
Emergence and Genomic Features of a mcr-1 Escherichia coli from Duck in Hungary
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Isolation and Identification of the Colistin Resistant E. coli Strain Ec45-2020
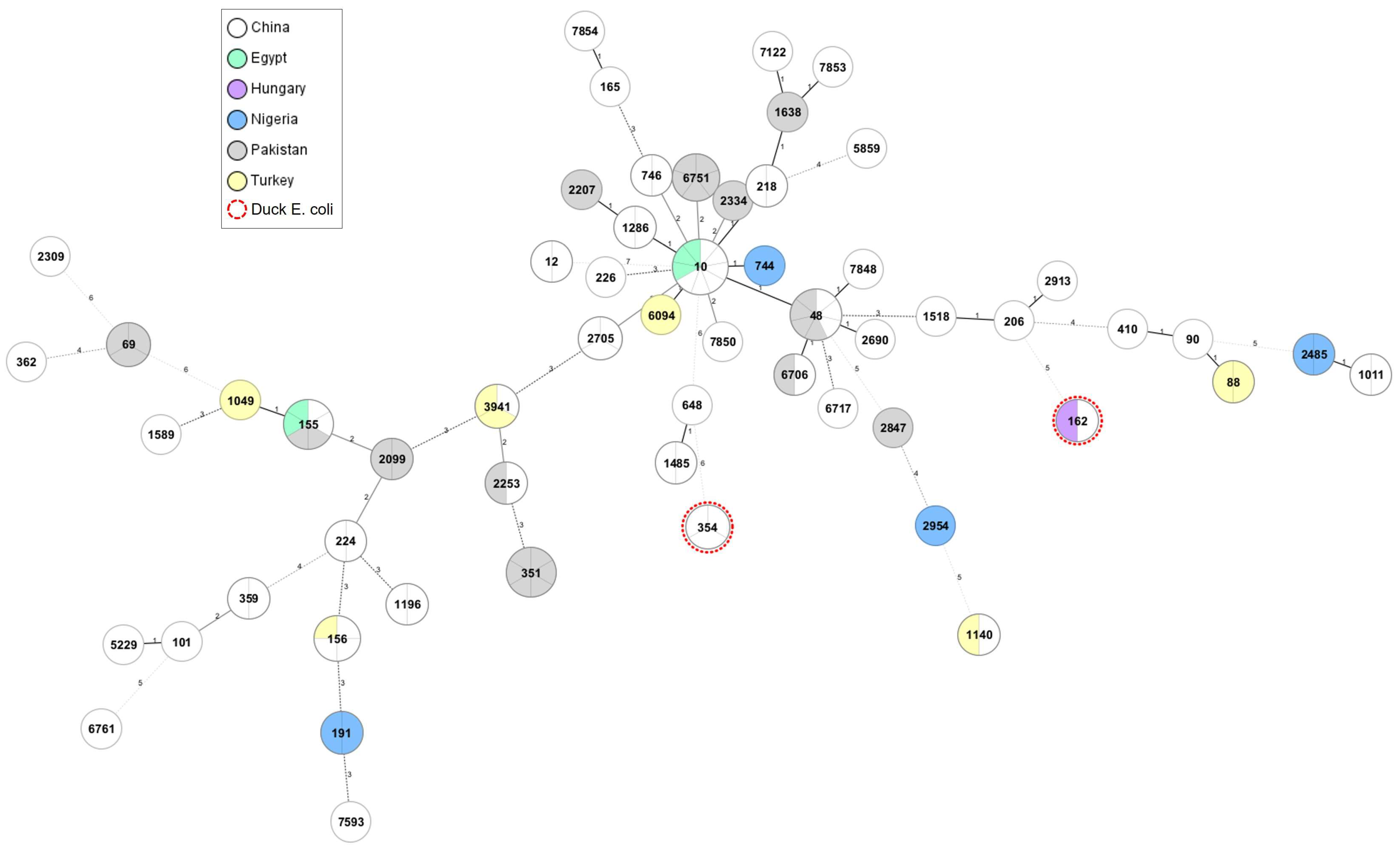
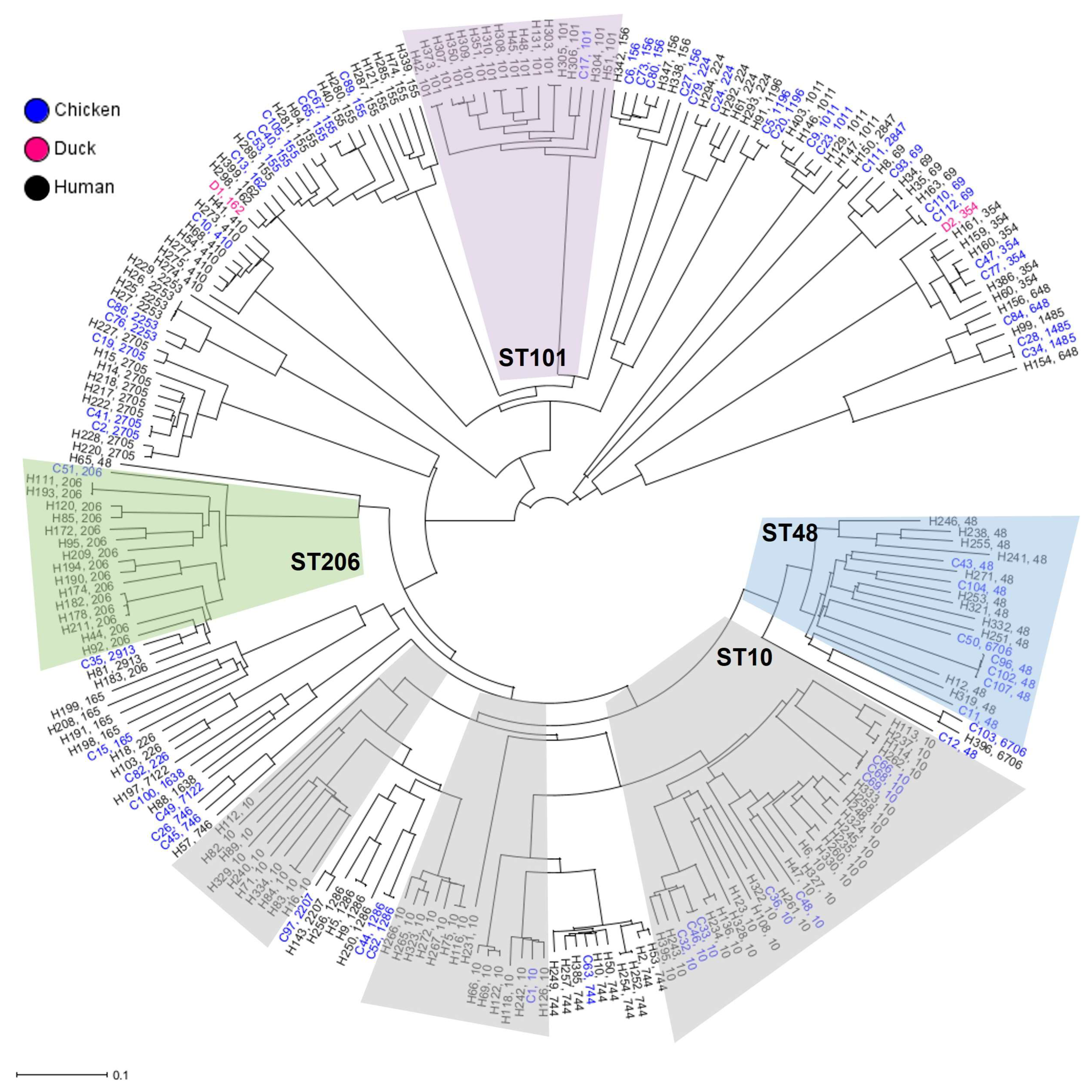
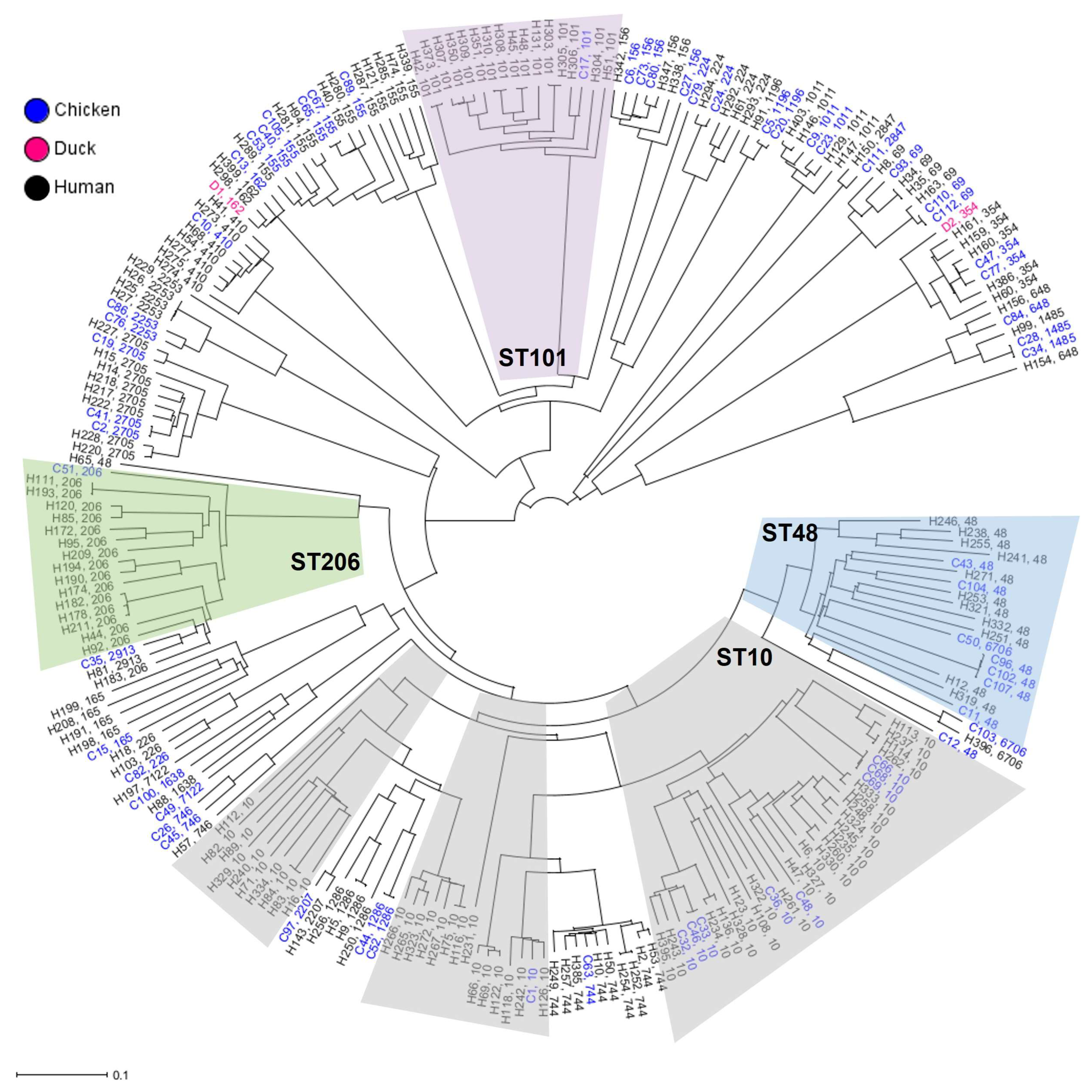
2.2. Phylogenetic Relation and Genomic Diversity of mcr-1-Positive E. coli Strains from Poultry and Humans
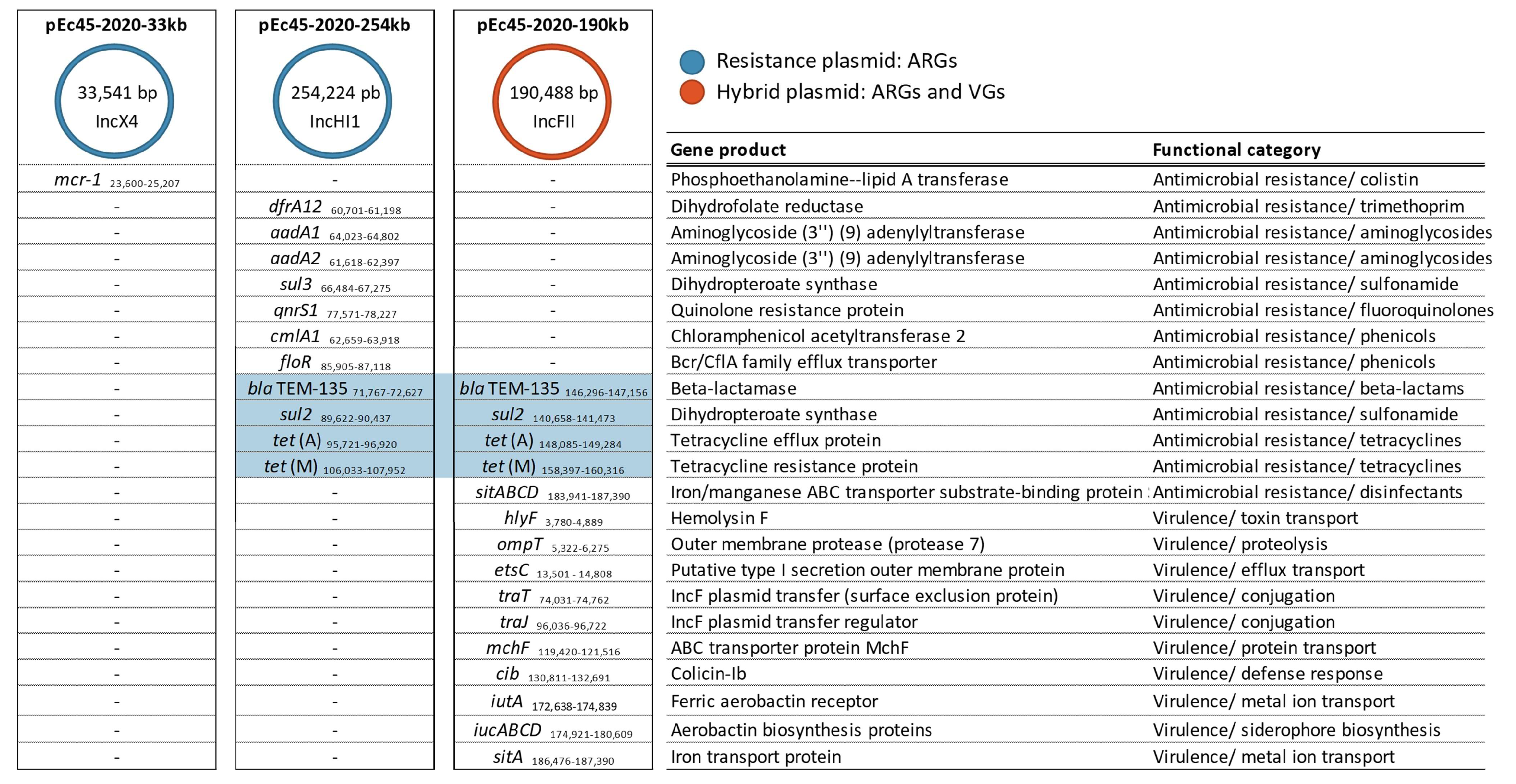
2.3. Genomic Determinants of Antimicrobial Resistance and Virulence
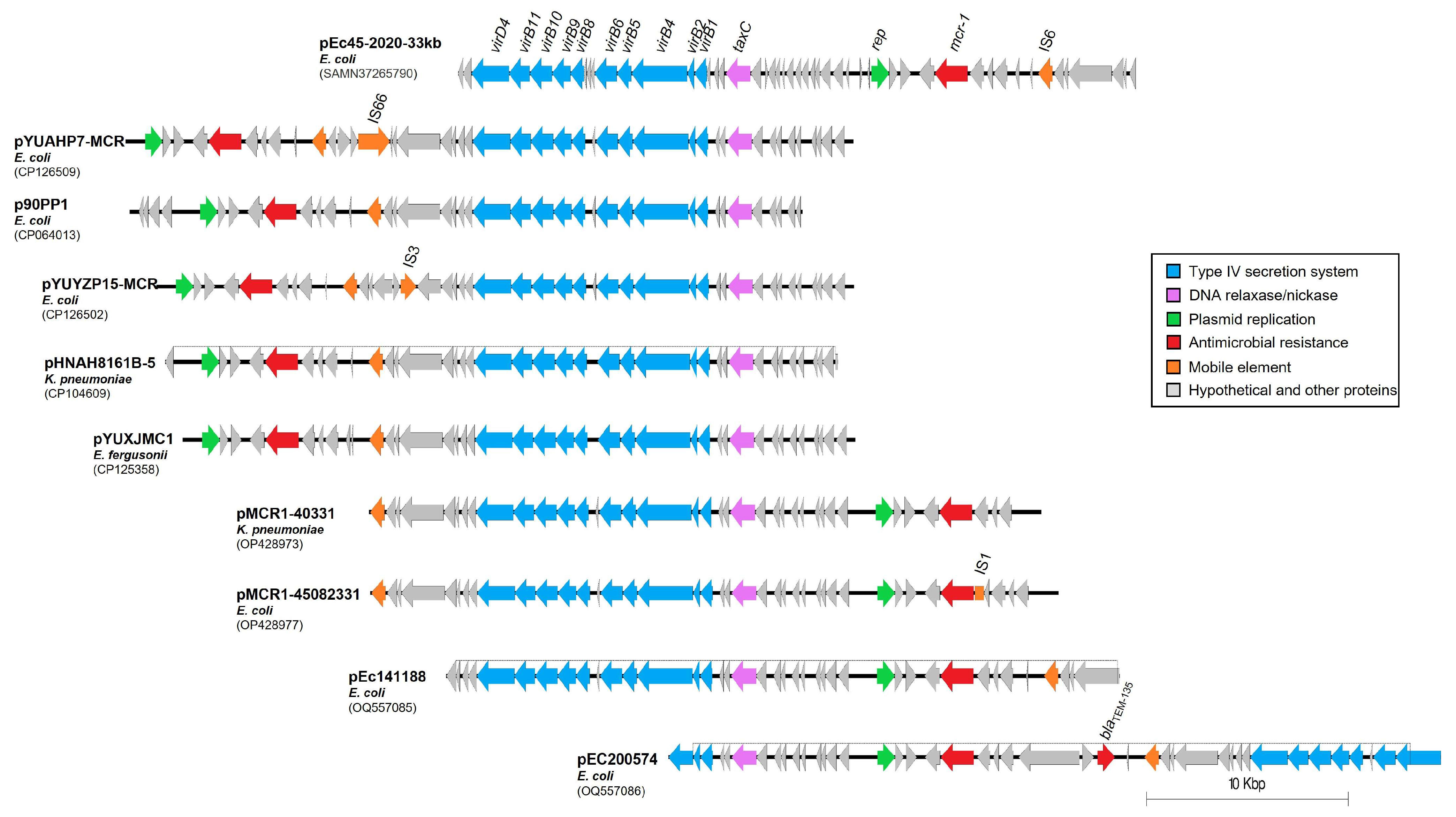
2.4. Genome Architecture of the mcr-1 Plasmid pEc45-2020-33kb Identified in Hungary
3. Discussion
4. Materials and Methods
4.1. Collection of E. coli Strains Subjected to mcr-1 Screening
4.2. Identification of the mcr-1 Gene and Antibiotic Susceptibility Testing
4.3. Whole-Genome Sequencing and Analysis
4.4. Core Genome-Based Multi-Locus Sequence Typing (cgMLST)
4.5. In Silico Analysis of the Antibiotic Resistance and Virulence Genotypes and Testing the Transferability of the mcr-1 Plasmid
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaper, J.B. Pathogenic Escherichia coli. Int. J. Med. Microbiol. 2005, 295, 355–356. [Google Scholar] [CrossRef] [PubMed]
- Kathayat, D.; Lokesh, D.; Ranjit, S.; Rajashekara, G. Avian pathogenic Escherichia coli (APEC): An overview of virulence and pathogenesis factors, zoonotic potential, and control strategies. Pathogens 2021, 10, 467. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, S.L.; Stegger, M.; Kudirkiene, E.; Lilje, B.; Poulsen, L.L.; Ronco, T.; Pires Dos Santos, T.; Kiil, K.; Bisgaard, M.; Pedersen, K.; et al. Diversity and population overlap between avian and human Escherichia coli belonging to sequence type 95. mSphere 2019, 4, e00333-18. [Google Scholar] [CrossRef] [PubMed]
