage characterized by high virulence but relatively low antimicrobial resistance, which is typical of community-acquired extraintestinal pathogenic E. coli (ExPEC). The pathogenic potential of ST-73 is largely driven by the presence of key virulence genes, particularly pap, kpsMII, and fyuA. The pap gene cluster encodes P fimbriae that mediate strong adhesion to uroepithelial cells, facilitating colonization of the urinary tract and ascent to the kidneys, and is therefore closely associated with symptomatic and severe UTIs. The kpsMII gene is involved in the synthesis of a group 2 polysaccharide capsule, which enhances resistance to host immune defenses such as complement-mediated killing and phagocytosis, promoting persistence and invasiveness. The fyuA gene encodes the receptor for the yersiniabactin siderophore system, enabling efficient iron acquisition in the iron-limited urinary environment and contributing to bacterial survival and competitiveness. Together, these virulence determinants act synergistically to enhance adhesion, immune evasion, and nutrient acquisition, explaining the high pathogenic potential of ST-73 despite its generally lower resistance profile [43].
While ST-174 is not as widely reported in the literature, its identification as a predominant sequence type in this study may indicate regional clonal expansion or a niche-specific adaptation. A study [44] recently reported the presence of ST-174 in neonatal sepsis cases in China, where it was associated with virulence traits including siderophore production and capsule expression. The emergence of ST-174 in this cohort suggests that previously underreported clones may be gaining clinical significance in specific geographical areas.
While mapping the epidemiology of UPEC isolates, circulating clades, and evolving resistance, efforts to assess treatment modalities go a long way in informing appropriate empirical management [45]. It was reported [46] that Fosfomycin resistant ESBL E. coli isolates belonged to the epidemic clone ST-131. Management of ST-1193 is more difficult. A study [47] reported that ST-69 (15%) and ST-131 (8%) predominated among UPEC isolates in Australia and were associated with 57% resistance to trimethoprim-sulfamethoxazole.
Beyond efflux-mediated resistance, antibiotic inactivation emerged as the second most prevalent resistance mechanism, driven largely by the acquisition of β-lactamase-encoding genes. A total of 15 distinct acquired genes were implicated in this mechanism amongst which five ESBL genes CTX-M-14, CTX-M-15, CTX-M-27, TEM-1 and SHV-12, predominated, with CTX-M-15 being the most common (65%), reaffirming its global role as a high-risk resistance determinant and SHV-12 the less prevalent (4%) [48]. The OXA-1 gene was found in five isolates. In addition, the co-production of both AmpC and ESBL was particularly detected in nine strains. Authors [48] reported that 31% of ESBL-producing E. coli strains carried at least one of the β-lactamase genes (blaCTX-M, blaTEM, blaSHV) in Iranian outpatients with UTI. A study from Nepal [46] utilized purified DNA from E. coli isolates as a template to detect ESBL genotypes, including CTX-M, TEM, and SHV β-lactamase genes. A study conducted in India investigated the coexistence of ESBL genes with carbapenemase, AmpC, and aminoglycoside resistance genes among uropathogens, highlighting blaCTX-M-15 as the most prevalent ESBL gene. Furthermore, it was found that ESBL genes co-existed with carbapenemase genes (blaNDM-5 and blaOXA-48), AmpC genes (blaCIT and blaDHA-1), and aminoglycoside resistance genes (rmtB, rmtA, rmtC and armA) [49]. A recent study from Croatia reported that in E. coli and K. pneumoniae, the dominant resistance mechanisms are ESBLs belonging to the CTX-M, TEM, and SHV families; p-AmpC; and carbapenemases belonging to classes A, B, and D [50]. Different research groups reported the detection of OXA-1 in E. coli isolates. The resistance genes blaCTX-M-15, blaNDM-5, blaOXA-48, blaCIT, blaDHA-1, OXA-1, rmtA, rmtB, rmtC, and armA do not directly encode classical virulence factors, but they have a major indirect impact on pathogenicity, clinical outcomes, and public health by severely limiting effective treatment options and promoting bacterial persistence and spread. blaCTX-M-15 confers resistance to third-generation cephalosporins and is widely associated with successful high-risk clones, leading to frequent empiric treatment failure and increased use of broader-spectrum antibiotics. blaNDM-5 and blaOXA-48 encode carbapenemases that compromise last-line β-lactams, markedly increasing morbidity, mortality, and outbreak potential in healthcare and community settings. Plasmid-mediated AmpC genes (blaCIT, blaDHA-1) further reduce cephalosporin efficacy and complicate laboratory detection, while OXA-1 can undermine β-lactam/β-lactamase inhibitor combinations, limiting carbapenem-sparing strategies. The 16S rRNA methyltransferase genes (rmtA, rmtB, rmtC, armA) confer high-level resistance to aminoglycosides, eliminating an important option for severe and combination therapy. Collectively, these genes increase the clinical impact of infections by delaying appropriate therapy, prolonging infection and colonization, facilitating horizontal gene transfer via mobile genetic elements, and driving the emergence and dissemination of multidrug- and carbapenem-resistant strains, representing a significant threat to both individual patient outcomes and public health [51,52,53].
The marA gene—known for enhancing efflux pump activity and reducing membrane permeability—was detected in 11 isolates, making it one of the most frequently encountered efflux-associated genes [54]. Overall, efflux mechanisms emerged as the dominant resistance strategy, with nearly 20 different genes contributing to this class. The universal presence of genes like evgA and H-NS suggests a conserved role in baseline efflux regulation.
Moreover, six efflux genes—YojI, PmrF, evgS, acrD, AcrS, and emrA—were identified as unique, each occurring in only one isolate, which may reflect isolate-specific adaptations or rare resistance elements.
Plasmid-mediated resistance is a significant concern, as drug resistance genes can spread rapidly and efficiently between bacterial species through horizontal gene transfer [55]. Among the seven plasmids detected Col (BS512), IncFII (pRSB107), IncFIA, IncFII, IncFIB (AP001918), IncFIB (H89-PhagePlasmid) and IncFIB (K), Col(BS512) (31%) and IncFII (pRSB107) (27%) predominated. A recent study in Oman reported the detection of IncFIA, IncFII, IncY, IncI1-I(Alpha), and IncX in E. coli [56]. IncF, IncFII plasmids are known carriers of broad spectrum of antibiotic resistance genes in E. coli [57]. Another study reported that most of the E. coli isolates carried IncFIA, IncFII, IncFIB, Col(BS512), IncL1, IncX3, and IncH plasmids [58].
Antibiotic efflux was the most common mechanism of resistance, with nearly 20 genes being implicated. In particular, the marA gene, responsible for antibiotic efflux and reduction of cellular permeability to antibiotics, was detected in 11 E. coli isolates. The efflux genes belonged to several efflux families, like resistance-nodulation-cell division (RND), ATP-binding cassette (ABC) antibiotic efflux pumps, and major facilitator superfamily (MFS) [59,60]. Antibiotic inactivation, coded by 15 acquired genes, was predominantly mediated by β-lactamase enzymes (EC-5, ampC beta-lactamase, TEM-1 and CTX-M genes) [61]. Antibiotic target replacement was mediated by sul1, sul2, drfA14, drfA17, and PmrF.
The identification of multiple efflux pump genes and plasmid replicons in E. coli isolates has important implications for human and public health, as these mechanisms contribute to multidrug resistance, reduced treatment efficacy, and the dissemination of resistance within communities and healthcare settings. Although several efflux genes (yojI, pmrF, evgS, acrD, acrS, and emrA) were detected only once, their presence suggests isolate-specific adaptive responses to antimicrobial pressure. More importantly, the detection of marA in multiple isolates is clinically significant, as this global regulator enhances antibiotic efflux while simultaneously decreasing outer membrane permeability, resulting in broad, low-level resistance across multiple drug classes and facilitating the selection of additional resistance mechanisms. The predominance of IncF-family plasmids (IncFIA, IncFII, IncFIB) and Col(BS512) further amplifies this risk, as these plasmids are well-established vehicles for horizontal gene transfer and are frequently associated with ESBL, AmpC, and other resistance determinants, a pattern consistent with reports from Oman and other regions. The co-occurrence of efflux-mediated resistance, plasmid-borne genes, and enzymatic mechanisms such as β-lactamase production and target replacement increase the likelihood of multidrug-resistant infections, delayed effective therapy, prolonged colonization, and onward transmission, highlighting the need for integrated genomic surveillance to inform antimicrobial stewardship and public-health interventions [62].
A total of 57 virulence factors (VFs) were identified in this study, with the highest prevalence observed in the ST-73 and ST-998 lineages. This observation aligns with findings from other studies that have consistently highlighted ST-73 as a high-virulence clone associated with extraintestinal pathogenic E. coli (ExPEC). In a large-scale study on community-acquired UTIs, ST-73 was reported as the predominant clone in male patients (50%) and the second most frequent in females (12%), exhibiting the highest virulence score among tested clones, with a median and mean score of 9 [63]. The virulence of ST-73 is attributed to the presence of multiple factors, including adhesins (fimH, pap), siderophores (fyuA, irp2), and protectins such as iss, which collectively enhance colonization and immune evasion in the urinary tract.
While ST-998 has been less frequently reported in the literature, ST-73 was similarly prevalent in this analysis suggests a noteworthy virulence profile. The presence of numerous virulence genes in ST-998 indicates its potential for causing extraintestinal infections, although its full pathogenic capacity requires further investigation. Recent molecular surveillance studies have begun to detect ST-998 in clinical settings, suggesting an emerging role in UTI epidemiology and warranting closer attention [42]. The detection of ST-998 with a high VF load in this study may represent a localized clonal expansion or an underrecognized lineage with evolving clinical significance.
The ability of E. coli to cause infections relies on its arsenal of VFs, particularly those that mediate attachment to host tissues. Among the most prevalent attachment genes identified in the isolates was fimH, which encodes the adhesin component of Type 1 fimbriae [63]. This gene was universally detected in all isolates, supporting its critical role in urinary tract colonization. According to a study [64], fimH is strongly associated with adhesion to uroepithelial cells and is a key determinant in cystitis and pyelonephritis. In E. coli O159, the entire YHD fimbriae cluster—comprising yehA, yehC, and yehD—was present. These genes encode an outer membrane lipoprotein, a chaperone, and a major pilin subunit, respectively, and are implicated in biofilm formation and epithelial attachment [64]. Additionally, IpfA, encoding long polar fimbriae, was also exclusive to E. coli O159 and has been linked to intestinal colonization and interaction with Peyer’s patches [65]. Other notable fimbrial genes included papA_F43 in E. coli 31751, papA_F11 and papA_fsiA_F16 in E. coli 9535—all of which code for P fimbriae pilin subunits. These fimbriae are critical for adhesion to kidney tissues and are frequently associated with upper urinary tract infections [66,67].
Another central virulence mechanism involves iron acquisition, a vital process given the iron-limited environment of the human host. Several siderophore-associated genes were found in subsets of isolates. fyuA, encoding the yersiniabactin receptor, facilitates iron uptake and is associated with enhanced virulence and stress response. Banerjee et al. (2023) showed that fyuA is commonly present in multidrug-resistant uropathogenic E. coli (UPEC) strains. irp2, also detected in this study, is essential for yersiniabactin biosynthesis and has been linked to bloodstream infections [68]. Furthermore, chuA, which encodes a hemin receptor, allows E. coli to scavenge iron from hemoproteins and has been correlated with survival in urine [69]. The sitA gene, encoding an iron/manganese transporter, enhances intracellular survival by aiding in oxidative stress resistance, as noted by Ahmad et al. [70]. Together, these genes illustrate the diversity of siderophore systems employed by E. coli to overcome host nutritional immunity.
In terms of resistance to serum bactericidal activity, iss (increased serum survival) was found in multiple isolates. This gene enhances bacterial resistance to the complement system, contributing to the development of bloodstream infections and sepsis. It has been reported [71] that high prevalence of iss in invasive E. coli infection, including strains responsible for neonatal meningitis. Additionally, kpsMII_k23, which was uniquely present in E. coli 7402, encodes a polysialic acid capsule transport protein. This component of the Group 2 capsule facilitates immune evasion by protecting against phagocytosis and complement-mediated lysis [72]. These genes contribute to systemic virulence, particularly in extraintestinal infections.
Although classical cytotoxins such as hlyA (hemolysin) or cnf1 (cytotoxic necrotizing factor) were not detected, the isolates showed the presence of mchC and mchF in E. coli 5367, which are part of the microcin H47 operon. mchC is involved in the biosynthesis of the microcin peptide, while mchF encodes an ABC transporter that exports it. These microcins act as bacteriocins, inhibiting competing bacteria and enhancing niche colonization. Another study [73] highlighted the ecological advantage conferred by such systems during intestinal colonization, especially in mixed microbial environments.
The findings of the current study align with recently reported data on VFs in E. coli for UTIs. It was reported that several VFs such as P fimbriae (pap), type1 fimbriae, afimbrial adhesin I (afaI), hemolysin (hly), cytotoxic necrotizing factor 1 (cnf 1), aerobactin (aer), S fimbriae (sfa), adhesins, fimbriae, kpsMT, ompT, usp, iroN, iha, set 1, astA, group II capsule synthesis; sfa/foc, S and F1C fimbriae; iutA, traT, serum resistance; and fimH were identified in E. coli isolates causing UTIs [65,66]. In addition, a study [33] assessed the genetic relation and screening of VFs among carbapenemase- producing E. coli from UTI and found that the predominant virulence genes included iutA (97.66%), fyuA (85.33%), inh (83%), traT (82.33%), papП (96, 32%), fimH (93, 31%) and csgA (30.66%). Another study reported that the papG class II gene plays a critical role in the development of E. coli for UTIs, and fimH adhesion plays a role by acting synergistically with P papG class II adhesion [67].
4. Materials and Methods
4.1. Study Period
This multicentric study was conducted from September 2022 to August 2023 in the Department of Microbiology and Immunology, College of Medicine and Health Sciences, Sultan Qaboos University (SQU), Muscat, Oman, in collaboration with the Diagnostic Microbiology Laboratory at Sultan Qaboos University Hospital (SQUH). Clinical specimens were sourced from four centers—SQUH and the referring Khoula, Al Al-Nahdha, and Al-Masarrah Hospitals—with selected isolates processed and characterized at the SQUH Microbiology Diagnostic Laboratory for in-depth analysis. In total, 762 patients were included across these four centers. Ethical approval was granted by the Medical Research Ethics Committee (MREC) of the College of Medicine and Health Sciences, SQU (SQU-EC/377/2021).
The study assessed the demographic characteristics and antimicrobial susceptibility profiles of E. coli and K. pneumoniae isolated from adult females in the reproductive age group (15–50 years).
Eligible participants were women of reproductive age who met predefined clinical and microbiological criteria to ensure the study population represented sUTIs.
Inclusion criteria comprised two groups. First, non-pregnant women presenting with symptoms suggestive of acute uncomplicated UTI, including dysuria, urinary urgency, and increased frequency, and with no known structural, functional, or metabolic abnormalities of the urinary tract. Second, pregnant women, both symptomatic and asymptomatic, who were diagnosed with UTI during routine antenatal care, including cases of asymptomatic bacteriuria detected through screening, were included to capture the full spectrum of UTI presentations during pregnancy.