- Mellata, M. Human and avian extraintestinal pathogenic Escherichia coli: Infections, zoonotic risks, and antibiotic resistance trends. Foodborne Pathog. Dis. 2013, 10, 916–932. [Google Scholar] [CrossRef]
- Apostolakos, I.; Laconi, A.; Mughini-Gras, L.; Yapicier, Ö.Ş.; Piccirillo, A. Occurrence of colibacillosis in broilers and its relationship with avian pathogenic Escherichia coli (APEC) population structure and molecular characteristics. Front. Vet. Sci. 2021, 8, 737720. [Google Scholar] [CrossRef]
- Hu, J.; Afayibo, D.J.A.; Zhang, B.; Zhu, H.; Yao, L.; Guo, W.; Wang, X.; Wang, Z.; Wang, D.; Peng, H.; et al. Characteristics, pathogenic mechanism, zoonotic potential, drug resistance, and prevention of avian pathogenic Escherichia coli (APEC). Front. Microbiol. 2022, 13, 1049391. [Google Scholar] [CrossRef]
- Kemmett, K.; Williams, N.J.; Chaloner, G.; Humphrey, S.; Wigley, P.; Humphrey, T. The contribution of systemic Escherichia coli infection to the early mortalities of commercial broiler chickens. Avian Pathol. 2014, 43, 37–42. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Mead, A.; Toutain, P.L.; Richez, P.; Pelligand, L. Targeted dosing for susceptible heteroresistant subpopulations may improve rational dosage regimen prediction for colistin in broiler chickens. Sci. Rep. 2023, 13, 12822. [Google Scholar] [CrossRef]