Exclusion criteria were applied to minimize confounding factors that could influence antimicrobial susceptibility and clinical outcomes. Women were excluded if they had complicated UTIs, defined as infections associated with anatomical abnormalities, urinary obstruction, indwelling catheters, renal disease, or immunocompromising conditions. Patients who had received systemic antibiotic therapy within the preceding four weeks were excluded to avoid transient suppression of bacterial growth or selection bias toward resistant strains. In addition, women with significant comorbidities (such as diabetes mellitus, chronic kidney disease, or immunosuppressive disorders) and post-menopausal women were excluded, as these conditions are associated with altered UTI pathophysiology and resistance patterns distinct from those seen in women of reproductive age.
4.2. Bacterial Identification and Susceptibility Testing
All bacterial samples included in this study were collected, transported and processed as per standard guidelines for diagnostic purposes. None of the samples included in this study was collected exclusively for the purpose of the study. Only samples meeting the inclusion criteria were recruited for the analysis. All samples were sub-cultured in CLED agar (Oxoid, Basingstoke Hampshire, UK) including the samples transported from other hospitals, prior to preservation in sterile CryoBeads (Mast Diagnostics, Derby, UK) at −80 °C for further analysis. Bacterial identification was conducted using the Phoenix™ automated system (BD Diagnostics, Sparks Glencoe, MD, USA) and MALDI-TOF MS (Bruker, Bremen, Germany) at SQUH, in accordance with standard guidelines [15]. Antimicrobial susceptibility testing was performed and interpreted according to CLSI M100: Performance Standards for Antimicrobial Susceptibility Testing, 32nd ed. (2022) [74]. The automated system identified ESBL producers, while the presence of AmpC was initially estimated from the susceptibility profiles.
4.3. Phenotypic Detection of ESBL and AmpC
Thirty representative E. coli isolates previously validated as producers of extended-spectrum beta-lactamase (ESBL) through the BD Phoenix automated systems, were subjected to phenotypic detection of ESBL and AmpC. From the total cohort of 489 E. coli isolates, subsets were selected using a purposeful stratified approach to ensure representativeness of the overall population. For expanded antimicrobial susceptibility testing, 30 ESBL-producing and 82 non-ESBL isolates were chosen to capture diversity in resistance phenotypes, clinical sources, and collection periods, while minimizing over-representation of clonally similar strains.
ESBL Detection: Double-disk synergy test was used for detection of ESBLs. Two beta-lactam/beta-lactamase inhibitors (BL/BLI), piperacillin/tazobactam (TZP,110 µg) and amoxicillin/clavulanic Acid (AMC, 30 µg) (Oxoid, Basingstoke, UK), were utilized to detect ESBLs. The BL/BLIs were placed at the center of the Mueller Hinton agar plates and cefepime (FEP, 30 µg) (Oxoid, Basingstoke, UK), cefixime (CFM, 5 µg) (Oxoid, Basingstoke, UK), cefuroxime (CXM, 30 µg) (Oxoid, Basingstoke, UK), cefazolin (KZ, 30 µg) (Oxoid, Basingstoke, UK), cefaclor (CEC, 30 µg) (Oxoid, Basingstoke, UK), and ceftriaxone (CRO, 30 µg) (Oxoid, Basingstoke, UK) were placed at a distance of 20 mm edge to edge from them. The plates were incubated at 35 °C ± 2 °C for 18–24 h and examined for the potentiation of zones of inhibition between the BL/BLIs and the cephalosporins. An isolate is classified as an ESBL producer when the inhibition zone around the combination disc exceeds that of the cephalosporin-only disc by ≥5 mm (Figure 1).
AmpC Detection: Disk approximation test was employed to detect AmpC beta-lactamases. Imipenem (IPM, 10 µg), cefoxitin (FOX, 30 µg), amoxicillin-clavulanate (AMC, 30 µg), and piperacillin-tazobactam (TZP, 110 µg) were used as inducers and ceftazidime (CAZ, 30 µg) as a substrate in this test. The center-to-center distance between antibiotic disks was kept at 25 mm. The plate was incubated at 35 °C ± 2 °C for 18–24 h. The flattening or blunting of the zone of inhibition between the inducers and ceftazidime was evaluated the next day. A flattened or indented zone of inhibition (a “D” shape) around the cephalosporin disk, specifically on the side facing the inducer disk is considered positive for AmpC. This indentation shows the inducer (Imipenem and Cefoxitin) triggers the production of AmpC, which then diffuses out and inactivates the nearby cephalosporin, reducing its inhibition zone (Figure 1).
4.4. Antimicrobial Susceptibility Testing of Fosfomycin, Nitroxoline, Mecillinam, Temocillin
Representative 30 isolates of ESBL-producing E. coli in addition to 82 non-ESBL producing E. coli were selected for further investigation. The analysis of 30 representative ESBL-producing E. coli isolates was considered sufficient to address the study objectives while balancing feasibility and resource-intensive downstream analyses, such as molecular characterization. Similar sample sizes have been widely reported in ESBL epidemiological and genomic studies. These selected isolates were tested against temocillin (TMO, 30 µg) (Liofilchem, Roseto degli Abruzzi, Italy), fosfomycin (FOS, 200 µg) (LIOFILCHEM, Italy), nitroxoline (NI, 30 µg) (LIOFILCHEM, Italy), and mecillinam (MEC, 10 µg) (LIOFILCHEM, Italy) by the Kirby-Bauer disk diffusion method as follows. Three to five bacterial colonies from overnight pure cultures from CLED agar were suspended in normal saline (Fisher Chemical, Cramlington, UK) and adjusted to a 0.5 McFarland standard (approximately 1–2 × 108 CFU/mL) using a CrystalSpec nephelometer (BD Diagnostics, Sparks Glencoe, MD, USA), according to the manufacturer’s recommendations. After 15 min, the suspension was spread onto a Mueller–Hinton agar (MHA) surface (Oxoid, Hampshire, UK) and left for 1–2 min at room temperature to be absorbed. After 15 min, the selected antibiotic disks were placed on the inoculated MHA plates using sterile forceps. The plates were incubated at 37 °C for 18–24 h. Interpretation was based on the guidelines of the CLSI (2022) and the European Committee on Antimicrobial Susceptibility Testing for temocillin (Figure 1).
4.5. Genomic DNA Extraction and Purification and Whole Genome Sequencing (WGS)
Genomic DNA was extracted from 26 E. coli isolates (twenty-three ESBL producers and three wild-type strains). For whole-genome sequencing, 26 isolates (23 ESBL-producing and 3 non-ESBL) were selected based on phenotypic heterogeneity, resistance profiles, and temporal distribution to reflect the genetic diversity of ESBL-producing within the cohort. Isolates were drawn from multiple settings, reducing sampling bias and supporting generalizability of the genomic findings.
The DNA was extracted using Qiagen kit (QIAamp® genomic DNA kit, Hilden, Germany) as described in the manufacturer’s instructions with slight modifications. One to four colonies were suspended in 10 mL Mueller–Hinton broth (Oxoid, Hampshire, UK) and left overnight on a shaking incubator set to 250 rpm at 37 °C. Then, the bacterial suspension was centrifuged for 15 min at 4000× g. The samples were then incubated again at 37 °C at a 400-rpm shaking incubator for 30–60 min (Innova 4000, New Brunswick Scientific, Hertfordshire, UK). A volume of 100 μL of TE buffer was added to the sample to obtain a final volume of 200 μL in 1.5 mL microcentrifuge tube. Then, 200 μL of the sample was mixed with 400 μL of lysis solution and incubated at 65 °C for 15 min using a heat block (Eppendorf ThermoStat plus, Hamburg, Germany). As per the manufacturer’s instruction, the samples were washed twice using the columns. The DNA pellet was eluted in a final volume of 50–100 μL of nuclease-free H2O (QIAamp® genomic DNA kit, Hilden, Germany). The extracted DNA was aliquoted into 2 vials, which were stored at 4 °C and −80 °C for future use. The DNA quality was evaluated using a 1000 NanoDrop UV spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA), with an absorbance ratio of 260/280 nm between 1.8 and 2.0, which indicated pure DNA. Gel electrophoresis was performed to ensure there were no RNA or protein contaminants. A GeneRuler 1 kb DNA ladder was used to estimate the DNA fragment sizes.
The extracted DNA was submitted to microbesNG (Birmingham, UK) for whole-genome sequencing (WGS) using Illumina technology. The following steps were performed at the sequencing facility: DNA libraries were prepared with the Nextera XT Library Prep Kit (Illumina, San Diego, CA, USA) following the manufacturer’s instructions, with two modifications: the input DNA was doubled, and the PCR elongation step was extended to 45 s. DNA quantification and library preparation were performed using a Hamilton Microlab STAR automated liquid-handling platform (Hamilton Bonaduz AG, Bonaduz, Switzerland). Pooled libraries were quantified using the Kapa Biosystems Library Quantification Kit (Kapa Biosystems, Wilmington, MA, USA) for Illumina and sequenced on Illumina HiSeq/NovaSeq instruments with a 250 bp paired-end protocol.
Adaptor and quality trimming of raw reads was performed using Trimmomatic v0.30 with a sliding-window threshold of Q15 [75]. Genome assembly was carried out using SPAdes v3.7 [76], and annotation was performed with Prokka v1.11 [77]. The assembled genomes were delivered as contigs, with a minimum coverage depth of 30× to ensure high-quality reads suitable for downstream analyses. The final assemblies and annotations were uploaded for further bioinformatic processing. The sequences were subsequently registered in the MicrobesNG database (https://microbesng.com/ accessed on 16 July 2023). Genome sequences for all E. coli and Klebsiella pneumoniae isolates were deposited in the GenBank database under accession numbers SAMN47530948–SAMN47530980.
Bioinformatic analyses were performed using tools available through the Center for Genomic Epidemiology (CGE), the Comprehensive Antibiotic Resistance Database (CARD), and the National Center for Biotechnology Information (NCBI). Multi-locus sequence typing (MLST) was conducted via the CGE MLST server (https://www.genomicepidemiology.org, accessed on 4 November 2023) to determine allelic profiles and sequence types (STs) based on six or seven conserved housekeeping genes [78,79,80,81,82]. Whole-genome sequencing (WGS) data were further examined using ResFinder 4.0 and CARD (https://card.mcmaster.ca, accessed on 15 November 2023) to identify antimicrobial resistance genes and associated mechanisms [83,84]. Plasmid profiles of the 26 isolates were characterized using PlasmidFinder 2.1 [85], while virulence-associated genes were identified through VirulenceFinder [86,87,88].
A heat map illustrating the distribution of resistance genes and antimicrobial classes among E. coli isolates was generated using Microsoft Excel 2016. Phylogenetic relationships were inferred using the CGE SNP-based phylogeny tool [89,90], and the resulting alignments and tree were visualized in MEGA7 [90,91]. Final annotation and customization of the phylogenetic tree were performed using iTOL (https://itol.embl.de, accessed on 18 November 2023), following the approach described by Letunic and Bork (2007) [92].
4.6. Statistical Analysis
For descriptive purposes, categorical variables were presented as numbers and percentages, and continuous variables with mean ± standard deviation, unless otherwise indicated. For inference purposes, categorical variables were compared using the χ2 analysis. Antibiotic resistance rates were calculated as percentages. The Fisher test was used to calculate the differences between the isolates’ susceptibility profiles, with a p-value of <0.05 considered statistically significant. Sensitivity and specificity analyses of diagnostic tests were performed, and results were visualized using Microsoft Excel. All statistical analyses were performed using IBM Statistical Package for the Social Sciences (SPSS, version 27) software.
5. Conclusions
Our findings demonstrate that nitrofurantoin, mecillinam, fosfomycin, temocillin, and nitroxoline retain high in vitro activity against ESBL-producing E. coli causing uncomplicated cystitis, thereby meeting the primary objective of identifying effective oral therapeutic alternatives to broad-spectrum agents. From a human health perspective, the use of these agents offers a clinically meaningful strategy to ensure timely and effective treatment of common urinary tract infections while minimizing exposure to cephalosporins and fluoroquinolones, whose overuse is strongly associated with the acceleration of antimicrobial resistance. At the national level, these findings are particularly relevant for Oman, where ESBL-producing Enterobacterales are increasingly reported in both community and healthcare settings. Incorporating these agents into local treatment guidelines and institutional formularies would support evidence-based empirical therapy for simple urinary tract infections, reduce treatment failure, and lower the selective pressure that drives resistance to critically important antimicrobials. Such an approach directly strengthens antimicrobial stewardship programs and contributes to improved patient outcomes, reduced recurrence rates, and preservation of microbiome integrity. Regionally, the results align with resistance trends observed across Asia, where high rates of ESBL production limit oral treatment options for community-acquired infections. The demonstrated efficacy of these narrow-spectrum agents provides a scalable and practical framework for stewardship-driven empirical therapy in similar epidemiological contexts, particularly in resource-constrained settings where access to advanced diagnostics may be limited. Globally, the study supports World Health Organization (WHO) and international antimicrobial stewardship recommendations that advocate for the prioritization of Access-group antibiotics and the de-escalation from Watch and Reserve agents whenever clinically appropriate. By promoting the rational use of effective, lower-tier antimicrobials for uncomplicated infections, these findings contribute to global efforts to curb the emergence and dissemination of multidrug-resistant organisms and to safeguard the long-term effectiveness of last-line therapies. Overall, this study provides clinically actionable evidence that supports sustainable antimicrobial use, reinforces stewardship principles, and has direct implications for public health policy and clinical practice at local, regional, and global levels.
Author Contributions
Conceptualization, N.A.-K. and N.A.-T.; methodology, A.A.-M.; software, A.A.-M.; validation, M.R. and Z.A.-J.; formal analysis, A.A.-M.; investigation, A.A.-M.; resources, N.A.-K.; N.A.-T. and Z.A.-J.; data curation, A.A.-M.; N.A.-K. and N.A.-T.; writing—original draft preparation, A.A.-M.; writing—review and editing, M.R. and Z.A.-J.; visualization, A.A.-M.; supervision, M.R. and Z.A.-J.; project administration, Z.A.-J.; funding acquisition, Z.A.-J. All authors have read and agreed to the published version of the manuscript.
Funding
Funding was provided by a Sultan Qaboos University internal grant (project code: (IG/MED/MICR/24/01).
Institutional Review Board Statement
This study commenced after obtaining approval from the Medical Ethics Research Committee (MREC# EC/377/23 February 2021), College of Medicine and Health Sciences, Sultan Qaboos University, Muscat, Oman and the Health Studies and Research Approval Committee, Ministry of Health (MoH/CSR/21/24496).
Informed Consent Statement
No consent was obtained.
Data Availability Statement
All supporting data can be found in the manuscript. All whole-genome sequencing data are deposited in DDBJ/ENA/GenBank under the accessions SAMN47530948–SAMN47530980. The version described in this paper is version SAMN47530948–SAMN47530980. The raw data supporting the conclusions of this article will be made available by the authors without undue reservation.