- Rebelo, A.R.; Bortolaia, V.; Kjeldgaard, J.S.; Pedersen, S.K.; Leekitcharoenphon, P.; Hansen, I.M.; Guerra, B.; Malorny, B.; Borowiak, M.; Hammerl, J.A.; et al. Multiplex PCR for detection of plasmid-mediated colistin resistance determinants, mcr-1, mcr-2, mcr-3, mcr-4 and mcr-5 for surveillance purposes. Euro Surveill. 2018, 23, 17–00672. [Google Scholar] [CrossRef]
- Kimura, B. Will the emergence of core genome MLST end the role of in silico MLST? Food Microbiol. 2018, 75, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Juhász, E.; Iván, M.; Pintér, E.; Pongrácz, J.; Kristóf, K. Colistin resistance among blood culture isolates at a tertiary care centre in Hungary. J. Glob. Antimicrob. Resist. 2017, 11, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Adorján, A.; Makrai, L.; Mag, T.; Jánosi, S.; Könyves, L.; Tóth, I. High frequency of multidrug-resistant (MDR) atypical enteropathogenic Escherichia coli (aEPEC) in broilers in Hungary. Front. Vet. Sci. 2020, 7, 511. [Google Scholar] [CrossRef] [PubMed]
- Adorján, A.; Thuma, Á.; Könyves, L.; Tóth, I. First isolation of atypical enteropathogenic Escherichia coli from geese (Anser anser domestica) and first description of atypical EPEC from turkeys and pigeons in Hungary. BMC Vet. Res. 2021, 17, 263. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xie, Y.; Liu, M.; Tai, C.; Sun, J.; Deng, Z.; Ou, H.-Y. oriTfinder: A web-based tool for the identification of origin of transfers in DNA sequences of bacterial mobile genetic elements. Nucleic Acids Res. 2018, 46, gky352. [Google Scholar] [CrossRef] [PubMed]
- Olaitan, A.O.; Morand, S.; Rolain, J.-M. Mechanisms of polymyxin resistance: Acquired and intrinsic resistance in bacteria. Front. Microbiol. 2014, 5, 643. [Google Scholar] [CrossRef]