Acknowledgments
The authors would like to sincerely thank Nada Al-Siyabi and her team at Al Al-Nahdha Hospital for their valuable support and collaboration. We are grateful for their assistance in facilitating access to clinical data, ensuring data accuracy, and supporting institutional and ethical requirements. Their contribution was essential to the successful completion of this study. We would like to express our sincere thanks to the sequencing company provided by MicrobesNG (Birmingham, UK) (https://microbesng.com, accessed on 16 July 2023) for performing WGS and bioinformatics analysis.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ABC | ATP-binding cassette |
| AmpC | Ambler class C cephalosporinases |
| AMR | Antimicrobial Resistance |
| AMS | Antimicrobial stewardship |
| ATCC | American Type Culture Collection |
| AUC | Acute uncomplicated cystitis |
| BL | Beta-lactamase |
| BLI | Beta-lactamase inhibitor |
| CARD | Comprehensive Antibiotic Resistance Database |
| caUTI | Community-acquired urinary tract infection |
| CEC | Cefaclor |
| CFM | Cefixime |
| CGE | Genomic Epidemiology |
| CLSI | Clinical and Laboratory Standards Institute |
| CRO | Ceftriaxone |
| CXM | Cefuroxime |
| DDST | Double-Disk Synergy Test |
| E. coli | Escherichia coli |
| EB | Elution buffer |
| EDs | Emergency departments |
| ESBL | Extended-spectrum beta-lactamases |
| FAMCO | Family Medicine and Public Health |
| FOS | Fosfomycin |
| FQ | Fluoroquinolones |
| GBS | Group B Streptococcus |
| GC | Guanine-cytosine |
| ICU | Intensive care unit |
| KAP | Knowledge, Awareness and Practice |
| KPC | Klebsiella pneumoniae carbapenemase |
| KZ | Cefazolin |
| LB | Luria Broth |
| MDR | Multidrug resistant |
| MDRO | Multidrug resistant organisms |
| MEC | Mecillinam |
| MIC | Minimum inhibitory concentration |
| MLST | Multilocus sequence typing |
| MOH | Ministry of health |
| MPH | Macrolide phosphotransferase |
| MREC | Medical Research Ethics Committee |
| MRN | Medical records number |
| MRSA | Methicillin-resistant Staphylococcus aureus |
| MSU | Midstream Specimen of Urine |
| NCBI | National Center for Biotechnology Information |
| NFT | Nitrofurantoin |
| NI | Nitroxoline |
| NS | Non-susceptible |
| OPD | Outpatient departments |
| PDR | Pan-drug resistance |
| PRO | patient-reported outcome |
| RND | resistance-nodulation-cell division |
| SNPs | Single Nucleotide Polymorphisms |
| SQUH | Sultan Qaboos University Hospital |
| TMO | Temocillin |
| TMP-SMX | Trimethoprim-sulfamethoxazole |
| TPB | Theory Planned Behavior |
| UPEC | Uropathogenic E. coli |
| UTI | Urinary tract infection |
| WGS | Whole Genome Sequencing |
| WHO | World Health Organization |
| XDR | Extensive drug resistance |
References
- Mancuso, G.; Midiri, A.; Gerace, E.; Marra, M.; Zummo, S.; Biondo, C. Urinary Tract Infections: The Current Scenario and Future Prospects. Pathogens 2023, 12, 623. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Medina, M.; Castillo-Pino, E. An introduction to the epidemiology and burden of urinary tract infections. Ther. Adv. Urol. 2019, 11, 1756287219832172. [Google Scholar] [CrossRef] [PubMed]
- American College of Obstetricians and Gynecologists (ACOG). Urinary Tract Infections in Pregnant Individuals. 2023. Available online: https://www.acog.org/clinical/clinical-guidance/clinical-consensus/articles/2023/08/urinary-tract-infections-in-pregnant-individuals (accessed on 21 September 2024).
- Chu, C.M.; Lowder, J.L. Diagnosis and treatment of urinary tract infections across age groups. Am. J. Obstet. Gynecol. 2018, 219, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.C.; Lee, M. AntibiogramDSM: A combined local antibiogram and educational intervention. Antimicrob. Steward. Healthc. Epidemiol. 2023, 3, e179. [Google Scholar] [CrossRef]
- Tanwar, J.; Das, S.; Fatima, Z.; Hameed, S. Multidrug resistance: An emerging crisis. Interdiscip. Perspect. Infect. Dis. 2014, 2014, 541340. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Husna, A.; Rahman, M.M.; Badruzzaman, A.T.M.; Sikder, M.H.; Islam, M.R.; Rahman, M.T.; Alam, J.; Ashour, H.M. Extended-Spectrum β-Lactamases (ESBL): Challenges and Opportunities. Biomedicines 2023, 11, 2937. [Google Scholar] [CrossRef] [PubMed Central]
- Alghoribi, M.F.; Gibreel, T.M.; Farnham, G.; Al Johani, S.M.; Balkhy, H.H.; Upton, M. Antibiotic-resistant ST38, ST131 and ST405 strains are the leading uropathogenic Escherichia coli clones in Riyadh, Saudi Arabia. J. Antimicrob. Chemother. 2015, 70, 2757–2762. [Google Scholar] [CrossRef]
- Al Shuhoumi, M.A.; Ghafri, S.A.; Hameed, F.J.; Hinai, A.A.; Ghafri, A.A.; Mony, S.R.; Sawafi, B.A.; Govindaraj, G.; Yaqoobi, A.A.; Alawi, B.A.; et al. Burden of Multidrug-Resistant Organisms in Oman: A Six-Year Single-Study Calling for Urgent Actions. Microbiol. Res. 2025, 16, 45. [Google Scholar] [CrossRef]
- Basak, S.; Singh, P.; Rajurkar, M. Multidrug resistant and extensively drug resistant bacteria: A study. J. Pathog. 2016, 2016, 4065603. [Google Scholar] [CrossRef]
- Guggenbichler, J.P.; Assadian, O.; Kramer, A. Nitroxoline: Treatment and prevention of urinary tract infections from the urologist’s perspective. Ther. Adv. Urol. 2024, 16, 1756287223122426. [Google Scholar] [CrossRef]
- Lodise, T.P.; Kaye, K.S.; Henriksen, A.S.; Kahlmeter, G. Review of the In Vitro Microbiological Activity of Mecillinam Against Common Uropathogens in Uncomplicated Urinary Tract Infection: Focus on Resistant Pathogens. Open Forum Infect. Dis. 2024, 11, ofae296. [Google Scholar] [CrossRef]
- Lupia, T.; De Benedetto, I.; Stroffolini, G.; Di Bella, S.; Pinna, S.M.; Zerbato, V.; Rizzello, B.; Bosio, R.; Shbaklo, N.; Corcione, S.; et al. Temocillin: Applications in Antimicrobial Stewardship as a Potential Carbapenem-Sparing Antibiotic. Antibiotics 2022, 11, 493. [Google Scholar] [CrossRef] [PubMed]
- Kresken, M.; Pfeifer, Y.; Wagenlehner, F.; Werner, G.; Wohlfarth, E. on behalf of Study Group ‘Antimicrobial Resistance‘ of the Paul Ehrlich Society for Infection Therapy Resistance to Mecillinam and Nine Other Antibiotics for Oral Use in Escherichia coli Isolated from Urine Specimens of Primary Care Patients in Germany, 2019/20. Antibiotics 2022, 11, 751. [Google Scholar] [CrossRef]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Alshyarba, M.H.M.; Alamri, A.; Assiri, A.A. Economic impacts of the Bacillus Calmette-Guérin (BCG) therapy shortage and the proposed solutions for patients with non-muscle invasive bladder Cancer in Aseer Province, Saudi Arabia. J. Fam. Med. Prim. Care 2020, 9, 2758–2762. [Google Scholar] [CrossRef]
- Foxman, B. Urinary Tract Infection Syndromes: Occurrence, recurrence, bacteriology, risk factors, and disease burden. Infect. Dis. Clin. North Am. 2014, 28, 1–13. [Google Scholar] [CrossRef]
- Vimala, P.B.; Vajravelu, L.K.; Thulukanam, J.; Lathakumari, R.H.; Panneerselvam, V.P.; Nair, D.M. Clinical presentation and antibiotic resistance trends of Escherichia coli isolated from clinical samples in South India: A two-year study (2022–2023). Infect. Dis. Health 2025, 30, 183–193. [Google Scholar] [CrossRef]
- Kahlmeter, G. An international survey of the antimicrobial susceptibility of pathogens from uncomplicated urinary tract infections: The ECO·SENS Project. J. Antimicrob. Chemother. 2003, 51, 69–76. [Google Scholar] [CrossRef]
- Schneeberger, C.; Kazemier, B.M.; Geerlings, S.E. Asymptomatic bacteriuria and urinary tract infections in special patient groups: Women with diabetes mellitus and pregnant women. Curr. Opin. Infect. Dis. 2014, 27, 108–114. [Google Scholar] [CrossRef]
- Linhares, I.; Raposo, T.; Rodrigues, A.; Almeida, A. Frequency and antimicrobial resistance patterns of bacteria implicated in community urinary tract infections: A ten-year surveillance study (2000–2009). BMC Infect. Dis. 2013, 13, 19. [Google Scholar] [CrossRef]
- Abate, D.; Marami, D.; Letta, S. Prevalence, Antimicrobial Susceptibility Pattern, and Associated Factors of Urinary Tract Infections among Pregnant and Nonpregnant Women at Public Health Facilities, Harar, Eastern Ethiopia: A Comparative Cross-Sectional Study. Can. J. Infect. Dis. Med Microbiol. 2020, 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Clarke, K.; Hall, C.L.; Wiley, Z.; Tejedor, S.C.; Kim, J.S.; Reif, L.; Witt, L.; Jacob, J.T. Catheter-Associated Urinary Tract Infections in Adults: Diagnosis, Treatment, and Prevention. J. Hosp. Med. 2020, 15, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Dash, M.; Padhi, S.; Mohanty, I.; Panda, P.; Parida, B. Antimicrobial resistance in pathogens causing urinary tract infections in a rural community of Odisha, India. J. Fam. Community Med. 2013, 20, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Fourie, J.L.; Claassen, F.M.; Myburgh, J.J. Causative pathogens and antibiotic resistance in community-acquired urinary tract infections in central South Africa. S. Afr. Med. J. 2021, 111, 124–128. [Google Scholar] [CrossRef]
- Thompson, D.; Xu, J.; Ischia, J.; Bolton, D. Fluoroquinolone resistance in urinary tract infections: Epidemiology, mechanisms of action and management strategies. BJUI Compass 2023, 5, 5–11. [Google Scholar] [CrossRef]
- Oumer, Y.; Dadi, B.R.; Seid, M.; Biresaw, G.; Manilal, A. Catheter-Associated Urinary Tract Infection: Incidence, Associated Factors and Drug Resistance Patterns of Bacterial Isolates in Southern Ethiopia. Infect. Drug Resist. 2021, ume 14, 2883–2894. [Google Scholar] [CrossRef]
- van Duin, D.; Bonomo, R.A. Ceftazidime/avibactam and meropenem/vaborbactam: Emerging treatment options for multidrug-resistant Gram-negative infections. Clin. Infect. Dis. 2016, 63, 234–241. [Google Scholar] [CrossRef]
- Arooj, I.; Sehar, A.; Javaid, A. Prevalence and Antibiotic Sensitivity pattern of Bacterial Isolates from Catheterized UTI Patients: A Local Study. Pak. J. Biochem. Biotechnol. 2021, 2, 229–235. [Google Scholar] [CrossRef]
- Gupta, K.; Hooton, T.M.; Naber, K.G.; Wullt, B.; Colgan, R.; Miller, L.G.; Moran, G.J.; Nicolle, L.E.; Raz, R.; Schaeffer, A.J.; et al. International Clinical Practice Guidelines for the Treatment of Acute Uncomplicated Cystitis and Pyelonephritis in Women: A 2010 Update by the Infectious Diseases Society of America and the European Society for Microbiology and Infectious Diseases. Clin. Infect. Dis. 2011, 52, e103–e120. [Google Scholar] [CrossRef]
- Rando, E.; Giovannenze, F.; Murri, R.; Sacco, E. A review of recent advances in the treatment of adults with complicated urinary tract infection. Expert Rev. Clin. Pharmacol. 2022, 15, 1053–1066. [Google Scholar] [CrossRef]
- Salm, J.; Salm, F.; Arendarski, P.; Kramer, T.S. High antimicrobial resistance in urinary tract infections in male outpatients in routine laboratory data, Germany, 2015 to 2020. Eurosurveillance 2022, 27, 2101012. [Google Scholar] [CrossRef]
- Zykov, I.N. Old Antibiotics as Alternative Treatment Options for Urinary Tract Infections Caused by ESBL-, AmpC-and Carbapenemase-Producing Escherichia coli. Ph.D. Thesis, UiT Norges Arktiske Universitet, Tromsø, Norway, 2020. [Google Scholar]
- Rech, M.A.; Faine, B.; Vakkalanka, P.; A Talan, D. 1426. Empiric Antimicrobial Prescribing for Urinary Tract Infections in Patients Discharged from the Emergency Department. Open Forum Infect. Dis. 2021, 8, S796. [Google Scholar] [CrossRef]
- Kresken, M.; Körber-Irrgang, B.; Pfeifer, Y.; Wichelhaus, T.A.; Leistner, R.; Pfennigwerth, N. Antimicrobial susceptibility of Escherichia coli from uncomplicated urinary tract infections in women across Europe: Results from the ECO.SENS III study. J. Glob. Antimicrob. Resist. 2021, 24, 190–195. [Google Scholar] [CrossRef]
- Frimodt-Møller, N.; Simonsen, G.S.; Larsen, A.R.; Kahlmeter, G. Pivmecillinam, the paradigm of an antibiotic with low resistance rates in Escherichia coli urine isolates despite high consumption. J. Antimicrob. Chemother. 2022, 78, 289–295. [Google Scholar] [CrossRef] [PubMed]
- NICE. Urinary Tract Infection (Lower): Antimicrobial Prescribing [NG109]; National Institute for Health and Care Excellence: London, UK; Available online: https://www.nice.org.uk/guidance/ng109 (accessed on 14 November 2025).