- Jansen, W.; van Hout, J.; Wiegel, J.; Iatridou, D.; Chantziaras, I.; De Briyne, N. Colistin use in european livestock: Veterinary field data on trends and perspectives for further reduction. Vet. Sci. 2022, 9, 650. [Google Scholar] [CrossRef]
- European Medicines Agency, European Surveillance of Veterinary Antimicrobial Consumption. Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2019 and 2020; EMA/58183/2021; European Surveillance of Veterinary Antimicrobial Consumption: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Ali, M.W.; Utsho, K.S.; Karmakar, S.; Hoque, M.N.; Rahman, M.T.; Hassan, J. First report on the molecular characteristics of mcr-1 colistin resistant E. coli isolated from retail broiler meat in Bangladesh. Int. J. Food Microbiol. 2023, 388, 110065. [Google Scholar] [CrossRef]
- Díaz-Jiménez, D.; García-Meniño, I.; Fernández, J.; García, V.; Mora, A. Chicken and turkey meat: Consumer exposure to multidrug-resistant Enterobacteriaceae including mcr-carriers, uropathogenic E. coli and high-risk lineages such as ST131. Int. J. Food Microbiol. 2020, 331, 108750. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority) and ECDC (European Centre for Disease Prevention and Control), 2022. The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2019–2020. EFSA J. 2022, 20, 7209. [Google Scholar] [CrossRef]
- Gelbicova, T.; Kolackova, I.; Krutova, M.; Karpiskova, R. The emergence of mcr-1-mediated colistin-resistant Escherichia coli and Klebsiella pneumoniae in domestic and imported turkey meat in the Czech Republic 2017–2018. Folia Microbiol. 2020, 65, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Santiago, J.; Cornejo-Juárez, P.; Silva-Sánchez, J.; Garza-Ramos, U. Polymyxin resistance in enterobacterales: Overview and epidemiology in the Americas. Int. J. Antimicrob. Agents 2021, 58, 106426. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.-D.; Jin, W.-J.; Li, R.-B.; Zhang, R.-M.; Sun, J.; Liu, Y.-H.; Wang, M.-G.; Liao, X.-P. Prevalence and molecular characteristics of mcr-1-positive Escherichia coli isolated from duck farms and the surrounding environments in coastal China. Microbiol. Res. 2023, 270, 127348. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhang, R.; Yang, Y.; Li, H.; Wang, J.; Lan, J.; Li, P.; Zhu, Y.; Xie, Z.; Jiang, S. Occurrence and molecular characteristics of mcr-1-positive Escherichia coli from healthy meat ducks in Shandong Province of China. Animals 2020, 10, 1299. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.-G.; Yu, Y.; Wang, D.; Yang, R.-S.; Jia, L.; Cai, D.-T.; Zheng, S.-L.; Fang, L.-X.; Sun, J.; Liu, Y.-H.; et al. The emergence and molecular characteristics of New Delhi metallo β-lactamase-producing Escherichia coli from ducks in Guangdong, China. Front. Microbiol. 2021, 12, 677633. [Google Scholar] [CrossRef]
- Hu, X.; Chen, Y.; Xu, H.; Qiao, J.; Ge, H.; Liu, R.; Zheng, B. Genomic epidemiology and transmission characteristics of mcr1-positive colistin-resistant Escherichia coli strains circulating at natural environment. Sci. Total Environ. 2023, 882, 163600. [Google Scholar] [CrossRef]
- Biswas, S.; Li, Y.; Elbediwi, M.; Yue, M. Emergence and Dissemination of Mcr-Carrying Clinically Relevant Salmonella Typhimurium Monophasic Clone ST34. Microorganisms 2019, 7, 298. [Google Scholar] [CrossRef]