- Bader, M.S.; Loeb, M.; Leto, D.; Brooks, A.A. Treatment of urinary tract infections in the era of antimicrobial resistance and new antimicrobial agents. Postgrad. Med. 2020, 132, 234–250. [Google Scholar] [CrossRef]
- Plambeck, L.; Fuchs, F.; Sattler, J.; Hamprecht, A. In vitro activity of mecillinam, temocillin and nitroxoline against MDR Enterobacterales. JAC Antimicrob. Resist. 2022, 4, dlac059. [Google Scholar] [CrossRef]
- Sirwan, K.A.; Safin, H.; Karzan, Q.; Radhwan, H.I.; Abdulmalik, F.; Kochr, A.M.; Mona, G.M. Antimicrobial resistance: Impacts, challenges, and future prospects. J. Med. Surg. Public Health 2024, 2, 100081. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, S.; Zhang, S.; Zhao, Z.; Cao, Y.; Chen, M.; Li, B. A Comparative Study of Fluoroquinolone-Resistant Escherichia coli Lineages Portrays Indistinguishable Pathogenicity- and Survivability-Associated Phenotypic Characteristics Between ST1193 and ST131. Infect. Drug Resist. 2020, 13, 4167–4175. [Google Scholar] [CrossRef]
- Feng, C.; Jia, H.; Yang, Q.; Zou, Q. Genetic Evolution of Antibiotic Resistance and Virulence Genes in Escherichia coli Isolates from a Chinese Hospital over a 12-Year Period. Microorganisms 2025, 13, 954. [Google Scholar] [CrossRef]
- Villavicencio-Carrisoza, O.; Grobeisen-Duque, O.; Garcia-Correa, A.L.; Monroy-Muñoz, I.E.; Villeda-Gabriel, G.; Sosa-González, I.E.; Flores-Herrera, H.; Figueroa-Damian, R.; Cerna-Cortes, J.F.; Rivera-Gutierrez, S.; et al. Advancing Understanding of Escherichia coli Pathogenicity in Preterm Neonatal Sepsis. Microorganisms 2025, 13, 219. [Google Scholar] [CrossRef]
- Zeng, Q.; Xiao, S.; Gu, F.; He, W.; Xie, Q.; Yu, F.; Han, L. Antimicrobial resistance and molecular epidemiology of uropathogenic Escherichia coli isolated from female patients in Shanghai, China. Front. Cell. Infect. Microbiol. 2021, 11, 653983. [Google Scholar] [CrossRef] [PubMed]
- Mirkalantari, S.; Masjedian, F.; Irajian, G.; Siddig, E.E.; Fattahi, A. Determination of the frequency of β-lactamase genes (bla SHV, bla TEM, bla CTX-M) and phylogenetic groups among ESBL-producing uropathogenic Escherichia coli isolated from outpatients. J. Lab. Med. 2019, 44, 27–33. [Google Scholar] [CrossRef]
- Pandit, R.; Awal, B.; Shrestha, S.S.; Joshi, G.; Rijal, B.P.; Parajuli, N.P. Extended-spectrum β-lactamase (ESBL) genotypes among multidrug-resistant uropathogenic Escherichia coli clinical isolates from a teaching hospital of Nepal. Interdiscip. Perspect. Infect. Dis. 2020, 2020, 6525826. [Google Scholar] [CrossRef] [PubMed]
- Puljko, A.; Barišić, I.; Rozman, S.D.; Križanović, S.; Babić, I.; Jelić, M.; Maravić, A.; Udiković-Kolić, N. Molecular epidemiology and mechanisms of carbapenem and colistin resistance in Klebsiella and other Enterobacterales from treated wastewater in Croatia. Environ. Int. 2024, 185, 108554. [Google Scholar] [CrossRef]
- Bedenić, B.; Meštrović, T. Mechanisms of resistance in gram-negative urinary pathogens: From country-specific molecular insights to global clinical relevance. Diagnostics 2021, 11, 800. [Google Scholar] [CrossRef]
- HHoashi, K.; Hayama, B.; Suzuki, M.; Sakurai, A.; Takehana, K.; Enokida, T.; Takeda, K.; Ohkushi, D.; Doi, Y.; Harada, S. Comparison of the Treatment Outcome of Piperacillin-Tazobactam versus Carbapenems for Patients with Bacteremia Caused by Extended-Spectrum β-Lactamase-Producing Escherichia coli in Areas with Low Frequency of Coproduction of OXA-1: A Preliminary Analysis. Microbiol. Spectr. 2022, 10, e02206–e02222. [Google Scholar]
- Jamali, S.; Tavakoly, T.; Mojtahedi, A.; Shenagari, M. The phylogenetic relatedness of bla NDM-1 harboring extended-spectrum β-lactamase producing uropathogenic Escherichia coli and Klebsiella pneumoniae in the North of Iran. Infect. Drug Resist. 2020, 13, 651–657. [Google Scholar] [CrossRef]
- Kiemde, D.; Ribeiro, I.; Sanou, S.; Coulibaly, B.; Sie, A.; Ouedraogo, A.-S.; Eibach, D. Molecular characterization of beta-lactamase genes produced by community-acquired uropathogenic Escherichia coli in Nouna. J. Infect. Dev. Ctries. 2020, 14, 1274–1280. [Google Scholar] [CrossRef]
- Talebi, M.; Najar-Peerayeh, S.; Bakhshi, B. Hidden carbapenem resistance in the community-and hospital-associated OXA-48 gene-carrying uropathogenic Escherichia coli. Gene Rep. 2020, 21, 100897. [Google Scholar] [CrossRef]
- Sharma, P.; Haycocks, J.R.J.; Middlemiss, A.D.; Kettles, R.A.; Sellars, L.E.; Ricci, V.; Piddock, L.J.V.; Grainger, D.C. The multiple antibiotic resistance operon of enteric bacteria controls DNA repair and outer membrane integrity. Nat. Commun. 2017, 8, 1–12. [Google Scholar] [CrossRef]
- Harris, M.; Fasolino, T.; Ivankovic, D.; Davis, N.J.; Brownlee, N. Genetic Factors That Contribute to Antibiotic Resistance through Intrinsic and Acquired Bacterial Genes in Urinary Tract Infections. Microorganisms 2023, 11, 1407. [Google Scholar] [CrossRef] [PubMed]
- Al Mamari, A.M.K.; Al Jabri, Z.; Sami, H.; Rizvi, S.G.A.; Chan, M.F.; Al Siyabi, T.; Rizvi, M. Evaluation of six commercial and in-house phenotypic tests for detection of AmpC β-lactamases: Is routine detection possible? JAC-Antimicrob. Resist. 2023, 5, dlad101. [Google Scholar] [CrossRef] [PubMed]
- Lyimo, B.; Buza, J.; Subbiah, M.; Temba, S.; Kipasika, H.; Smith, W.; Call, D.R. IncF plasmids are commonly carried by antibiotic resistant Escherichia coli isolated from drinking water sources in northern Tanzania. Int. J. Microbiol. 2016, 2016, 3103672. [Google Scholar] [CrossRef]
- Khezri, A.; Avershina, E.; Ahmad, R. Plasmid identification and plasmid-mediated antimicrobial gene detection in Norwegian isolates. Microorganisms 2020, 9, 52. [Google Scholar] [CrossRef] [PubMed]
- Al-Lami, R.A.H.; Shehab, Z.H.; Almohaidi, A.M.S. The Role of Efflux Pump for Antibiotic Resistance in Bacteria: The Role of Efflux Pump for Antibiotic Resistance in Bacteria. Iraqi J. Mark. Res. Consum. Prot. 2022, 14, 127–134. [Google Scholar] [CrossRef]
- Kharel, S.; Rimal, S.; Neupane, B.; Sharma, B.K.; Bashyal, N.S.; Shah, Y.; Chalise, B.S.; Rawal, M.; Tun, M.M.N.; Dumre, S.P. Detection of AcrAB efflux pump mediated ciprofloxacin resistance in Escherichia coli and Klebsiella pneumoniae in Nepal. BMC Infect. Dis. 2025, 25, 1227. [Google Scholar] [CrossRef]
- Kornelsen, V.; Kumar, A. Update on Multidrug Resistance Efflux Pumps in Acinetobacter spp. Antimicrob. Agents Chemother. 2021, 65. [Google Scholar] [CrossRef]