- Li, A.; Yang, Y.; Miao, M.; Chavda, K.D.; Mediavilla, J.R.; Xie, X.; Feng, P.; Tang, Y.-W.; Kreiswirth, B.N.; Chen, L.; et al. Complete sequences of mcr-1 -harboring plasmids from extended-spectrum-β-lactamase- and carbapenemase-producing Enterobacteriaceae. Antimicrob. Agents Chemother. 2016, 60, 4351–4354. [Google Scholar] [CrossRef]
- Bai, F.; Li, X.; Niu, B.; Zhang, Z.; Malakar, P.K.; Liu, H.; Pan, Y.; Zhao, Y. A mcr-1-carrying conjugative IncX4 plasmid in colistin-resistant Escherichia coli ST278 strain isolated from dairy cow feces in Shanghai, China. Front. Microbiol. 2018, 9, 2833. [Google Scholar] [CrossRef]
- Smillie, C.; Garcillán-Barcia, M.; Francia, M.; Rocha, E.P.; de la Cruz, F. Mobility of plasmids. Microbiol. Mol. Biol. Rev. MMBR 2010, 74, 434–452. [Google Scholar] [CrossRef]
- Guzman-Otazo, J.; Gonzales-Siles, L.; Poma, V.; Bengtsson-Palme, J.; Thorell, K.; Flach, C.-F.; Iñiguez, V.; Sjöling, Å. Diarrheal bacterial pathogens and multi-resistant Enterobacteria in the Choqueyapu River in La Paz, Bolivia. PLoS ONE 2019, 14, e0210735. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, W.; Ohsaki, Y.; Taniguchi, Y.; Koide, S.; Kawamura, K.; Suzuki, M.; Kimura, K.; Wachino, J.; Nagano, Y.; Arakawa, Y.; et al. High prevalence of Bla CTX-M-14 among genetically diverse Escherichia coli recovered from retail raw chicken meat portions in Japan. Int. J. Food Microbiol. 2018, 284, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Szmolka, A.; Wami, H.; Dobrindt, U. Comparative genomics of emerging lineages and mobile resistomes of contemporary broiler strains of Salmonella Infantis and E. coli. Front. Microbiol. 2021, 12, 642125. [Google Scholar] [CrossRef]
- Yoon, S.; Lee, Y.J. Molecular Characteristics of ESBL-producing Escherichia coli isolated from chickens with colibacillosis. J. Vet. Sci. 2021, 23, e37. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro-Almeida, M.; Mourão, J.; Novais, Â.; Pereira, S.; Freitas-Silva, J.; Ribeiro, S.; da Costa, P.M.; Peixe, L.; Antunes, P. High diversity of pathogenic Escherichia coli clones carrying mcr-1 among gulls underlines the need for strategies at the environment–livestock–human interface. Environ. Microbiol. 2022, 24, 4702–4713. [Google Scholar] [CrossRef]
- Zhuge, X.; Jiang, M.; Tang, F.; Sun, Y.; Ji, Y.; Xue, F.; Ren, J.; Zhu, W.; Dai, J. Avian-source mcr-1-positive Escherichia coli is phylogenetically diverse and shares virulence characteristics with E. coli causing human extra-intestinal infections. Vet. Microbiol. 2019, 239, 108483. [Google Scholar] [CrossRef] [PubMed]
- Zhuge, X.; Ji, Y.; Tang, F.; Sun, Y.; Jiang, M.; Hu, W.; Wu, Y.; Xue, F.; Ren, J.; Zhu, W.; et al. Population structure and antimicrobial resistance traits of avian-origin mcr-1-positive Escherichia coli in Eastern China, 2015 to 2017. Transbound. Emerg. Dis. 2019, 66, 1920–1929. [Google Scholar] [CrossRef]
- Lindsey, R.L.; Garcia-Toledo, L.; Fasulo, D.; Gladney, L.M.; Strockbine, N. Multiplex polymerase chain reaction for identification of Escherichia coli, Escherichia albertii and Escherichia fergusonii. J. Microbiol. Methods. 2017, 140, 1–4. [Google Scholar] [CrossRef]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 8.0. 2018. Available online: https://www.eucast.org/ast_of_bacteria/previous_versions_of_documents (accessed on 17 July 2023).