- Alobaidallah, M.S.A.; García, V.; De Mets, R.; Wellner, S.M.; Thomsen, L.E.; Herrero-Fresno, A.; Olsen, J.E. Uncovering the Important Genetic Factors for Growth during Cefotaxime-Gentamicin Combination Treatment in blaCTX-M-1 Encoding Escherichia coli. Antibiotics 2023, 12, 993. [Google Scholar] [CrossRef]
- Manges, A.R.; Geum, H.M.; Guo, A.; Edens, T.J.; Fibke, C.D.; Pitout, J.D.D. Global Extraintestinal Pathogenic Escherichia coli (ExPEC) Lineages. Clin. Microbiol. Rev. 2019, 32, e00135-18. [Google Scholar] [CrossRef]
- Mousa, N.; Aiesh, B.M.; Jomaa, R.; Zouneh, Y.; Namrouti, A.; Abutaha, A.; Al-Jabi, S.W.; Sabateen, A.; Zyoud, S.H. Antibiotic resistance profiles and risk factors of multidrug-resistant Escherichia coli in a large tertiary care hospital in a low- and middle-income country. Sci. Rep. 2025, 15, 1–10. [Google Scholar] [CrossRef]
- Rezatofighi, S.E.; Mirzarazi, M.; Salehi, M. Virulence genes and phylogenetic groups of uropathogenic Escherichia coli isolates from patients with urinary tract infection and uninfected control subjects: A case-control study. BMC Infect. Dis. 2021, 21, 361. [Google Scholar] [CrossRef]
- Gonyar, L.A.; Sauder, A.B.; Mortensen, L.; Willsey, G.G.; Kendall, M.M. The yad and yeh fimbrial loci influence gene expression and virulence in enterohemorrhagic Escherichia coli O157:H7. mSphere 2024, 9, e0012424. [Google Scholar] [CrossRef] [PubMed]
- Ross, B.N.; Rojas-Lopez, M.; Cieza, R.J.; McWilliams, B.D.; Torres, A.G. The Role of Long Polar Fimbriae in Escherichia coli O104:H4 Adhesion and Colonization. PLoS ONE 2015, 10, e0141845. [Google Scholar] [CrossRef] [PubMed]
- Wullt, B.; Bergsten, G.; Samuelsson, M.; Svanborg, C. The role of P fimbriae for Escherichia coli establishment and mucosal inflammation in the human urinary tract. Int. J. Antimicrob. Agents 2002, 19, 522–538. [Google Scholar] [CrossRef] [PubMed]
- Mahapatra, A.; Behera, B.; Sarathi, S.; Nayak, S.; Naim, M.S.; Dash, M.S.; Patra, M.B.; Sahoo, M.S. Molecular mechanism of virulence, microbiological susceptibility, host factor and clinical outcome of blood stream infection caused by Escherichia coli. Int. J. Infect. Dis. 2025, 152, 107665. [Google Scholar] [CrossRef]
- Nagy, G.; Dobrindt, U.; Kupfer, M.; EmödY, L.; Karch, H.; Hacker, J. Expression of Hemin Receptor Molecule ChuA Is Influenced by RfaH in Uropathogenic Escherichia coli Strain 536. Infect. Immun. 2001, 69, 1924–1928. [Google Scholar] [CrossRef]
- Sabri, M.; Caza, M.; Proulx, J.; Lymberopoulos, M.H.; Brée, A.; Moulin-Schouleur, M.; Curtiss, R.; Dozois, C.M. Contribution of the SitABCD, MntH, and FeoB Metal Transporters to the Virulence of Avian Pathogenic Escherichia coli O78 Strain χ7122. Infect. Immun. 2008, 76, 601–611. [Google Scholar] [CrossRef]
- Porter, S.B.; Johnston, B.D.; Kisiela, D.; Clabots, C.; Sokurenko, E.V.; Johnson, J.R. Bacteriophage Cocktail and Microcin-Producing Probiotic Escherichia coli Protect Mice Against Gut Colonization with Multidrug-Resistant Escherichia coli Sequence Type 131. Front. Microbiol. 2022, 13, 887799. [Google Scholar] [CrossRef]
- Fatima, S.; Akbar, A.; Irfan, M.; Shafee, M.; Ali, A.; Ishaq, Z.; Hassan, S.S. Virulence Factors and Antimicrobial Resistance of Uropathogenic Escherichia coli EQ101 UPEC Isolated from UTI Patient in Quetta, Balochistan, Pakistan. BioMed Res. Int. 2023, 2023, 7278070. [Google Scholar] [CrossRef]
- Ravan, H.; Amandadi, M. Analysis of yeh Fimbrial Gene Cluster in Escherichia coli O157:H7 in Order to Find a Genetic Marker for this Serotype. Curr. Microbiol. 2015, 71, 274–282. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing A CLSI Supplement for Global Application; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Pevzner, P.A. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.; Cosentino, S.; Rasmussen, S.; Rundsten, C.; Hasman, H.; Marvig, R.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.; Aarestrup, F.; et al. Multilocus Sequence Typing of Total Genome Sequenced Bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef]
- Clausen, P.; Aarestrup, F.; Lund, O. Rapid and precise alignment of raw reads against redundant databases with KMA. BMC Bioinform. 2018, 19, 307. [Google Scholar] [CrossRef]
- Bartual, S.; Seifert, H.; Hippler, C.; Luzon, M.; Wisplinghoff, H.; Rodríguez-Valera, F. Development of a multilocus sequence typing scheme for characterization of clinical isolates of Acinetobacter baumannii. J. Clin. Microbiol. 2005, 43, 4382–4390. [Google Scholar] [CrossRef]
- Griffiths, D.; Fawley, W.; Kachrimanidou, M.; Bowden, R.; Crook, D.; Fung, R.; Golubchik, T.; Harding, R.; Jeffery, K.; Jolley, K. Multilocus sequence typing of Clostridium difficile. J. Clin. Microbiol. 2010, 48, 770–778. [Google Scholar] [CrossRef]
- Lemee, L.; Dhalluin, A.; Pestel-Caron, M.; Lemeland, J.; Pons, J. Multilocus sequence typing analysis of human and animal Clostridium difficile isolates of various toxigenic types. J. Clin. Microbiol. 2004, 42, 2609–2617. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic Resistome Surveillance with the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; Garcia-Fernandez, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. PlasmidFinder and pMLST: In silico detection and typing of plasmids. Antimicrob. Agents Chemother 2014, 58, 3895–3903. [Google Scholar] [CrossRef]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.; Karch, H.; Reeves, P.; Maiden, M.; Ochman, H.; et al. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef]
- Jaureguy, F.; Landraud, L.; Passet, V.; Diancourt, L.; Frapy, E.; Guigon, G.; Carbonnelle, E.; Lortholary, O.; Clermont, O.; Denamur, E.; et al. Phylogenetic and genomic diversity of human bacteremic Escherichia coli strains. BMC Genom. 2008, 9, 560. [Google Scholar] [CrossRef]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-time whole-genome sequencing for routine typing, surveillance, and outbreak detection of verotoxigenic Escherichia coli. J. Clin. Micobiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef]
- Malberg Tetzschner, A.M.; Johnson, J.R.; Johnston, B.D.; Lund, O.; Scheutz, F. In Silico Genotyping of Escherichia coli Isolates for Extraintestinal Virulence Genes by Use of Whole-Genome Sequencing Data. J. Clin. Micobiol. 2020, 58, e01269-20. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0. Molecular Biology and Evolution. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://www.R-project.org/ (accessed on 30 November 2023).
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL): An Online Tool for Phylogenetic Tree Display and Annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.