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Jünemann, S.; Sedlazeck, F.J.; Prior, K.; Albersmeier, A.; John, U.; Kalinowski, J.; Mellmann, A.; Goesmann, A.; von Haeseler, A.; Stoye, J.; et al. Updating benchtop sequencing performance comparison. Nat. Biotechnol. 2013, 31, 294–296. [Google Scholar] [CrossRef]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.; Ochman, H.; et al. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef] [PubMed]
- Joensen, K.G.; Tetzschner, A.M.M.; Iguchi, A.; Aarestrup, F.M.; Scheutz, F. Rapid and easy in silico serotyping of Escherichia coli isolates by use of whole-genome sequencing data. J. Clin. Microbiol. 2015, 53, 2410–2426. [Google Scholar] [CrossRef] [PubMed]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Tetzschner, A.M.M.; Johnson, J.R.; Johnston, B.D.; Lund, O.; Scheutz, F. In silico genotyping of Escherichia coli isolates for extraintestinal virulence genes by use of whole-genome sequencing data. J. Clin. Microbiol. 2020, 58, e01269-20. [Google Scholar] [CrossRef]
- Carattoli, A.; Hasman, H. Horizontal gene transfer, methods and protocols. Methods Mol. Biol. 2019, 2075, 285–294. [Google Scholar] [CrossRef]
- Rodriguez-Grande, J.; Fernandez-Lopez, R. Measuring plasmid conjugation using antibiotic selection. Methods Mol. Biol. 2019, 2075, 93–98. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pigeon | Chicken | Turkey | Goose | Duck | |
|---|---|---|---|---|---|
| Number of sampled farms (dead animals) | 5 (n = 38) | 8 (n = 27) | 1 (n = 4) | 9 (n = 53) | 8 (n = 36) * |
| Number of sampled slaughterhouses (culled animals) | - | 1 (n = 132) | 1 (n = 51) | 1 (n = 48) | 1 (n = 51) |
| Number of sampled outdoor keeping farms (live animals) | - | 1 (n = 18) | 1 (n = 6) | - | - |
| Number of sampled backyard farms (live animals) | - | 1 (n = 15) | - | - | - |
| Genotype | Poultry (%) | Human (%) | Total % (no.) |
|---|---|---|---|
| ST10 | 13.85 | 73.85 | 23.95 (57) |
| ST48 | 10.77 | 18.46 | 7.98 (19) |
| ST101 | 1.54 | 24.62 | 7.14 (17) |
| ST206 | 1.54 | 24.62 | 7.14 (17) |
| ST155 | 9.23 | 15.38 | 6.72 (16) |
| ST2705 | 4.62 | 12.31 | 4.62 (11) |
| ST744 | 1.54 | 13.85 | 4.20 (10) |
| ST354 * | 4.62 | 7.69 | 3.36 (8) |
| ST410 | 1.54 | 10.77 | 3.36 (8) |
| ST69 | 4.62 | 6.15 | 2.94 (7) |
| ST156 | 6.15 | 4.62 | 2.94 (7) |
| ST224 | 3.08 | 6.15 | 2.52 (6) |
| ST1286 | 3.08 | 6.15 | 2.52 (6) |
| ST1011 | 3.08 | 6.15 | 2.52 (6) |
| ST2253 | 3.08 | 6.15 | 2.52 (6) |
| ST165 | 1.54 | 6.15 | 2.10 (5) |
| ST162 * | 3.08 | 3.08 | 1.68 (4) |
| ST226 | 1.54 | 3.08 | 1.26 (3) |
| ST648 | 1.54 | 3.08 | 1.26 (3) |
| ST746 | 3.08 | 1.54 | 1.26 (3) |
| ST1196 | 3.08 | 1.54 | 1.26 (3) |
| ST1485 | 3.08 | 1.54 | 1.26 (3) |
| ST6706 | 3.08 | 1.54 | 1.26 (3) |
| ST1638 | 1.54 | 1.54 | 0.84 (2) |
| ST2207 | 1.54 | 1.54 | 0.84 (2) |
| ST2847 | 1.54 | 1.54 | 0.84 (2) |
| ST2913 | 1.54 | 1.54 | 0.84 (2) |
| ST7122 | 1.54 | 1.54 | 0.84 (2) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szmolka, A.; Gellért, Á.; Szemerits, D.; Rapcsák, F.; Spisák, S.; Adorján, A. Emergence and Genomic Features of a mcr-1 Escherichia coli from Duck in Hungary. Antibiotics 2023, 12, 1519. https://doi.org/10.3390/antibiotics12101519
Szmolka A, Gellért Á, Szemerits D, Rapcsák F, Spisák S, Adorján A. Emergence and Genomic Features of a mcr-1 Escherichia coli from Duck in Hungary. Antibiotics. 2023; 12(10):1519. https://doi.org/10.3390/antibiotics12101519
Chicago/Turabian StyleSzmolka, Ama, Ákos Gellért, Dóra Szemerits, Fanni Rapcsák, Sándor Spisák, and András Adorján. 2023. "Emergence and Genomic Features of a mcr-1 Escherichia coli from Duck in Hungary" Antibiotics 12, no. 10: 1519. https://doi.org/10.3390/antibiotics12101519
APA StyleSzmolka, A., Gellért, Á., Szemerits, D., Rapcsák, F., Spisák, S., & Adorján, A. (2023). Emergence and Genomic Features of a mcr-1 Escherichia coli from Duck in Hungary. Antibiotics, 12(10), 1519. https://doi.org/10.3390/antibiotics12101519